Note that corporate logos (such as the USQ Phoenix, and any other company represented) and branding are specifically excluded from the Creative Commons Attribution ShareAlike License 4.0 of this work, and may not be reproduced under any circumstances without the express written permission of the copyright holders.
Copyright
This work, Fundamentals of Anatomy and Physiology, is a derivative of Anatomy and Physiology by J. Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble and Peter DeSaix. The original text is used under a Creative Commons Attribution 4.0 License, and can be found here: https://openstax.org/details/books/anatomy-and-physiology
Dr Anna Chruścikis an academic at the University of Southern Queensland, Springfield, Australia. Anna has taught courses in human anatomy and physiology; histopathology and cytology; techniques in comparative physiology; pathophysiology; cells, tissues and regeneration; metabolism; immunology; biomolecular sciences laboratory; biochemistry and biochemical pathways in Australian universities.Her research background focused on the relationship between transforming growth factor and colon cancer stem cells. Anna strives to excite and inspire students about science by providing relatable guidance, support and knowledge.
Dr Kate Kauter is an Associate Professor in biomedical science at the University of Southern Queensland. She has taught anatomy, physiology, pathophysiology, pharmacology and microbiology to students from many disciplines including nursing, biomedical sciences, food sciences and agricultural sciences, among others. Kate has developed a number of digital activities to increase student interaction with these content areas and has deployed the use of current technologies, including use of 3D and animations in practical classes to inspire students’ learning. The latest venture is the provision of an open education resource for the study of anatomy and physiology to allow all students access to the fundamental information needed in the understanding of the human body.
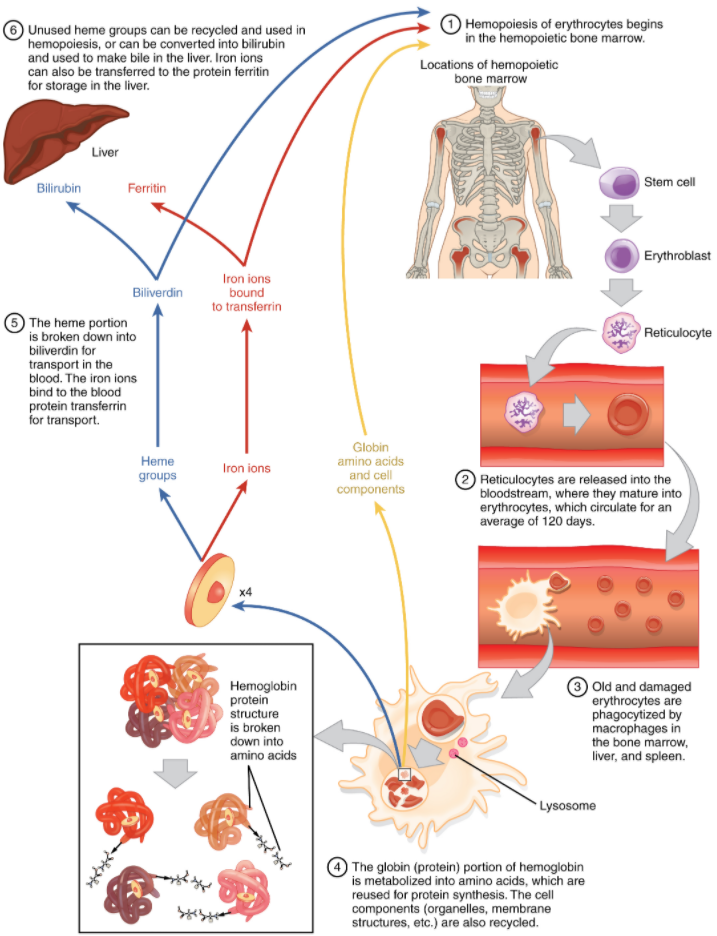
Dr Louisa Windus is a lecturer and researcher in the school of Health and Wellbeing (Biomedical Sciences) at the University of Southern Queensland, Australia. Louisa has a passion for developing methods that foster engagement and empower students to learn. Outside of the classroom, Louisa’s research focusses on biomarkers or molecular factors that mediate cancer progression and growth. She has collaborated extensively with research institutes across Australia and has been influential in developing novel 3D in vitro models that have helped expediate the drug discovery pipeline.
Dr Eliza Whitesideis a biomedical science researcher and Associate Professor at the University of Southern Queensland, Toowoomba, Australia. For the past two decades, Eliza has taught courses in introductory biomedical science, cell and molecular biology, anatomy and physiology, laboratory methods, pathophysiology andbiotechnology in universities in Australia and in the United Kingdom.Eliza’s passion is in improving the lives of others through accessible knowledge building, using learning and teaching scholarship, research and community outreach. Her research background is investigating dysregulated cell biology in cancer and chronic wounds. Her community outreach includes cancer education to the public and‘hands on’ science, particularly in underserved communitiessuch as regional and remote schools.
Acknowledgments
2
This book is predominantly based on the open textbook Anatomy and Physiology by J. Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble and Peter DeSaix. Our team of biomedical science academics and librarians at the University of Southern Queensland have a desire to help students succeed in their studies of anatomy and physiology by providing equitable access to information resources. This has driven us to adapt Anatomy and Physiologyfor Australian tertiary students in biomedical and health disciplines.
In Fundamentals of Anatomy and Physiology, you will find an Australian perspective on the fundamentals of anatomy and physiology along with career perspectives, with content especially re-worked for Australian students. Our team has also added new contributions including interactive quizzes, appendices and a glossary. Our sincere thanks to each team member for their tireless efforts, enthusiastic support of the project and their valued work on this book. Please see About the Authors for the list of our contributors. We would also like to thank Leanne Dooley for her review and original contributions to Chapter 6 Blood.
This book would also not have been possible without the outstanding contribution of our Open Education Content Librarian Nikki Andersen. We offer our sincere thanks to Nikki for her expertise, insightful guidance and dedicated approach in stepping us through this new process. Nikki, you made the journey enjoyable, exciting and achievable. Thank you. We also extend our appreciation to Adrian Stagg, Manager (Open Educational Practice) for his support, advice and expertise in open textbooks.
Finally, we thank Dr Anna Chruscik and Associate Professor Kate Kauter who committed their substantial skills and enthusiasm to create a seamless project which harnessed the writing talents of our team. Their writing and editing expertise supported us in producing this open textbook.
Acknowledgment of Country
The authors wish to acknowledge the Aboriginal and Torres Strait Islander peoples of this nation. We acknowledge the traditional owners of the country throughout Australia and their continuing connection to the land, culture and community. We acknowledge the traditional custodians of the lands on which we live and work, and where the book was written. We acknowledge the cultural diversity of all Aboriginal and Torres Strait Islander peoples and pay respect to Elders past, present and future. We celebrate the continuous living cultures of First Australians and acknowledge the important contributions Aboriginal and Torres Strait Islander people have and continue to make in Australian society.
Preface
3
The University of Southern Queensland (USQ) is committed to advancing the use of open textbooks in higher education. This textbook is a tool to support first year anatomy and physiology courses taught in Australia, aiming to provide students with an increased access to free, high-quality learning materials.
The material in this textbook is largely based on OpenStax’s Anatomy & Physiology textbook, however, has been modified for Australian course curriculum.
The coverage and scope of the current text includes:
Levels of organisation, homeostasis and nomenclature
Cells: structure and reproduction
Tissues, organs, and Systems
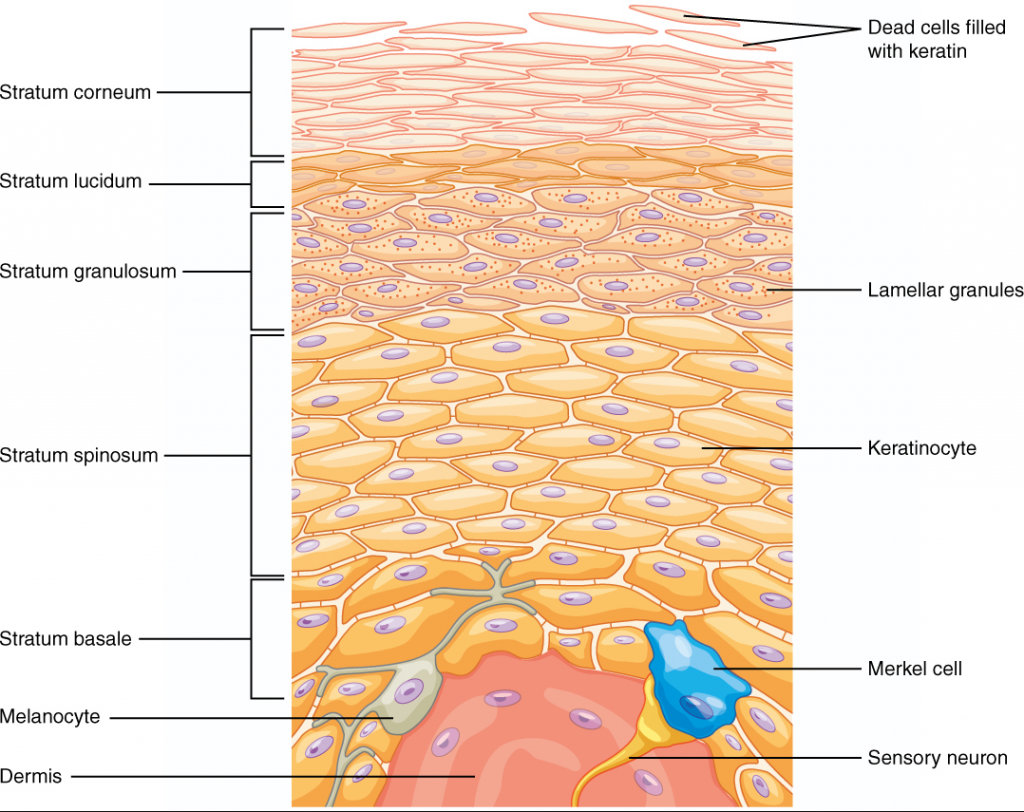
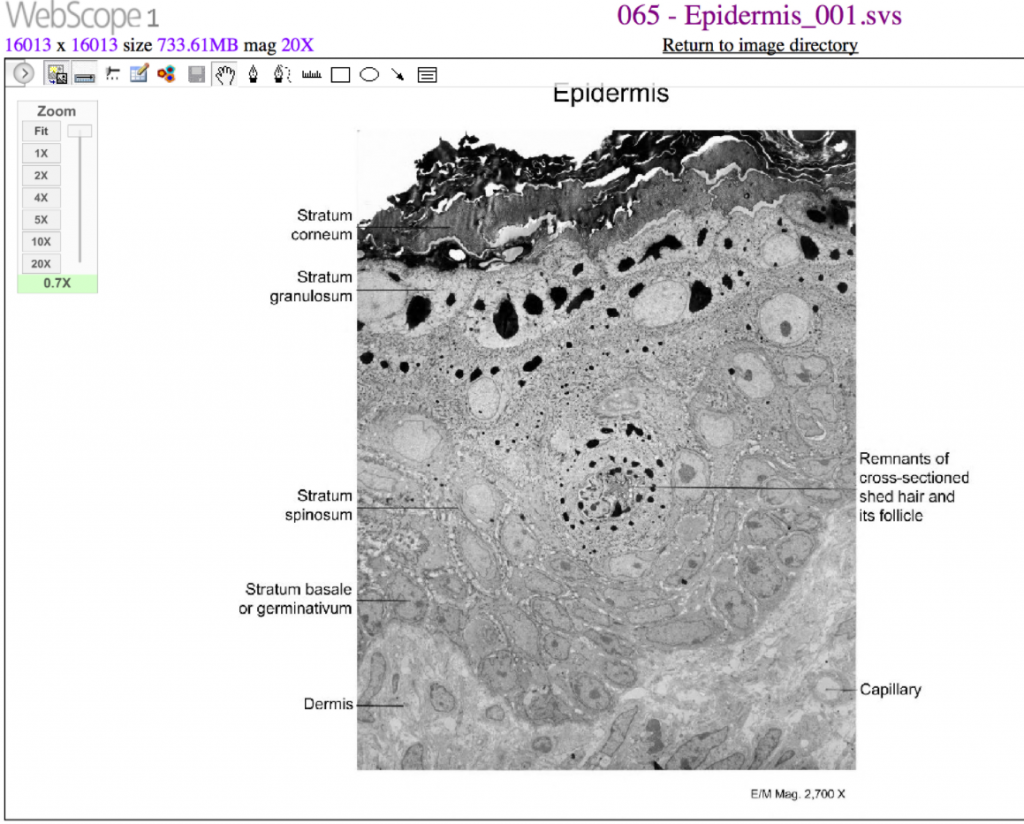
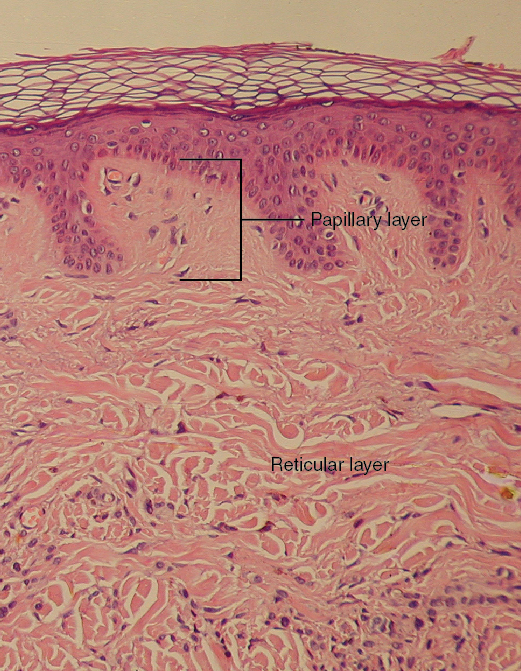
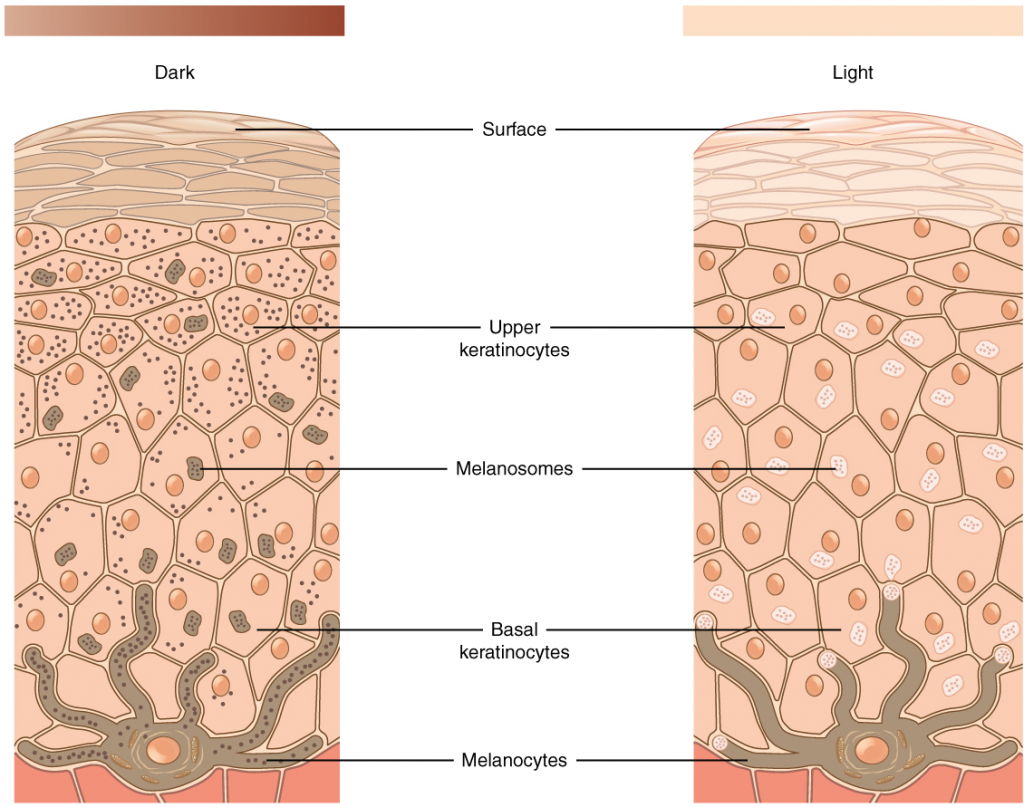
Integumentary System
Blood
Cardiovascular system
Lymphatic system and immunity
Respiratory systems
Muscle system
Skeletal system
Musculoskeletal system
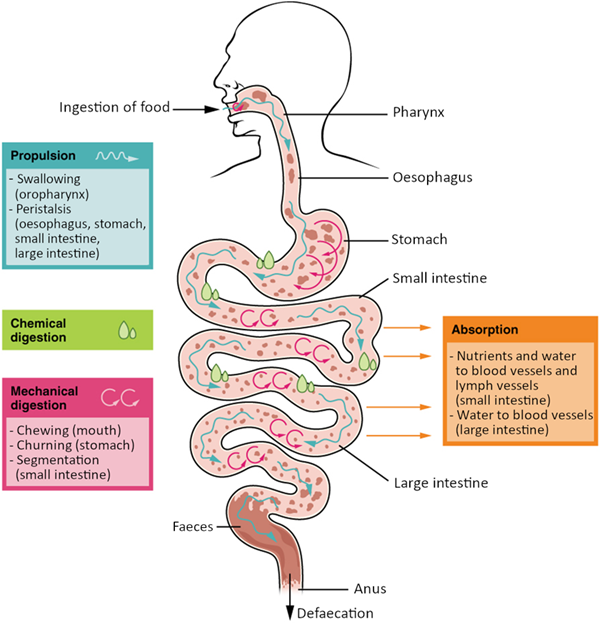
Digestive system
Nervous system
Endocrine system
Reproductive System
Pregnancy and human development
Urinary system
Levels of Organisation, Homeostasis and Nomenclature
I
1.1 Overview of Anatomy and Physiology
Learning Objectives
By the end of this section you will be able to:
Compare and contrast anatomy and physiology, including their specialisations and methods of study
Discuss the fundamental relationship between anatomy and physiology
Human anatomy is the scientific study of the body’s structures. Some of these structures are very small and can only be observed and analysed with the assistance of a microscope. Other larger structures can readily be seen, manipulated, measured, and weighed. The word “anatomy” comes from a Greek root that means “to cut apart.” Human anatomy was first studied by observing the exterior of the body and observing the wounds of soldiers and other injuries. Later, physicians could dissect bodies of the dead anatomy to augment their knowledge. When a body is dissected, its structures are cut apart in order to observe their physical attributes and their relationships to one another. Dissection is still used in medical schools, anatomy courses, and in pathology labs. In order to observe structures in living people, however, several imaging techniques have been developed. These techniques allow clinicians to visualise structures inside the living body such as a cancerous tumour or a fractured bone.
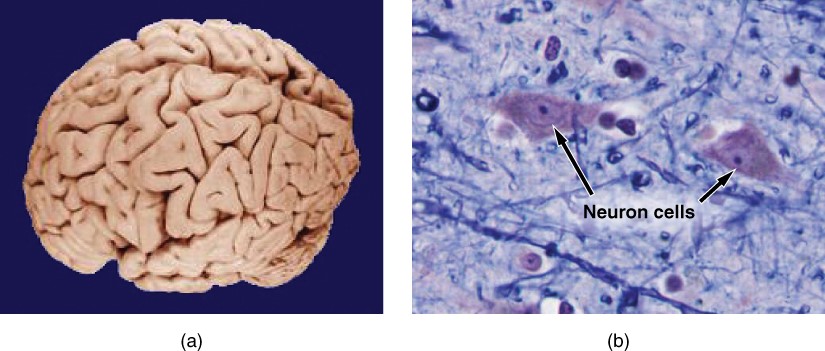
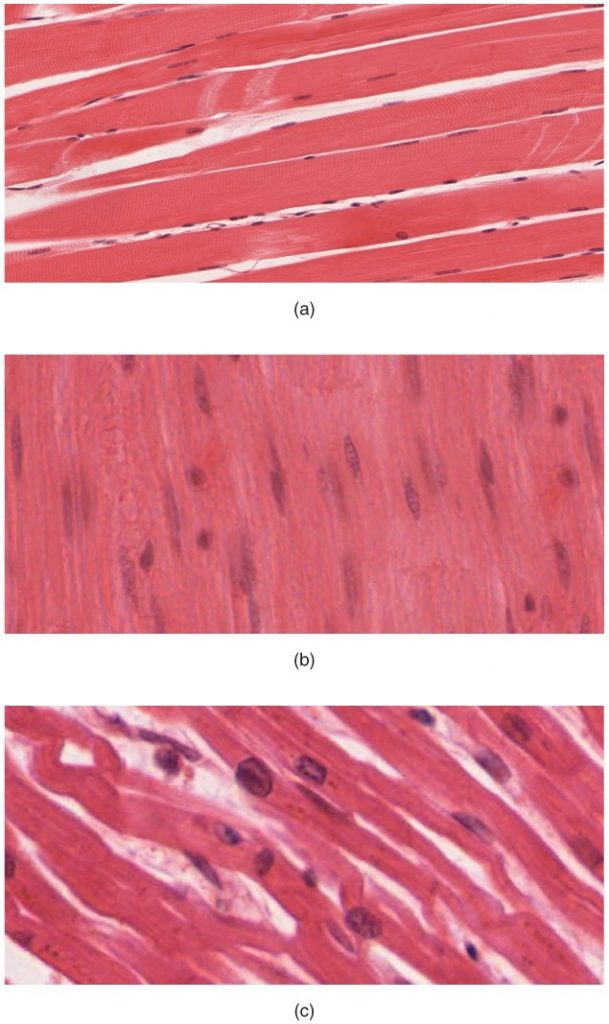
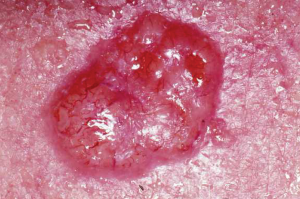
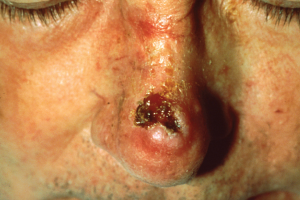
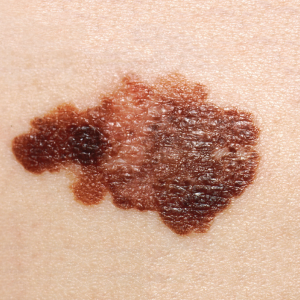
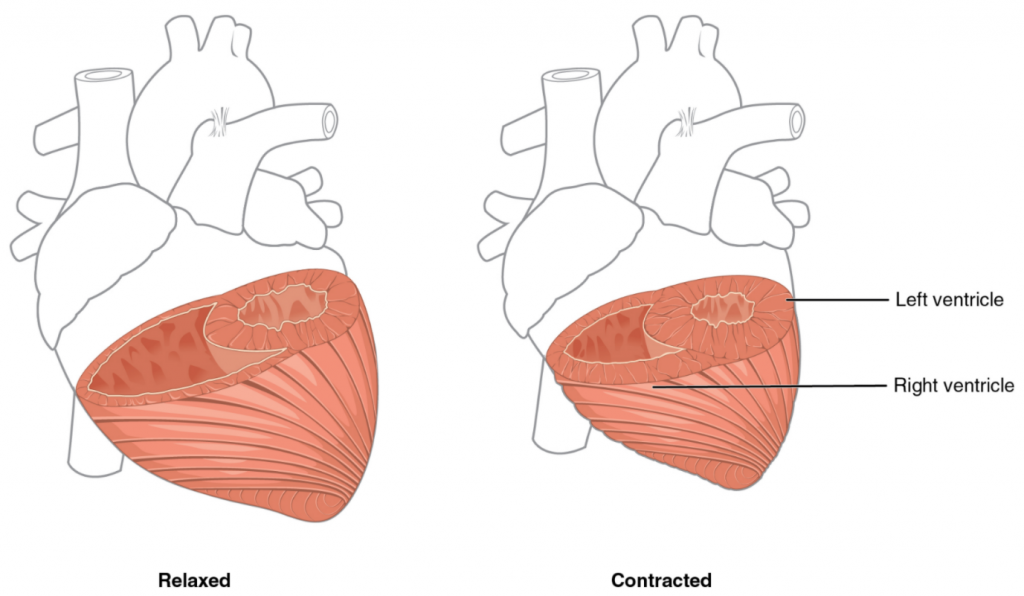
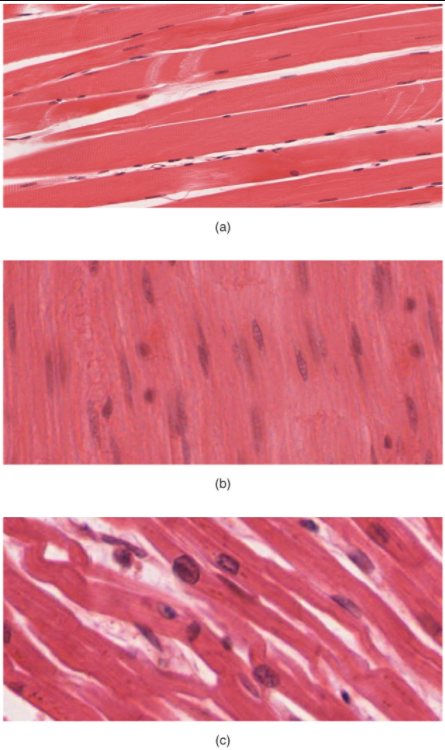
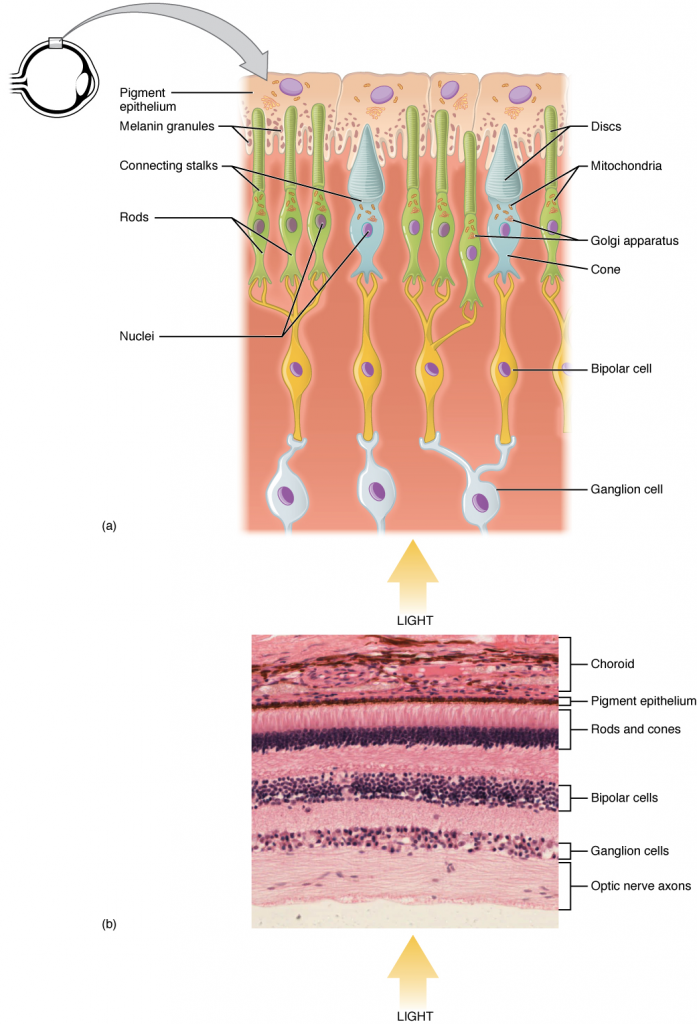
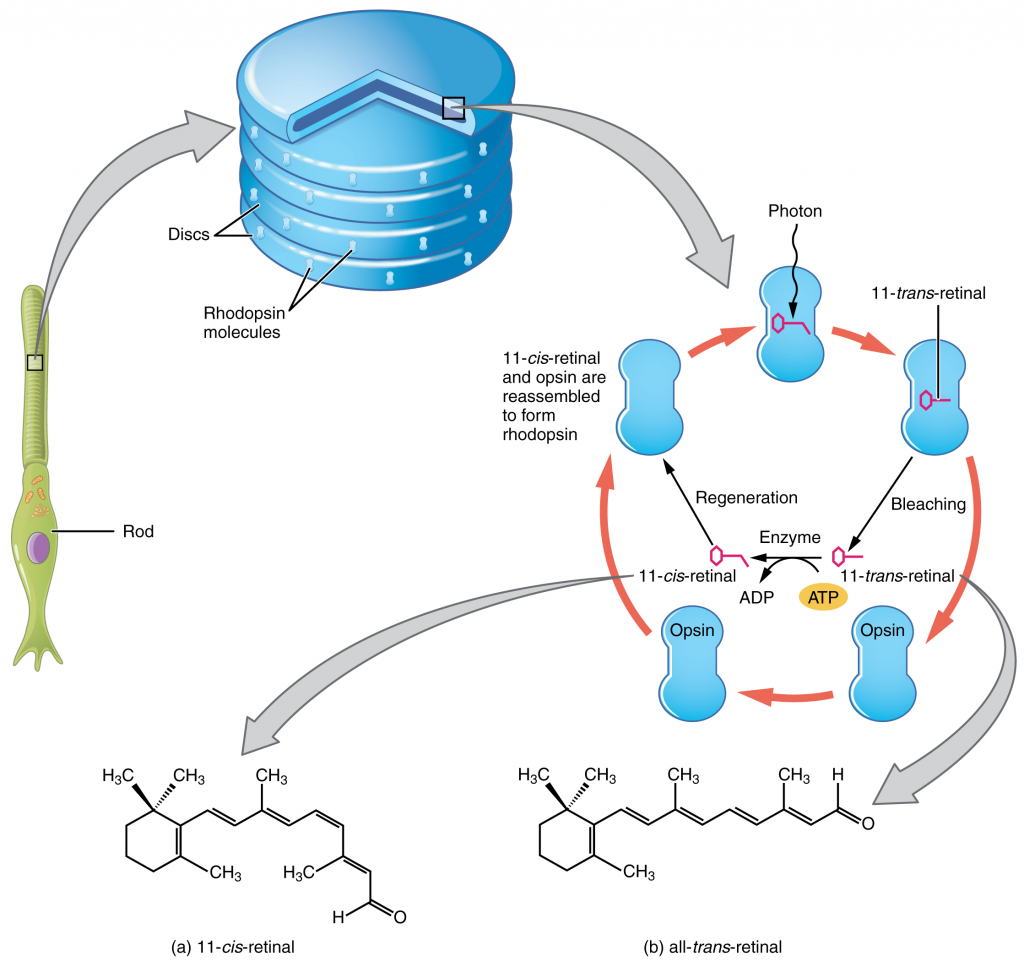
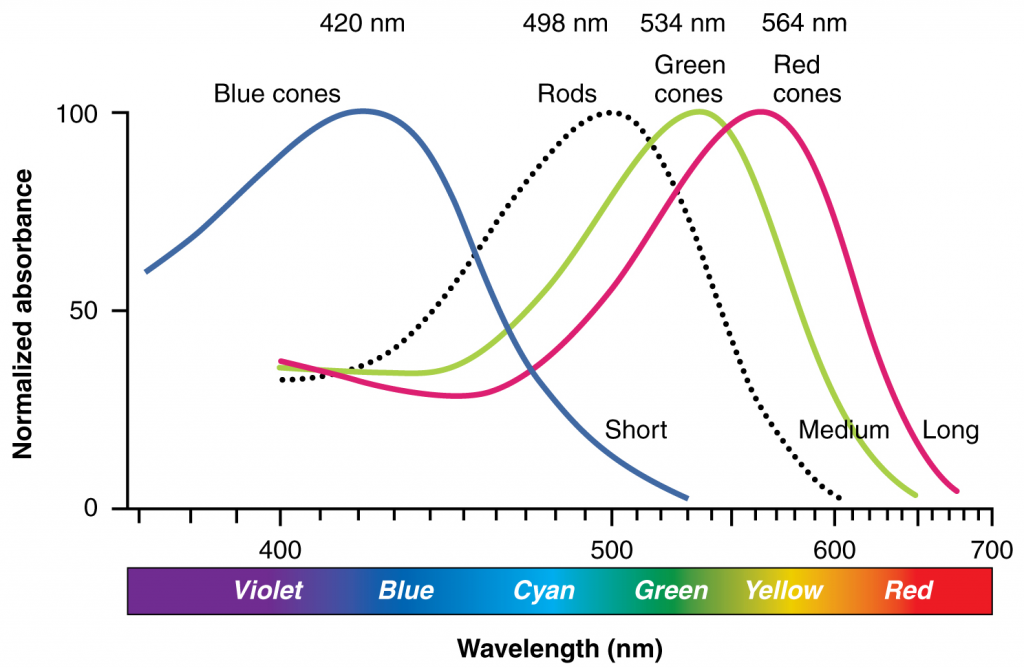
Like most scientific disciplines, anatomy has areas of specialisation. Gross anatomy is the study of the larger structures of the body, those visible without the aid of magnification (Figure 1.1.1a). Macro- means “large,” thus, gross anatomy is also referred to as macroscopic anatomy. In contrast, micro- means “small,” and microscopic anatomy is the study of structures that can be observed only with the use of a microscope or other magnification devices (Figure 1.1.1b). Microscopic anatomy includes cytology, the study of cells and histology, the study of tissues. As the technology of microscopes has advanced, anatomists have been able to observe smaller and smaller structures of the body, from slices of large structures like the heart, to the three-dimensional structures of large molecules in the body.
Anatomists take two general approaches to the study of the body’s structures: regional and systemic. Regional anatomy is the study of the interrelationships of all the structures in a specific body region, such as the abdomen. Studying regional anatomy helps us appreciate the interrelationships of body structures, such as how muscles, nerves, blood vessels, and other structures work together to serve a particular body region. In contrast, systemic anatomy is the study of the structures that make up a discrete body system—that is, a group of structures that work together to perform a unique body function, for example, a systemic anatomical study of the muscular system would consider all the skeletal muscles of the body.
Whereas anatomy is about structure, physiology is about function. Human physiology is the scientific study of the chemistry and physics of the structures of the body and the ways in which they work together to support the functions of life. Much of the study of physiology centres on the body’s tendency toward homeostasis. Homeostasis is the state of steady internal conditions maintained by living things. The study of physiology certainly includes observation, both with the naked eye and with microscopes, as well as manipulations and measurements. However, current advances in physiology usually depend on carefully designed laboratory experiments that reveal the functions of the many structures and chemical compounds that make up the human body.
Like anatomists, physiologists typically specialise in a particular branch of physiology, for example, neurophysiology is the study of the brain, spinal cord, and nerves and how they work together to perform functions as complex and diverse as vision, movement, and thinking. Physiologists may work from the organ level (exploring, for example, what different parts of the brain do) to the molecular level (such as exploring how an electrochemical signal travels along nerves).
Form is closely related to function in all living things, for example, the thin flap of your eyelid can snap down to clear away dust particles and almost instantaneously slide back up to allow you to see again. At the microscopic level, the arrangement and function of the nerves and muscles that serve the eyelid allow for its quick action and retreat. At a smaller level of analysis, the function of these nerves and muscles likewise relies on the interactions of specific molecules and ions. Even the three-dimensional structure of certain molecules is essential to their function.
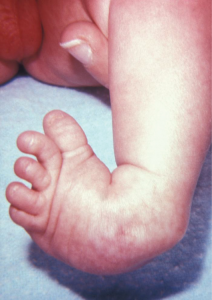
Your study of anatomy and physiology will make more sense if you continually relate the form of the structures you are studying to their function. In fact, it can be somewhat frustrating to attempt to study anatomy without an understanding of the physiology that a body structure supports. Imagine, for example, trying to appreciate the unique arrangement of the bones of the human hand if you had no conception of the function of the hand. Fortunately, your understanding of how the human hand manipulates tools—from pens to cell phones—helps you appreciate the unique alignment of the thumb in opposition to the four fingers, making your hand a structure that allows you to pinch and grasp objects and type text messages.
Section Review
Human anatomy is the scientific study of the body’s structures. In the past, anatomy has primarily been studied via observing injuries, and later by the dissection of anatomical structures of cadavers, but in the past century, computer-assisted imaging techniques have allowed clinicians to look inside the living body. Human physiology is the scientific study of the chemistry and physics of the structures of the body. Physiology explains how the structures of the body work together to maintain life. It is difficult to study structure (anatomy) without knowledge of function (physiology). The two disciplines are typically studied together because form and function are closely related in all living things.
Describe the structure of the human body in terms of six levels of organisation
List the eleven organ systems of the human body and identify at least one organ and one major function of each
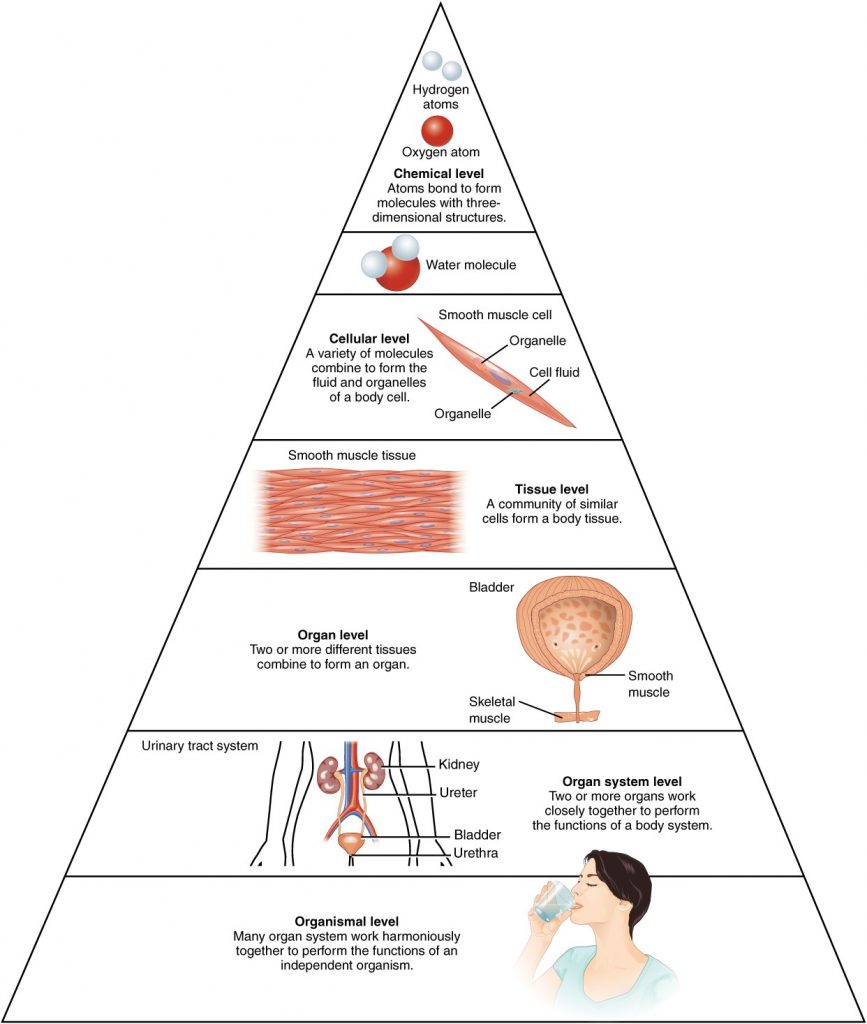
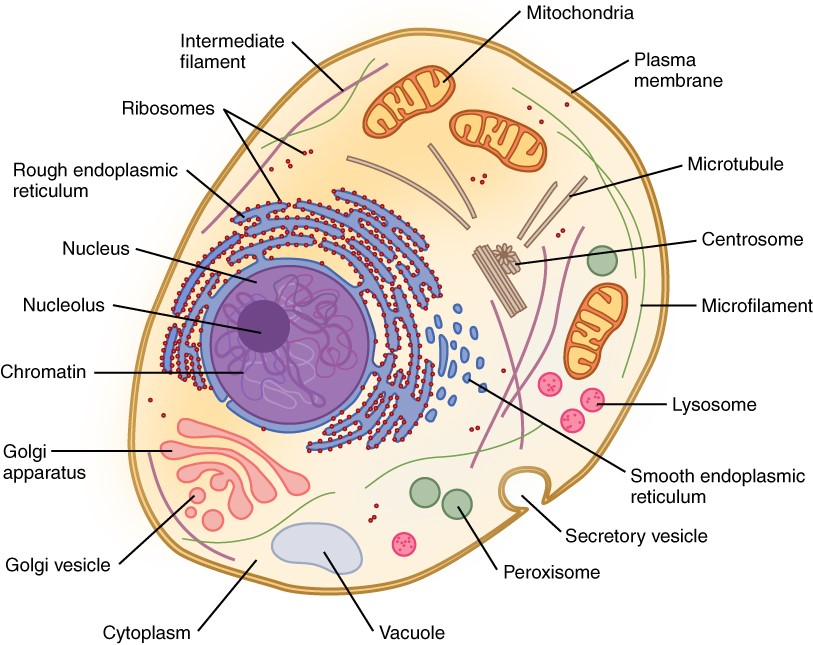
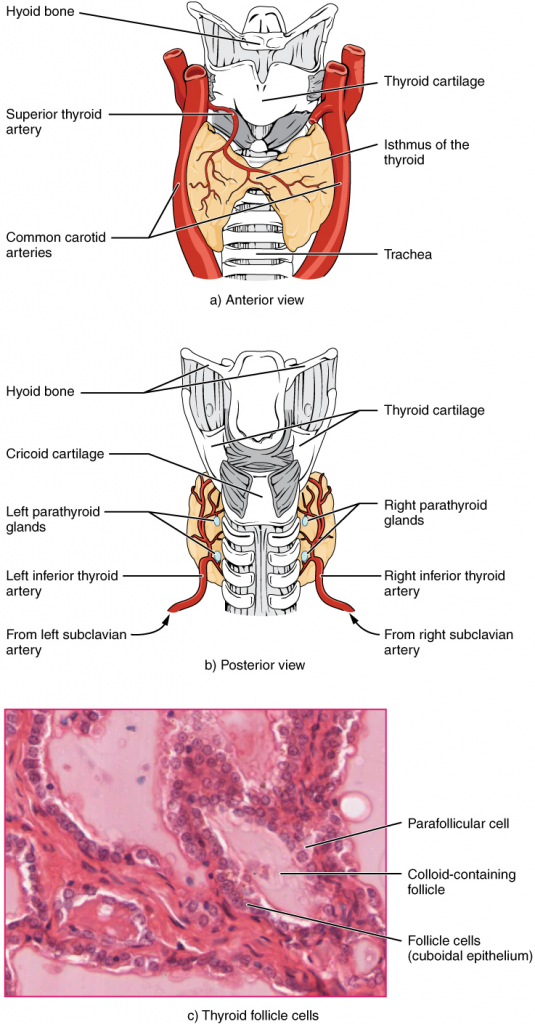
Before you begin to study the different structures and functions of the human body, it is helpful to consider its basic architecture; that is, how its smallest parts are assembled into larger structures. It is convenient to consider the structures of the body in terms of fundamental levels of organisation that increase in complexity: subatomic particles, atoms, molecules, organelles, cells, tissues, organs, organ systems, organisms and biosphere (Figure 1.2.1).
Figure 1.2.1. Levels of structural organisation of the human body. The organisation of the body often is discussed in terms of six distinct levels of increasing complexity, from the smallest chemical building blocks to a unique human organism.
The Levels of Organisation
To study the chemical level of organisation, scientists consider the simplest building blocks of matter: subatomic particles, atoms and molecules. All matter in the universe is composed of one or more unique pure substances called elements, familiar examples of which are hydrogen, oxygen, carbon, nitrogen, calcium, and iron. The smallest unit of any of these pure substances (elements) is an atom. Atoms are made up of subatomic particles such as the proton, electron and neutron. Two or more atoms combine to form a molecule, such as the water molecules, proteins, and sugars found in living things. Molecules are the chemical building blocks of all body structures.
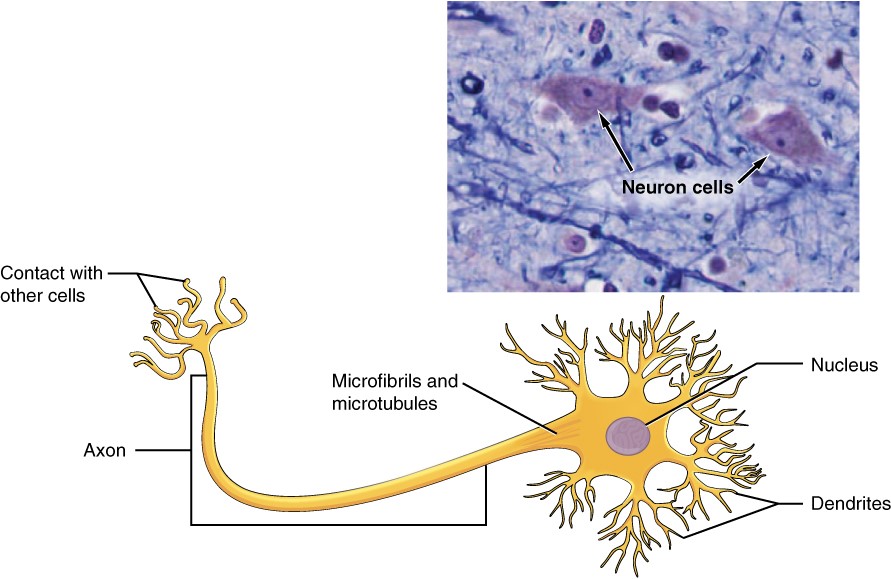
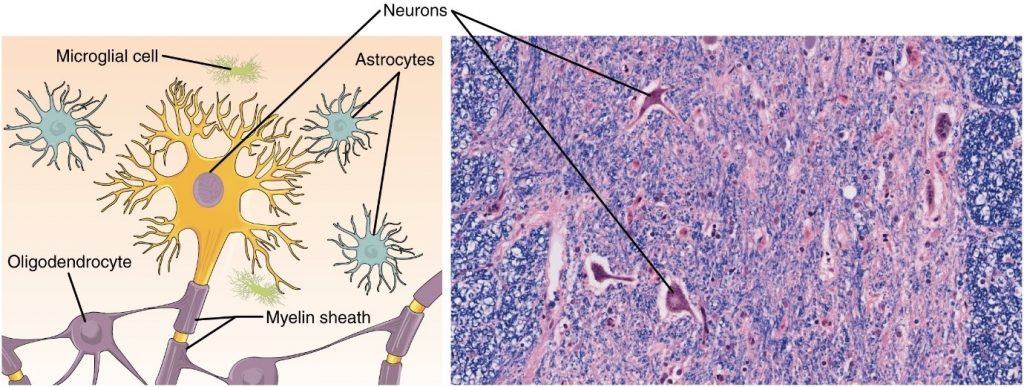
A cell is the smallest independently functioning unit of a living organism. Even bacteria, which are extremely small, independently-living organisms, have a cellular structure. Each bacterium is a single cell. All living structures of human anatomy contain cells, and almost all functions of human physiology are performed in cells or are initiated by cells.
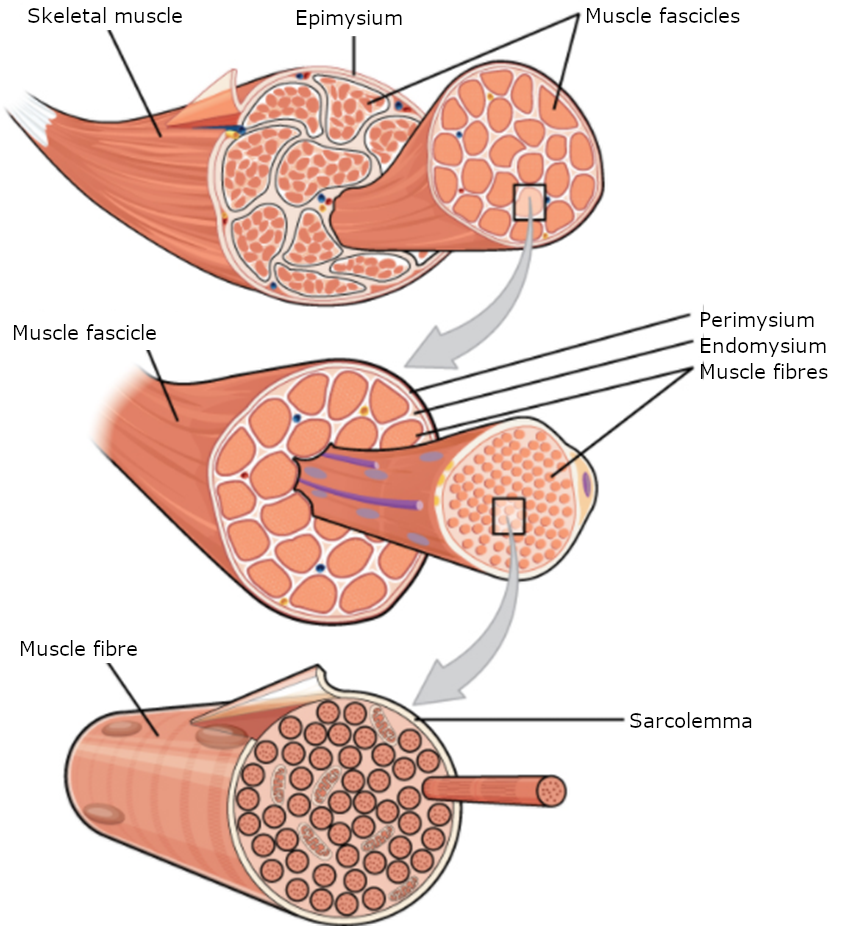
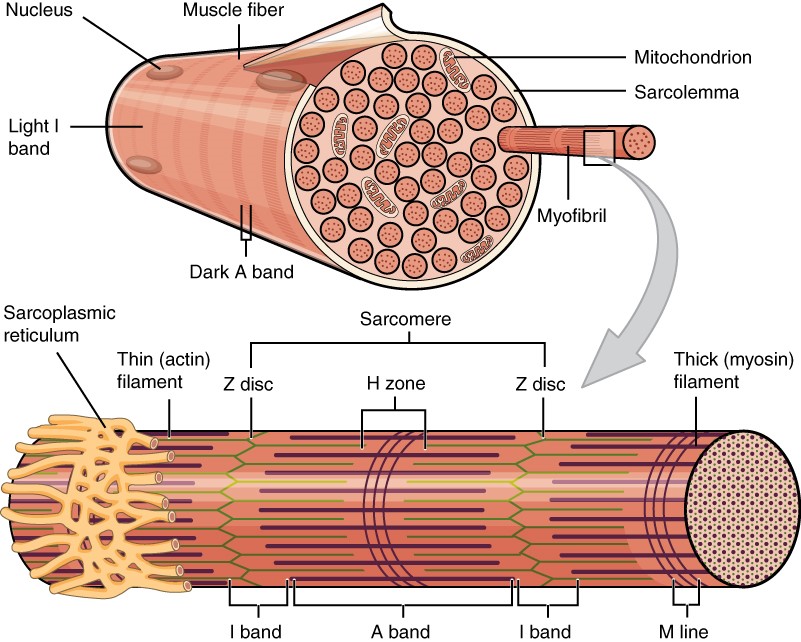
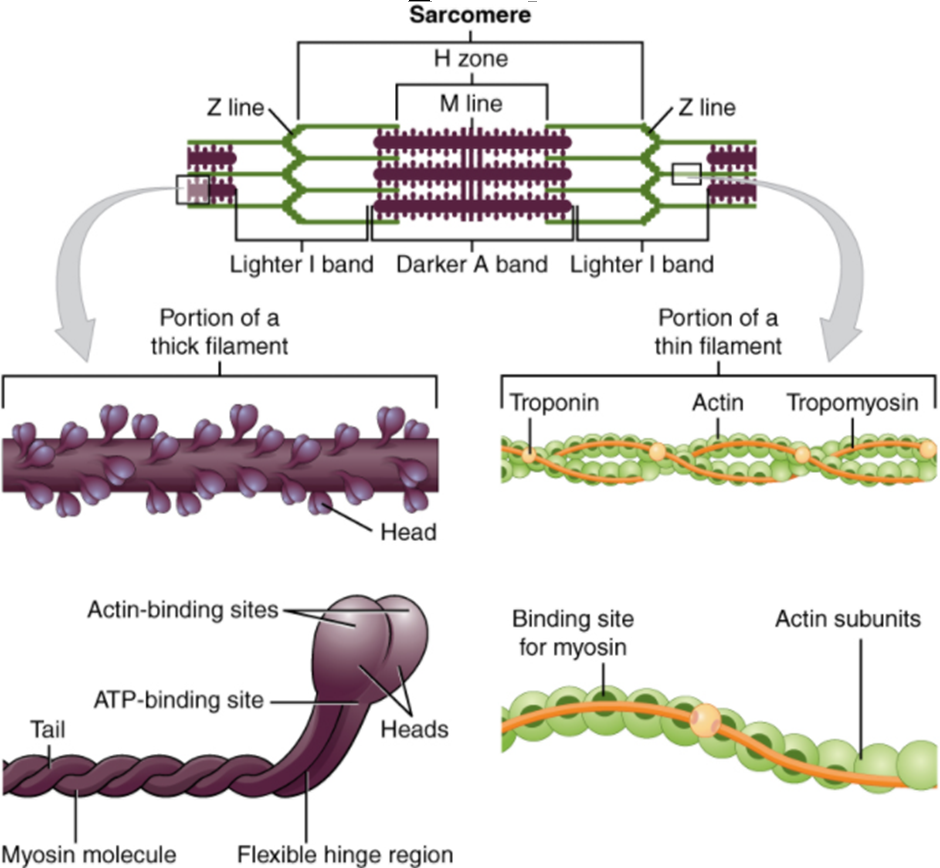
A human cell typically consists of flexible membranes that enclose cytoplasm, a water-based cellular fluid together with a variety of tiny functioning units called organelles. In humans, as in all organisms, cells perform all functions of life. A tissue is a group of many similar cells (though sometimes composed of a few related types) that work together to perform a specific function. An organ is an anatomically distinct structure of the body composed of two or more tissue types. Each organ performs one or more specific physiological functions. An organ system is a group of organs that work together to perform major functions or meet physiological needs of the body.
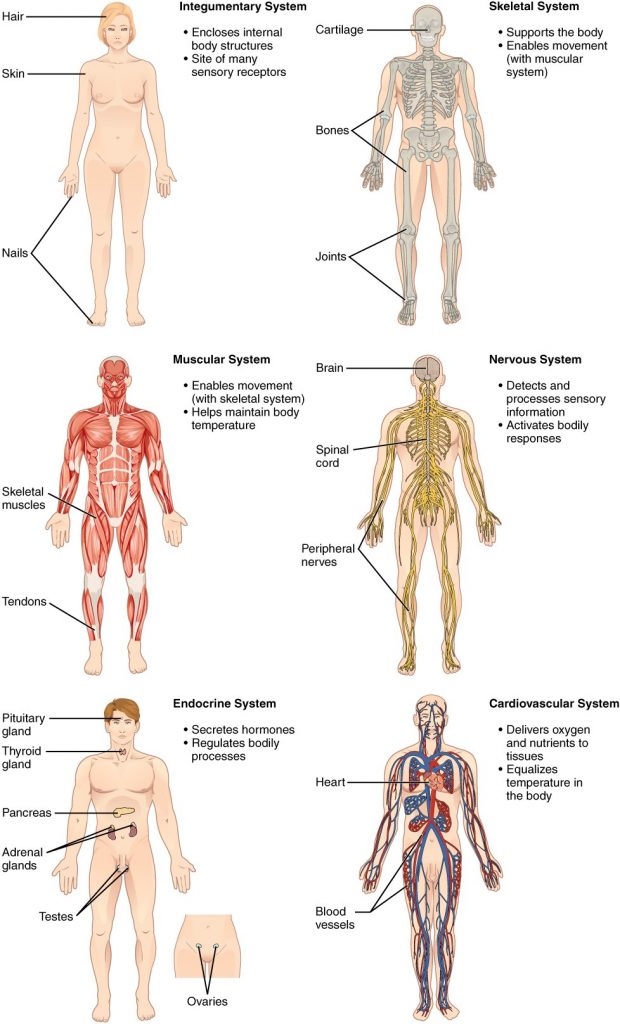
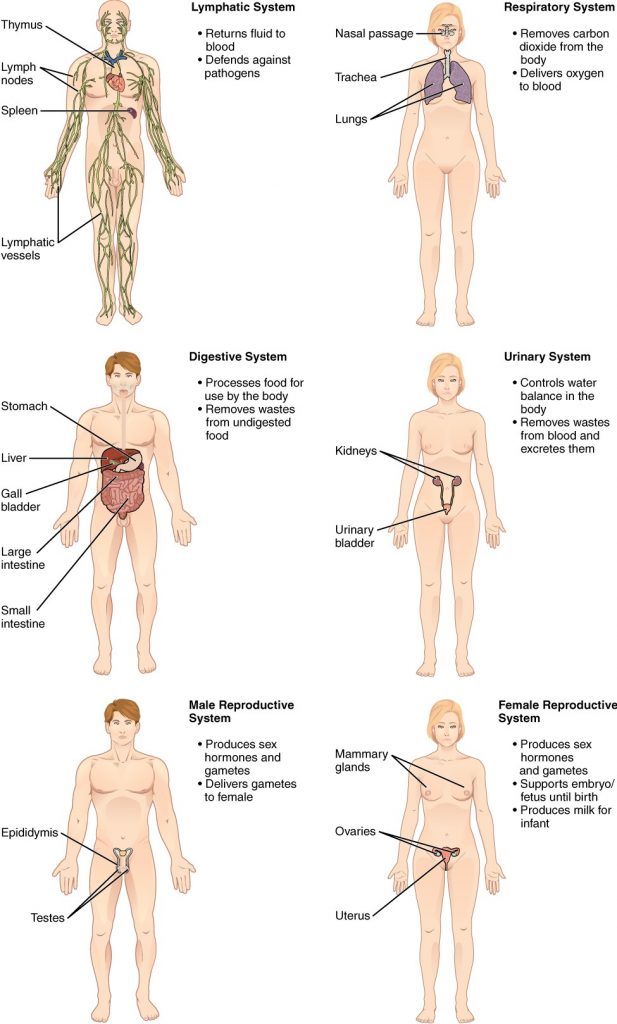
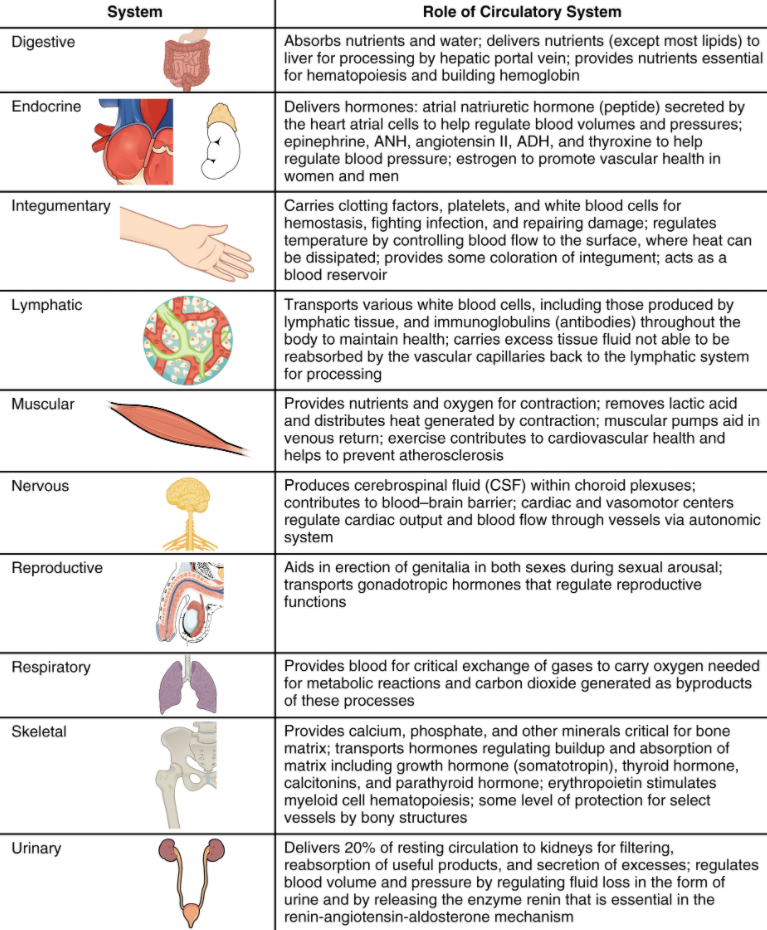
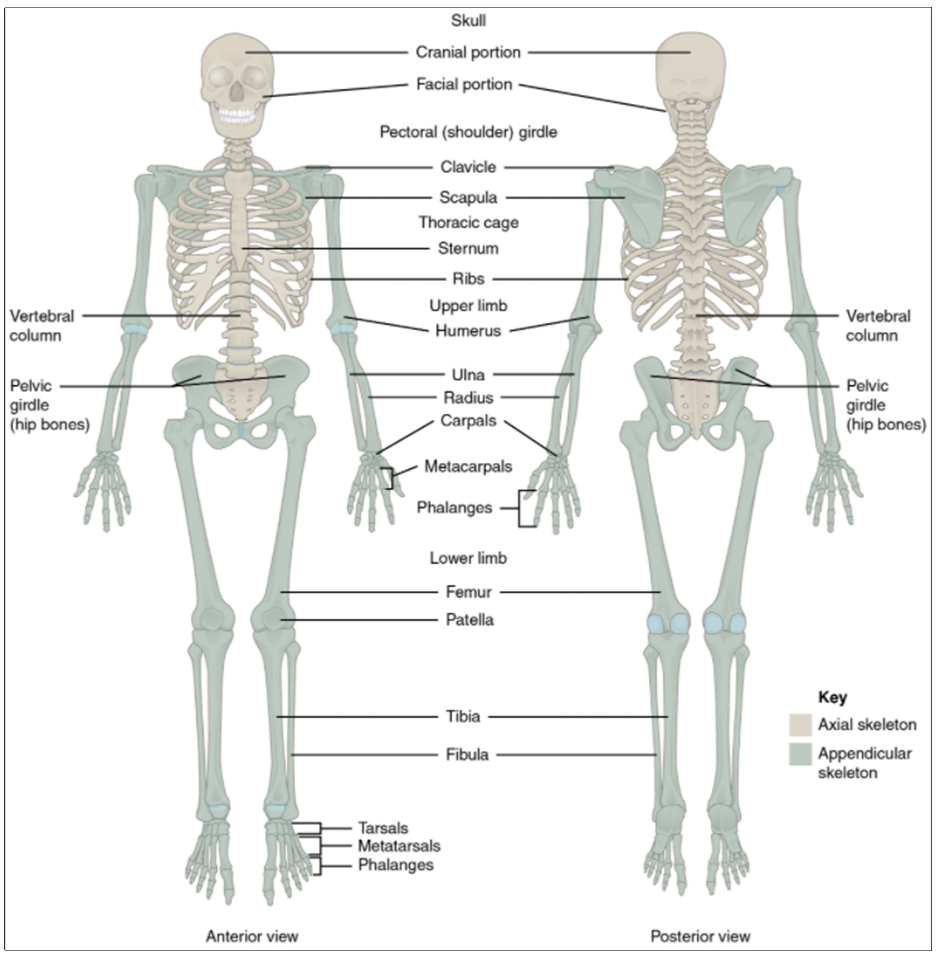
This book covers eleven distinct organ systems in the human body (Figure 1.2.2 and Figure 1.2.3). Assigning organs to organ systems can be imprecise since organs that “belong” to one system can also have functions integral to another system. In fact, most organs contribute to more than one system.
Figure 1.2.2. Organ systems of the human body. Organs that work together are grouped into organ systems.
Figure 1.2.3. Organ systems of the human body (continued). Organs that work together are grouped into organ systems.
The organism level is the highest level of organisation. An organism is a living being that has a cellular structure and that can independently perform all physiologic functions necessary for life. In multicellular organisms, including humans, all cells, tissues, organs, and organ systems of the body work together to maintain the life and health of the organism.
Section Review
Life processes of the human body are maintained at several levels of structural organisation. These include the chemical, cellular, tissue, organ, organ system, and the organism level. Higher levels of organisation are built from lower levels. Therefore, molecules combine to form cells, cells combine to form tissues, tissues combine to form organs, organs combine to form organ systems and organ systems combine to form organisms.
Discuss the role of homeostasis in healthy functioning
Recognise negative and positive feedback, giving one physiologic example of each mechanism
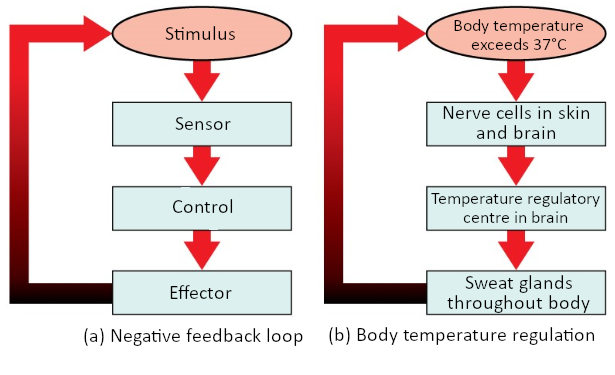
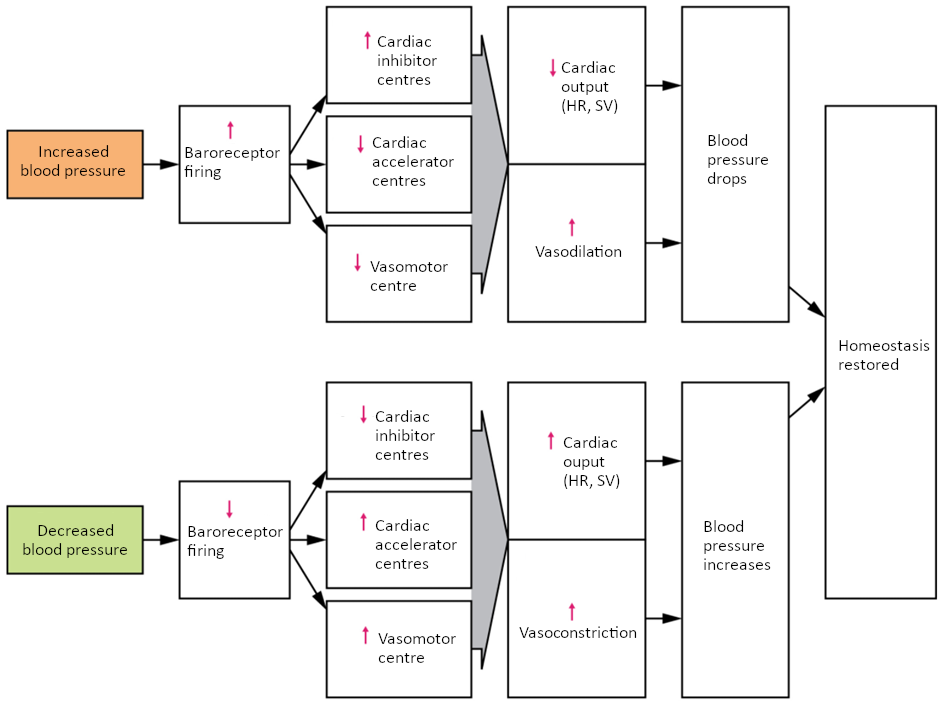
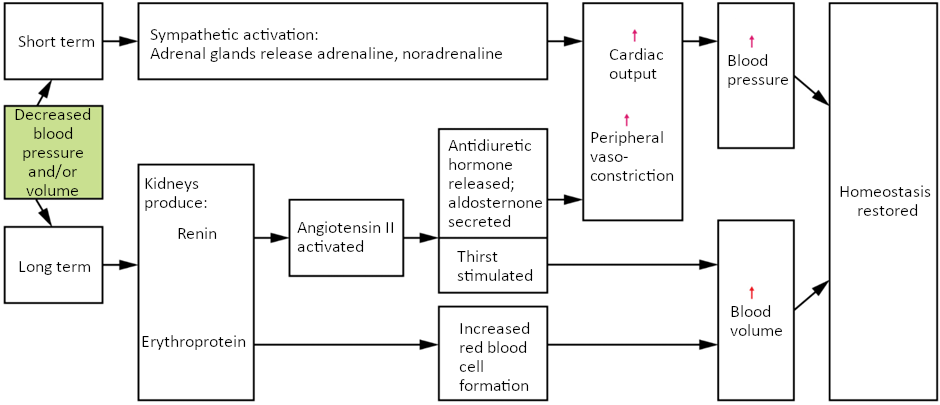
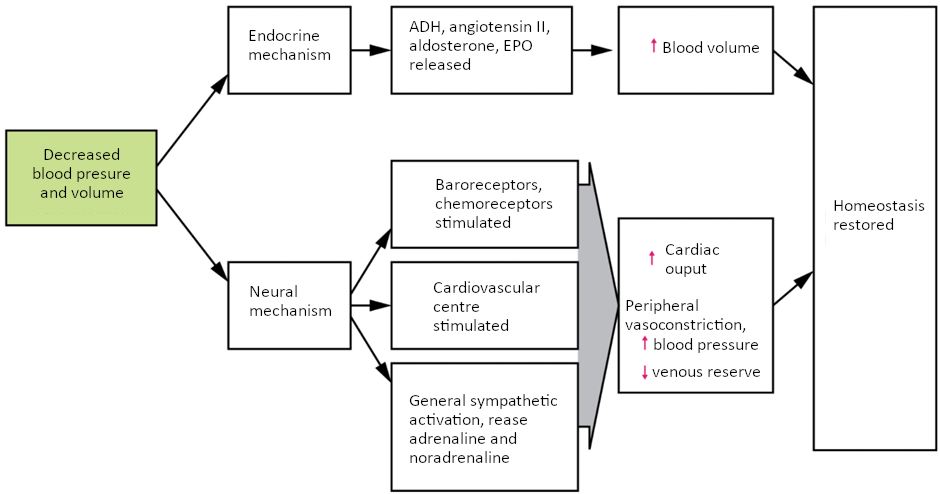
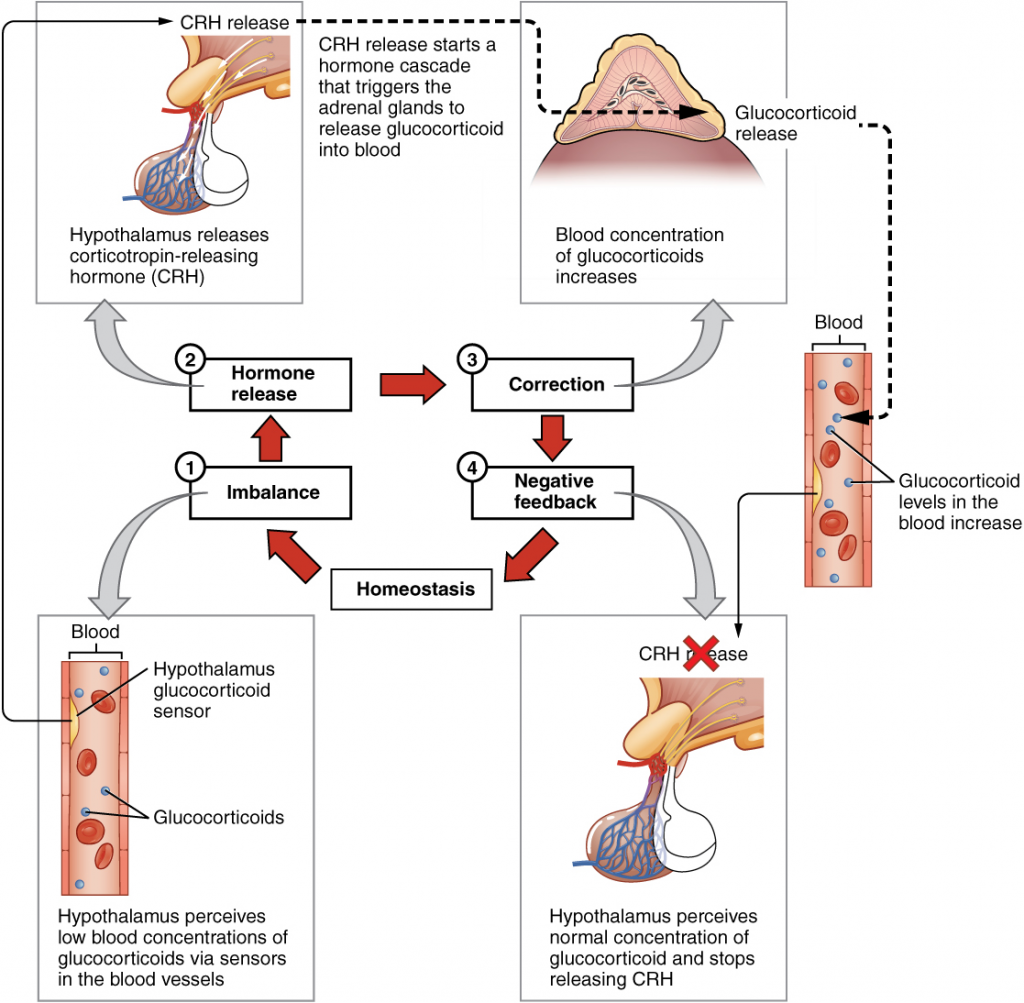
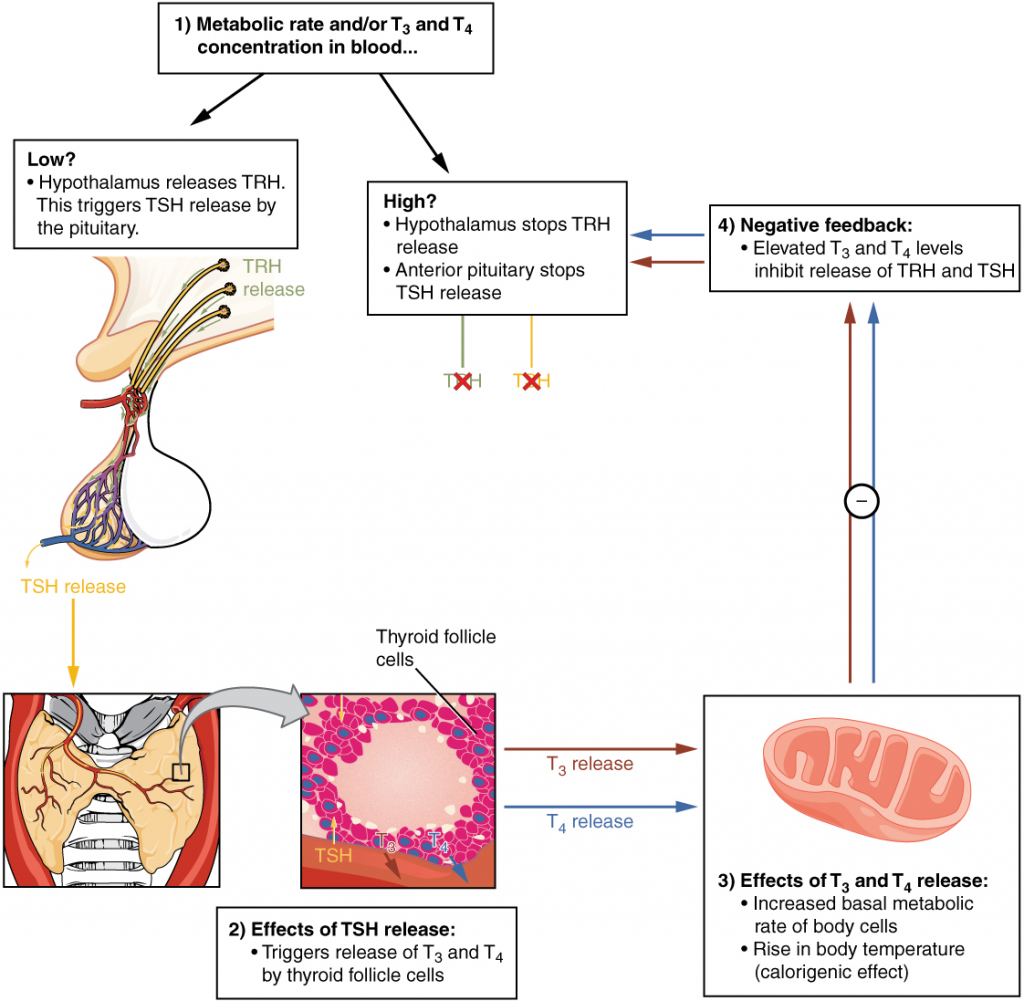
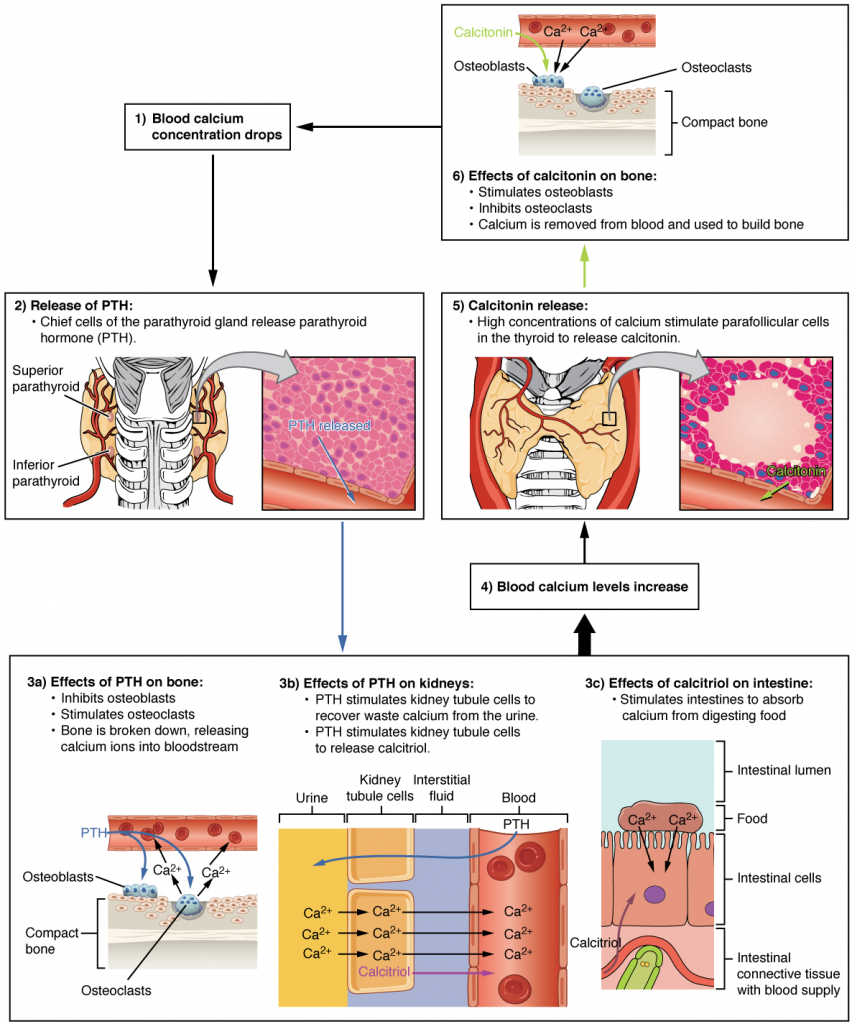
Maintaining homeostasis requires that the body continuously monitor its internal conditions. From body temperature to blood pressure to levels of certain nutrients, each physiological condition has a particular set point. A set point is the physiological value around which the normal range fluctuates. A normal range is the restricted set of values that is optimally healthful and stable. For example, the set point for normal human body temperature is approximately 37°C (98.6°F) Physiological parameters, such as body temperature and blood pressure, tend to fluctuate within a normal range a few degrees above and below that point. Control centres in the brain and other parts of the body monitor and react to deviations from homeostasis using negative feedback. Negative feedback is a mechanism that reverses a deviation from the set point. Therefore, negative feedback maintains body parameters within their normal range. The maintenance of homeostasis by negative feedback goes on throughout the body, at all times, and an understanding of negative feedback is thus fundamental to an understanding of human physiology.
Negative Feedback
A negative feedback system has three basic components (Figure 1.3.1). A sensor, also referred to a receptor, is a component of a feedback system that monitors a physiological value. This value is reported to the control centre. The control centre is the component in a feedback system that compares the value to the normal range. If the value deviates too much from the set point, then the control centre activates an effector. An effector is the component in a feedback system that causes a change to reverse the situation and return the value to the normal range.
Figure 1.3.1. Negative feedback loop. In a negative feedback loop, a stimulus—a deviation from a set point—is resisted through a physiological process that returns the body to homeostasis. (a) A negative feedback loop has four basic parts. (b) Body temperature is regulated by negative feedback.
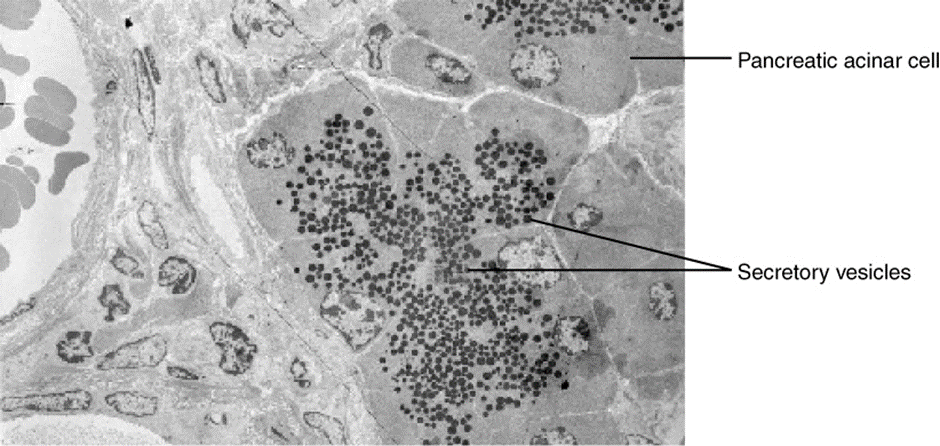
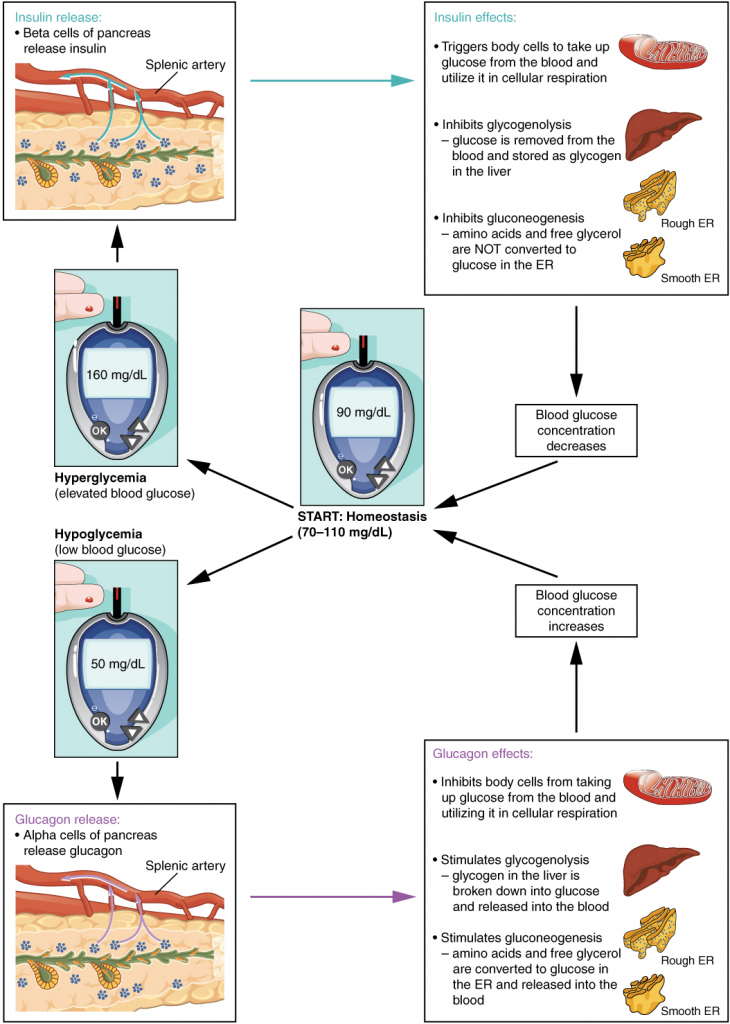
In order to set the system in motion, a stimulus must drive a physiological parameter beyond its normal range (that is, beyond homeostasis). This stimulus is “heard” by a specific sensor. For example, in the control of blood glucose, specific endocrine cells in the pancreas detect excess glucose (the stimulus) in the bloodstream. These pancreatic beta cells respond to the increased level of blood glucose by increased secretion of the hormone insulin into the bloodstream (ie more than the normal basal secretion). The insulin signals skeletal muscle fibres, fat cells (adipocytes), and liver cells to take up the excess glucose, removing it from the bloodstream. As glucose concentration in the bloodstream drops, the decrease in concentration—the actual negative feedback—is detected by pancreatic alpha cells, and insulin release stops. This prevents blood glucose levels from continuing to drop below the normal range.
Humans have a similar temperature regulation feedback system that works by promoting either heat loss or heat gain (Figure 1.3.1b). When the brain’s temperature regulation centre receives data from the sensors indicating that the body’s temperature exceeds its normal range, it stimulates a cluster of brain cells referred to as the “heat-loss centre.” This stimulation has three major effects:
Blood vessels in the skin begin to dilate allowing more blood from the body core to flow to the surface of the skin allowing the heat to radiate into the environment.
As blood flow to the skin increases, sweat glands are activated to increase their output. As the sweat evaporates from the skin surface into the surrounding air, it takes heat with it.
The depth of respiration increases, and a person may breathe through an open mouth instead of through the nasal passageways. This further increases heat loss from the lungs.
In contrast, activation of the brain’s heat-gain centre by exposure to cold reduces blood flow to the skin, and blood returning from the limbs is diverted into a network of deep veins. This arrangement traps heat closer to the body core and restricts heat loss. If heat loss is severe, the brain triggers an increase in random signals to skeletal muscles, causing them to contract and producing shivering. The muscle contractions of shivering release heat while using up ATP. The brain triggers the thyroid gland in the endocrine system to release thyroid hormone, which increases metabolic activity and heat production in cells throughout the body. The brain also signals the adrenal glands to release adrenaline (US: epinephrine), a hormone that causes the breakdown of glycogen into glucose, which can be used as an energy source. The breakdown of glycogen into glucose also results in increased metabolism and heat production.
Positive Feedback
Positive feedback intensifies a change in the body’s physiological condition rather than reversing it. A deviation from the normal range results in more change, and the system moves farther away from the normal range. Positive feedback in the body is normal only when there is a definite end point. Childbirth and the body’s response to blood loss are two examples of positive feedback loops that are normal but are activated only when needed.
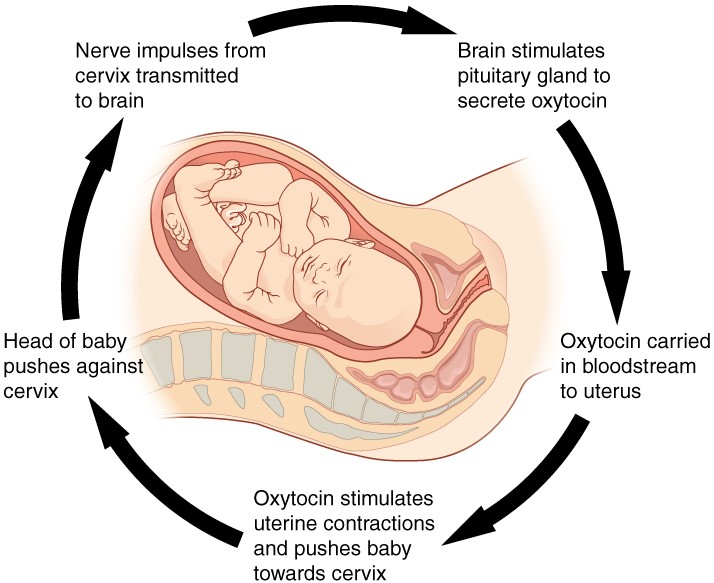
Childbirth at full term is an example of a situation in which the maintenance of the existing body state is not desired. Enormous changes in the mother’s body are required to expel the baby at the end of pregnancy, and the events of childbirth, once begun, must progress rapidly to a conclusion or the life of the mother and the baby are at risk. The extreme muscular work of labour and delivery are the result of a positive feedback system (Figure 1.3.2).
Figure 1.3.2. Positive feedback loop. Normal childbirth is driven by a positive feedback loop. A positive feedback loop results in a change in the body’s status, rather than a return to homeostasis.
The first contractions of labour (the stimulus) push the baby toward the cervix (the lowest part of the uterus). The cervix contains stretch-sensitive nerve cells that monitor the degree of stretching (the sensors). These nerve cells send messages to the brain, which in turn causes the pituitary gland at the base of the brain to release the hormone oxytocin into the bloodstream. Oxytocin causes stronger contractions of the smooth muscles in of the uterus (the effectors), pushing the baby further down the birth canal. This causes even greater stretching of the cervix. The cycle of stretching, oxytocin release, and increasingly more forceful contractions stops only when the baby is born. At this point, the stretching of the cervix halts, stopping the release of oxytocin.
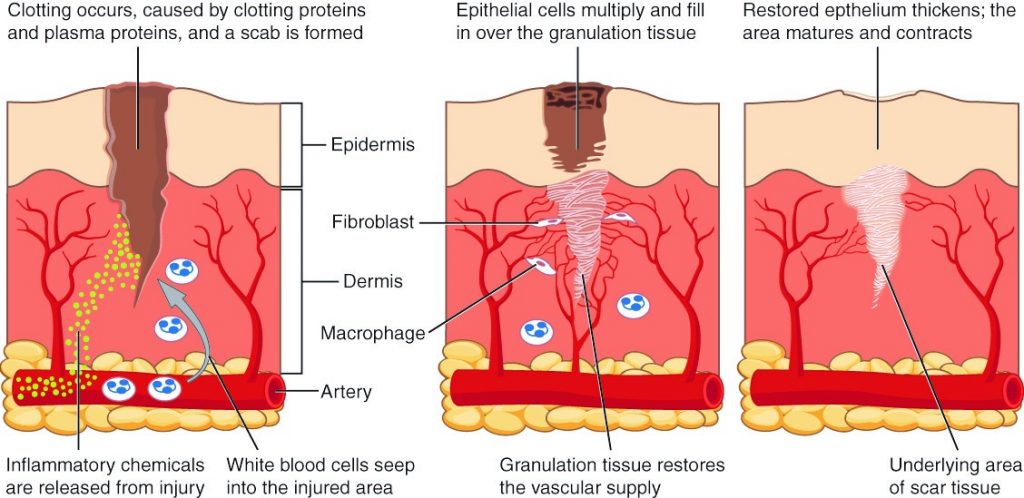
A second example of positive feedback centres on reversing extreme damage to the body. Following a penetrating wound, the most immediate threat is excessive blood loss. Less blood circulating means reduced blood pressure and reduced perfusion (penetration of blood) to the brain and other vital organs. If perfusion is severely reduced, vital organs will shut down and the person will die. The body responds to this potential catastrophe by releasing substances in the injured blood vessel wall that begin the process of blood clotting. As each step of clotting occurs, it stimulates the release of more clotting substances. This accelerates the processes of clotting and sealing off the damaged area. Clotting is contained in a local area based on the tightly controlled availability of clotting proteins. This is an adaptive, life-saving cascade of events.
Thermoregulation
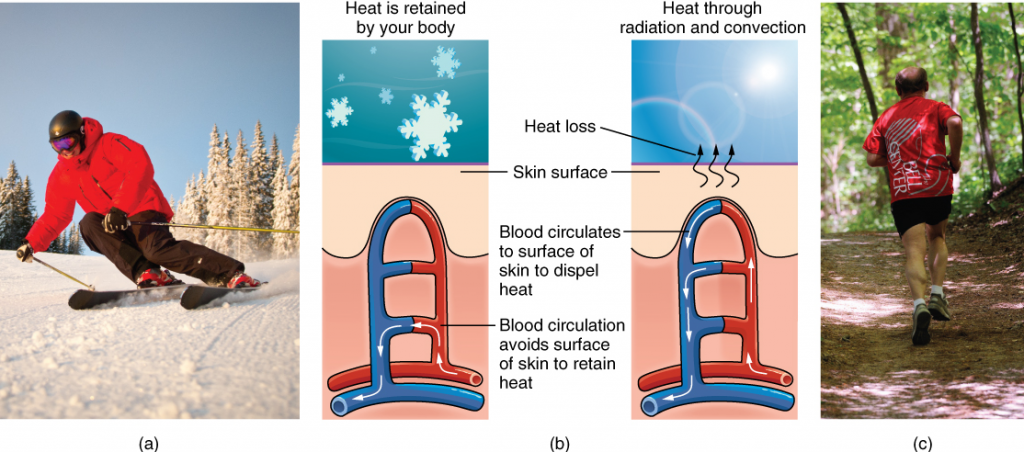
Animals, such as humans, that maintain a constant body temperature in the face of differing environmental temperatures, are called endotherms. We are able to maintain this temperature by generating internal heat (a waste product of the cellular chemical reactions of metabolism) that keeps the cellular processes operating optimally even when the environment is cold.
Endotherms use their circulatory systems to help maintain body temperature. Vasodilation, the opening up of arteries to the skin by relaxation of their smooth muscles, brings more blood and heat to the body surface, facilitating radiation and evaporative heat loss, cooling the body. Vasoconstriction, the narrowing of blood vessels to the skin by contraction of their smooth muscles, reduces blood flow in peripheral blood vessels, forcing blood toward the core and vital organs, conserving heat.
Thermoregulation is coordinated by the nervous system. The processes of temperature control are centred in a region of the brain called the hypothalamus. The hypothalamus maintains the set point for body temperature through reflexes that cause vasodilation or vasoconstriction and shivering or sweating. The hypothalamus directs the responses that effect the changes in temperature loss or gain that return the body to the set point. The set point may be adjusted in some instances. During an infection, compounds called pyrogens are produced and circulate to the hypothalamus resetting the thermostat to a higher value. This allows the body’s temperature to increase to a new homeostatic equilibrium point in what is commonly called a fever. The increase in body heat makes the body less optimal for bacterial growth and increases the activities of cells so they are better able to fight the infection.
Section Review
Homeostasis is the activity of cells throughout the body to maintain the physiological state within a narrow range that is compatible with life. Homeostasis is regulated by negative feedback loops and, much less frequently, by positive feedback loops. Both have the same components of a stimulus, sensor, control centre, and effector; however, negative feedback loops work to prevent an excessive response to the stimulus, whereas positive feedback loops intensify the response until an end point is reached.
Describe the human body using directional and regional terms
Identify three planes most commonly used in the study of anatomy
Recognise the posterior (dorsal) and the anterior (ventral) body cavities, identifying their subdivisions and representative organs found in each
Describe serous membranes and explain the functions
Anatomists and health care providers use terminology that can be bewildering to the uninitiated. However, the purpose of this language is not to confuse, but rather to increase precision and reduce medical errors. For example, is a scar “above the wrist” located on the forearm two or three inches away from the hand? Or is it at the base of the hand? Is it on the palm-side or back-side? By using precise anatomical terminology, we eliminate ambiguity. Anatomical terms derive from ancient Greek and Latin words. Because these languages are no longer used in everyday conversation, the meaning of their words does not change.
Anatomical terms are made up of roots, prefixes, and suffixes. The root of a term often refers to an organ, tissue, or condition, whereas the prefix or suffix often describes the root. For example, in the disorder hypertension, the prefix “hyper-” means “high” or “over,” and the root word “tension” refers to pressure, so the word “hypertension” refers to abnormally high blood pressure.
Anatomical Position
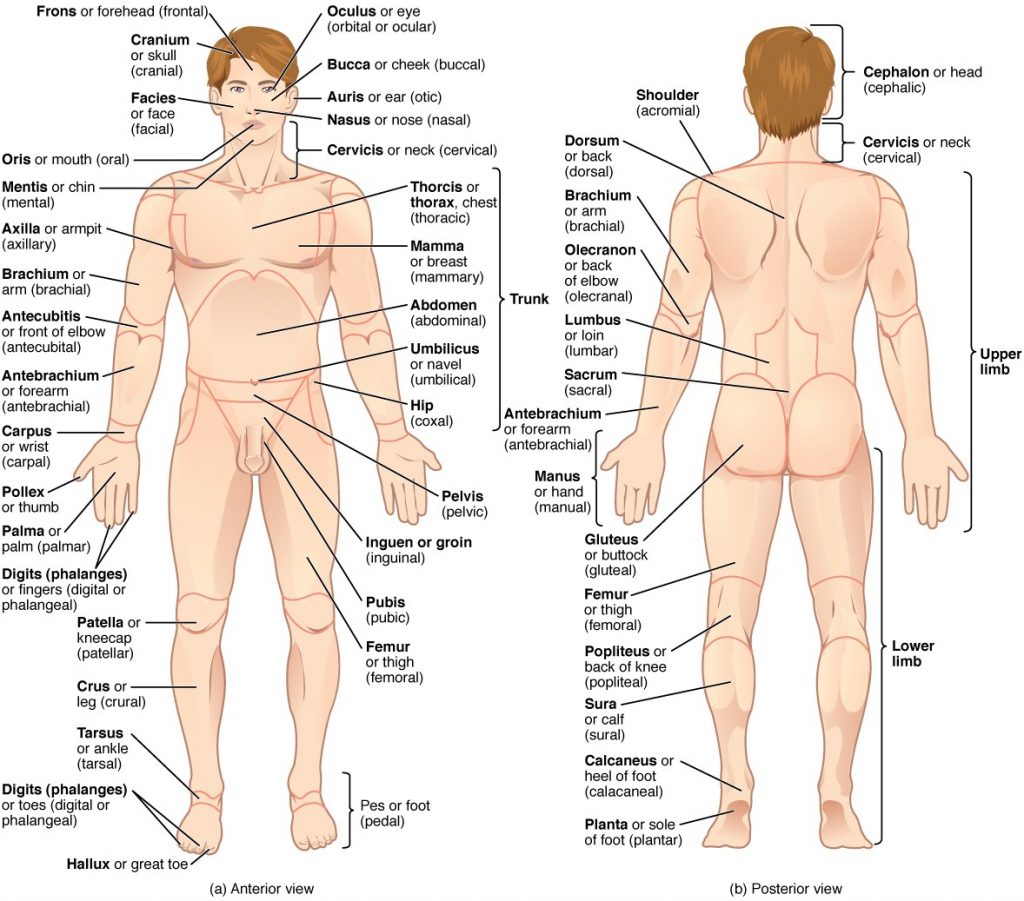
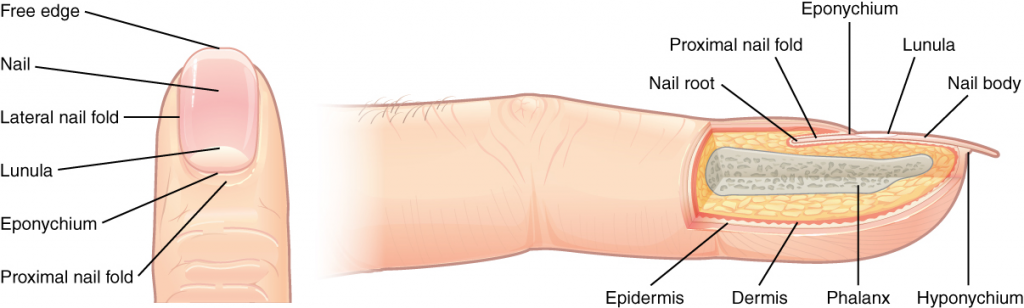
To further increase precision, anatomists standardise the way in which they view the body. Just as maps are normally oriented with north at the top, the standard body “map,” or anatomical position, is that of the body standing upright, with the feet at shoulder width and parallel, toes forward. The upper limbs are held out to each side, and the palms of the hands face forward as illustrated in Figure 1.4.1. Using this standard position reduces confusion. It does not matter how the body being described is oriented, the terms are used as if it is in anatomical position. For example, a scar in the “anterior (front) carpal (wrist) region” would be present on the palm side of the wrist. The term “anterior” would be used even if the hand were palm down on a table.
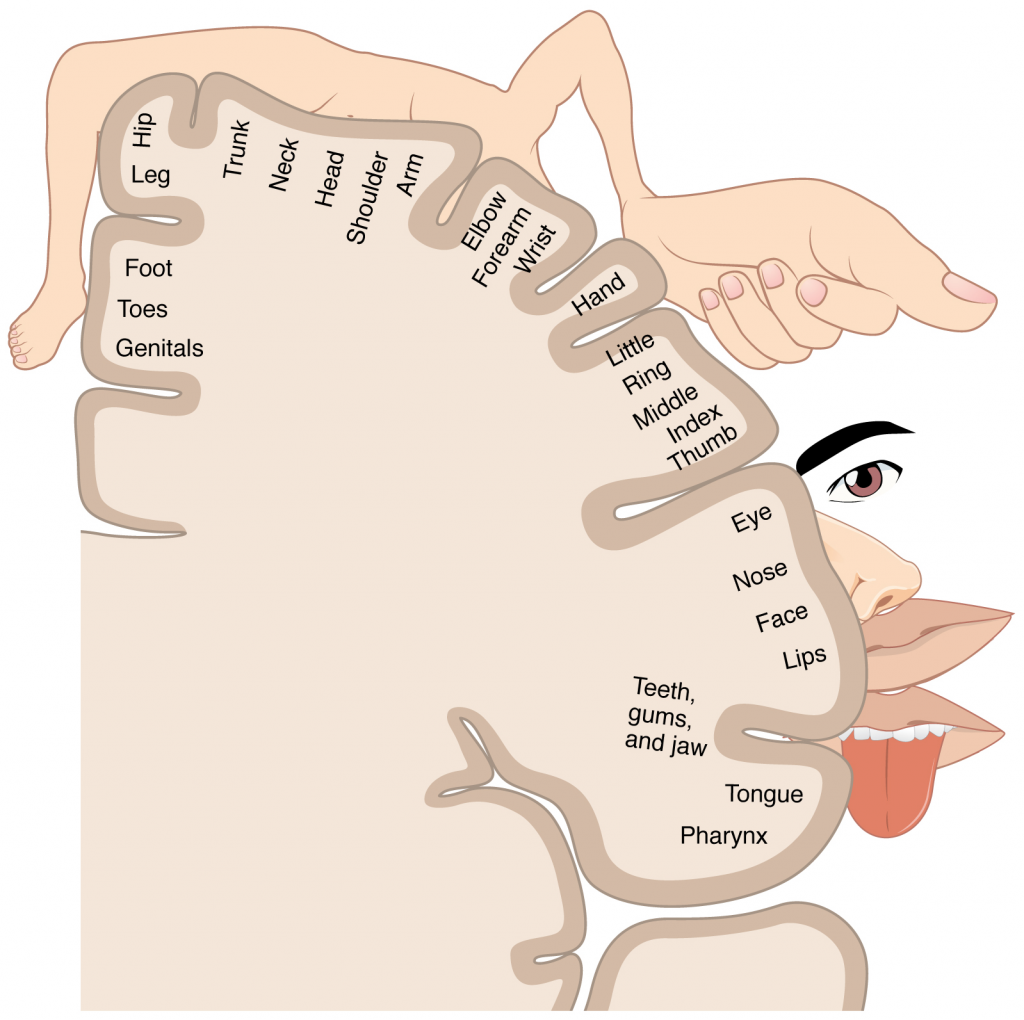
Figure 1.4.1. Regions of the human body. The human body is shown in anatomical position in an (a) anterior view and a (b) posterior view. The regions of the body are labelled in boldface.
A body that is lying down is described as either prone or supine. Prone describes a face-down orientation and supine describes a face up orientation. These terms are sometimes used in describing the position of the body during specific physical examinations or surgical procedures.
Regional Terms
The human body’s numerous regions have specific terms to help increase precision (see Figure 1.4.1). Notice that the term “brachium” or “arm” is reserved for the “upper arm” and “antebrachium” or “forearm” is used rather than “lower arm.” Similarly, “femur” or “thigh” is correct, and “leg” or “crus” is reserved for the portion of the lower limb between the knee and the ankle. You will be able to describe the body’s regions using the terms from the figure.
Directional Terms
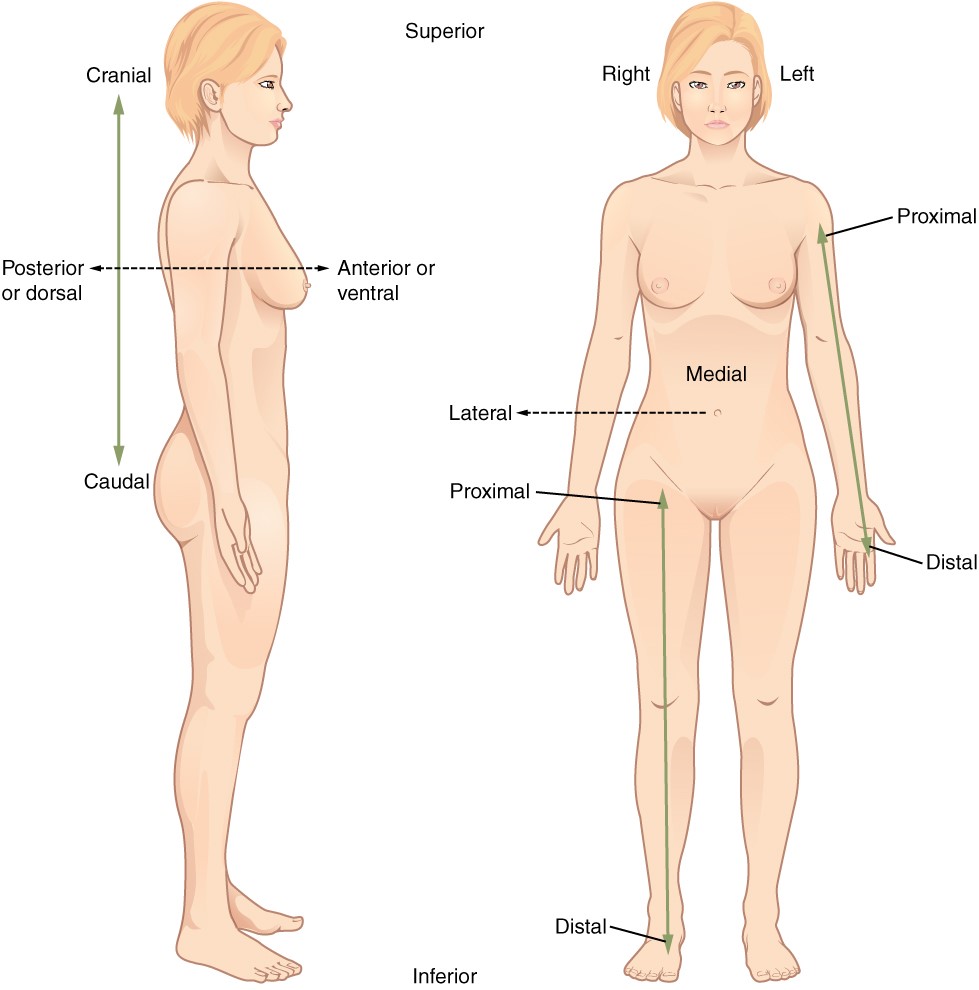
Certain directional anatomical terms appear throughout this and any other anatomy textbook (Figure 1.4.2). These terms are essential for describing the relative locations of different body structures. For instance, an anatomist might describe one band of tissue as “inferior to” another or a physician might describe a tumour as “superficial to” a deeper body structure. Commit these terms to memory to avoid confusion when you are studying or describing the locations of particular body parts.
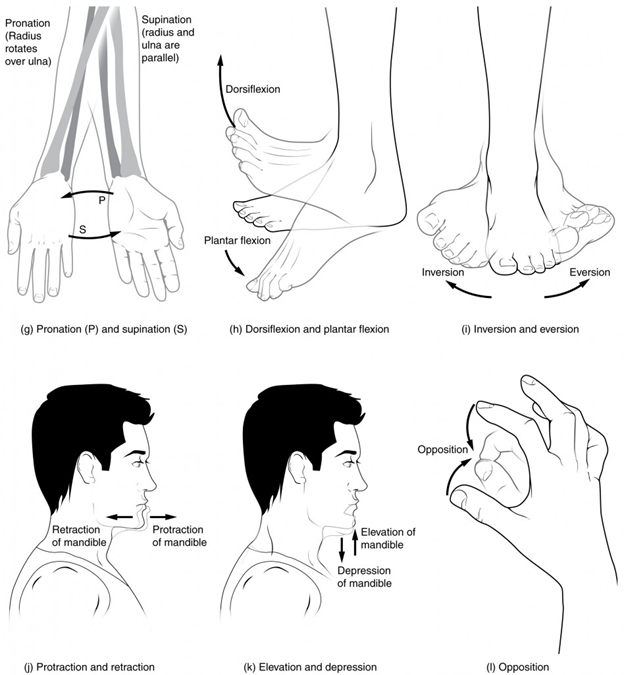
Anterior (or ventral) Describes the front or direction toward the front of the body. The toes are anterior to the foot.
Posterior (or dorsal) Describes the back or direction toward the back of the body. The popliteus is posterior to the patella.
Superior (or cranial) describes a position above or higher than another part of the body proper. The orbits are superior to the oris.
Inferior (or caudal) describes a position below or lower than another part of the body proper; near or toward the tail (in humans, the coccyx, or lowest part of the spinal column). The pelvis is inferior to the abdomen.
Lateral describes the side or direction toward the side of the body. The thumb (pollex) is lateral to the digits.
Medial describes the middle or direction toward the middle of the body. The hallux is the medial toe.
Proximal describes a position in a limb that is nearer to the point of attachment or the trunk of the body. The brachium is proximal to the antebrachium.
Distal describes a position in a limb that is farther from the point of attachment or the trunk of the body. The crus is distal to the femur.
Superficial describes a position closer to the surface of the body. The skin is superficial to the bones.
Deep describes a position farther from the surface of the body. The brain is deep to the skull.
Figure 1.4.2. Directional terms applied to the human body. Paired directional terms are shown as applied to the human body.
Body Planes
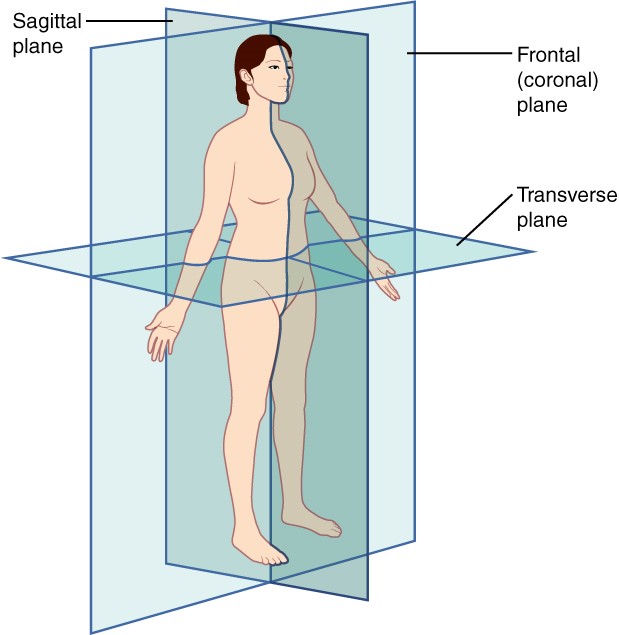
A section is a two-dimensional surface of a three-dimensional structure that has been cut. Modern medical imaging devices enable clinicians to obtain “virtual sections” of living bodies. We call these scans. Body sections and scans can be correctly interpreted, however, only if the viewer understands the plane along which the section was made. A plane is an imaginary two-dimensional surface that passes through the body. There are three planes commonly referred to in anatomy and medicine, as illustrated in Figure 1.4.3.
The sagittal plane is the plane that divides the body or an organ vertically into right and left sides. If this vertical plane runs directly down the middle of the body, it is called the midsagittal or median plane. If it divides the body into unequal right and left sides, it is called a parasagittal plane or less commonly a longitudinal section.
The frontal plane is the plane that divides the body or an organ into an anterior (front) portion and a posterior (rear) portion. The frontal plane is often referred to as a coronal plane. (“Corona” is Latin for “crown.”)
The transverse plane is the plane that divides the body or organ horizontally into upper and lower portions. Transverse planes produce images referred to as cross sections.
Figure 1.4.3. Planes of the body. The three planes most commonly used in anatomical and medical imaging are the sagittal, frontal (or coronal), and transverse plane.
Body Cavities and Serous Membranes
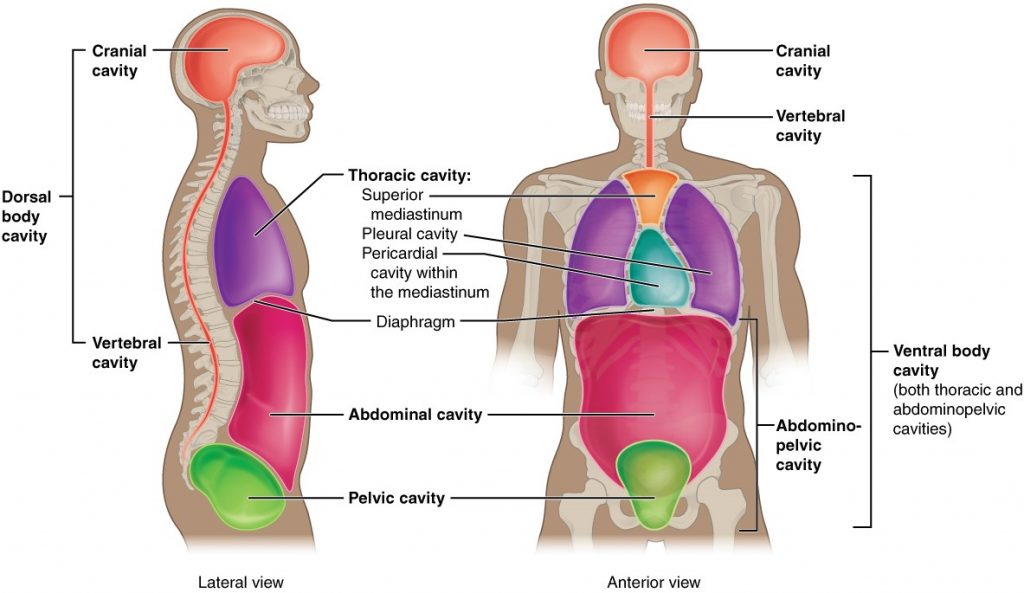
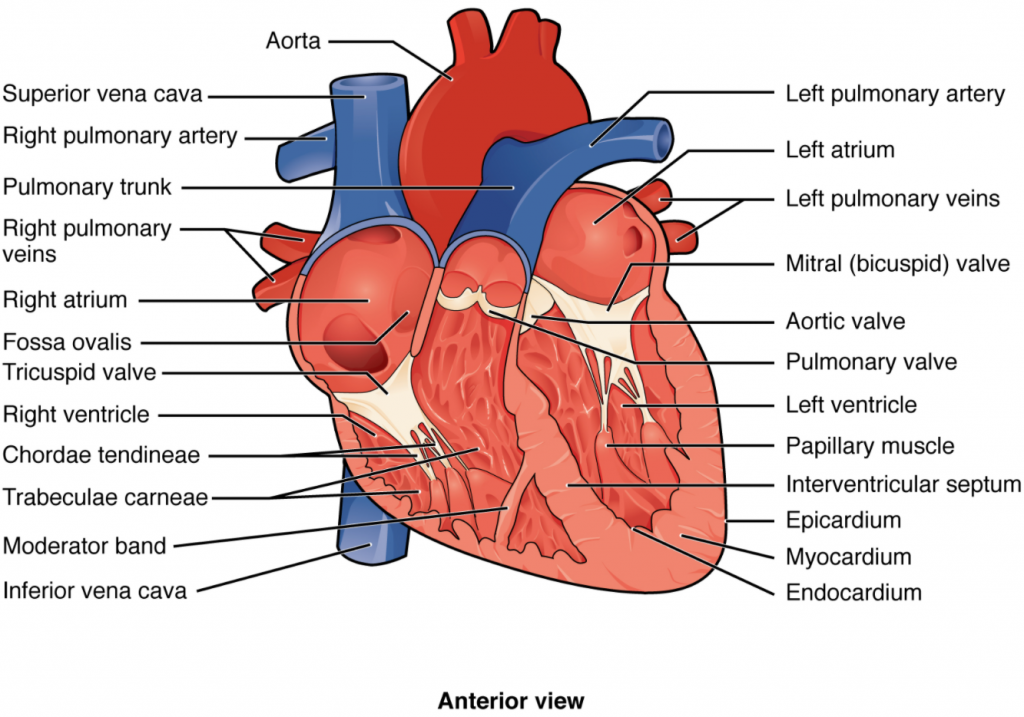
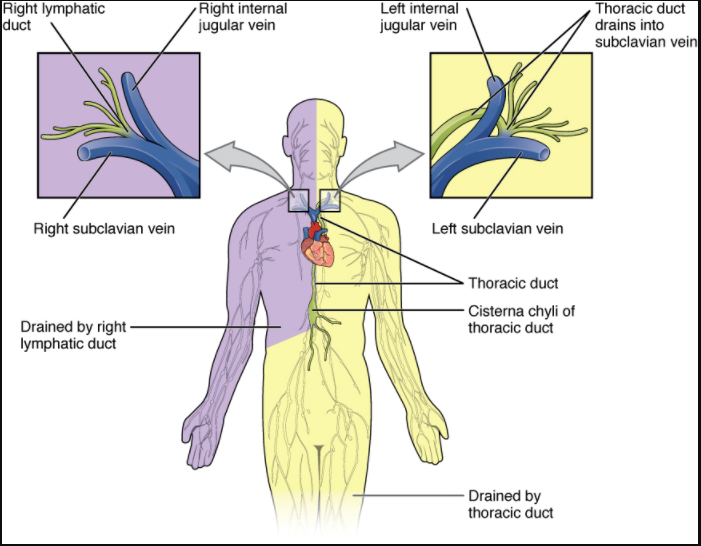
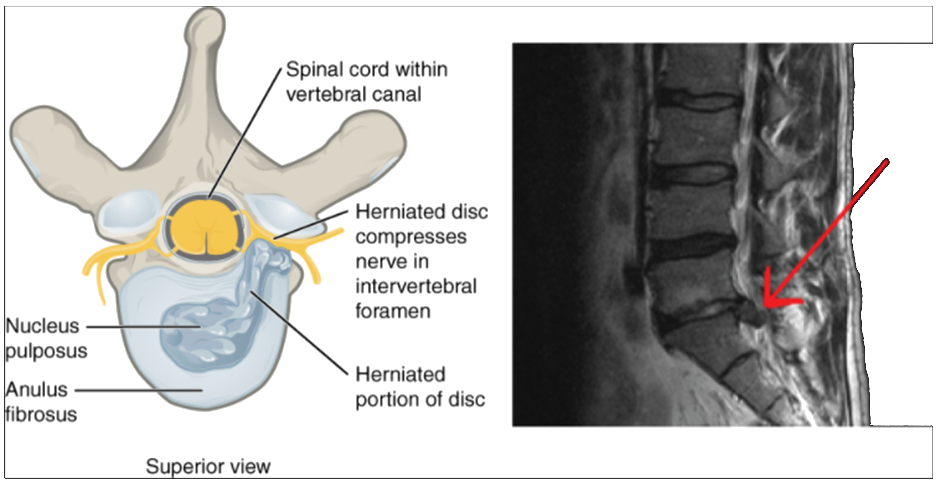
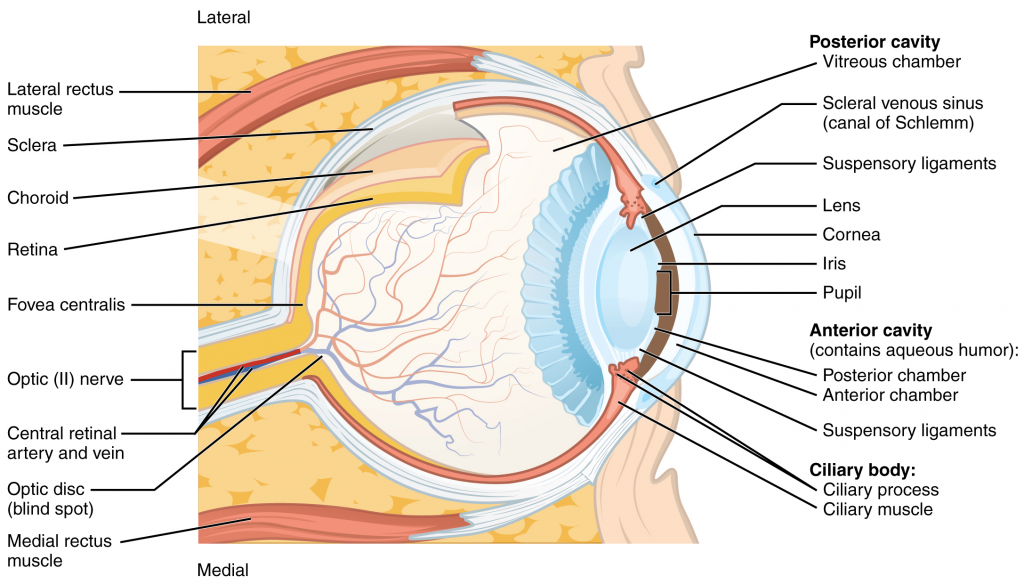
The body maintains its internal organisation by means of membranes, sheaths, and other structures that separate compartments. The dorsal (posterior) cavity and the ventral (anterior) cavity are the largest body compartments (Figure 1.4.4). These cavities contain and protect delicate internal organs, and the ventral cavity allows for significant changes in the size and shape of the organs as they perform their functions. The lungs, heart, stomach, and intestines, for example, can expand and contract without distorting other tissues or disrupting the activity of nearby organs.
Figure 1.4.4. Dorsal and ventral body cavities. The ventral cavity includes the thoracic and abdominopelvic cavities and their subdivisions. The dorsal cavity includes the cranial and spinal cavities.
Subdivisions of the Posterior (Dorsal) and Anterior (Ventral) Cavities
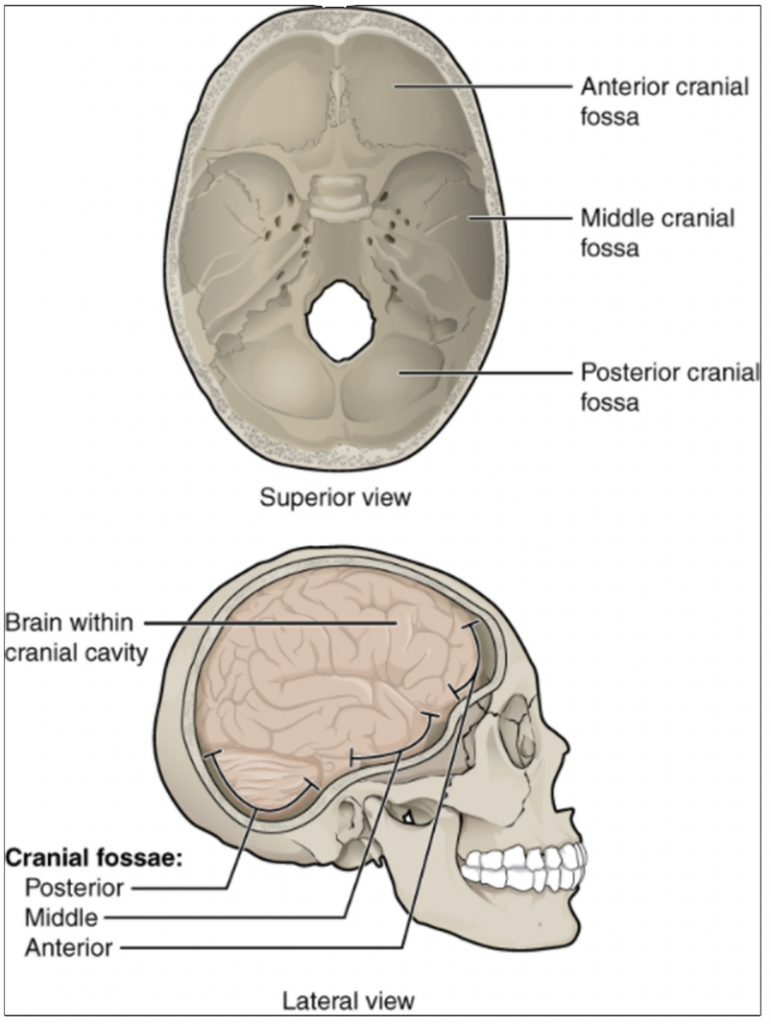
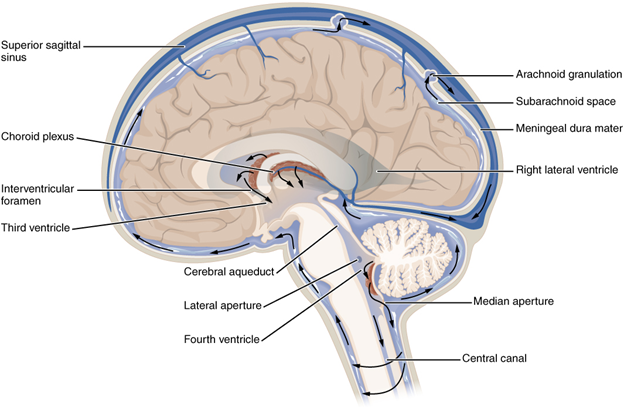
The posterior (dorsal) and anterior (ventral) cavities are each subdivided into smaller cavities. In the posterior (dorsal) cavity, the cranial cavity houses the brain, and the spinal cavity (or vertebral cavity) encloses the spinal cord. Just as the brain and spinal cord make up a continuous, uninterrupted structure, the cranial and spinal cavities that house them are also continuous. The brain and spinal cord are protected by the bones of the skull and vertebral column and by cerebrospinal fluid, a colourless fluid produced by the brain, which cushions the brain and spinal cord within the posterior (dorsal) cavity.
The anterior (ventral) cavity has two main subdivisions: the thoracic cavity and the abdominopelvic cavity (see Figure 1.4.4). The thoracic cavity is the more superior subdivision of the anterior cavity, and it is enclosed by the rib cage. The thoracic cavity contains the lungs and the heart, which is located in the mediastinum. The diaphragm forms the floor of the thoracic cavity and separates it from the more inferior abdominopelvic cavity. The abdominopelvic cavity is the largest cavity in the body. Although no membrane physically divides the abdominopelvic cavity, it can be useful to distinguish between the abdominal cavity, the division that houses the digestive organs, and the pelvic cavity, the division that houses the organs of reproduction.
Abdominal Regions and Quadrants
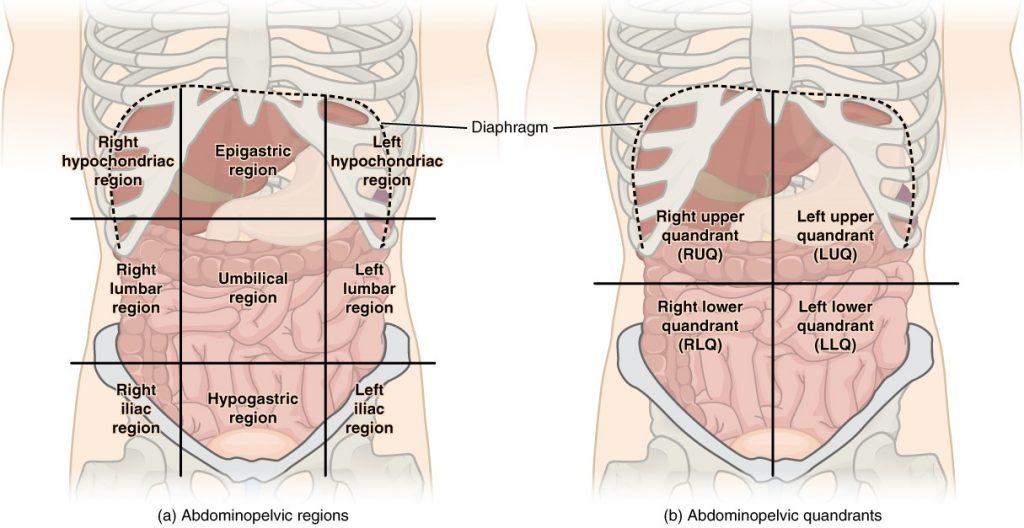
To promote clear communication, for instance about the location of a patient’s abdominal pain or a suspicious mass, health care providers typically divide up the cavity into either nine regions or four quadrants (Figure 1.4.5).
Figure 1.4.5. Regions and quadrants of the peritoneal cavity. There are (a) nine abdominal regions and (b) four abdominal quadrants in the peritoneal cavity.
The more detailed regional approach subdivides the cavity with one horizontal line immediately inferior to the ribs and one immediately superior to the pelvis, and two vertical lines drawn as if dropped from the midpoint of each clavicle (collarbone). There are nine resulting regions. The simpler quadrants approach, which is more commonly used in medicine, subdivides the cavity with one horizontal and one vertical line that intersect at the patient’s umbilicus (navel).
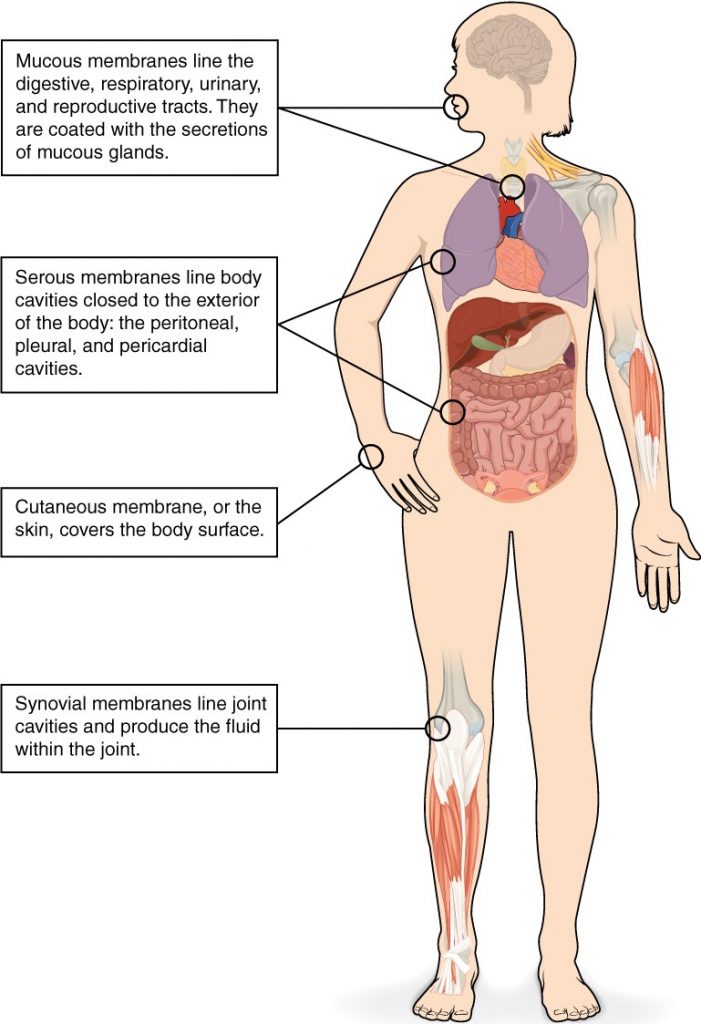
Membranes of the Anterior (Ventral) Body Cavity
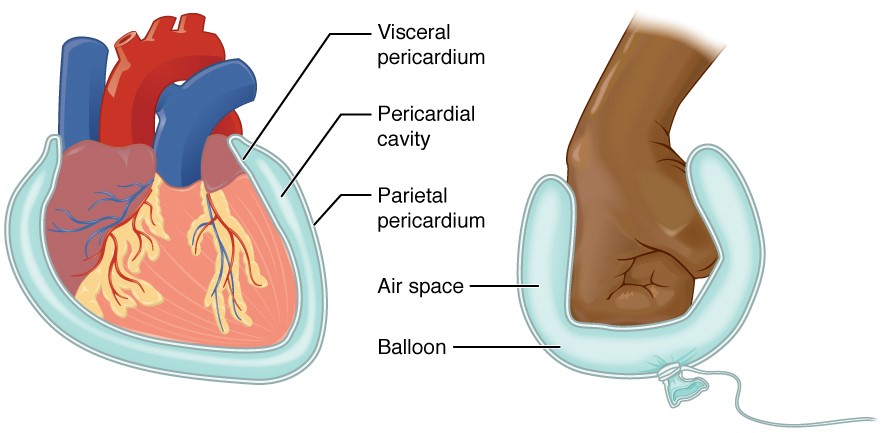
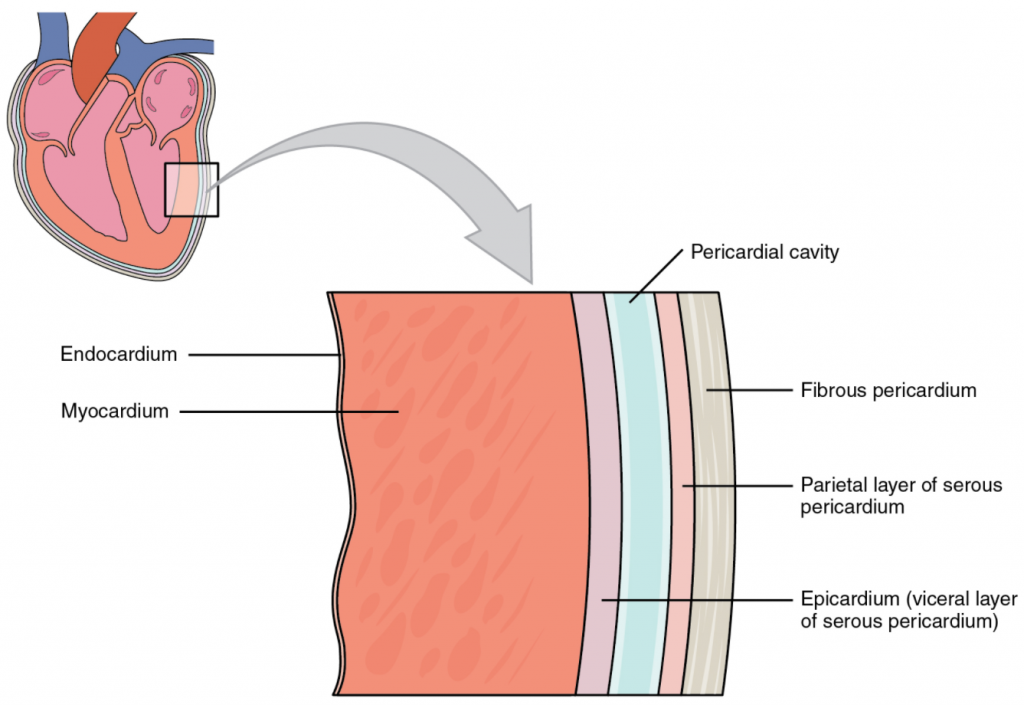
A serous membrane (also referred to a serosa) is one of the thin membranes that cover the walls and organs in the thoracic and abdominopelvic cavities. The parietal layers of the membranes line the walls of the body cavity (pariet- refers to a cavity wall). The visceral layer of the membrane covers the organs (the viscera). Between the parietal and visceral layers is a very thin, fluid-filled serous space, or cavity (Figure 1.4.6).
Figure 1.4.6. Serous membrane. Serous membrane lines the pericardial cavity and reflects back to cover the heart—much the same way that an underinflated balloon would form two layers surrounding a fist.
There are three serous cavities and their associated membranes. The pleura is the serous membrane that surrounds the lungs in the pleural cavity; the pericardium is the serous membrane that surrounds the heart in the pericardial cavity; and the peritoneum is the serous membrane that surrounds several organs in the abdominopelvic cavity. The serous membranes form fluid-filled sacs, or cavities, that are meant to cushion and reduce friction on internal organs when they move, such as when the lungs inflate or the heart beats. Both the parietal and visceral serosa secrete the thin, slippery serous fluid located within the serous cavities. The pleural cavity reduces friction between the lungs and the body wall. Likewise, the pericardial cavity reduces friction between the heart and the wall of the pericardium. The peritoneal cavity reduces friction between the abdominal and pelvic organs and the body wall. Therefore, serous membranes provide additional protection to the viscera they enclose by reducing friction that could lead to inflammation of the organs.
Section Review
Ancient Greek and Latin words are used to build anatomical terms. A standard reference position for mapping the body’s structures is the normal anatomical position. Regions of the body are identified using terms such as “occipital” that are more precise than common words and phrases such as “the back of the head.” Directional terms such as anterior and posterior are essential for accurately describing the relative locations of body structures. Images of the body’s interior commonly align along one of three planes: the sagittal, frontal, or transverse. The body’s organs are organised in one of two main cavities—dorsal (also referred to posterior) and ventral (also referred to anterior)—which are further sub-divided according to the structures present in each area. The serous membranes have two layers—parietal and visceral—surrounding a fluid filled space. Serous membranes cover the lungs (pleural serosa), heart (pericardial serosa), and some abdominopelvic organs (peritoneal serosa).
Explain dehydration (or condensation) and hydrolysis reactions
As you have learned, biological macromolecules are large molecules, necessary for life, that are built from smaller organic molecules. There are four major biological macromolecule classes (carbohydrates, lipids, proteins, and nucleic acids). Each is an important cell component and performs a wide array of functions. Combined, these molecules make up most of the cell’s dry mass (recall that water makes up most of its complete mass). Biological macromolecules are organic, meaning they contain carbon. In addition, they may contain hydrogen, oxygen, nitrogen, and additional minor elements.
Dehydration Synthesis
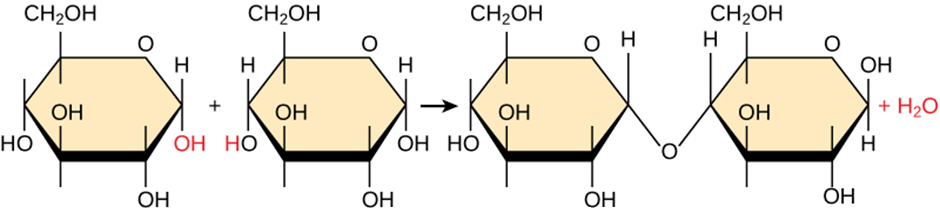
Most macromolecules are made from single subunits, or building blocks, called monomers. The monomers combine with each other using covalent bonds to form larger molecules known as polymers. In doing so, monomers release water molecules as by-products. This type of reaction is dehydration synthesis, which means “to put together while losing water.”
Figure 2.1.1.Dehydration synthesis. In the dehydration synthesis reaction above, two glucose molecules link to form the disaccharide maltose. In the process, it forms a water molecule.
In a dehydration synthesis reaction (Figure 2.1.1), the hydrogen of one monomer combines with the hydroxyl group of another monomer, releasing a water molecule. At the same time, the monomers share electrons and form covalent bonds. As additional monomers join, this chain of repeating monomers forms a polymer. Different monomer types can combine in many configurations, giving rise to a diverse group of macromolecules. Even one kind of monomer can combine in a variety of ways to form several different polymers. For example, glucose monomers are the constituents of starch, glycogen, and cellulose.
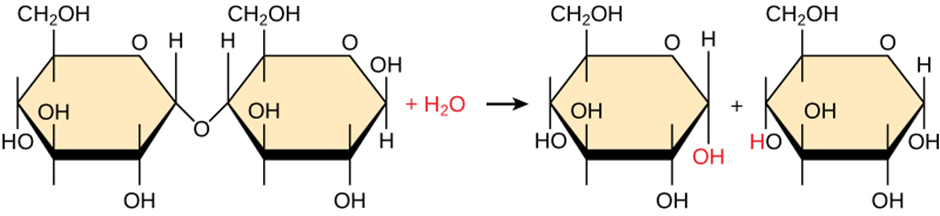
Hydrolysis
Polymers break down into monomers during hydrolysis. A chemical reaction occurs when inserting a water molecule across the bond. Breaking a covalent bond with this water molecule in the compound achieves this (Figure 2.1.2). During these reactions, the polymer breaks into two components: one part gains a hydrogen atom (H+) and the other gains a hydroxyl molecule (OH–) from a split water molecule.
Figure 2.1.2. Hydrolysis reaction. In the hydrolysis reaction here, the disaccharide maltose breaks down to form two glucose monomers by adding a water molecule. Note that this reaction is the reverse of the synthesis reaction in Figure 2.1.1.
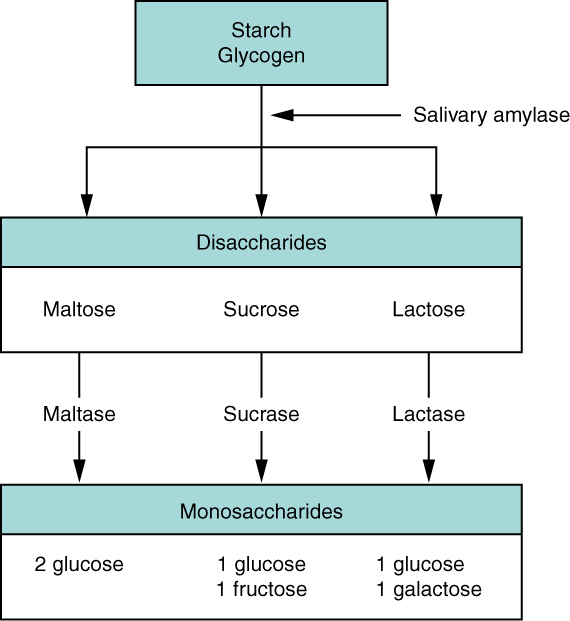
Dehydration and hydrolysis reactions are catalysed, or “sped up,” by specific enzymes; dehydration reactions involve the formation of new bonds, requiring energy, while hydrolysis reactions break bonds and release energy. These reactions are similar for most macromolecules, but each monomer and polymer reaction is specific for its class, for example, catalytic enzymes in the digestive system hydrolyse or break down the food we ingest into smaller molecules. This allows cells in our body to easily absorb nutrients in the intestine. A specific enzyme breaks down each macromolecule. For instance, amylase, sucrase, lactase, or maltase break down carbohydrates. Enzymes called proteases, such as pepsin and peptidase, and hydrochloric acid break down proteins. Lipases break down lipids. These broken-down macromolecules provide energy for cellular activities.
Section Review
Carbohydrates, lipids, proteins and nucleic acids are the four major classes of biological macromolecules—large molecules necessary for life that are built from smaller organic molecules. Macromolecules are comprised of single units scientists call monomers that are joined by covalent bonds to form larger polymers. The polymer is more than the sum of its parts: it acquires new characteristics and leads to an osmotic pressure that is much lower than that formed by its ingredients. This is an important advantage in maintaining cellular osmotic conditions. A monomer joins with another monomer with water molecule release, leading to a covalent bond forming. Scientists call these dehydration or condensation reactions. When polymers break down into smaller units (monomers), they use a water molecule for each bond broken by these reactions. Such reactions are hydrolysis reactions. Dehydration and hydrolysis reactions are similar for all macromolecules, but each monomer and polymer reaction is specific to its class. Dehydration reactions typically require an investment of energy for new bond formation, while hydrolysis reactions typically release energy by breaking bonds.
Note: Content and images from this chapter have been adapted from Biology 2ndedition by Mary Ann Clark, Jung Choi and Matthew Douglas, used under a CC-BY license.
2.2 Carbohydrates
Learning Objectives
By the end of this section, you will be able to:
Discuss the role of carbohydrates in cells and in the extracellular materials of animals and plants
Explain carbohydrate classifications
List common monosaccharides, disaccharides and polysaccharides
Most people are familiar with carbohydrates, one type of macromolecule, especially when it comes to what we eat. To lose weight, some individuals adhere to “low-carb” diets. Athletes, in contrast, often “carb-load” before important competitions to ensure that they have enough energy to compete at a high level. Carbohydrates are, in fact, an essential part of our diet. Grains, fruits, and vegetables are all-natural carbohydrate sources that provide energy to the body, particularly through glucose, a simple sugar that is a component of starch and an ingredient in many staple foods. Carbohydrates also have other important functions in humans, animals, and plants.
Molecular Structures
The stoichiometric formula (CH2O)n, where n is the number of carbons in the molecule represents carbohydrates. In other words, the ratio of carbon to hydrogen to oxygen is 1:2:1 in carbohydrate molecules. This formula also explains the origin of the term “carbohydrate”: the components are carbon (“carbo”) and the components of water (hence, “hydrate”). Scientists classify carbohydrates into three subtypes: monosaccharides, disaccharides and polysaccharides.
Monosaccharides
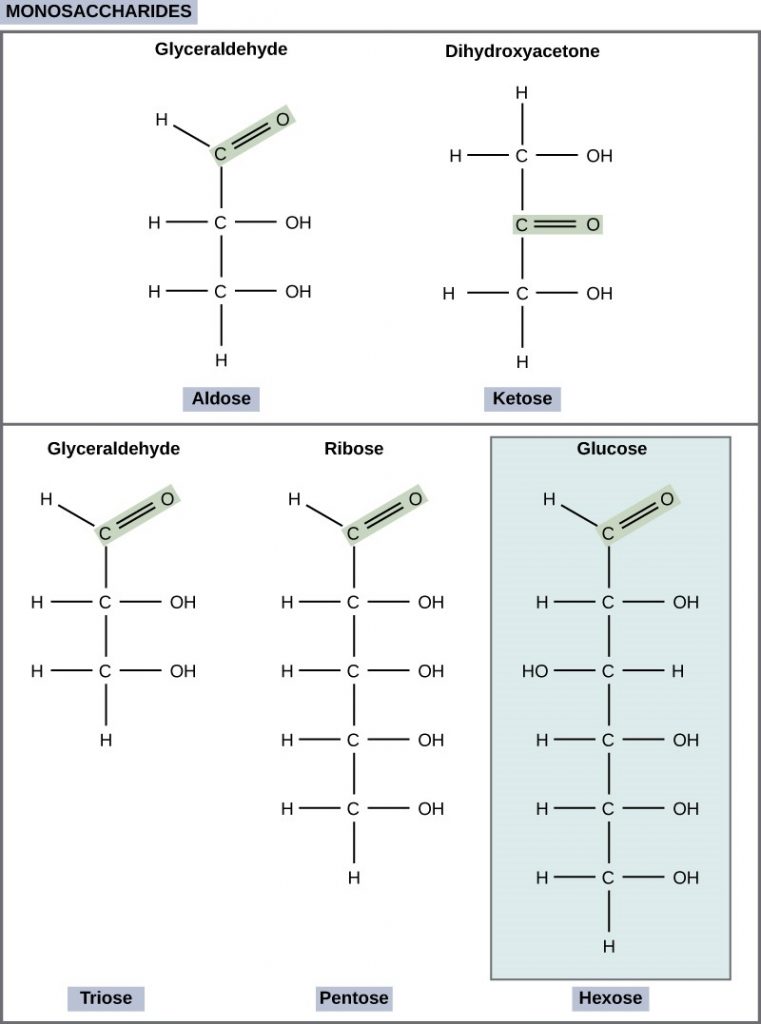
Monosaccharides (mono- = “one”; sacchar- = “sweet”) are simple sugars, the most common of which is glucose. In monosaccharides, the number of carbons usually ranges from three to seven. Most monosaccharide names end with the suffix-ose. If the sugar has an aldehyde group (the functional group with the structure R-CHO), it is an aldose, and if it has a ketone group (the functional group with the structure RC(=O)R’), it is a ketose. Depending on the number of carbons in the sugar, they can be trioses (three carbons), pentoses (five carbons), and/or hexoses (six carbons). Figure 2.2.1 illustrates monosaccharides.
Figure 2.2.1.Classifications of monosaccharides. Scientists classify monosaccharides based on the position of their carbonyl group and the number of carbons in the backbone. Aldoses have a carbonyl group (indicated in green) at the end of the carbon chain, and ketoses have a carbonyl group in the middle of the carbon chain. Trioses, pentoses, and hexoses have three-, five-, and six- carbon backbones, respectively.
The chemical formula for glucose is C6H12O6. In humans, glucose is an important source of energy. During cellular respiration, energy releases from glucose, and that energy helps make adenosine triphosphate (ATP). Plants synthesise glucose using carbon dioxide and water, and glucose in turn provides energy requirements for the plant. Humans and other animals that feed on plants often store excess glucose as catabolised (cell breakdown of larger molecules) starch.
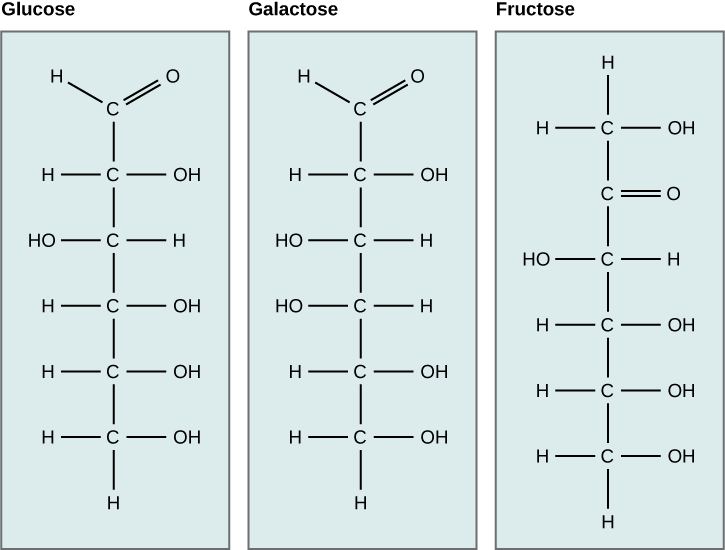
Galactose (part of lactose, or milk sugar) and fructose (found in sucrose, in fruit) are other common monosaccharides. Although glucose, galactose, and fructose all have the same chemical formula (C6H12O6), they differ structurally and chemically (and are isomers) because of the different arrangement of functional groups around the asymmetric carbon. All these monosaccharides have more than one asymmetric carbon (Figure 2.2.2).
Figure 2.2.2. Glucose isomers. Glucose, galactose, and fructose are all hexoses. They are structural isomers, meaning they have the same chemical formula (C6H12O6) but a different atom arrangement.
What kind of sugars are these, aldose or ketose?
Glucose, galactose, and fructose are isomeric monosaccharides (hexoses), meaning they have the same chemical formula but have slightly different structures. Glucose and galactose are aldoses, and fructose is a ketose.
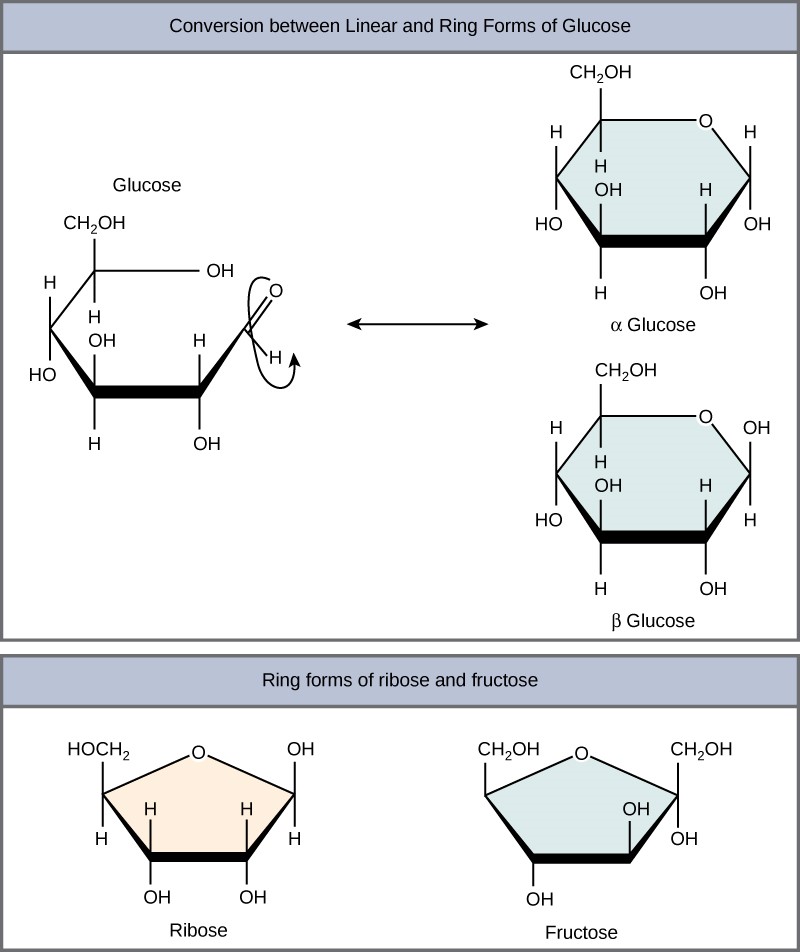
Monosaccharides can exist as a linear chain or as ring-shaped molecules. In aqueous solutions they are usually in ring forms (Figure 2.2.3). Glucose in a ring form can have two different hydroxyl group arrangements (OH) around the anomeric carbon (carbon 1 that becomes asymmetric in the ring formation process). If the hydroxyl group is below carbon number 1 in the sugar, it is in the alpha (α) position, and if it is above the plane, it is in the beta (β) position.
Figure 2.2.3.Ring forms of glucose. Five and six carbon monosaccharides exist in equilibrium between linear and ring forms. When the ring forms, the side chain it closes on locks into an α or β position. Fructose and ribose also form rings, although they form five-membered rings as opposed to the six-membered ring of glucose.
Disaccharides
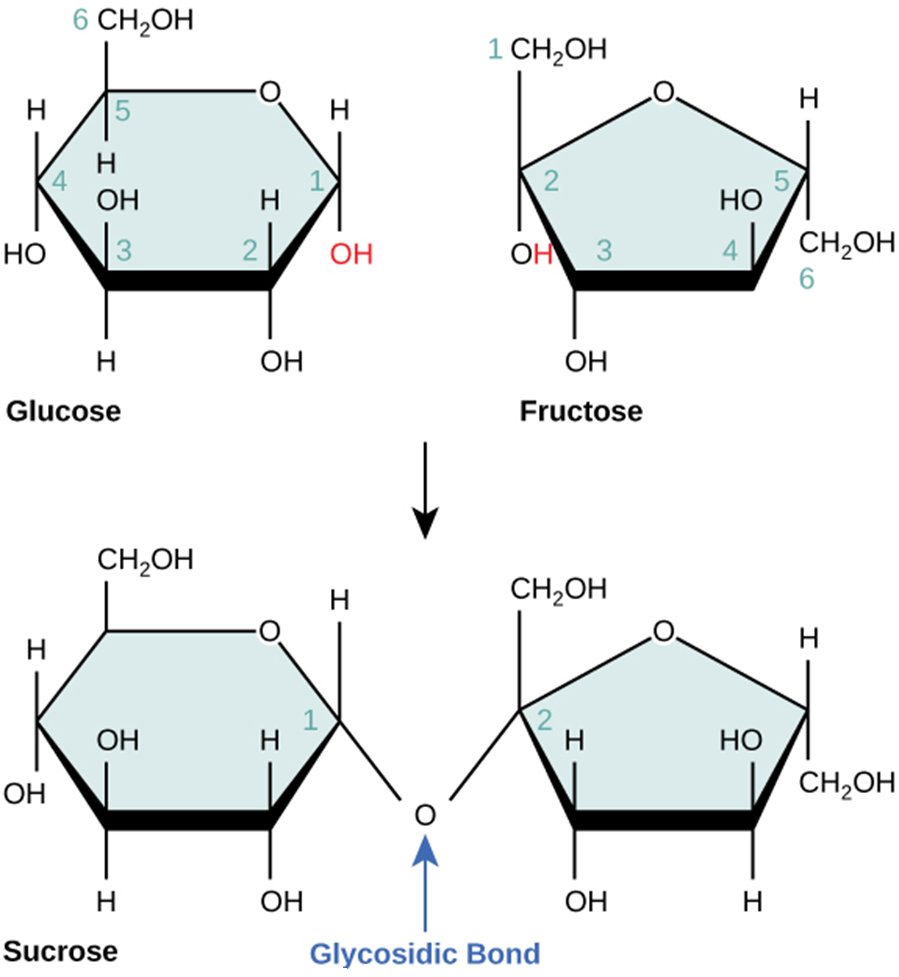
Disaccharides (di- = “two”) form when two monosaccharides undergo a dehydration reaction (or a condensation reaction or dehydration synthesis). During this process, one monosaccharide’s hydroxyl group combines with another monosaccharide’s hydrogen, releasing a water molecule and forming a covalent bond. A covalent bond forms between a carbohydrate molecule and another molecule (in this case, between two monosaccharides). Scientists call this a glycosidic bond (Figure 2.2.4). Glycosidic bonds (or glycosidic linkages) can be an alpha or beta type. An alpha bond is formed when the OH group on the carbon-1 of the first glucose is below the ring plane, and a beta bond is formed when the OH group on the carbon-1 is above the ring plane.
Figure 2.2.4.Sucrose. Sucrose forms when a glucose monomer and a fructose monomer join in a dehydration reaction to form a glycosidic bond. In the process, a water molecule is lost. By convention, the carbon atoms in a monosaccharide are numbered from the terminal carbon closest to the carbonyl group. In sucrose, a glycosidic linkage forms between carbon 1 in glucose and carbon 2 in fructose.
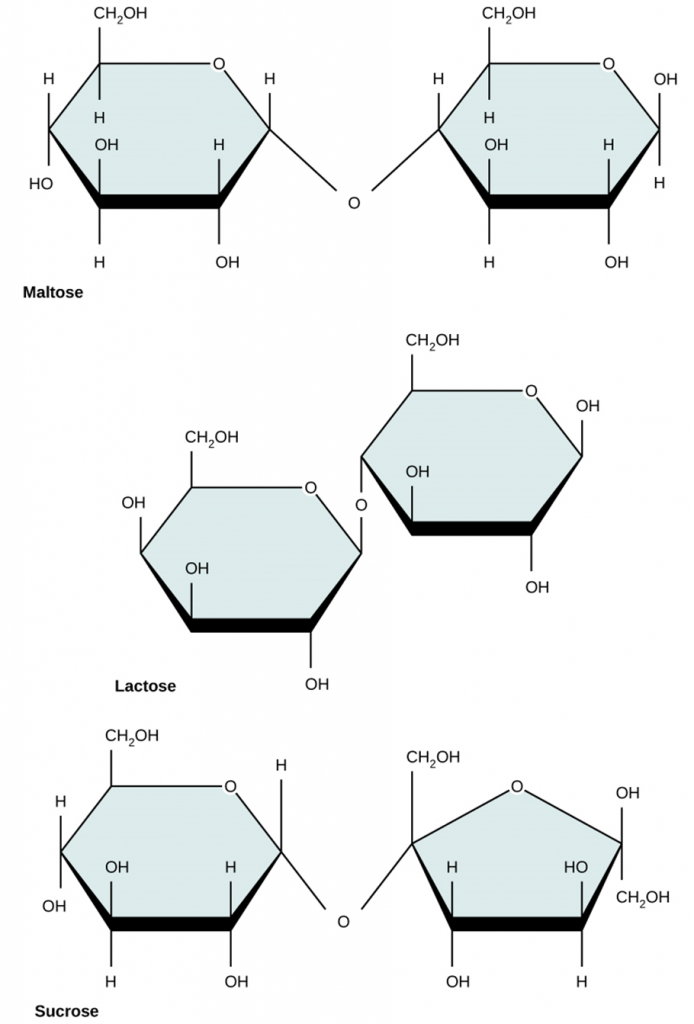
Common disaccharides include lactose, maltose, and sucrose (Figure 2.2.5). Lactose is a disaccharide consisting of the monomers; glucose and galactose. It is naturally in milk. Maltose, or malt sugar, is a disaccharide formed by a dehydration reaction between two glucose molecules. The most common disaccharide is sucrose, or table sugar, which is comprised of glucose and fructose monomers.
Figure 2.2.5.Common disaccharides. Common disaccharides include maltose (grain sugar), lactose (milk sugar) and sucrose (table sugar).
Polysaccharides
A long chain of monosaccharides linked by glycosidic bonds is a polysaccharide (poly- = “many”). The chain may be branched or unbranched, and it may contain different types of monosaccharides. The molecular weight (MW) may be 100,000 Daltons or more depending on the number of joined monomers. Starch, glycogen, cellulose and chitin are primary examples of polysaccharides.
Plants store sugars in the form of starch. In plants, an amylose and amylopectin mixture (both glucose polymers) comprise these sugars. Plants can synthesise glucose, and they store the excess glucose, beyond their immediate energy needs, as starch in different plant parts, including roots and seeds. The starch in the seeds provides food for the embryo as it germinates and can also act as a food source for humans and animals. Enzymes break down the starch that humans consume, for example, an amylase present in saliva catalyses, or breaks down this starch into smaller molecules, such as maltose and glucose. The cells can then absorb the glucose.
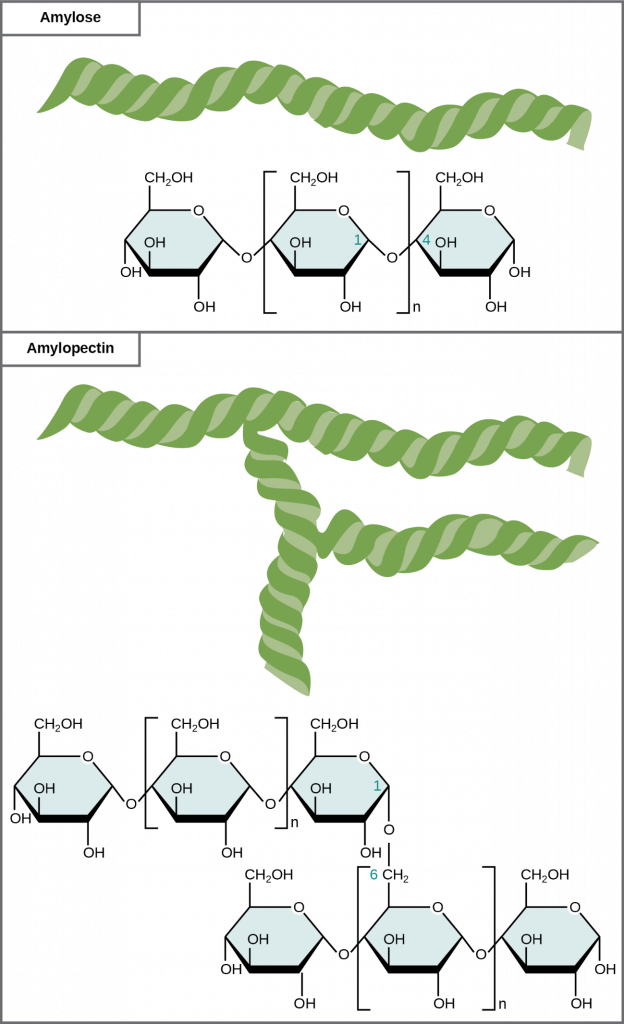
Glucose starch comprises monomers that are joined by α 1-4 or α 1-6 glycosidic bonds. The numbers 1-4 and 1-6 refer to the carbon number of the two residues that have joined to form the bond. As Figure 2.2.6 illustrates, unbranched glucose monomer chains (only α 1-4 linkages) form the starch; whereas, amylopectin is a branched polysaccharide (α 1-6 linkages at the branch points).
Figure 2.2.6. Starch forms. Amylose and amylopectin are two different starch forms. Unbranched glucose monomer chains comprise amylose by α 1-4 glycosidic linkages. Unbranched glucose monomer chains comprise amylopectin by α 1-4 and α 1-6 glycosidic linkages. Because of the way the subunits are joined, the glucose chains have a helical structure. Glycogen (not shown) is similar in structure to amylopectin but more highly branched.
Glycogen is the storage form of glucose in humans and other vertebrates and is comprised of monomers of glucose. Glycogen is the animal equivalent of starch and is a highly branched molecule usually stored in liver and muscle cells. Whenever blood glucose levels decrease, glycogen breaks down to release glucose in a process scientists call glycogenolysis.
Cellulose is the most abundant natural biopolymer. Cellulose mostly comprises a plant’s cell wall. This provides the cell structural support. Wood and paper are mostly cellulosic in nature. Glucose monomers comprise cellulose that β 1-4 glycosidic bonds link (Figure 2.2.7).
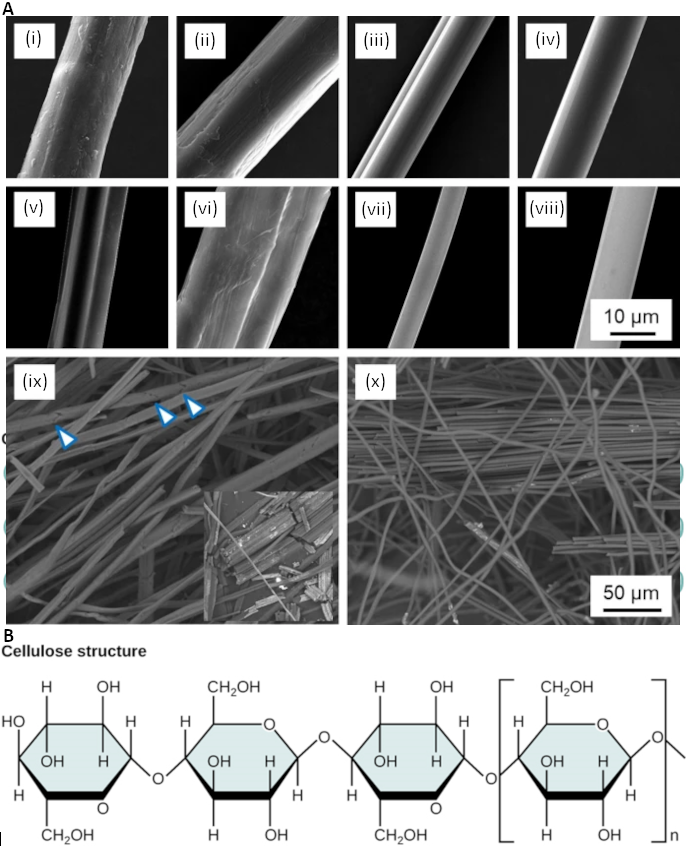
Figure 2.2.7. Figure 2.2.7. A) Scanning electron microscopy of single cellulose fibres (i) hemp, (ii) ramie, (iii) viscose, (iv) Tencel and their carbonised counterparts (v-viii) together with a low-magnification image of carbonised natural hemp fibres (ix) and Tencel regenerated cellulose fibres (x). Arrowheads in (ix) indicate cracks across the diameter of carbonised fibres leading to easy fragmentation as shown in the inset (note that a pair of fibres is shown in vi) (https://link.springer.com/article/10.1007/s10853-020-04743-y used under CC-BY licence); B) Cellulose structure in cellulose, glucose monomers are linked in unbranched chains by β 1-4 glycosidic linkages. Because of the way the glucose subunits are joined, every glucose monomer is flipped relative to the next one resulting in a linear, fibrous structure.
As Figure 2.2.7 shows, every other glucose monomer in cellulose is flipped over, and the monomers are packed tightly as extended long chains. This gives cellulose its rigidity and high tensile strength—which is so important to plant cells. While human digestive enzymes cannot break down the β 1-4 linkage, herbivores such as cows, koalas, and buffalos are able, with the help of the specialised microorganisms in their stomach, to digest plant material that is rich in cellulose and use it as a food source. In some of these animals, certain species of bacteria and protists reside in the rumen (part of the herbivore’s digestive system) and secrete the enzyme cellulase. The appendix of grazing animals also contains bacteria that digest cellulose, giving it an important role in ruminants’ digestive systems. Cellulases can break down cellulose into glucose monomers that animals use as an energy source. Termites are also able to break down cellulose because of the presence of other organisms in their bodies that secrete cellulases.
Figure 2.2.8.Bee exoskeleton. Insects have a hard-outer exoskeleton made of chitin, a type of polysaccharide. (credit: Louise Docker).
Carbohydrates serve various functions in different animals. Arthropods (insects, crustaceans, and others) have an outer skeleton, the exoskeleton, which protects their internal body parts (as we see in the bee in Figure 2.2.8). This exoskeleton is made of the biological macromolecule chitin, which is a nitrogen-containing polysaccharide. It is made of repeating N-acetyl-β-d-glucosamine units, which are a modified sugar. Chitin is also a major component of fungal cell walls. Fungi are neither animals nor plants and form a kingdom of their own in the domain Eukarya.
Benefits of Carbohydrates
Are carbohydrates good for you? Some people believe that carbohydrates are bad and they should avoid them. Some diets completely forbid carbohydrate consumption, claiming that a low-carbohydrate diet helps people to lose weight faster. However, carbohydrates have been an important part of the human diet for thousands of years. Artifacts from ancient civilizations show the presence of wheat, rice, and corn in our ancestors’ storage areas.
As part of a well-balanced diet, we should supplement carbohydrates with proteins, vitamins, and fats. Calorie-wise, a gram of carbohydrate provides 4.3 Kcal. For comparison, fats provide 9 Kcal/g, a less desirable ratio. Carbohydrates contain soluble and insoluble elements. The insoluble part, fibre, is mostly cellulose. Fibre has many uses. It promotes regular bowel movement by adding bulk, and it regulates the blood glucose consumption rate. Fibre also helps to remove excess cholesterol from the body. Fibre binds to the cholesterol in the small intestine, then attaches to the cholesterol and prevents the cholesterol particles from entering the bloodstream. Cholesterol then exits the body via the faeces. Fibre-rich diets also have a protective role in reducing the occurrence of colon cancer. In addition, a meal containing whole grains and vegetables gives a feeling of fullness. As an immediate source of energy, glucose breaks down during the cellular respiration process, which produces ATP, the cell’s energy currency. Without consuming carbohydrates, we reduce the availability of “instant energy”. Eliminating carbohydrates from the diet may be necessary for some people, but such a step may not be healthy for everyone.
Career Connections: Registered Dietitian
Obesity is a worldwide health concern, and many diseases such as diabetes and heart disease are becoming more prevalent because of obesity. This is one of the reasons why people increasingly seek out registered dietitians for advice. Registered dietitians help plan nutrition programs for individuals in various settings. They often work with patients in health care facilities, designing nutrition plans to treat and prevent diseases. For example, dietitians may teach a patient with diabetes how to manage blood sugar levels by eating the correct types and amounts of carbohydrates. Dietitians may also work in nursing homes, schools, and private practices.
To become a registered dietitian, one needs to earn at least a bachelor’s degree in dietetics, nutrition, food technology, or a related field. In addition, registered dietitians must complete a supervised internship program and pass a national exam. Those who pursue careers in dietetics take courses in nutrition, chemistry, biochemistry, biology, microbiology, and human physiology. Dietitians must become experts in the chemistry and physiology (biological functions) of food (proteins, carbohydrates, and fats).
Biomedical Scientist
A biomedical scientist has at least a bachelor’s degree in biomedical science or science with a major in biomedical science. The majority of biomedical scientists also have an Honours degree (popular in countries such as Australia, Singapore, India and the United Kingdom) or a Masters degree (United States, Canada, Americas, Europe, parts of Asia and Africa), with PhDs also common. Biomedical scientists work in university, research institutes and commercial laboratories undertaking basic through to translational research projects aimed at improving our understanding of the human body in health and disease and investigating better ways to prevent, treat and diagnose diseases. Funding may be public in the form of government grants or commercial investment from the pharmaceutical and biotechnology industries.
Section Review
Carbohydrates are a group of macromolecules that are a vital energy source for the cell and provide structural support to plant cells, fungi, and all of the arthropods that include lobsters, crabs, shrimp, insects, and spiders. Scientists classify carbohydrates as monosaccharides, disaccharides, and polysaccharides depending on the number of monomers in the molecule. Monosaccharides are linked by glycosidic bonds that form as a result of dehydration reactions, forming disaccharides and polysaccharides with eliminating a water molecule for each bond formed. Glucose, galactose, and fructose are common monosaccharides; whereas, common disaccharides include lactose, maltose, and sucrose. Starch and glycogen, examples of polysaccharides, are the storage forms of glucose in plants and animals, respectively. The long polysaccharide chains may be branched or unbranched. Cellulose is an example of an unbranched polysaccharide; whereas, amylopectin, a constituent of starch, is a highly branched molecule. Glucose storage, in the form of polymers like starch of glycogen, makes it slightly less accessible for metabolism; however, this prevents it from leaking out of the cell or creating a high osmotic pressure that could cause the cell to uptake excessive water.
Note: Content and images from this chapter have been adapted from Biology 2nd edition by Mary Ann Clark, Jung Choi and Matthew Douglas, used under a CC-BY license.
2.3 Lipids
Learning Objectives
By the end of this section, you will be able to:
Describe the four major types of lipids
Explain the role of fats in storing energy
Differentiate between saturated and unsaturated fatty acids
Describe phospholipids and their role in cells
Define the basic structure of a steroid and some steroid functions
Explain how cholesterol helps maintain the plasma membrane’s fluid nature
Figure 2.3.1. Otter fur. Hydrophobic lipids in aquatic mammals’ fur, such as this river otter, protect them from the elements. (credit: Ken Bosma).
Lipids include a diverse group of compounds that are largely nonpolar in nature. This is because they are hydrocarbons that include mostly nonpolar carbon–carbon or carbon–hydrogen bonds. Non-polar molecules are hydrophobic (“water fearing”), or insoluble in water. Lipids perform many different functions in a cell. Cells store energy for long-term use in the form of fats. Lipids also provide insulation from the environment for plants and animals (Figure 2.3.1). For example, they help keep aquatic birds and mammals dry when forming a protective layer over fur or feathers because of their water-repellent hydrophobic nature. Lipids are also the building blocks of many hormones and are an important constituent of all cellular membranes. Lipids include fats and oils, waxes, phospholipids and steroids.
Fats and Oils
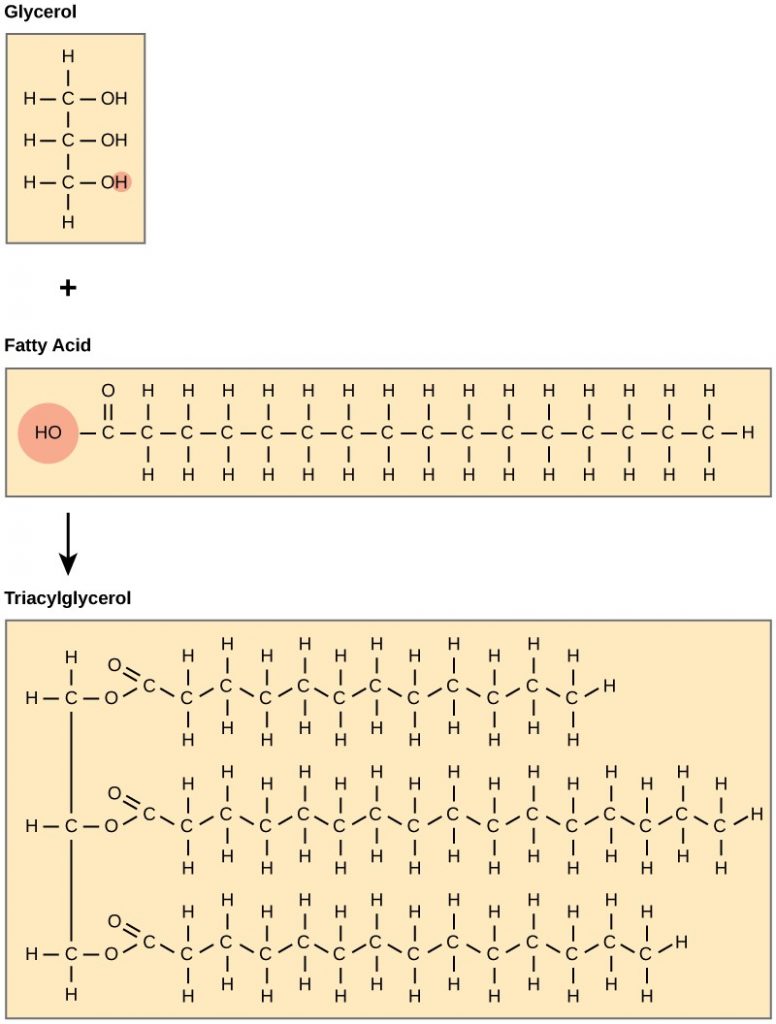
A fat molecule consists of two main components—glycerol and fatty acids. Glycerol is an organic compound (alcohol) with three carbons, five hydrogens, and three hydroxyl (OH) groups. Fatty acids have a long chain of hydrocarbons to which a carboxyl group is attached, hence the name “fatty acid.” The number of carbons in the fatty acid may range from 4 to 36. The most common are those containing 12–18 carbons. In a fat molecule, the fatty acids attach to each of the glycerol molecule’s three carbons with an ester bond through an oxygen atom (Figure 2.3.2).
Figure 2.3.2.Fatty acid dehydration reaction. Joining three fatty acids to a glycerol backbone in a dehydration reaction forms triacylglycerol. Three water molecules release in the process.
During this ester bond formation, three water molecules are released. The three fatty acids in the triacylglycerol may be similar or dissimilar. We also call fats triacylglycerols (TAGs) or triglycerides (TGs) because of their chemical structure. Some fatty acids have common names that specify their origin, for example, palmitic acid, a saturated fatty acid, is derived from the palm tree. Arachidic acid (also known as eicosanoic acid) is derived from Arachis hypogea, the scientific name for groundnuts or peanuts.
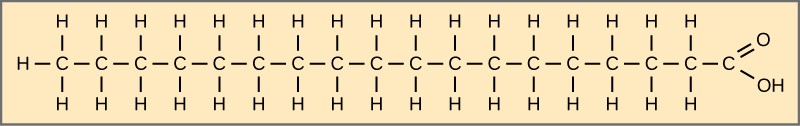
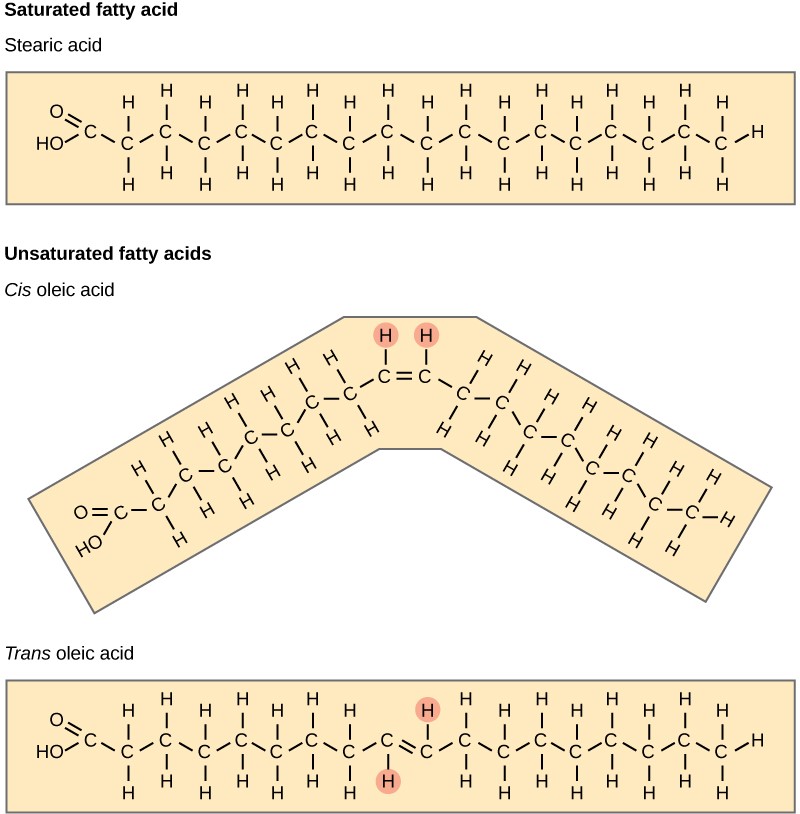
Fatty acids may be saturated or unsaturated. In a fatty acid chain, if there are only single bonds between neighbouring carbons in the hydrocarbon chain, the fatty acid is saturated. Saturated fatty acids are saturated with hydrogen. In other words, the number of hydrogen atoms attached to the carbon skeleton is maximised. Stearic acid is an example of a saturated fatty acid (Figure 2.3.3).
Figure 2.3.3.Stearic acid. Stearic acid is a common saturated fatty acid.
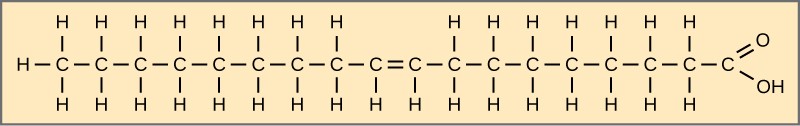
When the hydrocarbon chain contains a double bond, the fatty acid is unsaturated. Oleic acid is an example of an unsaturated fatty acid (Figure 2.3.4).
Figure 2.3.4.Oleic acid. Oleic acid is a common unsaturated fatty acid.
Most unsaturated fats are liquid at room temperature. We call these oils. If there is one double bond in the molecule, then it is a monounsaturated fat (e.g., olive oil), and if there is more than one double bond, then it is a polyunsaturated fat (e.g., canola oil).
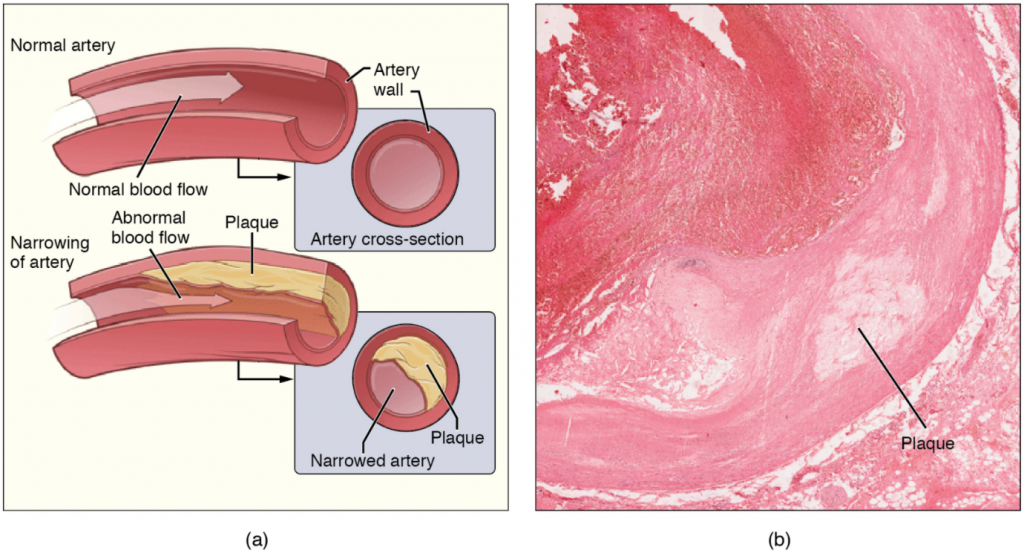
When a fatty acid has no double bonds, it is a saturated fatty acid because it is not possible to add more hydrogen to the chain’s carbon atoms. A fat may contain similar or different fatty acids attached to glycerol. Long straight fatty acids with single bonds generally pack tightly and are solid at room temperature. Animal fats with stearic acid and palmitic acid (common in meat) and the fat with butyric acid (common in butter) are examples of saturated fats. Mammals store fats in specialised cells, or adipocytes, where fat globules occupy most of the cell’s volume. Plants store fat or oil in many seeds and use them as a source of energy during seedling development. Unsaturated fats or oils are usually of plant origin and contain cis unsaturated fatty acids. Cis and trans indicate the configuration of the molecule around the double bond. If hydrogens are present in the same plane, it is a cis fat. If the hydrogen atoms are on two different planes, it is a trans-fat. The cis double bond causes a bend or a “kink” that prevents the fatty acids from packing tightly, keeping them liquid at room temperature (Figure 2.3.5). Olive oil, corn oil, canola oil, and cod liver oil are examples of unsaturated fats. Unsaturated fats help to lower blood cholesterol levels; whereas, saturated fats contribute to plaque formation in the arteries.
Figure 2.3.5.Saturated fatty acids. Saturated fatty acids have hydrocarbon chains connected by single bonds only. Unsaturated fatty acids have one or more double bonds. Each double bond may be in a cis or trans configuration. In the cis configuration, both hydrogens are on the same side of the hydrocarbon chain. In the trans configuration, the hydrogens are on opposite sides. A cis double bond causes a kink in the chain.
Trans Fats
The food industry artificially hydrogenates oils to make them semi-solid and of a consistency desirable for many processed food products. Simply speaking, hydrogen gas is bubbled through oils to solidify them. During this hydrogenation process, double bonds of the cis– conformation in the hydrocarbon chain may convert to double bonds in the trans– conformation.
Margarine, some types of peanut butter, and shortening are examples of artificially hydrogenated trans fats. Recent studies have shown that an increase in trans fats in the human diet may lead to higher levels of low-density lipoproteins (LDL), or “bad” cholesterol, which in turn may lead to plaque deposition in the arteries, resulting in heart disease. Many fast-food restaurants have recently banned using trans fats, and food labels are required to display the trans-fat content.
Omega Fatty Acids
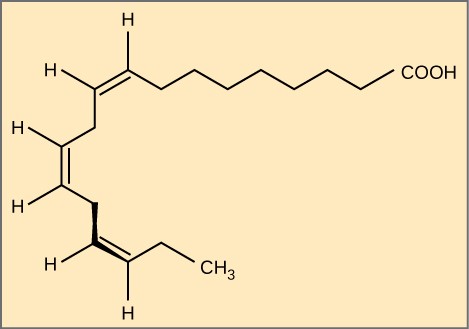
Essential fatty acids are those that the human body requires but does not synthesise. Consequently, they must be supplemented through ingestion via the diet. Omega-3 fatty acids (like those in Figure 2.3.6) fall into this category and are one of only two known for humans (the other is omega-6 fatty acid). These are polyunsaturated fatty acids and are omega-3 because a double bond connects the third carbon from the hydrocarbon chain’s end to its neighbouring carbon.
Figure 2.3.6. Fatty acid omega-3. Alpha-linolenic acid is an example of an omega-3 fatty acid. It has three cis double bonds and, as a result, a curved shape. For clarity, the diagram does not show the carbons. Each singly bonded carbon has two hydrogens associated with it, which the diagram also does not show.
The farthest carbon away from the carboxyl group is numbered as the omega (ω) carbon, and if the double bond is between the third and fourth carbon from that end, it is an omega-3 fatty acid. Nutritionally important because the body does not make them, omega-3 fatty acids include alpha-linoleic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA), all of which are polyunsaturated. Salmon, trout, and tuna are good sources of omega-3 fatty acids. Research indicates that omega-3 fatty acids reduce the risk of sudden death from heart attacks, lower triglycerides in the blood, decrease blood pressure, and prevent thrombosis by inhibiting blood clotting. They also reduce inflammation and may help lower the risk of some cancers in animals.
Like carbohydrates, fats have received considerable bad publicity. It is true that eating an excess of fried foods and other “fatty” foods leads to weight gain. However, fats do have important functions. Many vitamins are fat soluble, and fats serve as a long-term storage form of fatty acids: a source of energy. They also provide insulation for the body. Therefore, we should consume “healthy” fats in moderate amounts on a regular basis.
Waxes
Figure 2.3.7.Waxy leaves. Lipids comprise waxy coverings on some leaves. (Credit: Roger Griffith).
Wax covers some aquatic birds’ feathers and some plants’ leaf surfaces. Because of waxes’ hydrophobic nature, they prevent water from sticking on the surface (Figure 2.3.7). Long fatty acid chains esterified to long-chain alcohols comprise waxes.
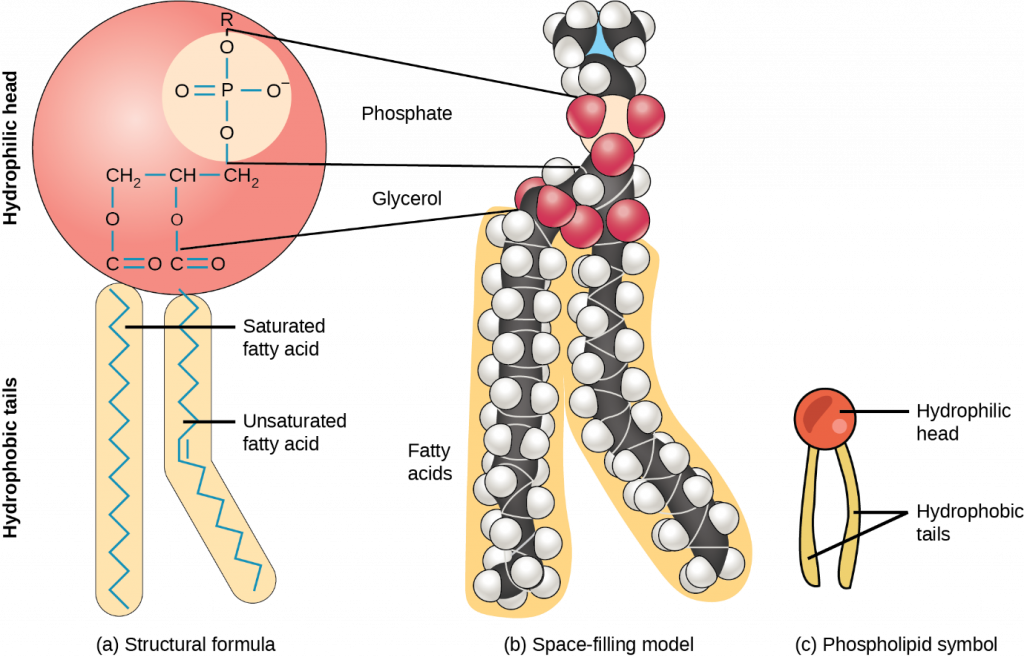
Phospholipids
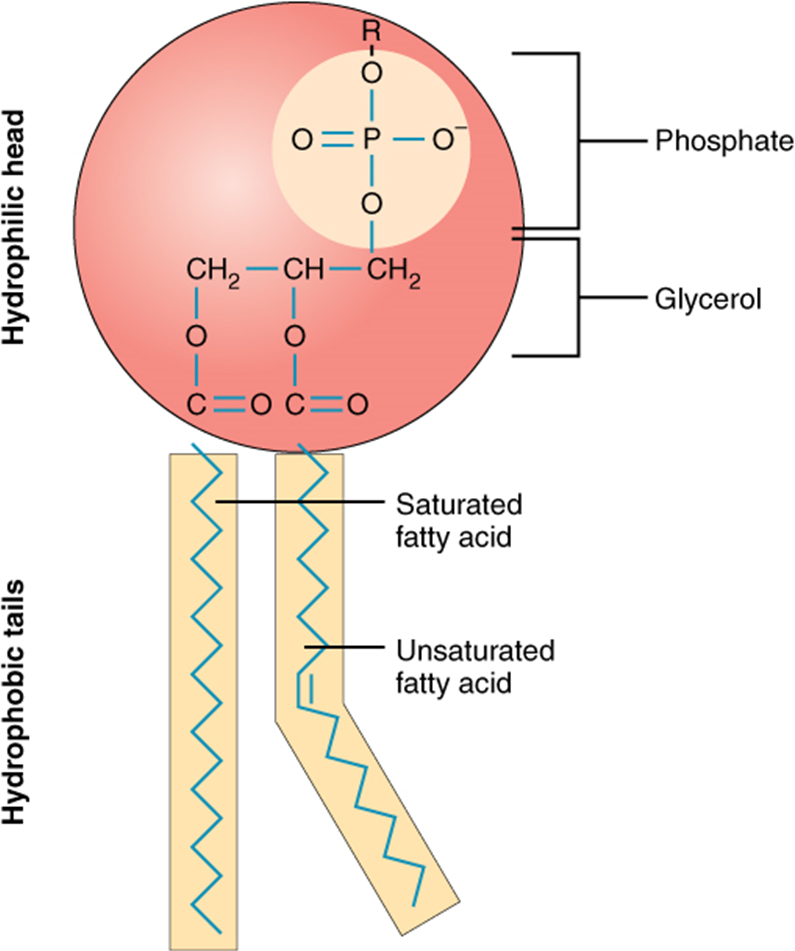
Phospholipids are major plasma membrane constituents that comprise cells’ outermost layer. Like fats, they are comprised of fatty acid chains attached to a glycerol or sphingosine backbone. However, instead of three fatty acids attached as in triglycerides, there are two fatty acids forming diacylglycerol, and a modified phosphate group occupies the glycerol backbone’s third carbon (Figure 2.3.8). A phosphate group alone attached to a diacylglycerol does not qualify as a phospholipid. It is phosphatidate (diacylglycerol 3-phosphate), the precursor of phospholipids. An alcohol modifies the phosphate group. Phosphatidylcholine and phosphatidylserine are two important phospholipids that are in plasma membranes.
Figure 2.3.8.Phospholipid. A phospholipid is a molecule with two fatty acids and a modified phosphate group attached to a glycerol backbone. Adding a charged or polar chemical group may modify the phosphate.
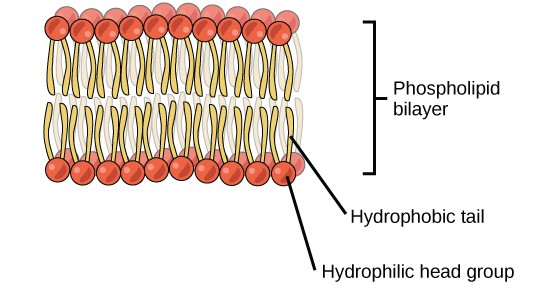
A phospholipid is an amphipathic molecule, meaning it has a hydrophobic and a hydrophilic part. The fatty acid chains are hydrophobic and cannot interact with water; whereas, the phosphate-containing group is hydrophilic and interacts with water (Figure 2.3.9).
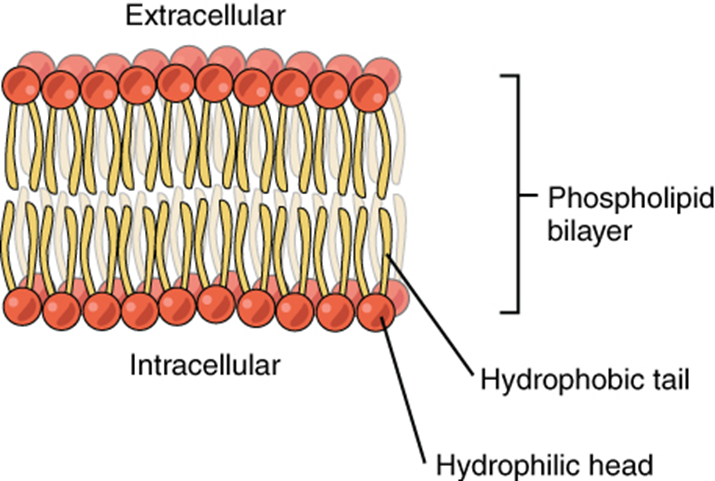
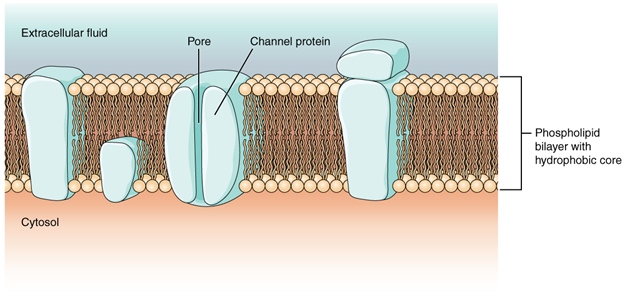
Figure 2.3.9. Phospholipid bilayer. The phospholipid bilayer is the major component of all cellular membranes. The hydrophilic head groups of the phospholipids face the aqueous solution. The hydrophobic tails are sequestered in the middle of the bilayer.
The head is the hydrophilic part, and the tail contains the hydrophobic fatty acids. In a membrane, a bilayer of phospholipids forms the structure’s matrix, phospholipids’ fatty acid tails face inside, away from water; whereas, the phosphate group faces the outside, aqueous side (Figure 2.3.9).
Phospholipids are responsible for the plasma membrane’s dynamic nature. If a drop of phospholipids is placed in water, it spontaneously forms a structure that scientists call a micelle, where the hydrophilic phosphate heads face the outside and the fatty acids face the structure’s interior.
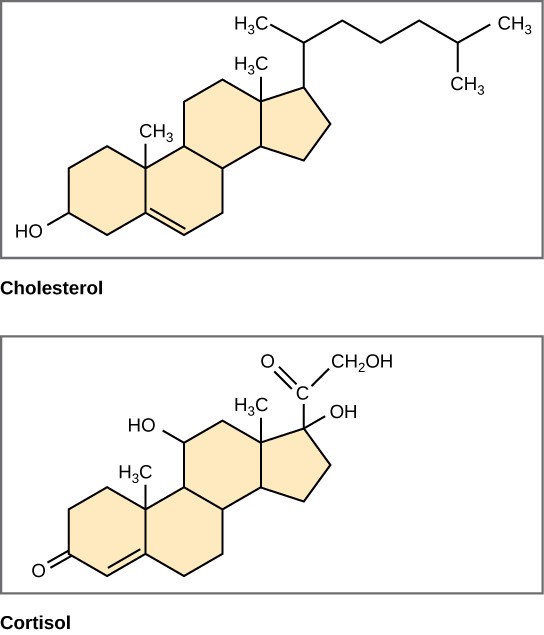
Steroids
Unlike the phospholipids and fats that was discussed earlier, steroids have a fused ring structure. Although they do not resemble the other lipids, scientists group them with them because they are also hydrophobic and insoluble in water. All steroids have four linked carbon rings and several of them, like cholesterol, have a short tail (Figure 2.3.10). Many steroids also have the –OH functional group, which puts them in the alcohol classification (sterols).
Figure 2.3.10.Steroids. Four fused hydrocarbon rings comprise steroids such as cholesterol and cortisol.
Cholesterol is the most common steroid. The liver synthesises cholesterol and is the precursor to many steroid hormones such as testosterone and oestradiol, which gonads and endocrine glands secrete. It is also the precursor to Vitamin D. Cholesterol is also the precursor of bile salts, which help emulsifying fats and their subsequent absorption by cells. Although lay people often speak negatively about cholesterol, it is necessary for the body’s proper functioning. Sterols (cholesterol in animal cells, phytosterol in plants) are components of the plasma membrane of cells and are found within the phospholipid bilayer.
Section Review
Lipids are a class of macromolecules that are nonpolar and hydrophobic in nature. Major types include fats and oils, waxes, phospholipids and steroids. Fats are a stored form of energy and are also known as triacylglycerols or triglycerides. Fats are comprised of fatty acids and either glycerol or sphingosine. Fatty acids may be unsaturated or saturated, depending on the presence or absence of double bonds in the hydrocarbon chain. If only single bonds are present, they are saturated fatty acids. Unsaturated fatty acids may have one or more double bonds in the hydrocarbon chain. Phospholipids comprise the membrane’s matrix. They have a glycerol or sphingosine backbone to which two fatty acid chains and a phosphate-containing group are attached. Steroids are another class of lipids. Their basic structure has four fused carbon rings. Cholesterol is a type of steroid and is an important constituent of the plasma membrane, where it helps to maintain the membrane’s fluid nature. It is also the precursor of steroid hormones such as testosterone.
Note: Content and images from this chapter have been adapted from Biology 2ndedition by Mary Ann Clark, Jung Choi and Matthew Douglas, used under a CC-BY license.
2.4 Protein
Learning Objectives
By the end of this section, you will be able to:
Describe the functions proteins perform in the cell and in tissues
Discuss the relationship between amino acids and proteins
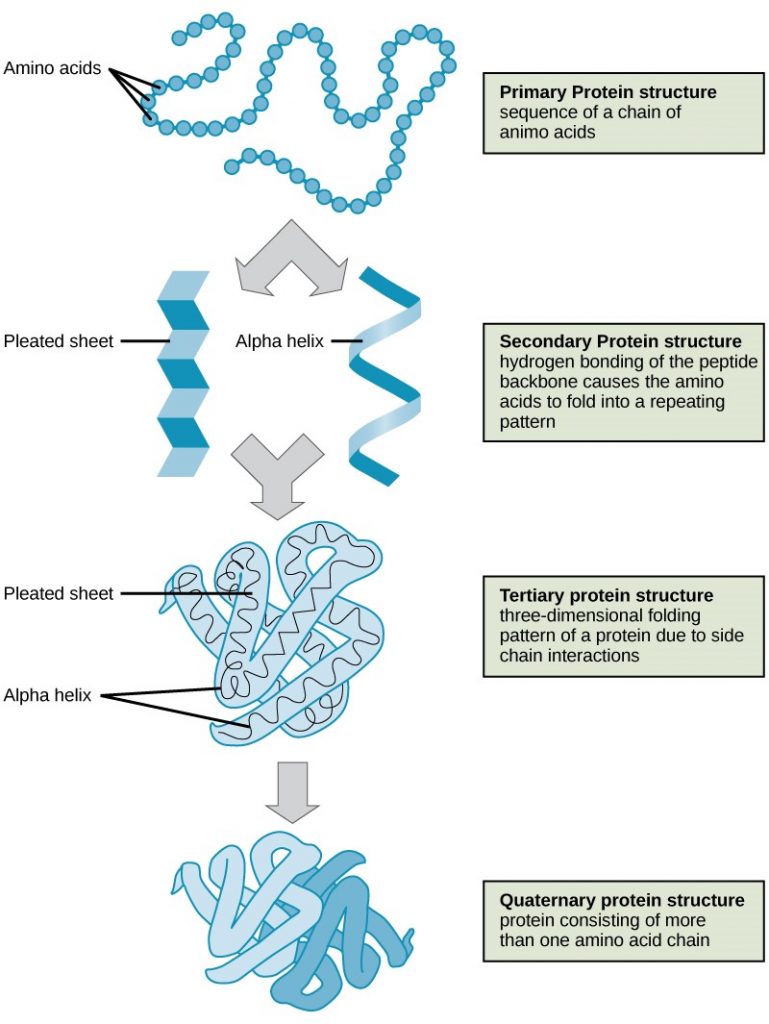
Explain the four levels of protein organisation
Describe the ways in which protein shape and function are linked
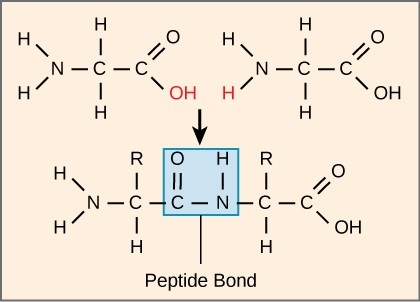
Proteins are one of the most abundant organic molecules in living systems and have the most diverse range of functions of all macromolecules. Proteins may be structural, regulatory, contractile, or protective. They may serve in transport, storage, or membranes; or they may be toxins or enzymes. Each cell in a living system may contain thousands of proteins, each with a unique function. Their structures, like their functions, vary greatly. They are all, however, amino acid polymers arranged in a linear sequence.
Types and Functions of Proteins
Enzymes, which living cells produce, are catalysts in biochemical reactions (like digestion) and are usually complex or conjugated proteins. Each enzyme is specific for the substrate (a reactant that binds to an enzyme) upon which it acts. The enzyme may help in breakdown, rearrangement, or synthesis reactions. We call enzymes that break down their substrates catabolic enzymes. Those that build more complex molecules from their substrates are anabolic enzymes, and enzymes that affect the rate of reaction are catalytic enzymes. Note that all enzymes increase the reaction rate and, therefore, are organic catalysts. An example of an enzyme is salivary amylase, which hydrolyses its substrate amylose, a component of starch.
Hormones are chemical-signalling molecules, usually small proteins or steroids, secreted by endocrine cells that act to control or regulate specific physiological processes, including growth, development, metabolism, and reproduction. For example, insulin is a protein hormone that helps regulate the blood glucose level. Table 2.4.1 lists the primary types and functions of proteins.
Protein Types and Functions
Table 2.4.1. Types of proteins and their functions.
Type
Examples
Functions
Digestive Enzymes
Amylase, lipase, pepsin, trypsin
Help in food by catabolising nutrients into monomeric units
Transport
Haemoglobin, albumin
Carry substances in the blood or lymph throughout the body
Structural
Actin, tubulin, keratin
Construct different structures, like the cytoskeleton
Hormones
Insulin, thyroxine
Coordinate different body systems’ activity
Defences
Immunoglobulins
Protect the body from foreign pathogens
Contractile
Actin, myosin
Effect muscle contraction
Storage
Legume storage proteins, egg white (albumin)
Provide nourishment in early embryo development and the seeding
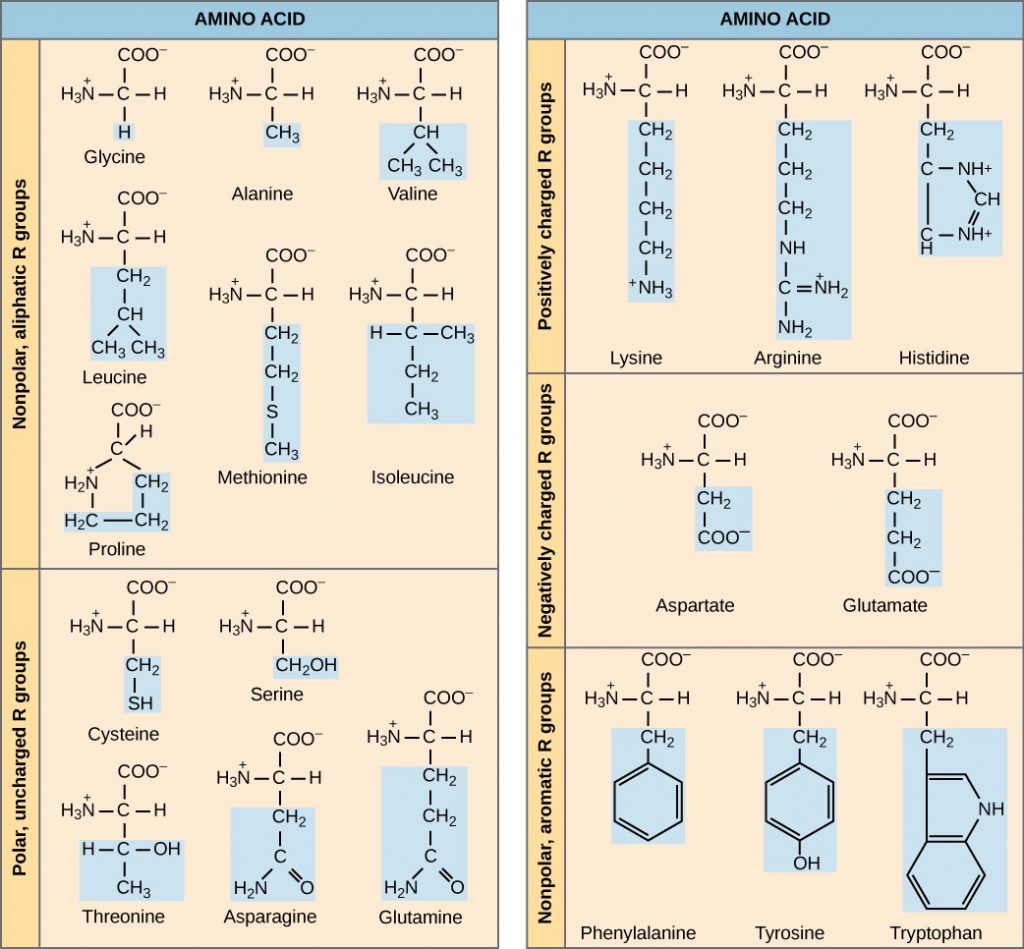
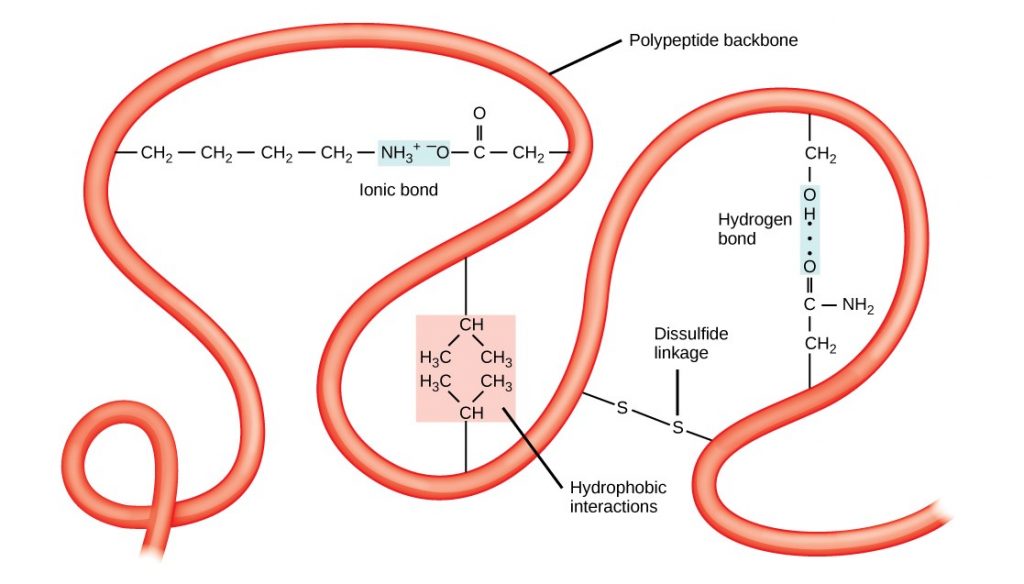
Proteins have different shapes and molecular weights. Some proteins are globular in shape; whereas, others are fibrous in nature, for example, haemoglobin is a globular protein, but collagen, located in our skin, is a fibrous protein. Protein shape is critical to its function, and many different types of chemical bonds maintain this shape. Changes in temperature, pH, and exposure to chemicals may lead to permanent changes in the protein’s shape, leading to loss of function, or denaturation. Different arrangements of the same 20 types of amino acids comprise all proteins. Two rare new amino acids were discovered recently (selenocystein and pirrolysine), and additional new discoveries may be added to the list.
Amino Acids
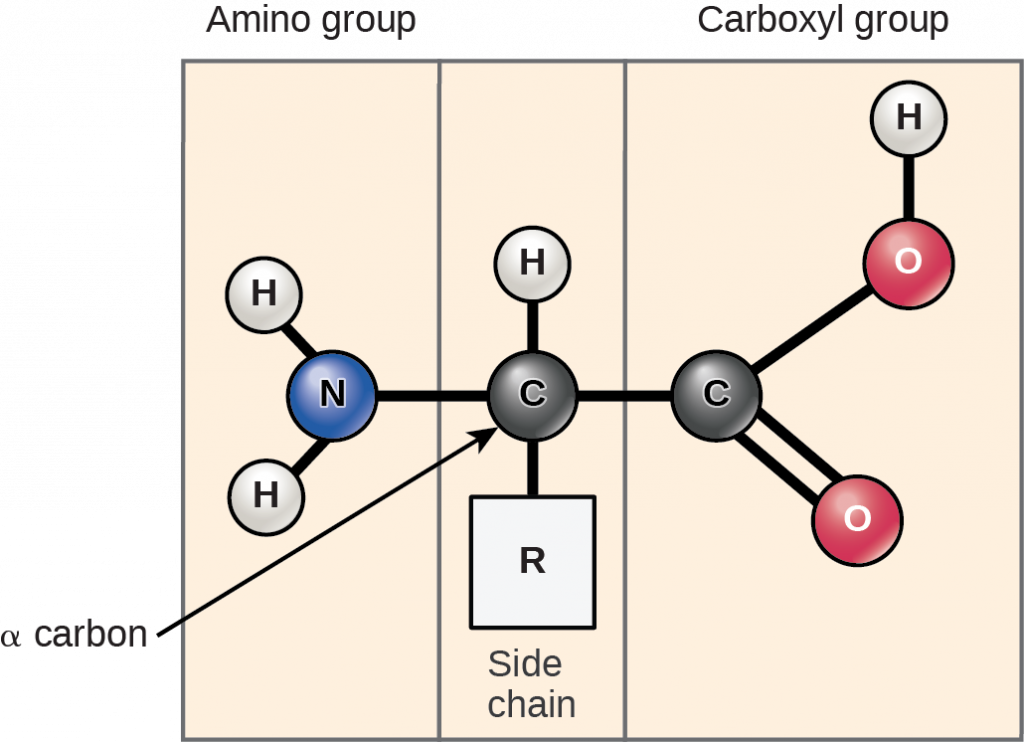
Amino acids are the monomers that comprise proteins. Each amino acid has the same fundamental structure, which consists of a central carbon atom, or the alpha (α) carbon, bonded to an amino group (-NH2), a carboxyl group (-COOH), and to a hydrogen atom. Every amino acid also has another atom or group of atoms bonded to the central atom known as the R group (Figure 2.4.1).
Figure 2.4.1.Amino acids. Amino acids have a central asymmetric carbon to which an amino group, a carboxyl group, a hydrogen atom, and a side chain (R group) are attached.
Scientists use the name “amino acid” because these acids contain both amino group and carboxyl-acid-group in their basic structure. As we mentioned, there are 20 common amino acids present in proteins. Nine of these are essential amino acids in humans because the human body cannot produce them and we obtain them from our diet. For each amino acid, the R group (or side chain) is different (Figure 2.4.2).
Figure 2.4.2. Common amino acids. There are 20 common amino acids commonly found in proteins, each with a different R group (variant group) that determines its chemical nature.
Which categories of amino acid would you expect to find on a soluble protein’s surface, and which would you expect to find in the interior? What distribution of amino acids would you expect to find in a protein embedded in a lipid bilayer?
The chemical nature of the side chain determines the amino acid’s nature (that is, whether it is acidic, basic, polar, or nonpolar). For example, the amino acid glycine has a hydrogen atom as the R group. Amino acids such as valine, methionine, and alanine are nonpolar or hydrophobic in nature, while amino acids such as serine, threonine, and cysteine are polar and have hydrophilic side chains. The side chains of lysine and arginine are positively charged, and therefore these amino acids are also basic amino acids. Proline has an R group that is linked to the amino group, forming a ring-like structure. Proline is an exception to the amino acid’s standard structure since its amino group is not separate from the side chain (Figure 2.4.2).
A single upper-case letter or a three-letter abbreviation represents amino acids, for example, the letter ‘V’ or the three-letter symbol ‘val’ represent the amino acid, valine. Just as some fatty acids are essential to a diet, some amino acids also are necessary. These essential amino acids in humans include isoleucine, leucine, and cysteine. Essential amino acids refer to those necessary to build proteins in the body, but not those that the body produces. Which amino acids are essential varies from organism to organism.
The sequence and the number of amino acids ultimately determine the protein’s shape, size, and function. A covalent bond, or peptide bond, attaches to each amino acid, which a dehydration reaction forms. One amino acid’s carboxyl group and the incoming amino acid’s amino group combine, releasing a water molecule. The resulting bond is the peptide bond (Figure 2.4.3).
Figure 2.4.3.Peptide bond. Peptide bond formation is a dehydration synthesis reaction. The carboxyl group of one amino acid is linked to the incoming amino acid’s amino group. In the process, it releases a water molecule.
The products that such linkages form are peptides. As more amino acids join to this growing chain, the resulting chain is a polypeptide. Each polypeptide has a free amino group at one end. This end the N terminal, or the amino terminal, and the other end has a free carboxyl group, also the C or carboxyl terminal. While the terms polypeptide and protein are sometimes used interchangeably, a polypeptide is technically a polymer of amino acids, whereas the term protein is used for a polypeptide or polypeptides that have combined together, often have bound non-peptide prosthetic groups, have a distinct shape, and have a unique function. After protein synthesis (translation), most proteins are modified. These are known as post-translational modifications. They may undergo cleavage, phosphorylation, or may require adding other chemical groups. Only after these modifications is the protein completely functional.
Evolution Connection
The Evolutionary Significance of Cytochrome c
Cytochrome c is an important component of the electron transport chain, a part of cellular respiration, and it is normally located in the cellular organelle, the mitochondrion. This protein has a haem prosthetic group, and the haem’s central ion alternately reduces and oxidises during electron transfer. Because this essential protein’s role in producing cellular energy is crucial, it has changed very little over millions of years. Protein sequencing has shown that there is a considerable amount of cytochrome c amino acid sequence homology among different species. In other words, we can assess evolutionary kinship by measuring the similarities or differences among various species’ DNA or protein sequences.
Scientists have determined that human cytochrome c contains 104 amino acids. For each cytochrome c molecule from different organisms that scientists have sequenced to date, 37 of these amino acids appear in the same position in all cytochrome c samples. This indicates that there may have been a common ancestor. On comparing the human and chimpanzee protein sequences, scientists did not find a sequence difference. When researchers compared human and rhesus monkey sequences, the single difference was in one amino acid. In another comparison, human to yeast sequencing shows a difference in the 44th position.
Protein Structure
As was discussed earlier, a protein’s shape is critical to its function, for example, an enzyme can bind to a specific substrate at an active site. If this active site is altered because of local changes or changes in overall protein structure, the enzyme may be unable to bind to the substrate. To understand how the protein gets its final shape or conformation, we need to understand the four levels of protein structure: primary, secondary, tertiary, and quaternary.
Primary Structure
Amino acids’ unique sequence in a polypeptide chain is its primary structure. For example, the pancreatic hormone insulin has two polypeptide chains, A and B, and they are linked together by disulphide bonds. The N terminal amino acid of the A chain is glycine; whereas, the C terminal amino acid is asparagine (Figure 2.4.4). The amino acid sequences in the A and B chains are unique to insulin.
Figure 2.4.4.Bovine serum insulin amino acid sequence. Bovine serum insulin is a protein hormone comprised of two peptide chains, A (21 amino acids long) and B (30 amino acids long). In each chain, three-letter abbreviations that represent the amino acids’ names in the order they are present indicate primary structure. The amino acid cysteine (cys) has a sulfhydryl (SH) group as a side chain. Two sulfhydryl groups can react in the presence of oxygen to form a disulphide (S-S) bond. Two disulphide bonds connect the A and B chains together, and a third helps the A chain fold into the correct shape. Note that all disulphide bonds are the same length, but we have drawn them different sizes for clarity.
The gene encoding the protein ultimately determines the unique sequence for every protein. A change in nucleotide sequence of the gene’s coding region may lead to adding a different amino acid to the growing polypeptide chain, causing a change in protein structure and function. In sickle cell anaemia, the haemoglobin β chain (a small portion of which we show in Figure 2.4.5) has a single amino acid substitution, causing a change in protein structure and function. Specifically, valine in the β chain substitutes the amino acid glutamic. What is most remarkable to consider is that a haemoglobin molecule is comprised of two alpha and two beta chains that each consist of about 150 amino acids. The molecule, therefore, has about 600 amino acids. The structural difference between a normal haemoglobin molecule and a sickle cell molecule—which dramatically decreases life expectancy—is a single amino acid of the 600. What is even more remarkable is that three nucleotides each encode those 600 amino acids, and a single base change (point mutation), 1 in 1800 bases causes the mutation.
Figure 2.4.5.Haemoglobin amino acid composition. The beta chain of haemoglobin is 147 residues in length, yet a single amino acid substitution leads to sickle cell anaemia. In normal haemoglobin, the amino acid at position seven is glutamate. In sickle cell haemoglobin, a valine replaces glutamate.
Because of this change of one amino acid in the chain, haemoglobin molecules form long fibres that distort the biconcave, or disc-shaped, red blood cells and causes them to assume a crescent or “sickle” shape, which clogs blood vessels (Figure 2.4.6). This can lead to myriad serious health problems such as breathlessness, dizziness, headaches and abdominal pain for those affected by this disease.
Figure 2.4.6.Blood smear. In this blood smear, visualised at 535x magnification using bright field microscopy, sickle cells are crescent shaped, while normal cells are disc-shaped. (Credit: modification of work by Ed Uthman; scale-bar data from Matt Russell).
Secondary Structure
The local folding of the polypeptide in some regions gives rise to the secondary structure of the protein. The most common are the α-helix and β-pleated sheet structures (Figure 2.4.7). Both structures are held in shape by hydrogen bonds. The hydrogen bonds form between the oxygen atom in the carbonyl group in one amino acid and another amino acid that is four amino acids farther along the chain.
Figure 2.4.7. Secondary protein structure. The α-helix and β-pleated sheet are secondary structures of proteins that form because of hydrogen bonding between carbonyl and amino groups in the peptide backbone. Certain amino acids have a propensity to form an α-helix, while others have a propensity to form a β-pleated sheet.
Every helical turn in an alpha helix has 3.6 amino acid residues. The polypeptide’s R groups (the variant groups) protrude out from the α-helix chain. In the β-pleated sheet, hydrogen bonding between atoms on the polypeptide chain’s backbone form the “pleats”. The R groups are attached to the carbons and extend above and below the pleat’s folds. The pleated segments align parallel or antiparallel to each other, and hydrogen bonds form between the partially positive hydrogen atom in the amino group and the partially negative oxygen atom in the peptide backbone’s carbonyl group. The α-helix and β-pleated sheet structures are in most globular and fibrous proteins and they play an important structural role.
Tertiary Structure
The polypeptide’s unique three-dimensional structure is its tertiary structure (Figure 2.4.8). This structure is in part due to chemical interactions at work on the polypeptide chain. Primarily, the interactions among R groups create the protein’s complex three-dimensional tertiary structure. The nature of the R groups in the amino acids involved can counteract forming the hydrogen bonds we described for standard secondary structures. For example, R groups with like charges repel each other and those with unlike charges are attracted to each other (ionic bonds). When protein folding takes place, the nonpolar amino acids’ hydrophobic R groups lie in the protein’s interior; whereas, the hydrophilic R groups lie on the outside. Scientists also call the former interaction types hydrophobic interactions. Interaction between cysteine side chains forms disulphide linkages in the presence of oxygen, the only covalent bond that forms during protein folding.
Figure 2.4.8.Protein tertiary structure. A variety of chemical interactions determine the proteins’ tertiary structure. These include hydrophobic interactions, ionic bonding, hydrogen bonding, and disulphide linkages.
All these interactions, weak and strong, determine the protein’s final three-dimensional shape. When a protein loses its three-dimensional shape, it may no longer be functional.
Quaternary Structure
In nature, some proteins form from several polypeptides, or subunits, and the interaction of these subunits forms the quaternary structure. Weak interactions between the subunits help to stabilise the overall structure. For example, insulin (a globular protein) has a combination of hydrogen and disulphide bonds that cause it to mostly clump into a ball shape. Insulin starts out as a single polypeptide and loses some internal sequences in the presence of post-translational modification after forming the disulphide linkages that hold the remaining chains together. Silk (a fibrous protein), however, has a β-pleated sheet structure that is the result of hydrogen bonding between different chains.
Figure 2.4.9 illustrates the four levels of protein structure (primary, secondary, tertiary, and quaternary).
Figure 2.4.9. Protein structure levels. Observe the four levels of protein structure in these illustrations. (Credit: modification of work by National Human Genome Research Institute).
Denaturation and Protein Folding
Each protein has its own unique sequence and shape that chemical interactions hold together. If the protein is subject to changes in temperature, pH, or exposure to chemicals, the protein structure may change, losing its shape without losing its primary sequence in what scientists call denaturation. Denaturation is often reversible because the polypeptide’s primary structure is conserved in the process if the denaturing agent is removed, allowing the protein to resume its function. Sometimes denaturation is irreversible, leading to loss of function. One example of irreversible protein denaturation is frying an egg. The albumin protein in the liquid egg white denatures when placed in a hot pan. Not all proteins denature at high temperatures. For instance, bacteria that survive in hot springs have proteins that function at temperatures close to boiling. The stomach is also very acidic, has a low pH and denatures proteins as part of the digestion process; however, the stomach’s digestive enzymes retain their activity under these conditions.
Protein folding is critical to its function. Scientists originally thought that the proteins themselves were responsible for the folding process. Only recently researchers discovered that often they receive assistance in the folding process from protein helpers, or chaperones (or chaperonins) that associate with the target protein during the folding process. They act by preventing polypeptide aggregation that comprise the complete protein structure, and they disassociate from the protein once the target protein is folded.
Section Review
Proteins are a class of macromolecules that perform a diverse range of functions for the cell. They help in metabolism by acting as enzymes, carriers, or hormones, and provide structural support. The building blocks of proteins (monomers) are amino acids. Each amino acid has a central carbon that bonds to an amino group, a carboxyl group, a hydrogen atom, and an R group or side chain. There are 20 commonly occurring amino acids, each of which differs in the R group. A peptide bond links each amino acid to its neighbours. A long amino acid chain is a polypeptide.
Proteins are organised at four levels: primary, secondary, tertiary, and (optional) quaternary. The primary structure is the amino acids’ unique sequence. The polypeptide’s local folding to form structures such as the α-helix and β-pleated sheet constitutes the secondary structure. The overall three-dimensional structure is the tertiary structure. When two or more polypeptides combine to form the complete protein structure, the configuration is the protein’s quaternary structure. Protein shape and function are intricately linked. Any change in shape caused by changes in temperature or pH may lead to protein denaturation and a loss in function.
Note: Content and images from this chapter have been adapted from Biology 2ndedition by Mary Ann Clark, Jung Choi and Matthew Douglas, used under a CC-BY license.
2.5 Nucleic Acid
Learning Objectives
By the end of this section, you will be able to:
Describe nucleic acids’ structure and define the two types of nucleic acids
Explain DNA structure and role
Explain RNA structure and roles
Nucleic acids are the most important macromolecules for the continuity of life. They carry the cell’s genetic blueprint and carry instructions for its functioning.
DNA and RNA
The two main types of nucleic acids are deoxyribonucleic acid(DNA) and ribonucleic acid (RNA). DNA is the genetic material in all living organisms, ranging from single-celled bacteria to multicellular mammals. It is in the nucleus of eukaryotes and in the organelles, chloroplasts, and mitochondria. In prokaryotes, the DNA is not enclosed in a membranous envelope.
The cell’s entire genetic content is its genome, and the study of genomes is genomics. In eukaryotic cells but not in prokaryotes, DNA forms a complex with histone proteins to form chromatin, the substance of eukaryotic chromosomes. A chromosome may contain tens of thousands of genes. Many genes contain the information to make protein products. Other genes code for RNA products. DNA controls all the cellular activities by turning the genes “on” or “off.”
The other type of nucleic acid, RNA, is mostly involved in protein synthesis. The DNA molecules never leave the nucleus but instead use an intermediary to communicate with the rest of the cell. This intermediary is the messenger RNA (mRNA). Other types of RNA—like rRNA, tRNA, and microRNA—are involved in protein synthesis and its regulation.
DNA and RNA are comprised of monomers that scientists call nucleotides. The nucleotides combine with each other to form a polynucleotide, DNA or RNA. Three components comprise each nucleotide: a nitrogenous base, a pentose (five-carbon) sugar, and a phosphate group (Figure 2.5.1). Each nitrogenous base in a nucleotide is attached to a sugar molecule, which is attached to one or more phosphate groups.
Figure 2.5.1.Nucleotide components. Three components comprise a nucleotide: a nitrogenous base, a pentose sugar, and one or more phosphate groups. Carbon residues in the pentose are numbered 1′ through 5′ (the prime distinguishes these residues from those in the base, which are numbered without using a prime notation). The base is attached to the ribose’s 1′ position, and the phosphate is attached to the 5′ position. When a polynucleotide forms, the incoming nucleotide’s 5′ phosphate attaches to the 3′ hydroxyl group at the end of the growing chain. Two types of pentose are in nucleotides, deoxyribose (found in DNA) and ribose (found in RNA). Deoxyribose is similar in structure to ribose, but it has an H instead of an OH at the 2′ position. We can divide bases into two categories: purines and pyrimidines. Purines have a double ring structure, and pyrimidines have a single ring.
The nitrogenous bases, important components of nucleotides, are organic molecules and are so named because they contain carbon and nitrogen. They are bases because they contain an amino group that has the potential of binding an extra hydrogen, and thus decreasing the hydrogen ion concentration in its environment, making it more basic. Each nucleotide in DNA contains one of four possible nitrogenous bases: adenine (A), guanine (G) cytosine (C), and thymine (T).
Scientists classify adenine and guanine as purines. The purine’s primary structure is two carbon-nitrogen rings. Scientists classify cytosine, thymine, and uracil as pyrimidines which have a single carbon-nitrogen ring as their primary structure (Figure 2.5.1). Each of these basic carbon-nitrogen rings has different functional groups attached to it. In molecular biology shorthand, we know the nitrogenous bases by their symbols A, T, G, C, and U. DNA contains A, T, G, and C; whereas, RNA contains A, U, G, and C.
The pentose sugar in DNA is deoxyribose, and in RNA, the sugar is ribose (Figure 3.5.1). The difference between the sugars is the presence of the hydroxyl group on the ribose’s second carbon and hydrogen on the deoxyribose’s second carbon. The carbon atoms of the sugar molecule are numbered as 1′, 2′, 3′, 4′, and 5′ (1′ is read as “one prime”). The phosphate residue attaches to the hydroxyl group of the 5′ carbon of one sugar and the hydroxyl group of the 3′ carbon of the sugar of the next nucleotide, which forms a 5′–3′ phosphodiester linkage. A simple dehydration reaction like the other linkages connecting monomers in macromolecules does not form the phosphodiester linkage. Its formation involves removing two phosphate groups. A polynucleotide may have thousands of such phosphodiester linkages.
DNA Double-Helix Structure
Figure 2.5.2. Native DNA is an antiparallel double helix. The phosphate backbone (indicated by the curvy lines) is on the outside, and the bases are on the inside. Each base from one strand interacts via hydrogen bonding with a base from the opposing strand. (Credit: Jerome Walker/Dennis Myts).
DNA has a double-helix structure (Figure 2.5.2). The sugar and phosphate lie on the outside of the helix, forming the DNA’s backbone. The nitrogenous bases are stacked in the interior, like a pair of staircase steps. Hydrogen bonds bind the pairs to each other. Every base pair in the double helix is separated from the next base pair by 0.34 nm. The helix’s two strands run in opposite directions, meaning that the 5′ carbon end of one strand will face the 3′ carbon end of its matching strand. (Scientists call this an antiparallel orientation and is important to DNA replication and in many nucleic acid interactions.)
Only certain types of base pairing are allowed. For example, a certain purine can only pair with a certain pyrimidine. This means A can pair with T, and G can pair with C, as Figure 2.5.3 shows. This is the base complementary rule. In other words, the DNA strands are complementary to each other. If the sequence of one strand is AATTGGCC, the complementary strand would have the sequence TTAACCGG. During DNA replication, each strand copies itself, resulting in a daughter DNA double helix containing one parental DNA strand and a newly synthesised strand.
Figure 2.5.3. Hydrogen bonds in a double stranded DNA molecule. In a double stranded DNA molecule, the two strands run antiparallel to one another so that one strand runs 5′ to 3′ and the other 3′ to 5′. The phosphate backbone is located on the outside, and the bases are in the middle. Adenine forms hydrogen bonds (or base pairs) with thymine, and guanine base pairs with cytosine.
A mutation occurs, and adenine replaces cytosine. What impact do you think this will have on the DNA structure?
RNA
Ribonucleic acid, or RNA, is mainly involved in the process of protein synthesis under the direction of DNA. RNA is usually single-stranded and is comprised of ribonucleotides that are linked by phosphodiester bonds. A ribonucleotide in the RNA chain contains ribose (the pentose sugar), one of the four nitrogenous bases (A, U, G, and C), and the phosphate group.
There are four major types of RNA: messenger RNA (mRNA), ribosomal RNA (rRNA), transfer RNA (tRNA), and microRNA (miRNA). The first, mRNA, carries the message from DNA, which controls all the cellular activities in a cell. If a cell requires synthesising a certain protein, the gene for this product turns “on” and the messenger RNA synthesises in the nucleus. The RNA base sequence is complementary to the DNA’s coding sequence from which it has been copied. However, in RNA, the base T is absent and U is present instead. If the DNA strand has a sequence AATTGCGC, the sequence of the complementary RNA is UUAACGCG. In the cytoplasm, the mRNA interacts with ribosomes and other cellular machinery (Figure 2.5.4).
Figure 2.5.4.Ribosome. A ribosome has two parts: a large subunit and a small subunit. The mRNA sits in between the two subunits. A tRNA molecule recognises a codon on the mRNA, binds to it by complementary base pairing, and adds the correct amino acid to the growing peptide chain.
The mRNA is read in sets of three bases known as codons. Each codon codes for a single amino acid. In this way, the mRNA is read and the protein product is made. Ribosomal RNA (rRNA) is a major constituent of ribosomes on which the mRNA binds. The rRNA ensures the proper alignment of the mRNA and the Ribosomes. The ribosome’s rRNA also has an enzymatic activity (peptidyl transferase) and catalyses peptide bond formation between two aligned amino acids. Transfer RNA (tRNA) is one of the smallest of the four types of RNA, usually 70–90 nucleotides long. It carries the correct amino acid to the protein synthesis site. It is the base pairing between the tRNA and mRNA that allows for the correct amino acid to insert itself in the polypeptide chain. MicroRNAs are the smallest RNA molecules and their role involves regulating gene expression by interfering with the expression of certain mRNA messages. Table 2.5.1 summarises DNA and RNA features.
Table 2.5.1 DNA and RNA Features
DNA
RNA
Function
Carries genetic information
Involved in protein synthesis
Location
Remains in the nucleus
Leaves the nucleus
Structure
Double helix
Usually single-stranded
Sugar
Deoxyribose
Ribose
Pyrimidines
Cytosine, thymine
Cytosine, uracil
Purines
Adenine, guanine
Adenine, guanine
Even though the RNA is single stranded, most RNA types show extensive intramolecular base pairing between complementary sequences, creating a predictable three-dimensional structure essential for their function.
As you have learned, information flow in an organism takes place from DNA to RNA to protein. DNA dictates the structure of mRNA in a process scientists call transcription, and RNA dictates the protein’s structure in a process scientists call translation. This is the Central Dogma of Life, which holds true for all organisms; however, exceptions to the rule occur in connection with viral infections.
Section Review
Nucleic acids are molecules comprised of nucleotides that direct cellular activities such as cell division and protein synthesis. Pentose sugar, a nitrogenous base, and a phosphate group comprise each nucleotide. There are two types of nucleic acids: DNA and RNA. DNA carries the cell’s genetic blueprint and passes it on from parents to offspring (in the form of chromosomes). It has a double-helical structure with the two strands running in opposite directions, connected by hydrogen bonds, and complementary to each other. RNA is a single-stranded polymer composed of linked nucleotides made up of a pentose sugar (ribose), a nitrogenous base, and a phosphate group. RNA is involved in protein synthesis and its regulation. Messenger RNA (mRNA) copies from the DNA, exports itself from the nucleus to the cytoplasm, and contains information for constructing proteins. Ribosomal RNA (rRNA) is a part of the ribosomes at the site of protein synthesis; whereas, transfer RNA (tRNA) carries the amino acid to the site of protein synthesis. The microRNA regulates using mRNA for protein synthesis.
Note: Content and images from this chapter have been adapted from Biology 2nd edition by Mary Ann Clark, Jung Choi and Matthew Douglas, used under a CC-BY license.
2.6 The Cell Membrane
Learning Objectives
By the end of this section, you will be able to:
Describe the molecular components that make up the cell membrane
Explain the major features and properties of the cell membrane
Differentiate between materials that can and cannot diffuse through the lipid bilayer
Recognise different types of passive transport and active transport, providing examples of each
Despite differences in structure and function, all living cells in multicellular organisms have a surrounding cell membrane. As the outer layer of your skin separates your body from its environment, the cell membrane (also known as the plasma membrane) separates the inner contents of a cell from its exterior environment. This cell membrane provides a protective barrier around the cell and regulates which materials can pass in or out.
Structure and Composition of the Cell Membrane
The cell membrane is an extremely pliable structure composed primarily of back-to-back phospholipids (a “bilayer”). Cholesterol is also present, which contributes to the fluidity of the membrane, and there are various proteins embedded within the membrane that have a variety of functions.
A single phospholipid molecule has a phosphate group on one end, called the “head,” and two side-by-side chains of fatty acids that make up the lipid tails (Figure 2.6.1). The phosphate group is negatively charged, making the head polar and hydrophilic—or “water loving.” A hydrophilic molecule (or region of a molecule) is one that is attracted to water. The phosphate heads are thus attracted to the water molecules of both the extracellular and intracellular environments. The lipid tails, on the other hand, are uncharged, or nonpolar, and are hydrophobic—or “water fearing.” A hydrophobic molecule (or region of a molecule) repels and is repelled by water. Some lipid tails consist of saturated fatty acids and some contain unsaturated fatty acids. This combination adds to the fluidity of the tails that are constantly in motion. Phospholipids are thus amphipathic molecules. An amphipathic molecule is one that contains both a hydrophilic and a hydrophobic region. In fact, soap works to remove oil and grease stains because it has amphipathic properties. The hydrophilic portion can dissolve in water while the hydrophobic portion can trap grease in micelles that then can be washed away.
Figure 2.6.1.Phospholipid structure. A phospholipid molecule consists of a polar phosphate “head,” which is hydrophilic and a non-polar lipid “tail,” which is hydrophobic. Unsaturated fatty acids result in kinks in the hydrophobic tails.
The cell membrane consists of two adjacent layers of phospholipids. The lipid tails of one layer face the lipid tails of the other layer, meeting at the interface of the two layers. The phospholipid heads face outward, one layer exposed to the interior of the cell and one layer exposed to the exterior (Figure 2.6.2). Because the phosphate groups are polar and hydrophilic, they are attracted to water in the intracellular fluid. Intracellular fluid (ICF) is the fluid interior of the cell. The phosphate groups are also attracted to the extracellular fluid. Extracellular fluid (ECF) is the fluid environment outside the enclosure of the cell membrane. Interstitial fluid (IF) is the term given to extracellular fluid not contained within blood vessels. Because the lipid tails are hydrophobic, they meet in the inner region of the membrane, excluding watery intracellular and extracellular fluid from this space. The cell membrane has many proteins, as well as other lipids (such as cholesterol), that are associated with the phospholipid bilayer. An important feature of the membrane is that it remains fluid; the lipids and proteins in the cell membrane are not rigidly locked in place.
Figure 2.6.2. Phospholipid bilayer. The phospholipid bilayer consists of two adjacent sheets of phospholipids, arranged tail to tail. The hydrophobic tails associate with one another, forming the interior of the membrane. The polar heads contact the fluid inside and outside of the cell.
Membrane Proteins
The lipid bilayer forms the basis of the cell membrane, but it is peppered throughout with various proteins. Two different types of proteins that are commonly associated with the cell membrane are the integral proteins and peripheral protein (Figure 2.6.3). As its name suggests, an integral protein is a protein that is embedded in the membrane. A channel protein is an example of an integral protein that selectively allows particular materials, such as certain ions, to pass into or out of the cell.
Figure 2.6.3. Cell Membrane. The cell membrane of the cell is a phospholipid bilayer containing many different molecular components, including proteins and cholesterol, some with carbohydrate groups attached.
Another important group of integral proteins are cell recognition proteins, which serve to mark a cell’s identity so that it can be recognised by other cells. A receptor is a type of recognition protein that can selectively bind a specific molecule outside the cell, and this binding induces a chemical reaction within the cell. A ligand is the specific molecule that binds to and activates a receptor. Some integral proteins serve dual roles as both a receptor and an ion channel. One example of a receptor-ligand interaction is the receptors on nerve cells that bind neurotransmitters, such as dopamine. When a dopamine molecule binds to a dopamine receptor protein, a channel within the transmembrane protein opens to allow certain ions to flow into the cell.
Some integral membrane proteins are glycoproteins. A glycoprotein is a protein that has carbohydrate molecules attached, which extend into the extracellular matrix. The attached carbohydrate tags on glycoproteins aid in cell recognition. The carbohydrates that extend from membrane proteins and even from some membrane lipids collectively form the glycocalyx. The glycocalyx is a fuzzy-appearing coating around the cell formed from glycoproteins and other carbohydrates attached to the cell membrane. The glycocalyx can have various roles. For example, it may have molecules that allow the cell to bind to another cell, it may contain receptors for hormones, or it might have enzymes to break down nutrients. The glycocalyces found in a person’s body are products of that person’s genetic makeup. They give each of the individual’s trillions of cells the “identity” of belonging in the person’s body. This identity is the primary way that a person’s immune defence cells “know” not to attack the person’s own body cells, but it also is the reason organs donated by another person might be rejected.
Peripheral proteins are typically found on the inner or outer surface of the lipid bilayer but can also be attached to the internal or external surface of an integral protein. These proteins typically perform a specific function for the cell. Some peripheral proteins on the surface of intestinal cells, for example, act as digestive enzymes to break down nutrients to sizes that can pass through the cells and into the bloodstream.
Transport Across the Cell
One of the great wonders of the cell membrane is its ability to regulate the concentration of substances inside the cell. These substances include ions such as Ca2+, Na+, K+ and Cl–; nutrients including sugars, fatty acids, and amino acids; and waste products, particularly carbon dioxide (CO2), which must leave the cell.
The membrane’s lipid bilayer structure provides the first level of control. The phospholipids are tightly packed together, and the membrane has a hydrophobic interior. This structure causes the membrane to be selectively permeable. A membrane that has selective permeability allows only substances meeting certain criteria to pass through it unaided. In the case of the cell membrane, only relatively small, nonpolar materials can move through the lipid bilayer (remember, the lipid tails of the membrane are nonpolar). Some examples of these are other lipids, oxygen and carbon dioxide gases, and alcohol. However, water-soluble materials—like glucose, amino acids, and electrolytes—need some assistance to cross the membrane because they are repelled by the hydrophobic tails of the phospholipid bilayer. All substances that move through the membrane do so by one of two general methods, which are categorised based on whether or not energy is required. Passive transport is the movement of substances across the membrane without the expenditure of cellular energy. In contrast, active transport is the movement of substances across the membrane using energy from adenosine triphosphate (ATP).
Passive Transport
In order to understand how substances move passively across a cell membrane, it is necessary to understand concentration gradients and diffusion. A concentration gradient is the difference in concentration of a substance across a space. Molecules (or ions) will spread/diffuse from where they are more concentrated to where they are less concentrated until they are equally distributed in that space. (When molecules move in this way, they are said to move down their concentration gradient.) Diffusion is the movement of particles from an area of higher concentration to an area of lower concentration. A couple of common examples will help to illustrate this concept. Imagine being inside a closed bathroom. If a bottle of perfume were sprayed, the scent molecules would naturally diffuse from the spot where they left the bottle to all corners of the bathroom, and this diffusion would go on until no more concentration gradient remains. Another example is a spoonful of sugar placed in a cup of tea. Eventually the sugar will diffuse throughout the tea until no concentration gradient remains. In both cases, if the room is warmer or the tea hotter, diffusion occurs even faster as the molecules are bumping into each other and spreading out faster than at cooler temperatures. Having an internal body temperature around 37oC (98.6° F) thus also aids in diffusion of particles within the body.
Whenever a substance exists in greater concentration on one side of a semipermeable membrane, such as the cell membranes, any substance that can move down its concentration gradient across the membrane will do so. Consider substances that can easily diffuse through the lipid bilayer of the cell membrane, such as the gases oxygen (O2) and CO2. O2 generally diffuses into cells because it is more concentrated outside of them, and CO2 typically diffuses out of cells because it is more concentrated inside of them. Neither of these examples requires any energy on the part of the cell, and therefore they use passive transport to move across the membrane.
Before moving on, you need to review the gases that can diffuse across a cell membrane. Because cells rapidly use up oxygen during metabolism, there is typically a lower concentration of O2 inside the cell than outside. As a result, oxygen will diffuse from the interstitial fluid directly through the lipid bilayer of the membrane and into the cytoplasm within the cell. On the other hand, because cells produce CO2 as a by-product of metabolism, CO2 concentrations rise within the cytoplasm; therefore, CO2 will move from the cell through the lipid bilayer and into the interstitial fluid, where its concentration is lower. This mechanism of molecules moving across a cell membrane from the side where they are more concentrated to the side where they are less concentrated is a form of passive transport called simple diffusion (Figure 2.6.4).
Figure 2.6.4. Simple diffusion across the cell (plasma) membrane. The structure of the lipid bilayer allows small, uncharged substances such as oxygen and carbon dioxide, and hydrophobic molecules such as lipids, to pass through the cell membrane, down their concentration gradient, by simple diffusion.
Large polar or ionic molecules, which are hydrophilic, cannot easily cross the phospholipid bilayer. Very small polar molecules, such as water, can cross via simple diffusion due to their small size. Charged atoms or molecules of any size cannot cross the cell membrane via simple diffusion as the charges are repelled by the hydrophobic tails in the interior of the phospholipid bilayer. Solutes dissolved in water on either side of the cell membrane will tend to diffuse down their concentration gradients, but because most substances cannot pass freely through the lipid bilayer of the cell membrane, their movement is restricted to protein channels and specialised transport mechanisms in the membrane. Facilitated diffusion is the diffusion process used for those substances that cannot cross the lipid bilayer due to their size, charge, and/or polarity (Figure 2.6.5). A common example of facilitated diffusion is the movement of glucose into the cell, where it is used to make ATP. Although glucose can be more concentrated outside of a cell, it cannot cross the lipid bilayer via simple diffusion because it is both large and polar. To resolve this, a specialised carrier protein called the glucose transporter will transfer glucose molecules into the cell to facilitate its inward diffusion.
Figure 2.6.5. Facilitated diffusion. (a) Facilitated diffusion of substances crossing the cell (plasma) membrane takes place with the help of proteins such as channel proteins and carrier proteins. Channel proteins are less selective than carrier proteins, and usually mildly discriminate between their cargo based on size and charge. (b) Carrier proteins are more selective, often only allowing one particular type of molecule to cross.
As an example, even though sodium ions (Na+) are highly concentrated outside of cells, these electrolytes are charged and cannot pass through the nonpolar lipid bilayer of the membrane. Their diffusion is facilitated by membrane proteins that form sodium channels (or “pores”), so that Na+ ions can move down their concentration gradient from outside the cells to inside the cells. There are many other solutes that must undergo facilitated diffusion to move into a cell, such as amino acids, or to move out of a cell, such as wastes. Because facilitated diffusion is a passive process, it does not require energy expenditure by the cell.
Water also can move freely across the cell membrane of all cells, either through protein channels or by slipping between the lipid tails of the membrane itself. Osmosis is the diffusion of water through a semipermeable membrane (Figure 2.6.6).
Figure 2.6.6. Osmotic diffusion. Osmosis is the diffusion of water through a semipermeable membrane down its concentration gradient. If a membrane is permeable to water, though not to a solute, water will equalise its own concentration by diffusing to the side of lower water concentration (and thus the side of higher solute concentration). In the beaker on the left, the solution on the right side of the membrane is hypertonic.
The movement of water molecules is not itself regulated by cells, so it is important that cells are exposed to an environment in which the concentration of solutes outside of the cells (in the extracellular fluid) is equal to the concentration of solutes inside the cells (in the cytoplasm). Two solutions that have the same concentration of solutes are said to be isotonic (equal tension). When cells and their extracellular environments are isotonic, the concentration of water molecules is the same outside and inside the cells, and the cells maintain their normal shape (and function).
Osmosis occurs when there is an imbalance of solutes outside of a cell versus inside the cell. A solution that has a higher concentration of solutes than another solution is said to be hypertonic, and water molecules tend to diffuse into a hypertonic solution (Figure 2.6.7). Cells in a hypertonic solution will shrivel as water leaves the cell via osmosis. In contrast, a solution that has a lower concentration of solutes than another solution is said to be hypotonic, and water molecules tend to diffuse out of a hypotonic solution. Cells in a hypotonic solution will take on too much water and swell, with the risk of eventually bursting. A critical aspect of homeostasis in living things is to create an internal environment in which all of the body’s cells are in an isotonic solution. Various organ systems, particularly the kidneys, work to maintain this homeostasis.
Figure 2.6.7. Concentration of solutions. A hypertonic solution has a solute concentration higher than another solution. An isotonic solution has a solute concentration equal to another solution. A hypotonic solution has a solute concentration lower than another solution.
Another mechanism besides diffusion to passively transport materials between compartments is filtration. Unlike diffusion of a substance from where it is more concentrated to less concentrated, filtration uses a hydrostatic pressure gradient that pushes the fluid—and the solutes within it—from a higher pressure area to a lower pressure area. Filtration is an extremely important process in the body. For example, the circulatory system uses filtration to move plasma and substances across the endothelial lining of capillaries and into surrounding tissues, supplying cells with the nutrients. Filtration pressure in the kidneys provides the mechanism to remove wastes from the bloodstream.
Active Transport
For all the transport methods described above, the cell expends no energy. Membrane proteins that aid in the passive transport of substances do so without the use of ATP. During active transport, ATP is required to move a substance across a membrane, often with the help of protein carriers, and usually against its concentration gradient.
One of the most common types of active transport involves proteins that serve as pumps. The word “pump” probably conjures up thoughts of using energy to pump up the tire of a bicycle or a basketball. Similarly, energy from ATP is required for these membrane proteins to transport substances—molecules or ions—across the membrane, usually against their concentration gradients (from an area of low concentration to an area of high concentration).
The sodium-potassium pump, which is also called Na+/K+ ATPase, transports sodium out of a cell while moving potassium into the cell. The Na+/K+ pump is an important ion pump found in the membranes of many types of cells. These pumps are particularly abundant in nerve cells, which are constantly pumping out sodium ions and pulling in potassium ions to maintain an electrical gradient across their cell membranes. An electrical gradient is a difference in electrical charge across a space. In the case of nerve cells, for example, the electrical gradient exists between the inside and outside of the cell, with the inside being negatively-charged (at around -70 mV) relative to the outside. The negative electrical gradient is maintained because each Na+/K+ pump moves three Na+ ions out of the cell and two K+ ions into the cell for each ATP molecule that is used (Figure 2.6.8). This process is so important for nerve cells that it accounts for the majority of their ATP usage.
Figure 2.6.8. Sodium-potassium pump. The sodium-potassium pump is found in many cell (plasma) membranes. Powered by ATP, the pump moves sodium and potassium ions in opposite directions, each against its concentration gradient. In a single cycle of the pump, three sodium ions are extruded from and two potassium ions are imported into the cell.
Active transport pumps can also work together with other active or passive transport systems to move substances across the membrane. For example, the sodium-potassium pump maintains a high concentration of sodium ions outside of the cell. Therefore, if the cell needs sodium ions, all it has to do is open a passive sodium channel, as the concentration gradient of the sodium ions will drive them to diffuse into the cell. In this way, the action of an active transport pump (the sodium-potassium pump) powers the passive transport of sodium ions by creating a concentration gradient. When active transport powers the transport of another substance in this way, it is called secondary active transport.
Symporters are secondary active transporters that move two substances in the same direction. For example, the sodium-glucose symporter uses sodium ions to “pull” glucose molecules into the cell. Because cells store glucose for energy, glucose is typically at a higher concentration inside of the cell than outside. However, due to the action of the sodium-potassium pump, sodium ions will easily diffuse into the cell when the symporter is opened. The flood of sodium ions through the symporter provides the energy that allows glucose to move through the symporter and into the cell, against its concentration gradient.
Conversely, antiporters are secondary active transport systems that transport substances in opposite directions. For example, the sodium-hydrogen ion antiporter uses the energy from the inward flood of sodium ions to move hydrogen ions (H+) out of the cell. The sodium-hydrogen antiporter is used to maintain the pH of the cell’s interior.
Other forms of active transport do not involve membrane carriers. Endocytosis (bringing “into the cell”) is the process of a cell ingesting material by enveloping it in a portion of its cell membrane, and then pinching off that portion of membrane (Figure 2.6.9). Once pinched off, the portion of membrane and its contents becomes an independent, intracellular vesicle. A vesicle is a membranous sac—a spherical and hollow organelle bounded by a lipid bilayer membrane. Endocytosis often brings materials into the cell that must be broken down or digested. Phagocytosis (“cell eating”) is the endocytosis of large particles. Many immune cells engage in phagocytosis of invading pathogens. Like little Pac-men, their job is to patrol body tissues for unwanted matter, such as invading bacterial cells, phagocytose them, and digest them. In contrast to phagocytosis, pinocytosis (“cell drinking”) brings fluid containing dissolved substances into a cell through membrane vesicles.
Figure 2.6.9. Three forms of endocytosis. Endocytosis is a form of active transport in which a cell envelopes extracellular materials using its cell membrane. (a) In phagocytosis, which is relatively nonselective, the cell takes in a large particle. (b) In pinocytosis, the cell takes in small particles in fluid. (c) In contrast, receptor-mediated endocytosis is quite selective. When external receptors bind a specific ligand, the cell responds by endocytosing the ligand.
Phagocytosis and pinocytosis take in large portions of extracellular material, and they are typically not highly selective in the substances they bring in. Cells regulate the endocytosis of specific substances via receptor-mediated endocytosis. Receptor-mediated endocytosis is endocytosis by a portion of the cell membrane that contains many receptors that are specific for a certain substance. Once the surface receptors have bound enough of the specific substance (the receptor’s ligand), the cell will endocytose the part of the cell membrane containing the receptor-ligand complexes. Iron, a required component of haemoglobin, is endocytosed by red blood cells in this way. Iron is bound to a protein called transferrin in the blood. Specific transferrin receptors on red blood cell surfaces bind the iron-transferrin molecules, and the cell endocytoses the receptor-ligand complexes.
In contrast with endocytosis, exocytosis (taking “out of the cell”) is the process of a cell exporting material using vesicular transport (Figure 2.6.10). Many cells manufacture substances that must be secreted, like a factory manufacturing a product for export. These substances are typically packaged into membrane-bound vesicles within the cell. When the vesicle membrane fuses with the cell membrane, the vesicle releases its contents into the interstitial fluid. The vesicle membrane then becomes part of the cell membrane. Cells of the stomach and pancreas produce and secrete digestive enzymes through exocytosis (Figure 2.6.11). Endocrine cells produce and secrete hormones that are sent throughout the body, and certain immune cells produce and secrete large amounts of histamine, a chemical important for immune responses.
Figure 2.6.10. Exocytosis. Exocytosis is much like endocytosis in reverse. Material destined for export is packaged into a vesicle inside the cell. The membrane of the vesicle fuses with the cell membrane, and the contents are released into the extracellular space.
Over 3,000 Australians are living with cystic fibrosis (CF), with an estimated 1 out of every 2,500 babies being born with CF every week. In the United States, approximately 1,000 new cases are reported each year, with an approximate 30,000 people affected by CF. The genetic disease is most well-known for its damage to the lungs, causing breathing difficulties and chronic lung infections, but it also affects the liver, pancreas, and intestines. Only about 50 years ago, the prognosis for children born with CF was very grim—a life expectancy rarely over 10 years. Today, with advances in medical treatment, many CF patients live into their 30s.
The symptoms of CF result from a malfunctioning membrane ion channel called the cystic fibrosis transmembrane conductance regulator, or CFTR. In healthy people, the CFTR protein is an integral membrane protein that transports Cl– ions out of the cell. In a person who has CF, the gene for the CFTR is mutated, thus, the cell manufactures a defective channel protein that typically is not incorporated into the membrane but is instead degraded by the cell.
The CFTR requires ATP in order to function, making its Cl– transport a form of active transport. This characteristic puzzled researchers for a long time because the Cl– ions are actually flowing down their concentration gradient when transported out of cells. Active transport generally pumps ions against their concentration gradient, but the CFTR presents an exception to this rule.
In normal lung tissue, the movement of Cl– out of the cell maintains a Cl–-rich, negatively charged environment immediately outside of the cell. This is particularly important in the epithelial lining of the respiratory system. Respiratory epithelial cells secrete mucus, which serves to trap dust, bacteria, and other debris. A cilium (plural = cilia) is one of the hair-like appendages found on certain cells. Cilia on the epithelial cells move the mucus and its trapped particles up the airways away from the lungs and toward the outside. In order to be effectively moved upward, the mucus cannot be too viscous; rather it must have a thin, watery consistency. The transport of Cl– and the maintenance of an electronegative environment outside of the cell attract positive ions such as Na+ to the extracellular space. The accumulation of both Cl– and Na+ ions in the extracellular space creates solute-rich mucus, which has a low concentration of water molecules. As a result, through osmosis, water moves from cells and extracellular matrix into the mucus, “thinning” it out. This is how, in a normal respiratory system, the mucus is kept sufficiently watered-down to be propelled out of the respiratory system.
If the CFTR channel is absent, Cl– ions are not transported out of the cell in adequate numbers, thus preventing them from drawing positive ions. The absence of ions in the secreted mucus results in the lack of a normal water concentration gradient. Thus, there is no osmotic pressure pulling water into the mucus. The resulting mucus is thick and sticky, and the ciliated epithelia cannot effectively remove it from the respiratory system. Passageways in the lungs become blocked with mucus, along with the debris it carries. Bacterial infections occur more easily because bacterial cells are not effectively carried away from the lungs.
Section Review
The cell membrane provides a barrier around the cell, separating its internal components from the extracellular environment. It is composed of a phospholipid bilayer, with hydrophobic internal lipid “tails” and hydrophilic external phosphate “heads.” Various membrane proteins are scattered throughout the bilayer, both inserted within it and attached to it peripherally. The cell membrane is selectively permeable, allowing only a limited number of materials to diffuse through its lipid bilayer. All materials that cross the membrane do so using passive (non energy-requiring) or active (energy-requiring) transport processes. During passive transport, materials move by simple diffusion or by facilitated diffusion through the membrane, down their concentration gradient. Water passes through the membrane in a diffusion process called osmosis. During active transport, energy is expended to assist material movement across the membrane in a direction against their concentration gradient. Active transport may take place with the help of protein pumps or through the use of vesicles.
Describe the structure and function of the cellular organelles associated with the endomembrane system, including the endoplasmic reticulum, Golgi apparatus, and lysosomes
Describe the structure and function of mitochondria and peroxisomes
Explain the three components of the cytoskeleton, including their composition and functions
Now that you have learned that the cell membrane surrounds all cells, you can dive inside of a prototypical human cell to learn about its internal components and their functions. All living cells in multicellular organisms contain an internal cytoplasmic compartment, and a nucleus within the cytoplasm. Cytosol, the jelly-like substance within the cell, provides the fluid medium necessary for biochemical reactions. Eukaryotic cells, including all animal cells, also contain various cellular organelles. An organelle (“little organ”) is one of several different types of membrane-enclosed bodies in the cell, each performing a unique function. Just as the various bodily organs work together in harmony to perform all of a human’s functions, the many different cellular organelles work together to keep the cell healthy and performing all of its important functions. The organelles and cytosol, taken together, compose the cell’s cytoplasm. The nucleus is a cell’s central organelle, which contains the cell’s DNA (Figure 2.7.1).
Figure 2.7.1. Prototypical human cell. While this image is not indicative of any one particular human cell, it is a prototypical example of a cell containing the primary organelles and internal structures.
Organelles of the Endomembrane System
A set of three major organelles together form a system within the cell called the endomembrane system. These organelles work together to perform various cellular jobs, including the task of producing, packaging, and exporting certain cellular products. The organelles of the endomembrane system include the endoplasmic reticulum, Golgi apparatus, and vesicles.
Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a system of channels that is continuous with the nuclear membrane (or “envelope”) covering the nucleus and composed of the same lipid bilayer material. The ER can be thought of as a series of winding thoroughfares like the waterway canals in Venice. The ER provides passages throughout much of the cell that function in transporting, synthesising, and storing materials. The winding structure of the ER results in a large membranous surface area that supports its many functions (Figure 2.7.2).
Endoplasmic reticulum can exist in two forms: rough ER and smooth ER. These two types of ER perform some very different functions and can be found in very different amounts depending on the type of cell. Rough ER (RER) is so-called because its membrane is dotted with embedded granules—organelles called ribosomes, giving the RER a bumpy appearance. A ribosome is an organelle that serves as the site of protein synthesis. It is composed of two ribosomal RNA subunits that wrap around mRNA to start the process of translation, followed by protein synthesis. Smooth ER (SER) lacks these ribosomes.
One of the main functions of the smooth ER is in the synthesis of lipids. The smooth ER synthesises phospholipids, the main component of biological membranes, as well as steroid hormones. For this reason, cells that produce large quantities of such hormones, such as those of the female ovaries and male testes, contain large amounts of smooth ER. In addition to lipid synthesis, the smooth ER also sequesters (i.e., stores) and regulates the concentration of cellular Ca2+, a function extremely important in cells of the nervous system where Ca2+ is the trigger for neurotransmitter release. The smooth ER additionally metabolises some carbohydrates and performs a detoxification role, breaking down certain toxins.
In contrast with the smooth ER, the primary job of the rough ER is the synthesis and modification of proteins destined for the cell membrane or for export from the cell. For this protein synthesis, many ribosomes attach to the ER (giving it the studded appearance of rough ER). Typically, a protein is synthesised within the ribosome and released inside the channel of the rough ER, where sugars can be added to it (by a process called glycosylation) before it is transported within a vesicle to the next stage in the packaging and shipping process: the Golgi apparatus.
The Golgi Apparatus
The Golgi apparatus is responsible for sorting, modifying, and shipping off the products that come from the rough ER, much like a post-office. The Golgi apparatus looks like stacked flattened discs, almost like stacks of oddly shaped pancakes. Like the ER, these discs are membranous. The Golgi apparatus has two distinct sides, each with a different role. One side of the apparatus receives products in vesicles. These products are sorted through the apparatus, and then they are released from the opposite side after being repackaged into new vesicles. If the product is to be exported from the cell, the vesicle migrates to the cell surface and fuses to the cell membrane, and the cargo is secreted (Figure 2.7.3).
Figure 2.7.3. Golgi apparatus. (a) The Golgi apparatus manipulates products from the rough ER, and also produces new organelles called lysosomes. Proteins and other products of the ER are sent to the Golgi apparatus, which organises, modifies, packages, and tags them. Some of these products are transported to other areas of the cell and some are exported from the cell through exocytosis. Enzymatic proteins are packaged as new lysosomes (or packaged and sent for fusion with existing lysosomes). (b) An electron micrograph of the Golgi apparatus.
Lysosomes
Some of the protein products packaged by the Golgi include digestive enzymes that are meant to remain inside the cell for use in breaking down certain materials. The enzyme-containing vesicles released by the Golgi may form new lysosomes, or fuse with existing, lysosomes. A lysosome is an organelle that contains enzymes that break down and digest unneeded cellular components, such as a damaged organelle. (A lysosome is like a wrecking crew that takes down old and unsound buildings in a neighbourhood.) Autophagy (“self-eating”) is the process of a cell digesting its own structures. Lysosomes are also important for breaking down foreign material, for example, when certain immune defence cells (white blood cells) phagocytose bacteria, the bacterial cell is transported into a lysosome and digested by the enzymes inside. As one might imagine, such phagocytic defence cells contain large numbers of lysosomes.
Under certain circumstances, lysosomes perform a more grand and dire function. In the case of damaged or unhealthy cells, lysosomes can be triggered to open up and release their digestive enzymes into the cytoplasm of the cell, killing the cell. This “self-destruct” mechanism is called autolysis, and makes the process of cell death controlled (a mechanism called “apoptosis”).
Organelles for the Energy Production and Detoxification
In addition to the jobs performed by the endomembrane system, the cell has many other important functions. Just as you must consume nutrients to provide yourself with energy, so must each of your cells take in nutrients, some of which convert to chemical energy that can be used to power biochemical reactions. Another important function of the cell is detoxification. Humans take in all sorts of toxins from the environment and also produce harmful chemicals as by-products of cellular processes. Cells called hepatocytes in the liver detoxify many of these toxins.
Mitochondria
A mitochondrion (plural = mitochondria) is a membranous, bean-shaped organelle that is the “energy transformer” of the cell. Mitochondria consist of an outer lipid bilayer membrane as well as an additional inner lipid bilayer membrane (Figure 2.7.4). The inner membrane is highly folded into winding structures with a great deal of surface area, called cristae. It is along this inner membrane that a series of proteins, enzymes, and other molecules perform the biochemical reactions of cellular respiration. These reactions convert energy stored in nutrient molecules (such as glucose) into adenosine triphosphate (ATP), which provides usable cellular energy to the cell. Cells use ATP constantly, and so the mitochondria are constantly at work. Oxygen molecules are required during cellular respiration, which is why you must constantly breathe it in. One of the organ systems in the body that uses huge amounts of ATP is the muscular system because ATP is required to sustain muscle contraction. As a result, muscle cells are packed full of mitochondria. Nerve cells also need large quantities of ATP to run their sodium-potassium pumps. Therefore, an individual neuron will be loaded with over a thousand mitochondria. On the other hand, a bone cell, which is not nearly as metabolically active, might only have a couple hundred mitochondria.
Like lysosomes, a peroxisome is a membrane-bound cellular organelle that contains mostly enzymes (Figure 2.7.5). Peroxisomes perform a couple of different functions, including lipid metabolism and chemical detoxification. In contrast to the digestive enzymes found in lysosomes, the enzymes within peroxisomes serve to transfer hydrogen atoms from various molecules to oxygen, producing hydrogen peroxide (H2O2). In this way, peroxisomes neutralise poisons such as alcohol. In order to appreciate the importance of peroxisomes, it is necessary to understand the concept of reactive oxygen species.
Figure 2.7.5. Peroxisome. Peroxisomes are membrane-bound organelles that contain an abundance of enzymes for detoxifying harmful substances and lipid metabolism.
Reactive oxygen species (ROS) such as peroxides and free radicals are the highly reactive products of many normal cellular processes, including the mitochondrial reactions that produce ATP and oxygen metabolism. Examples of ROS include the hydroxyl radical OH, H2O2, and superoxide (O2−). Some ROS are important for certain cellular functions, such as cell signalling processes and immune responses against foreign substances. Free radicals are reactive because they contain free unpaired electrons; they can easily oxidise other molecules throughout the cell, causing cellular damage and even cell death. Free radicals are thought to play a role in many destructive processes in the body, from cancer to coronary artery disease.
Peroxisomes, on the other hand, oversee reactions that neutralise free radicals. Peroxisomes produce large amounts of the toxic H2O2 in the process, but peroxisomes contain enzymes that convert H2O2 into water and oxygen. These by-products are safely released into the cytoplasm. Like miniature sewage treatment plants, peroxisomes neutralise harmful toxins so that they do not wreak havoc in the cells. The liver is the organ primarily responsible for detoxifying the blood before it travels throughout the body, and liver cells contain an exceptionally high number of peroxisomes.
Defence mechanisms such as detoxification within the peroxisome and certain cellular antioxidants serve to neutralise many of these molecules. Some vitamins and other substances, found primarily in fruits and vegetables, have antioxidant properties. Antioxidants work by being oxidised themselves, halting the destructive reaction cascades initiated by the free radicals. Sometimes though, ROS accumulate beyond the capacity of such defences.
Oxidative stress is the term used to describe damage to cellular components caused by ROS. Due to their characteristic unpaired electrons, ROS can set off chain reactions where they remove electrons from other molecules, which then become oxidised and reactive, and do the same to other molecules, causing a chain reaction. ROS can cause permanent damage to cellular lipids, proteins, carbohydrates, and nucleic acids. Damaged DNA can lead to genetic mutations and even cancer. A mutation is a change in the nucleotide sequence in a gene within a cell’s DNA, potentially altering the protein coded by that gene. Other diseases believed to be triggered or exacerbated by ROS include Alzheimer’s disease, cardiovascular diseases, diabetes, Parkinson’s disease, arthritis, Huntington’s disease and schizophrenia, among many others. It is noteworthy that these diseases are largely age-related. Many scientists believe that oxidative stress is a major contributor to the ageing process.
Ageing and the Cell: The Free Radical Theory
The free radical theory on ageing was originally proposed in the 1950s, and still remains under debate. Generally speaking, the free radical theory of ageing suggests that accumulated cellular damage from oxidative stress contributes to the physiological and anatomical effects of ageing. There are two significantly different versions of this theory: one states that the ageing process itself is a result of oxidative damage, and the other states that oxidative damage causes age-related disease and disorders. The latter version of the theory is more widely accepted than the former. However, many lines of evidence suggest that oxidative damage does contribute to the ageing process. Research has shown that reducing oxidative damage can result in a longer lifespan in certain organisms such as yeast, worms, and fruit flies. Conversely, increasing oxidative damage can shorten the lifespan of mice and worms. Interestingly, a manipulation called calorie-restriction (moderately restricting the caloric intake) has been shown to increase life span in some laboratory animals. It is believed that this increase is at least in part due to a reduction of oxidative stress. However, a long-term study of primates with calorie-restriction showed no increase in their lifespan. A great deal of additional research will be required to better understand the link between reactive oxygen species and ageing.
The Cytoskeleton
Much like the bony skeleton structurally supports the human body, the cytoskeleton helps the cells to maintain their structural integrity. The cytoskeleton is a group of fibrous proteins that provide structural support for cells, but this is only one of the functions of the cytoskeleton. Cytoskeletal components are also critical for cell motility, cell reproduction, and transportation of substances within the cell.
The cytoskeleton forms a complex thread-like network throughout the cell consisting of three different kinds of protein-based filaments: microfilaments, intermediate filaments, and microtubules (Figure 2.7.6). The thickest of the three is the microtubule, a structural filament composed of subunits of a protein called tubulin. Microtubules maintain cell shape and structure, help resist compression of the cell, and play a role in positioning the organelles within the cell. Microtubules also make up two types of cellular appendages important for motion: cilia and flagella. Cilia are found on many cells of the body, including the epithelial cells that line the airways of the respiratory system. Cilia move rhythmically; they beat constantly, moving waste materials such as dust, mucus, and bacteria upward through the airways, away from the lungs and toward the mouth. Beating cilia on cells in the female fallopian tubes move egg cells from the ovary towards the uterus. A flagellum (plural = flagella) is an appendage larger than a cilium and specialised for cell locomotion. The only flagellated cell in humans is the sperm cell that must propel itself towards female egg cells.
Figure 2.7.6. The three components of the cytoskeleton. The cytoskeleton consists of (a) microtubules, (b) microfilaments, and (c) intermediate filaments. The cytoskeleton plays an important role in maintaining cell shape and structure, promoting cellular movement, and aiding cell division.
A very important function of microtubules is to set the paths (somewhat like railroad tracks) along which the genetic material can be pulled (a process requiring ATP) during cell division, so that each new daughter cell receives the appropriate set of chromosomes. Two short, identical microtubule structures called centrioles are found near the nucleus of cells. A centriole can serve as the cellular origin point for microtubules extending outward as cilia or flagella or can assist with the separation of DNA during cell division. Microtubules grow out from the centrioles by adding more tubulin subunits, like adding additional links to a chain.
In contrast with microtubules, the microfilament is a thinner type of cytoskeletal filament (see Figure 2.7.6b). Actin, a protein that forms chains, is the primary component of these microfilaments. Actin fibres, twisted chains of actin filaments, constitute a large component of muscle tissue and, along with the protein myosin, are responsible for muscle contraction. Like microtubules, actin filaments are long chains of single subunits (called actin subunits). In muscle cells, these long actin strands, called thin filaments, are “pulled” by thick filaments of the myosin protein to contract the cell.
Actin also has an important role during cell division. When a cell is about to split in half during cell division, actin filaments work with myosin to create a cleavage furrow that eventually splits the cell down the middle, forming two new cells from the original cell.
The final cytoskeletal filament is the intermediate filament. As its name would suggest, an intermediate filament is a filament intermediate in thickness between the microtubules and microfilaments (see Figure 2.7.6c). Intermediate filaments are made up of long fibrous subunits of a protein called keratin that are wound together like the threads that compose a rope. Intermediate filaments, in concert with the microtubules, are important for maintaining cell shape and structure. Unlike the microtubules, which resist compression, intermediate filaments resist tension—the forces that pull apart cells. There are many cases in which cells are prone to tension, such as when epithelial cells of the skin are compressed, tugging them in different directions. Intermediate filaments help anchor organelles together within a cell and link cells to other cells by forming special cell-to-cell junctions.
Section Review
The internal environmental of a living cell is made up of a fluid, jelly-like substance called cytosol, which consists mainly of water, but also contains various dissolved nutrients and other molecules. The cell contains an array of cellular organelles, each one performing a unique function and helping to maintain the health and activity of the cell. The cytosol and organelles together compose the cell’s cytoplasm. Most organelles are surrounded by a lipid membrane like the cell membrane of the cell. The endoplasmic reticulum (ER), Golgi apparatus, and lysosomes share a functional connectivity and are collectively referred to as the endomembrane system. There are two types of ER: smooth and rough. While the smooth ER performs many functions, including lipid synthesis and ion storage, the rough ER is mainly responsible for protein synthesis using its associated ribosomes. The rough ER sends newly made proteins to the Golgi apparatus where they are modified and packaged for delivery to various locations within or outside of the cell. Some of these protein products are enzymes destined to break down unwanted material and are packaged as lysosomes for use inside the cell.
Cells also contain mitochondria and peroxisomes, which are the organelles responsible for producing the cell’s energy supply and detoxifying certain chemicals, respectively. Biochemical reactions within mitochondria transform energy-carrying molecules into the usable form of cellular energy known as ATP. Peroxisomes contain enzymes that transform harmful substances such as free radicals into oxygen and water. Cells also contain a miniaturised “skeleton” of protein filaments that extend throughout its interior. Three different kinds of filaments compose this cytoskeleton (in order of increasing thickness): microfilaments, intermediate filaments, and microtubules. Each cytoskeletal component performs unique functions as well as provides a supportive framework for the cell.
Describe the structure and features of the nuclear membrane
List the contents of the nucleus
Explain the organisation of the DNA molecule within the nucleus
Describe the process of DNA replication
The nucleus is the largest and most prominent of a cell’s organelles (Figure 2.8.1). The nucleus is generally considered the control centre of the cell because it stores all of the genetic instructions for manufacturing proteins. Interestingly, some cells in the body, such as muscle cells, contain more than one nucleus (Figure 2.8.2), which is known as multinucleated. Other cells, such as mammalian red blood cells (RBCs), do not contain nuclei at all. RBCs eject their nuclei as they mature, making space for the large numbers of haemoglobin molecules that carry oxygen throughout the body (Figure 2.8.3). Without nuclei, the life span of RBCs is short, and so the body must produce new ones constantly.
Figure 2.8.1. The nucleus. The nucleus is the control centre of the cell. The nucleus of living cells contains the genetic material that determines the entire structure and function of that cell.
Inside the nucleus lies the blueprint that dictates everything a cell will do and all the products it will make. This information is stored within DNA. The nucleus sends “commands” to the cell via molecular messengers that translate the information from DNA. Each cell in your body (with the exception of germ cells) contains the complete set of your DNA. When a cell divides, the DNA must be duplicated so that each new cell receives a full complement of DNA. The following section will explore the structure of the nucleus and its contents, as well as the process of DNA replication.
Organisation of the Nucleus and its DNA
Like most other cellular organelles, the nucleus is surrounded by a membrane called the nuclear envelope. This membranous covering consists of two adjacent lipid bilayers with a thin fluid space in between them. Spanning these two bilayers are nuclear pores. A nuclear pore is a tiny passageway for the passage of proteins, RNA, and solutes between the nucleus and the cytoplasm. Proteins called pore complexes lining the nuclear pores regulate the passage of materials into and out of the nucleus.
Inside the nuclear envelope is a gel-like nucleoplasm with solutes that include the building blocks of nucleic acids. There also can be a dark-staining mass often visible under a simple light microscope, called a nucleolus (plural = nucleoli). The nucleolus is a region of the nucleus that is responsible for manufacturing the RNA necessary for construction of ribosomes. Once synthesised, newly made ribosomal subunits exit the cell’s nucleus through the nuclear pores.
The genetic instructions that are used to build and maintain an organism are arranged in an orderly manner in strands of DNA. Within the nucleus are threads of chromatin composed of DNA and associated proteins (Figure 2.8.4). Along the chromatin threads, the DNA is wrapped around a set of histone proteins. A nucleosome is a single, wrapped DNA-histone complex. Multiple nucleosomes along the entire molecule of DNA appear like a beaded necklace, in which the string is the DNA and the beads are the associated histones. When a cell is in the process of division, the chromatin condenses into chromosomes, so that the DNA can be safely transported to the “daughter cells.” The chromosome is composed of DNA and proteins; it is the condensed form of chromatin. It is estimated that humans have almost 22,000 genes distributed on 46 chromosomes.
Figure 2.8.4. DNA macrostructure. Strands of DNA are wrapped around supporting histones. These proteins are increasingly bundled and condensed into chromatin, which is packed tightly into chromosomes when the cell is ready to divide.
DNA Replication
In order for an organism to grow, develop, and maintain its health, cells must reproduce themselves by dividing to produce two new daughter cells, each with the full complement of DNA as found in the original cell. Billions of new cells are produced in an adult human every day. Only very few cell types in the body do not divide, including nerve cells, skeletal muscle fibres and cardiac muscle cells. The division time of different cell types varies. Epithelial cells of the skin and gastrointestinal lining, for instance, divide very frequently to replace those that are constantly being rubbed off the surface by friction.
A DNA molecule is made of two strands that “complement” each other in the sense that the molecules that compose the strands fit together and bind to each other, creating a double-stranded molecule that looks much like a long, twisted ladder. Each side rail of the DNA ladder is composed of alternating sugar and phosphate groups (Figure 2.8.5). The two sides of the ladder are not identical, but are complementary. These two backbones are bonded to each other across pairs of protruding bases, each bonded pair forming one “rung,” or cross member. The four DNA bases are adenine (A), thymine (T), cytosine (C), and guanine (G). Because of their shape and charge, the two bases that compose a pair always bond together. Adenine always binds with thymine, and cytosine always binds with guanine. The particular sequence of bases along the DNA molecule determines the genetic code. Therefore, if the two complementary strands of DNA were pulled apart, you could infer the order of the bases in one strand from the bases in the other, complementary strand. For example, if one strand has a region with the sequence AGTGCCT, then the sequence of the complementary strand would be TCACGGA.
Figure 2.8.5. Molecular structure of DNA. The DNA double helix is composed of two complementary strands. The strands are bonded together via their nitrogenous base pairs using hydrogen bonds.
DNA replication is the copying of DNA that occurs before cell division can take place. After a great deal of debate and experimentation, the general method of DNA replication was deduced in 1958 by two scientists in California, Matthew Meselson and Franklin Stahl. This method is illustrated in Figure 2.8.6 and described below.
Figure 2.8.6. DNA replication. DNA replication faithfully duplicates the entire genome of the cell. During DNA replication, a number of different enzymes work together to pull apart the two strands so each strand can be used as a template to synthesise new complementary strands. The two new daughter DNA molecules each contain one pre-existing strand and one newly synthesised strand. Thus, DNA replication is said to be “semiconservative.”
Stage 1: Initiation. The two complementary strands are separated, much like unzipping a zipper. Special enzymes, including helicase, untwist and separate the two strands of DNA.
Stage 2:Elongation. Each strand becomes a template along which a new complementary strand is built. DNA polymerase brings in the correct bases to complement the template strand, synthesising a new strand base by base. A DNA polymerase is an enzyme that adds free nucleotides to the end of a chain of DNA, making a new double strand. This growing strand continues to be built until it has fully complemented the template strand.
Stage 3: Termination. Once the two original strands are bound to their own, finished, complementary strands, DNA replication is stopped and the two new identical DNA molecules are complete.
Each new DNA molecule contains one strand from the original molecule and one newly synthesised strand. The term for this mode of replication is “semiconservative,” because half of the original DNA molecule is conserved in each new DNA molecule. This process continues until the cell’s entire genome, the entire complement of an organism’s DNA, is replicated. As you might imagine, it is very important that DNA replication take place precisely so that new cells in the body contain the exact same genetic material as their parent cells. Mistakes made during DNA replication, such as the accidental addition of an inappropriate nucleotide, have the potential to render a gene dysfunctional or useless. Fortunately, there are mechanisms in place to minimise such mistakes. A DNA proofreading process enlists the help of special enzymes that scan the newly synthesised molecule for mistakes and corrects them. Once the process of DNA replication is complete, the cell is ready to divide. You will explore the process of cell division later in the chapter.
Section Review
The nucleus is the command centre of the cell, containing the genetic instructions for all the materials a cell will make (and thus all of its functions it can perform). The nucleus is encased within a membrane of two interconnected lipid bilayers, side-by-side. This nuclear envelope is studded with protein-lined pores that allow materials to be trafficked into and out of the nucleus. The nucleus contains one or more nucleoli, which serve as sites for ribosome synthesis. The nucleus houses the genetic material of the cell: DNA. DNA is normally found as a loosely contained structure called chromatin within the nucleus, where it is wound up and associated with a variety of histone proteins. When a cell is about to divide, the chromatin coils tightly and condenses to form chromosomes.
There is a pool of cells constantly dividing within your body. The result is billions of new cells being created each day. Before any cell is ready to divide, it must replicate its DNA so that each new daughter cell will receive an exact copy of the organism’s genome. A variety of enzymes are enlisted during DNA replication. These enzymes unwind the DNA molecule, separate the two strands, and assist with the building of complementary strands along each parent strand. The original DNA strands serve as templates from which the nucleotide sequence of the new strands are determined and synthesised. When replication is completed, two identical DNA molecules exist. Each one contains one original strand and one newly synthesised complementary strand.
Explain how the genetic code stored within DNA determines the protein that will form
Describe the process of transcription
Describe the process of translation
Discuss the function of ribosomes
It was mentioned earlier that DNA provides a “blueprint” for the cell structure and physiology. This refers to the fact that DNA contains the information necessary for the cell to build one very important type of molecule: the protein. Most structural components of the cell are made up, at least in part, by proteins and virtually all the functions that a cell carries out are completed with the help of proteins. One of the most important classes of proteins is enzymes, which help speed up necessary biochemical reactions that take place inside the cell. Some of these critical biochemical reactions include building larger molecules from smaller components (such as occurs during DNA replication or synthesis of microtubules) and breaking down larger molecules into smaller components (such as when harvesting chemical energy from nutrient molecules). Whatever the cellular process may be, it is almost sure to involve proteins. Just as the cell’s genome describes its full complement of DNA, a cell’s proteome is its full complement of proteins. Protein synthesis begins with genes. A gene is a functional segment of DNA that provides the genetic information necessary to build a protein. Each particular gene provides the code necessary to construct a particular protein. Gene expression, which transforms the information coded in a gene to a final gene product, ultimately dictates the structure and function of a cell by determining which proteins are made.
The interpretation of genes works in the following way. Recall that proteins are polymers, or chains, of many amino acid building blocks. The sequence of bases in a gene (that is, its sequence of A, T, C, G nucleotides) translates to an amino acid sequence. A triplet is a section of three DNA bases in a row that codes for a specific amino acid. Similar to the way in which the three-letter code d-o-g signals the image of a dog, the three-letter DNA base code signals the use of a particular amino acid. For example, the DNA triplet CAC (cytosine, adenine, and cytosine) specifies the amino acid valine. Therefore, a gene, which is composed of multiple triplets in a unique sequence, provides the code to build an entire protein, with multiple amino acids in the proper sequence (Figure 2.9.1). The mechanism by which cells turn the DNA code into a protein product is a two-step process, with an RNA molecule as the intermediate.
Figure 2.9.1. The genetic code. DNA holds all of the genetic information necessary to build a cell’s proteins. The nucleotide sequence of a gene is ultimately translated into an amino acid sequence of the gene’s corresponding protein.
From DNA to RNA: Transcription
DNA is housed within the nucleus, and protein synthesis takes place in the cytoplasm, thus there must be some sort of intermediate messenger that leaves the nucleus and manages protein synthesis. This intermediate messenger is messenger RNA (mRNA), a single-stranded nucleic acid that carries a copy of the genetic code for a single gene out of the nucleus and into the cytoplasm where it is used to produce proteins.
There are several different types of RNA, each having different functions in the cell. The structure of RNA is similar to DNA with a few small exceptions. For one thing, unlike DNA, most types of RNA, including mRNA, are single-stranded and contain no complementary strand. Second, the ribose sugar in RNA contains an additional oxygen atom compared with DNA. Finally, instead of the base thymine, RNA contains the base uracil. This means that adenine will always pair up with uracil during the protein synthesis process.
Gene expression begins with the process called transcription, which is the synthesis of a strand of mRNA that is complementary to the gene of interest. This process is called transcription because the mRNA is like a transcript, or copy, of the gene’s DNA code. Transcription begins in a fashion somewhat like DNA replication, in that a region of DNA unwinds and the two strands separate, however, only that small portion of the DNA will be split apart. The triplets within the gene on this section of the DNA molecule are used as the template to transcribe the complementary strand of RNA (Figure 2.9.2). A codon is a three-base sequence of mRNA, so-called because they directly encode amino acids. Like DNA replication, there are three stages to transcription: initiation, elongation, and termination.
Figure 2.9.2. Transcription: from DNA to mRNA. In the first of the two stages of making protein from DNA, a gene on the DNA molecule is transcribed into a complementary mRNA molecule.
Stage 1: Initiation. A region at the beginning of the gene called a promoter—a particular sequence of nucleotides—triggers the start of transcription.
Stage 2: Elongation. Transcription starts when RNA polymerase unwinds the DNA segment. One strand, referred to as the coding strand, becomes the template with the genes to be coded. The polymerase then aligns the correct nucleic acid (A, C, G, or U) with its complementary base on the coding strand of DNA. RNA polymerase is an enzyme that adds new nucleotides to a growing strand of RNA. This process builds a strand of mRNA.
Stage 3: Termination. When the polymerase has reached the end of the gene, one of three specific triplets (UAA, UAG, or UGA) codes a “stop” signal, which triggers the enzymes to terminate transcription and release the mRNA transcript.
Before the mRNA molecule leaves the nucleus and proceeds to protein synthesis, it is modified in several ways. For this reason, it is often called a pre-mRNA at this stage. For example, your DNA, and thus complementary mRNA, contains long regions called non-coding regions that do not code for amino acids. Their function is still a mystery, but the process called splicing removes these non-coding regions from the pre-mRNA transcript (Figure 2.9.3). A spliceosome—a structure composed of various proteins and other molecules—attaches to the mRNA and “splices” or cuts out the non-coding regions. The removed segment of the transcript is called an intron. The remaining exons are pasted together. An exon is a segment of RNA that remains after splicing. Interestingly, some introns that are removed from mRNA are not always non-coding. When different coding regions of mRNA are spliced out, different variations of the protein will eventually result, with differences in structure and function. This process results in a much larger variety of possible proteins and protein functions. When the mRNA transcript is ready, it travels out of the nucleus and into the cytoplasm.
Figure 2.9.3. Splicing DNA. In the nucleus, a structure called a spliceosome cuts out introns (noncoding regions) within a pre-mRNA transcript and reconnects the exons.
From RNA to Protein: Translation
Like translating a book from one language into another, the codons on a strand of mRNA must be translated into the amino acid alphabet of proteins. Translation is the process of synthesising a chain of amino acids called a polypeptide. Translation requires two major aids: first, a “translator,” the molecule that will conduct the translation, and second, a substrate on which the mRNA strand is translated into a new protein, like the translator’s “desk.” Both of these requirements are fulfilled by other types of RNA. The substrate on which translation takes place is the ribosome.
Remember that many of a cell’s ribosomes are found associated with the rough ER, and carry out the synthesis of proteins destined for the Golgi apparatus. Ribosomal RNA (rRNA) is a type of RNA that, together with proteins, composes the structure of the ribosome. Ribosomes exist in the cytoplasm as two distinct components, a small and a large subunit. When an mRNA molecule is ready to be translated, the two subunits come together and attach to the mRNA. The ribosome provides a substrate for translation, bringing together and aligning the mRNA molecule with the molecular “translators” that must decipher its code.
The other major requirement for protein synthesis is the translator molecules that physically “read” the mRNA codons. Transfer RNA (tRNA) is a type of RNA that ferries the appropriate corresponding amino acids to the ribosome, and attaches each new amino acid to the last, building the polypeptide chain one-by-one. Thus, tRNA transfers specific amino acids from the cytoplasm to a growing polypeptide. The tRNA molecules must be able to recognise the codons on mRNA and match them with the correct amino acid. The tRNA is modified for this function. On one end of its structure is a binding site for a specific amino acid. On the other end is a base sequence that matches the codon specifying its particular amino acid. This sequence of three bases on the tRNA molecule is called an anticodon. For example, a tRNA responsible for shuttling the amino acid glycine contains a binding site for glycine on one end. On the other end it contains an anticodon that complements the glycine codon (GGA is a codon for glycine, and so the tRNAs anticodon would read CCU). Equipped with its particular cargo and matching anticodon, a tRNA molecule can read its recognised mRNA codon and bring the corresponding amino acid to the growing chain (Figure 2.9.4).
Figure 2.9.4. Translation from RNA to protein. During translation, the mRNA transcript is “read” by a functional complex consisting of the ribosome and tRNA molecules. tRNAs bring the appropriate amino acids in sequence to the growing polypeptide chain by matching their anti-codons with codons on the mRNA strand.
Much like the processes of DNA replication and transcription, translation consists of three main stages: initiation, elongation, and termination. Initiation takes place with the binding of a ribosome to an mRNA transcript. The elongation stage involves the recognition of a tRNA anticodon with the next mRNA codon in the sequence. Once the anticodon and codon sequences are bound (remember, they are complementary base pairs), the tRNA presents its amino acid cargo and the growing polypeptide strand is attached to this next amino acid. This attachment takes place with the assistance of various enzymes and requires energy. The tRNA molecule then releases the mRNA strand, the mRNA strand shifts one codon over in the ribosome, and the next appropriate tRNA arrives with its matching anticodon. This process continues until the final codon on the mRNA is reached which provides a “stop” message that signals termination of translation and triggers the release of the complete, newly synthesised protein. Thus, a gene within the DNA molecule is transcribed into mRNA, which is then translated into a protein product (Figure 2.9.5).
Figure 2.9.5. From DNA to protein: transcription through translation. Transcription within the cell nucleus produces an mRNA molecule, which is modified and then sent into the cytoplasm for translation. The transcript is decoded into a protein with the help of a ribosome and tRNA molecules.
Commonly, an mRNA transcription will be translated simultaneously by several adjacent ribosomes. This increases the efficiency of protein synthesis. A single ribosome might translate an mRNA molecule in approximately one minute; so multiple ribosomes aboard a single transcript could produce multiple times the number of the same protein in the same minute. A polyribosome is a string of ribosomes translating a single mRNA strand.
Section Review
DNA stores the information necessary for instructing the cell to perform all its functions. Cells use the genetic code stored within DNA to build proteins, which ultimately determine the structure and function of the cell. This genetic code lies in the particular sequence of nucleotides that make up each gene along the DNA molecule. To “read” this code, the cell must perform two sequential steps. In the first step, transcription, the DNA code is converted into a RNA code. A molecule of messenger RNA that is complementary to a specific gene is synthesised in a process like DNA replication. The molecule of mRNA provides the code to synthesise a protein. In the process of translation, the mRNA attaches to a ribosome. Next, tRNA molecules shuttle the appropriate amino acids to the ribosome, one-by-one, coded by sequential triplet codons on the mRNA, until the protein is fully synthesised. When completed, the mRNA detaches from the ribosome, and the protein is released. Typically, multiple ribosomes attach to a single mRNA molecule at once such that multiple proteins can be manufactured from the mRNA concurrently.
Describe the implications of losing control over the cell cycle
Describe the stages of mitosis and cytokinesis, in order
So far in this chapter, you have read numerous times of the importance and prevalence of cell division. While there are a few cells in the body that do not undergo cell division (such as gametes, red blood cells, most neurons, and some muscle cells), most somatic cells divide regularly. A somatic cell is a general term for a body cell, and all human cells, except for the cells that produce eggs and sperm (which are referred to as germ cells), are somatic cells. Somatic cells contain two copies of each of their chromosomes (one copy received from each parent). A homologous pair of chromosomes is the two copies of a single chromosome found in each somatic cell. The human is a diploid organism, having 23 homologous pairs of chromosomes in each of the somatic cells. The condition of having pairs of chromosomes is known as diploidy.
Cells in the body replace themselves over the lifetime of a person. For example, the cells lining the gastrointestinal tract must be frequently replaced when constantly “worn off” by the movement of food through the gut. But what triggers a cell to divide, and how does it prepare for and complete cell division? The cell cycle is the sequence of events in the life of the cell from the moment it is created at the end of a previous cycle of cell division until it then divides itself, generating two new cells.
The Cell Cycle
One “turn” or cycle of the cell cycle consists of two general phases: interphase, followed by mitosis and cytokinesis. Interphase is the period of the cell cycle during which the cell is not dividing. The majority of cells are in interphase most of the time. Mitosis is the division of genetic material, during which the cell nucleus breaks down and two new, fully functional, nuclei are formed. Cytokinesis divides the cytoplasm into two distinctive cells.
Interphase
Figure 2.10.1. Cell cycle. The two major phases of the cell cycle include mitosis (cell division), and interphase, when the cell grows and performs all of its normal functions. Interphase is further subdivided into G1, S, and G2 phases.
A cell grows and carries out all normal metabolic functions and processes in a period called G1 (Figure 2.10.1). G1 phase (gap 1 phase) is the first gap, or growth phase in the cell cycle. For cells that will divide again, G1 is followed by replication of the DNA, during the S phase. The S phase (synthesis phase) is period during which a cell replicates its DNA.
After the synthesis phase, the cell proceeds through the G2 phase. The G2 phase is a second gap phase, during which the cell continues to grow and makes the necessary preparations for mitosis. Between G1, S, and G2 phases, cells will vary the most in their duration of the G1 phase. It is here that a cell might spend a couple of hours, or many days. The S phase typically lasts between 8-10 hours and the G2 phase approximately 5 hours. In contrast to these phases, the G0 phase is a resting phase of the cell cycle. Cells that have temporarily stopped dividing and are resting (a common condition) and cells that have permanently ceased dividing (like nerve cells) are said to be in G0.
The Structure of Chromosomes
Billions of cells in the human body divide every day. During the synthesis phase (S, for DNA synthesis) of interphase, the amount of DNA within the cell precisely doubles. Therefore, after DNA replication but before cell division, each cell contains two copies of each chromosome. Each copy of the chromosome is referred to as a sister chromatid and is physically bound to the other copy. The centromere is the structure that attaches one sister chromatid to another. Because a human cell has 46 chromosomes, during this phase, there are 92 chromatids (46 × 2) in the cell. Make sure not to confuse the concept of a pair of chromatids (one chromosome and its exact copy attached during mitosis) and a homologous pair of chromosomes (two paired chromosomes which were inherited separately, one from each parent) (Figure 2.10.2).
Figure 2.10.2. A homologous pair of chromosomes with their attached sister chromatids. The red and blue colours correspond to a homologous pair of chromosomes. Each member of the pair was separately inherited from one parent. Each chromosome in the homologous pair is also bound to an identical sister chromatid, which is produced by DNA replication, and results in the familiar “X” shape.
Mitosis and Cytokinesis
The mitotic phase of the cell typically takes between 1 and 2 hours. During this phase, a cell undergoes two major processes. First, it completes mitosis, during which the contents of the nucleus are equitably pulled apart and distributed between its two halves. Cytokinesis then occurs, dividing the cytoplasm and cell body into two new cells. Mitosis is divided into four major stages that take place after interphase (Figure 2.10.3) and in the following order: prophase, metaphase, anaphase, and telophase. The process is then followed by cytokinesis.
Figure 2.10.3. Cell division: mitosis followed by cytokinesis. The stages of cell division oversee the separation of identical genetic material into two new nuclei, followed by the division of the cytoplasm.
Prophase is the first phase of mitosis, during which the loosely packed chromatin coils and condenses into visible chromosomes. During prophase, each chromosome becomes visible with its identical partner attached, forming the familiar X-shape of sister chromatids. The nucleolus disappears early during this phase, and the nuclear envelope also disintegrates.
A major occurrence during prophase concerns a very important structure that contains the origin site for microtubule growth. Recall the cellular structures called centrioles that serve as origin points from which microtubules extend. These tiny structures also play a very important role during mitosis. A centrosome is a pair of centrioles together. The cell contains two centrosomes side-by-side, which begin to move apart during prophase. As the centrosomes migrate to two different sides of the cell, microtubules begin to extend from each like long fingers from two hands extending toward each other. The mitotic spindle is the structure composed of the centrosomes and their emerging microtubules.
Near the end of prophase there is an invasion of the nuclear area by microtubules from the mitotic spindle. The nuclear membrane has disintegrated, and the microtubules attach themselves to the centromeres that adjoin pairs of sister chromatids. The kinetochore is a protein structure on the centromere that is the point of attachment between the mitotic spindle and the sister chromatids. This stage is referred to as late prophase or “prometaphase” to indicate the transition between prophase and metaphase.
Metaphase is the second stage of mitosis. During this stage, the sister chromatids, with their attached microtubules, line up along a linear plane in the middle of the cell. A metaphase plate forms between the centrosomes that are now located at either end of the cell. The metaphase plate is the name for the plane through the centre of the spindle on which the sister chromatids are positioned. The microtubules are now poised to pull apart the sister chromatids and bring one from each pair to each side of the cell.
Anaphase is the third stage of mitosis. Anaphase takes place over a few minutes, when the pairs of sister chromatids are separated from one another, forming individual chromosomes once again. These chromosomes are pulled to opposite ends of the cell by their kinetochores, as the microtubules shorten. Each end of the cell receives one partner from each pair of sister chromatids, ensuring that the two new daughter cells will contain identical genetic material.
Telophase is the final stage of mitosis. Telophase is characterised by the formation of two new daughter nuclei at either end of the dividing cell. These newly formed nuclei surround the genetic material, which uncoils such that the chromosomes return to loosely packed chromatin. Nucleoli also reappear within the new nuclei, and the mitotic spindle breaks apart, each new cell receiving its own complement of DNA, organelles, membranes, and centrioles. At this point, the cell is already beginning to split in half as cytokinesis begins.
The cleavage furrow is a contractile band made up of microfilaments that forms around the midline of the cell during cytokinesis. (Recall that microfilaments consist of actin.) This contractile band squeezes the two cells apart until they finally separate. Two new cells are now formed. One of these cells (the “stem cell”) enters its own cell cycle; able to grow and divide again at some future time. The other cell transforms into the functional cell of the tissue, typically replacing an “old” cell there.
Imagine a cell that completed mitosis but never underwent cytokinesis. In some cases, a cell may divide its genetic material and grow in size, but fail to undergo cytokinesis. This results in larger cells with more than one nucleus. Usually this is an unwanted aberration and can be a sign of cancerous cells.
Cell Cycle Control
A very elaborate and precise system of regulation controls direct the way cells proceed from one phase to the next in the cell cycle and begin mitosis. The control system involves molecules within the cell as well as external triggers. These internal and external control triggers provide “stop” and “advance” signals for the cell. Precise regulation of the cell cycle is critical for maintaining the health of an organism, and loss of cell cycle control can lead to cancer.
Mechanisms of Cell Cycle Control
As the cell proceeds through its cycle, each phase involves certain processes that must be completed before the cell should advance to the next phase. A checkpoint is a point in the cell cycle at which the cycle can be signalled to move forward or stopped. At each of these checkpoints, different varieties of molecules provide the stop or go signals, depending on certain conditions within the cell. A cyclin is one of the primary classes of cell cycle control molecules (Figure 2.10.4). A cyclin-dependent kinase (CDK) is one of a group of molecules that work together with cyclins to determine progression past cell checkpoints. By interacting with many additional molecules, these triggers push the cell cycle forward unless prevented from doing so by “stop” signals, if for some reason the cell is not ready. At the G1 checkpoint, the cell must be ready for DNA synthesis to occur. At the G2 checkpoint the cell must be fully prepared for mitosis. Even during mitosis, a crucial stop and go checkpoint in metaphase ensures that the cell is fully prepared to complete cell division. The metaphase checkpoint ensures that all sister chromatids are properly attached to their respective microtubules and lined up at the metaphase plate before the signal is given to separate them during anaphase.
Figure 2.10.4. Control of the cell cycle. Cells proceed through the cell cycle under the control of a variety of molecules, such as cyclins and cyclin-dependent kinases. These control molecules determine whether or not the cell is prepared to move into the following stage.
The Cell Cycle Out of Control: Implications
Most people understand that cancer or tumours are caused by abnormal cells that multiply continuously. If the abnormal cells continue to divide unstopped, they can damage the tissues around them, spread to other parts of the body, and eventually result in death. In healthy cells, the tight regulation mechanisms of the cell cycle prevent this from happening, while failures of cell cycle control can cause unwanted and excessive cell division. Failures of control may be caused by inherited genetic abnormalities that compromise the function of certain “stop” and “go” signals. Environmental insult that damages DNA can also cause dysfunction in those signals. Often, a combination of both genetic predisposition and environmental factors lead to cancer.
The process of a cell escaping its normal control system and becoming cancerous may actually happen throughout the body quite frequently. Fortunately, certain cells of the immune system are capable of recognising cells that have become cancerous and destroying them. However, in certain cases the cancerous cells remain undetected and continue to proliferate. If the resulting tumour does not pose a threat to surrounding tissues, it is said to be benign and can usually be easily removed. If capable of damage, the tumour is considered malignant and the patient is diagnosed with cancer.
Homeostatic Imbalances
Cancer Arises from Homeostatic Imbalances
Cancer is an extremely complex condition, capable of arising from a wide variety of genetic and environmental causes. Typically, mutations or aberrations in a cell’s DNA that compromise normal cell cycle control systems lead to cancerous tumours. Cell cycle control is an example of a homeostatic mechanism that maintains proper cell function and health. While progressing through the phases of the cell cycle, a large variety of intracellular molecules provide stop and go signals to regulate movement forward to the next phase. These signals are maintained in an intricate balance so that the cell only proceeds to the next phase when it is ready. This homeostatic control of the cell cycle can be thought of like a car’s cruise control. Cruise control will continually apply just the right amount of acceleration to maintain a desired speed, unless the driver hits the brakes, in which case the car will slow down. Similarly, the cell includes molecular messengers, such as cyclins, that push the cell forward in its cycle.
In addition to cyclins, a class of proteins that are encoded by genes called proto-oncogenes provide important signals that regulate the cell cycle and move it forward. Examples of proto-oncogene products include cell-surface receptors for growth factors, or cell-signalling molecules, two classes of molecules that can promote DNA replication and cell division. In contrast, a second class of genes known as tumours suppressor genes sends stop signals during a cell cycle. For example, certain protein products of tumours suppressor genes signal potential problems with the DNA and thus stop the cell from dividing, while other proteins signal the cell to die if it is damaged beyond repair. Some tumours suppressor proteins also signal a sufficient surrounding cellular density, which indicates that the cell need not presently divide. The latter function is uniquely important in preventing tumours growth: normal cells exhibit a phenomenon called “contact inhibition;” thus, extensive cellular contact with neighbouring cells causes a signal that stops further cell division.
These two contrasting classes of genes, proto-oncogenes and tumour suppressor genes, are like the accelerator and brake pedal of the cell’s own “cruise control system,” respectively. Under normal conditions, these stop and go signals are maintained in a homeostatic balance. Generally speaking, there are two ways that the cell’s cruise control can lose control: a malfunctioning (overactive) accelerator, or a malfunctioning (underactive) brake. When compromised through a mutation, or otherwise altered, proto-oncogenes can be converted to oncogenes, which produce oncoproteins that push a cell forward in its cycle and stimulate cell division even when it is undesirable to do so. For example, a cell that should be programmed to self-destruct (a process called apoptosis) due to extensive DNA damage might instead be triggered to proliferate by an oncoprotein. On the other hand, a dysfunctional tumours suppressor gene may fail to provide the cell with a necessary stop signal, also resulting in unwanted cell division and proliferation.
A delicate homeostatic balance between the many proto-oncogenes and tumours suppressor genes delicately controls the cell cycle and ensures that only healthy cells replicate. Therefore, a disruption of this homeostatic balance can cause aberrant cell division and cancerous growths.
Section Review
The life of cell consists of stages that make up the cell cycle. After a cell is ‘born’, it passes through an interphase before it is ready to replicate itself and produce daughter cells. This interphase includes two gap phases (G1 and G2), as well as an S phase, during which its DNA is replicated in preparation for cell division. The cell cycle is under precise regulation by chemical messengers both inside and outside the cell that provide “stop” and “go” signals for movement from one phase to the next. Failures of these signals can result in cells that continue to divide uncontrollably, which can lead to cancer.
Once a cell has completed interphase and is ready for cell division, it proceeds through four separate stages of mitosis (prophase, metaphase, anaphase, and telophase). Telophase is followed by the division of the cytoplasm (cytokinesis), which generates two daughter cells. This process takes place in all normally dividing cells of the body except for the germ cells that produce eggs and sperm.
Discuss how the generalised cells of a developing embryo or the stem cells of an adult organism become differentiated into specialised cells
Distinguish between the categories of stem cells
How does a complex organism such as a human develop from a single cell—a fertilised egg—into the vast array of cell types such as nerve cells, muscle cells, and epithelial cells that characterise the adult? Throughout development and adulthood, the process of cellular differentiation leads cells to assume their final morphology and physiology. Differentiation is the process by which unspecialised cells become specialised to carry out distinct functions.
Stem Cells
A stem cell is an unspecialised cell that can divide without limit as needed and can, under specific conditions, differentiate into specialised cells. Stem cells are divided into several categories according to their potential to differentiate.
The first embryonic cells that arise from the division of the zygote are the ultimate stem cells; these stems cells are described as totipotent because they have the potential to differentiate into any of the cells needed to enable an organism to grow and develop.
The embryonic cells that develop from totipotent stem cells and are precursors to the fundamental tissue layers of the embryo are classified as pluripotent. A pluripotent stem cell is one that has the potential to differentiate into any type of human tissue but cannot support the full development of an organism. These cells then become slightly more specialised and are referred to as multipotent cells.
A multipotent stem cell has the potential to differentiate into different types of cells within a given cell lineage or small number of lineages, such as a red blood cell or white blood cell.
Finally, multipotent cells can become further specialised oligopotent cells. An oligopotent stem cell is limited to becoming one of a few different cell types. In contrast, a unipotent cell is fully specialised and can only reproduce to generate more of its own specific cell type.
Stem cells are unique in that they can also continually divide and regenerate new stem cells instead of further specialising. There are different stem cells present at different stages of a human’s life. They include the embryonic stem cells of the embryo, foetal stem cells of the foetus, and adult stem cells in the adult. One type of adult stem cell is the epithelial stem cell, which gives rise to the keratinocytes in the multiple layers of epithelial cells in the epidermis of skin. Adult bone marrow has three distinct types of stem cells: hematopoietic stem cells, which give rise to red blood cells, white blood cells and platelets (Figure 2.11.1); endothelial stem cells, which give rise to the endothelial cell types that line blood and lymph vessels; and mesenchymal stem cells, which give rise to the different types of muscle cells.
Figure 2.11.1. Haematopoiesis. The process of haematopoiesis involves the differentiation of multipotent cells into blood and immune cells. The multipotent hematopoietic stem cells give rise to many different cell types, including the cells of the immune system and red blood cells.
Differentiation
When a cell differentiates (becomes more specialised), it may undertake major changes in its size, shape, metabolic activity, and overall function. Because all cells in the body, beginning with the fertilised egg, contain the same DNA, how do the different cell types come to be so different? The answer is analogous to a movie script. The different actors in a movie all read from the same script, however, they are each only reading their own part of the script. Similarly, all cells contain the same full complement of DNA, but each type of cell only “reads” the portions of DNA that are relevant to its own function. In biology, this is referred to as the unique genetic expression of each cell.
In order for a cell to differentiate into its specialised form and function, it need only manipulate those genes (and thus those proteins) that will be expressed, and not those that will remain silent. The primary mechanism by which genes are turned “on” or “off” is through transcription factors. A transcription factor is one of a class of proteins that bind to specific genes on the DNA molecule and either promote or inhibit their transcription (Figure 2.11.2).
Figure 2.11.2. Transcription factors regulate gene expression. While each body cell contains the organism’s entire genome, different cells regulate gene expression with the use of various transcription factors. Transcription factors are proteins that affect the binding of RNA polymerase to a particular gene on the DNA molecule.
Everyday Connection
Stem Cell Research
Stem cell research aims to find ways to use stem cells to regenerate and repair cellular damage. Over time, most adult cells undergo the wear and tear of ageing and lose their ability to divide and repair themselves. Stem cells do not display a particular morphology or function. Adult stem cells, which exist as a small subset of cells in most tissues, keep dividing and can differentiate into a number of specialised cells generally formed by that tissue. These cells enable the body to renew and repair body tissues.
The mechanisms that induce a non-differentiated cell to become a specialised cell are poorly understood. In a laboratory setting, it is possible to induce stem cells to differentiate into specialised cells by changing the physical and chemical conditions of growth. Several sources of stem cells are used experimentally and are classified according to their origin and potential for differentiation. Human embryonic stem cells (hESCs) are extracted from embryos and are pluripotent. The adult stem cells that are present in many organs and differentiated tissues, such as bone marrow and skin, are multipotent, being limited in differentiation to the types of cells found in those tissues. The stem cells isolated from umbilical cord blood are also multipotent, as are cells from deciduous teeth (baby teeth). Researchers have recently developed induced pluripotent stem cells (iPSCs) from mouse and human adult stem cells. These cells are genetically reprogrammed multipotent adult cells that function like embryonic stem cells; they are capable of generating cells characteristic of all three germ layers.
Because of their capacity to divide and differentiate into specialised cells, stem cells offer a potential treatment for diseases such as diabetes and heart disease (Figure 2.11.3). Cell-based therapy refers to treatment in which stem cells induced to differentiate in a growth dish are injected into a patient to repair damaged or destroyed cells or tissues. Many obstacles must be overcome for the application of cell-based therapy. Although embryonic stem cells have a nearly unlimited range of differentiation potential, they are seen as foreign by the patient’s immune system and may trigger rejection. Also, the destruction of embryos to isolate embryonic stem cells raises considerable ethical and legal questions.
Figure 2.11.3. Stem cells. The capacity of stem cells to differentiate into specialised cells make them potentially valuable in therapeutic applications designed to replace damaged cells of different body tissues.
In contrast, adult stem cells isolated from a patient are not seen as foreign by the body, but they have a limited range of differentiation. Some individuals bank the cord blood or deciduous teeth of their child, storing away those sources of stem cells for future use, should their child need it. Induced pluripotent stem cells are considered a promising advance in the field because using them avoids the legal, ethical and immunological pitfalls of embryonic stem cells.
Section Review
One of the major areas of research in biology is that of how cells specialise to assume their unique structures and functions, since all cells essentially originate from a single fertilised egg. Cell differentiation is the process of cells becoming specialised as they body develops. A stem cell is an unspecialised cell that can divide without limit as needed and can, under specific conditions, differentiate into specialised cells. Stem cells are divided into several categories according to their potential to differentiate. While all somatic cells contain the exact same genome, different cell types only express some of those genes at any given time. These differences in gene expression ultimately dictate a cell’s unique morphological and physiological characteristics. The primary mechanism that determines which genes will be expressed and which ones will not is through the use of different transcription factor proteins, which bind to DNA and promote or hinder the transcription of different genes. Through the action of these transcription factors, cells specialise into one of hundreds of different cell types in the human body.
Relate the structure of each tissue type to their function
Discuss the embryonic origin of tissue
Identify the three major germ layers
Identify the main types of tissue membranes
The term tissue is used to describe a group of cells found together in the body. The cells within a tissue share a common embryonic origin. Microscopic observation reveals that the cells in a tissue share morphological features and are arranged in an orderly pattern that achieves the tissue’s functions. From the evolutionary perspective, tissues appear in more complex organisms. For example, multicellular protists, ancient eukaryotes, do not have cells organised into tissues.
Although there are many types of cells in the human body, they are organised into four broad categories of tissues: epithelial, connective, muscle, and nervous. Each of these categories is characterised by specific functions that contribute to the overall health and maintenance of the body. A disruption of the structure is a sign of injury or disease. Such changes can be detected through histology, the microscopic study of tissue appearance, organisation, and function.
The Four Types of Tissues
Epithelial tissue, also referred to as epithelium, refers to the sheets of cells that cover exterior surfaces of the body, lines internal cavities and passageways, and forms certain glands. Connective tissue, as its name implies, binds the cells and organs of the body together and functions in the protection, support, and integration of all parts of the body. Muscle tissue is excitable, responding to stimulation and contracting to provide movement, and occurs as three major types: skeletal (voluntary) muscle, smooth muscle, and cardiac muscle in the heart. Nervous tissue is also excitable, allowing the propagation of electrochemical signals in the form of nerve impulses that communicate between different regions of the body (Figure 3.1.1).
The next level of organisation is the organ, where several types of tissues come together to form a working unit. Just as knowing the structure and function of cells helps you in your study of tissues, knowledge of tissues will help you understand how organs function. The epithelial and connective tissues are discussed in detail in this chapter. Muscle and nervous tissues will be discussed only briefly in this chapter.
The zygote, or fertilised egg, is a single cell formed by the fusion of an egg and sperm. After fertilisation the zygote gives rise to rapid mitotic cycles, generating many cells to form the embryo. The first embryonic cells generated can differentiate into any type of cell in the body and, as such, are called totipotent, meaning each has the capacity to divide, differentiate, and develop into a new organism. As cell proliferation progresses, three major cell lineages are established within the embryo. Each of these lineages of embryonic cells forms the distinct germ layers from which all the tissues and organs of the human body eventually form. Each germ layer is identified by its relative position: ectoderm (ecto- = “outer”), mesoderm (meso- = “middle”), and endoderm (endo- = “inner”). Figure 3.1.2 shows the types of tissues and organs associated with the each of the three germ layers. Note that epithelial tissue originates in all three layers, whereas nervous tissue derives primarily from the ectoderm and muscle tissue from mesoderm.
Figure 3.1.2. Embryonic origin of tissues and major organs.
Tissue Membranes
A tissue membrane is a thin layer or sheet of cells that covers the outside of the body (for example, skin), the organs (for example, pericardium), internal passageways that lead to the exterior of the body (for example, abdominal mesenteries), and the lining of the moveable joint cavities. There are two basic types of tissue membranes: connective tissue and epithelial membranes (Figure 3.1.3).
Figure 3.1.3. Tissue membranes. The two broad categories of tissue membranes in the body are (1) connective tissue membranes, which include synovial membranes, and (2) epithelial membranes, which include mucous membranes, serous membranes, and the cutaneous membrane, in other words, the skin.
Connective Tissue Membranes
The connective tissue membrane is formed solely from connective tissue. These membranes encapsulate organs, such as the kidneys, and line our movable joints. A synovial membrane is a type of connective tissue membrane that lines the cavity of a freely movable joint. For example, synovial membranes surround the joints of the shoulder, elbow, and knee. Fibroblasts in the inner layer of the synovial membrane release hyaluronan into the joint cavity. The hyaluronan effectively traps available water to form the synovial fluid, a natural lubricant that enables the bones of a joint to move freely against one another without much friction. This synovial fluid readily exchanges water and nutrients with blood, as do all body fluids.
Epithelial Membranes
The epithelial membrane is composed of epithelium attached to a layer of connective tissue, for example, your skin. The mucous membrane is also a composite of connective and epithelial tissues. Sometimes called mucosae, these epithelial membranes line the body cavities and hollow passageways that open to the external environment, and include the digestive, respiratory, excretory, and reproductive tracts. Mucous, produced by the epithelial exocrine glands, covers the epithelial layer. The underlying connective tissue, called the lamina propria (literally “own layer”), help support the fragile epithelial layer.
A serous membrane is an epithelial membrane composed of mesodermally derived epithelium called the mesothelium that is supported by connective tissue. These membranes line the coelomic cavities of the body, that is, those cavities that do not open to the outside, and they cover the organs located within those cavities. They are essentially membranous bags, with mesothelium lining the inside and connective tissue on the outside. Serous fluid secreted by the cells of the thin squamous mesothelium lubricates the membrane and reduces abrasion and friction between organs. Serous membranes are identified according locations. Three serous membranes line the thoracic cavity; the two pleura that cover the lungs and the pericardium that covers the heart. A fourth, the peritoneum, is the serous membrane in the abdominal cavity that covers abdominal organs and forms double sheets of mesenteries that suspend many of the digestive organs.
The skin is an epithelial membrane also called the cutaneous membrane. It is a stratified squamous epithelial membrane resting on top of connective tissue. The apical surface of this membrane is exposed to the external environment and is covered with dead, keratinised cells that help protect the body from desiccation and pathogens.
Section Review
The human body contains more than 200 types of cells that can all be classified into four types of tissues: epithelial, connective, muscle, and nervous. Epithelial tissues act as coverings controlling the movement of materials across the surface. Connective tissue integrates the various parts of the body and provides support and protection to organs. Muscle tissue allows the body to move. Nervous tissues propagate information.
The study of the shape and arrangement of cells in tissue is called histology. All cells and tissues in the body derive from three germ layers in the embryo: the ectoderm, mesoderm, and endoderm.
Different types of tissues form membranes that enclose organs, provide a friction-free interaction between organs, and keep organs together. Synovial membranes are connective tissue membranes that protect and line the joints. Epithelial membranes are formed from epithelial tissue attached to a layer of connective tissue. There are three types of epithelial membranes: mucous, which contain glands; serous, which secrete fluid; and cutaneous which makes up the skin.
Explain the structure and function of epithelial tissue
Distinguish between tight junctions, anchoring junctions, and gap junctions
Distinguish between simple epithelia and stratified epithelia, as well as between squamous, cuboidal, and columnar epithelia
Describe the structure and function of endocrine and exocrine glands and their respective secretions
Most epithelial tissues are essentially large sheets of cells covering all the surfaces of the body exposed to the outside world and lining the outside of organs. Epithelium also forms much of the glandular tissue of the body. Skin is not the only area of the body exposed to the outside. Other areas include the airways, the digestive tract, as well as the urinary and reproductive systems, all of which are lined by an epithelium. Hollow organs and body cavities that do not connect to the exterior of the body, which includes, blood vessels and serous membranes, are lined by endothelium (plural = endothelia), which is a type of epithelium.
Epithelial cells derive from all three major embryonic layers. The epithelia lining the skin, parts of the mouth and nose, and the anus develop from the ectoderm. Cells lining the airways and most of the digestive system originate in the endoderm. The epithelium that lines vessels in the lymphatic and cardiovascular system derives from the mesoderm and is called an endothelium.
All epithelia share some important structural and functional features. This tissue is highly cellular, with little or no extracellular material present between cells. Adjoining cells form a specialised intercellular connection between their cell membranes called a cell junction. The epithelial cells exhibit polarity with differences in structure and function between the exposed or apical facing surface of the cell and the basal surface close to the underlying body structures. The basal lamina, a mixture of glycoproteins and collagen, provides an attachment site for the epithelium, separating it from underlying connective tissue. The basal lamina attaches to a reticular lamina, which is secreted by the underlying connective tissue, forming a basement membrane that helps hold it all together.
Epithelial tissues are nearly completely avascular. For instance, no blood vessels cross the basement membrane to enter the tissue, and nutrients must come by diffusion or absorption from underlying tissues or the surface. Many epithelial tissues are capable of rapidly replacing damaged and dead cells. Sloughing off damaged or dead cells is a characteristic of surface epithelium and allows our airways and digestive tracts to rapidly replace damaged cells with new cells.
Generalised Functions of Epithelial Tissue
Epithelial tissues provide the body’s first line of protection from physical, chemical, and biological wear and tear. The cells of an epithelium act as gatekeepers of the body controlling permeability and allowing selective transfer of materials across a physical barrier. All substances that enter the body must cross an epithelium. Some epithelia often include structural features that allow the selective transport of molecules and ions across their cell membranes.
Many epithelial cells are capable of secretion and release mucous and specific chemical compounds onto their apical surfaces. The epithelium of the small intestine releases digestive enzymes, for example. Cells lining the respiratory tract secrete mucous that traps incoming microorganisms and particles. A glandular epithelium contains many secretory cells.
The Epithelial Cell
Epithelial cells are typically characterised by the polarised distribution of organelles and membrane-bound proteins between their basal and apical surfaces. Particular structures found in some epithelial cells are an adaptation to specific functions. Certain organelles are segregated to the basal sides, whereas other organelles and extensions, such as cilia, when present, are on the apical surface.
Cilia are microscopic extensions of the apical cell membrane that are supported by microtubules. They beat in unison and move fluids as well as trapped particles. Ciliated epithelium lines the ventricles of the brain where it helps circulate the cerebrospinal fluid. The ciliated epithelium of your airway forms a mucociliary escalator that sweeps particles of dust and pathogens trapped in the secreted mucous toward the throat. It is called an escalator because it continuously pushes mucous with trapped particles upward. In contrast, nasal cilia sweep the mucous blanket down towards your throat. In both cases, the transported materials are usually swallowed, and end up in the acidic environment of your stomach.
Cell to Cell Junctions
Cells of epithelia are closely connected and are not separated by intracellular material. Three basic types of connections allow varying degrees of interaction between the cells: tight junctions, anchoring junctions, and gap junctions (Figure 3.2.1).
Figure 3.2.1. Types of cell junctions. The three basic types of cell-to-cell junctions are tight junctions, gap junctions, and anchoring junctions.
At one end of the spectrum is the tight junction, which separates the cells into apical and basal compartments. An anchoring junction includes several types of cell junctions that help stabilise epithelial tissues. Anchoring junctions are common on the lateral and basal surfaces of cells where they provide strong and flexible connections. There are three types of anchoring junctions: desmosomes, hemidesmosomes, and adherens. Desmosomes occur in patches on the membranes of cells. The patches are structural proteins on the inner surface of the cell’s membrane. The adhesion molecule, cadherin, is embedded in these patches and projects through the cell membrane to link with the cadherin molecules of adjacent cells. These connections are especially important in holding cells together. Hemidesmosomes, which look like half a desmosome, link cells to the extracellular matrix, for example, the basal lamina. While similar in appearance to desmosomes, they include the adhesion proteins called integrins rather than cadherins. Adherens junctions use either cadherins or integrins depending on whether they are linking to other cells or matrix. The junctions are characterised by the presence of the contractile protein actin located on the cytoplasmic surface of the cell membrane. The actin can connect isolated patches or form a belt-like structure inside the cell. These junctions influence the shape and folding of the epithelial tissue.
In contrast with the tight and anchoring junctions, a gap junction forms an intercellular passageway between the membranes of adjacent cells to facilitate the movement of small molecules and ions between the cytoplasm of adjacent cells. These junctions allow electrical and metabolic coupling of adjacent cells, which coordinates function in large groups of cells.
Classification of Epithelial Tissues
Epithelial tissues are classified according to the shape of the cells and number of the cell layers formed (Figure 3.2.2). Cell shapes can be squamous (flattened and thin), cuboidal (boxy, as wide as it is tall), or columnar (rectangular, taller than it is wide). Similarly, the number of cell layers in the tissue can be one—where every cell rests on the basal lamina—which is a simple epithelium, or more than one, which is a stratified epithelium and only the basal layer of cells rests on the basal lamina. Pseudostratified (pseudo- = “false”) describes tissue with a single layer of irregularly shaped cells that give the appearance of more than one layer. Transitional describes a form of specialised stratified epithelium in which the shape of the cells can vary.
Figure 3.2.2. Cells of epithelial tissue. Simple epithelial tissue is organised as a single layer of cells and stratified epithelial tissue is formed by several layers of cells.
Simple Epithelium
The shape of the cells in the single cell layer of simple epithelium reflects the functioning of those cells. The cells in simple squamous epithelium have the appearance of thin scales. Squamous cell nuclei tend to be flat, horizontal, and elliptical, mirroring the form of the cell. The endothelium is the epithelial tissue that lines vessels of the lymphatic and cardiovascular system, and it is made up of a single layer of squamous cells. Simple squamous epithelium, because of the thinness of the cell, is present where rapid passage of chemical compounds is observed. The alveoli of lungs where gases diffuse, segments of kidney tubules, and the lining of capillaries are also made of simple squamous epithelial tissue. The mesothelium is a simple squamous epithelium that forms the surface layer of the serous membrane that lines body cavities and internal organs. Its primary function is to provide a smooth and protective surface. Mesothelial cells are squamous epithelial cells that secrete a fluid that lubricates the mesothelium.
In simple cuboidal epithelium, the nucleus of the box-like cells appears round and is generally located near the centre of the cell. These epithelia are active in the secretion and absorptions of molecules. Simple cuboidal epithelia are observed in the lining of the kidney tubules and in the ducts of glands.
In simple columnar epithelium, the nucleus of the tall column-like cells tends to be elongated and located in the basal end of the cells. Like the cuboidal epithelia, this epithelium is active in the absorption and secretion of molecules. Simple columnar epithelium forms the lining of some sections of the digestive system and parts of the female reproductive tract. Ciliated columnar epithelium is composed of simple columnar epithelial cells with cilia on their apical surfaces. These epithelial cells are found in the lining of the fallopian tubes and parts of the respiratory system, where the beating of the cilia helps remove particulate matter.
Pseudostratified columnar epithelium is a type of epithelium that appears to be stratified but instead consists of a single layer of irregularly shaped and differently sized columnar cells. In pseudostratified epithelium, nuclei of neighbouring cells appear at different levels rather than clustered in the basal end. The arrangement gives the appearance of stratification; but in fact, all the cells are in contact with the basal lamina, although some do not reach the apical surface. Pseudostratified columnar epithelium is found in the respiratory tract, where some of these cells have cilia.
Both simple and pseudostratified columnar epithelia are heterogeneous epithelia because they include additional types of cells interspersed among the epithelial cells. For example, a goblet cell is a mucous-secreting unicellular “gland” interspersed between the columnar epithelial cells of mucous membranes (Figure 3.2.3).
A stratified epithelium consists of several stacked layers of cells. This epithelium protects against physical and chemical wear and tear. The stratified epithelium is named by the shape of the most apical layer of cells, closest to the free space. Stratified squamous epithelium is the most common type of stratified epithelium in the human body. The apical cells are squamous, whereas the basal layer contains either columnar or cuboidal cells. The top layer may be covered with dead cells filled with keratin. Mammalian skin is an example of this dry, keratinised, stratified squamous epithelium. The lining of the mouth cavity is an example of an unkeratinised, stratified squamous epithelium. Stratified cuboidal epithelium and stratified columnar epithelium can also be found in certain glands and ducts but are uncommon in the human body.
Another kind of stratified epithelium is transitional epithelium, so-called because of the gradual changes in the shapes of the apical cells as the bladder fills with urine. It is found only in the urinary system, specifically the ureters and urinary bladder. When the bladder is empty, this epithelium is convoluted and has cuboidal apical cells with convex, umbrella shaped, apical surfaces. As the bladder fills with urine, this epithelium loses its convolutions and the apical cells transition from cuboidal to squamous. It appears thicker and more multi-layered when the bladder is empty, and more stretched out and less stratified when the bladder is full and distended. Figure 3.2.4 summarises the different categories of epithelial cell tissue cells.
Figure 3.2.4. Summary of epithelial tissue cells.
Glandular Epithelium
A gland is a structure made up of one or more cells modified to synthesise and secrete chemical substances. Most glands consist of groups of epithelial cells. A gland can be classified as an endocrine gland, a ductless gland that releases secretions directly into surrounding tissues and fluids (endo- = “inside”), or an exocrine gland whose secretions leave through a duct that opens directly, or indirectly, to the external environment (exo- = “outside”).
Endocrine Glands
The secretions of endocrine glands are called hormones. Hormones are released into the interstitial fluid, diffused into the bloodstream, and delivered to targets, in other words, cells that have receptors to bind the hormones. The endocrine system is part of a major regulatory system coordinating the regulation and integration of body responses. A few examples of endocrine glands include the anterior pituitary, thymus, adrenal cortex, and gonads.
Exocrine Glands
Exocrine glands release their contents through a duct that leads to the epithelial surface. Mucous, sweat, saliva, and breast milk are all examples of secretions from exocrine glands. They are all discharged through tubular ducts. Secretions into the lumen of the gastrointestinal tract, technically outside of the body, are of the exocrine category.
Glandular Structure
Exocrine glands are classified as either unicellular or multicellular. The unicellular glands are scattered single cells, such as goblet cells, found in the mucous membranes of the small and large intestine.
The multicellular exocrine glands known as serous glands develop from simple epithelium to form a secretory surface that secretes directly into an inner cavity. These glands line the internal cavities of the abdomen and chest and release their secretions directly into the cavities. Other multicellular exocrine glands release their contents through a tubular duct. The duct is single in a simple gland but in compound glands is divided into one or more branches (Figure 3.2.5). In tubular glands, the ducts can be straight or coiled, whereas tubes that form pockets are alveolar (acinar), such as the exocrine portion of the pancreas. Combinations of tubes and pockets are known as tubuloalveolar (tubuloacinar) compound glands. In a branched gland, a duct is connected to more than one secretory group of cells.
Figure 3.2.5. Types of exocrine glands. Exocrine glands are classified by their structure.
Methods and Types of Secretion
Exocrine glands can be classified by their mode of secretion and the nature of the substances released, as well as by the structure of the glands and shape of ducts (Figure 3.2.6). Merocrine secretion is the most common type of exocrine secretion. The secretions are enclosed in vesicles that move to the apical surface of the cell where the contents are released by exocytosis. For example, watery mucous containing the glycoprotein mucin, a lubricant that offers some pathogen protection is a merocrine secretion. The eccrine glands that produce and secrete sweat are another example.
Figure 3.2.6. Modes of glandular secretion. (a) In merocrine secretion, the cell remains intact. (b) In apocrine secretion, the apical portion of the cell is released, as well. (c) In holocrine secretion, the cell is destroyed as it releases its product and the cell itself becomes part of the secretion.
Apocrine secretion accumulates near the apical portion of the cell. That portion of the cell and its secretory contents pinch off from the cell and are released. The sweat glands of the armpit are classified as apocrine glands. Both merocrine and apocrine glands continue to produce and secrete their contents with little damage caused to the cell because the nucleus and Golgi regions remain intact after secretion.
In contrast, the process of holocrine secretion involves the rupture and destruction of the entire gland cell. The cell accumulates its secretory products and releases them only when it bursts. New gland cells differentiate from cells in the surrounding tissue to replace those lost by secretion. The sebaceous glands that produce the oils on the skin and hair are holocrine glands/cells (Figure 3.2.7).
Glands are also named after the products they produce. The serous gland produces watery, blood-plasma-like secretions rich in enzymes such as alpha amylase, whereas the mucous gland releases watery to viscous products rich in the glycoprotein mucin. Both serous and mucous glands are common in the salivary glands of the mouth. Mixed exocrine glands contain both serous and mucous glands and release both types of secretions.
Section Review
In epithelial tissue, cells are closely packed with little or no extracellular matrix except for the basal lamina that separates the epithelium from underlying tissue. The main functions of epithelia are protection from the environment, coverage, secretion and excretion, absorption, and filtration. Cells are bound together by tight junctions that form an impermeable barrier. They can also be connected by gap junctions, which allow free exchange of soluble molecules between cells, and anchoring junctions, which attach cell to cell or cell to matrix. The different types of epithelial tissues are characterised by their cellular shapes and arrangements: squamous, cuboidal, or columnar epithelia. Single cell layers form simple epithelia, whereas stacked cells form stratified epithelia. Very few capillaries penetrate these tissues.
Glands are secretory tissues and organs that are derived from epithelial tissues. Exocrine glands release their products through ducts. Endocrine glands secrete hormones directly into the interstitial fluid and blood stream. Glands are classified both according to the type of secretion and by their structure. Merocrine glands secrete products as they are synthesised. Apocrine glands release secretions by pinching off the apical portion of the cell, whereas holocrine gland cells store their secretions until they rupture and release their contents. In this case, the cell becomes part of the secretion.
Identify and distinguish between the types of connective tissue: proper, supportive, and fluid
Explain the functions of connective tissues
As may be obvious from its name, one of the major functions of connective tissue is to connect tissues and organs. Unlike epithelial tissue, which is composed of cells closely packed with little or no extracellular space in between, connective tissue cells are dispersed in a matrix. The matrix usually includes a large amount of extracellular material produced by the connective tissue cells that are embedded within it. The matrix plays a major role in the functioning of this tissue. The major component of the matrix is a ground substance often crisscrossed by protein fibres. This ground substance is usually a fluid, but it can also be mineralised and solid, as in bones. Connective tissues come in a vast variety of forms, yet they typically have in common three characteristic components: cells, large amounts of amorphous ground substance, and protein fibres. The amount and structure of each component correlates with the function of the tissue, from the rigid ground substance in bones supporting the body to the inclusion of specialised cells; for example, a phagocytic cell that engulfs pathogens and rids tissue of cellular debris.
Functions of Connective Tissues
Connective tissues perform many functions in the body, but most importantly, they support and connect other tissues; from the connective tissue sheath that surrounds muscle cells, to the tendons that attach muscles to bones, and to the skeleton that supports the positions of the body. Protection is another major function of connective tissue, in the form of fibrous capsules and bones that protect delicate organs and, of course, the skeletal system. Specialised cells in connective tissue defend the body from microorganisms that enter the body. Transport of fluid, nutrients, waste, and chemical messengers is ensured by specialised fluid connective tissues, such as blood and lymph. Adipose cells store surplus energy in the form of fat and contribute to the thermal insulation of the body.
Embryonic Connective Tissue
All connective tissues derive from the mesodermal layer of the embryo. The first connective tissue to develop in the embryo is mesenchyme, the stem cell line from which all connective tissues are later derived. Clusters of mesenchymal cells are scattered throughout adult tissue and supply the cells needed for replacement and repair after a connective tissue injury. A second type of embryonic connective tissue forms in the umbilical cord, called mucous connective tissue or Wharton’s jelly. This tissue is no longer present after birth, leaving only scattered mesenchymal cells throughout the body.
Classification of Connective Tissues
The three broad categories of connective tissue are classified according to the characteristics of their ground substance and the types of fibres found within the matrix (Table 3.3.1). Connective tissue proper includes loose connective tissue and dense connective tissue. Both tissues have a variety of cell types and protein fibres suspended in a viscous ground substance. Dense connective tissue is reinforced by bundles of fibres that provide tensile strength, elasticity, and protection. In loose connective tissue, the fibres are loosely organised, leaving large spaces in between. Supportive connective tissue—bone and cartilage—provide structure and strength to the body and protect soft tissues. A few distinct cell types and densely packed fibres in a matrix characterise these tissues. In bone, the matrix is rigid and described as calcified because of the deposited calcium salts. In fluid connective tissue, in other words, lymph and blood, various specialised cells circulate in a watery fluid containing salts, nutrients, and dissolved proteins.
Table 3.3.1. Connective Tissue Examples
Connective Tissue Proper
Supportive Connective Tissue
Fluid Connective Tissue
Loose connective tissue
Cartilage
Blood
• Areolar
• Hyaline
• Adipose
• Fibrocartilage
• Reticular
• Elastic
Dense connective tissue
Bones
Lymph
• Regular elastic
• Compact bone
• Irregular elastic
• Cancellous bone
Connective Tissue Proper
Fibroblasts are present in all connective tissue proper (Figure 3.3.1). Fibrocytes, adipocytes, and mesenchymal cells are fixed cells, which means they remain within the connective tissue. Other cells move in and out of the connective tissue in response to chemical signals. Macrophages, mast cells, lymphocytes, plasma cells, and phagocytic cells are found in connective tissue proper but are actually part of the immune system protecting the body.
The most abundant cell in connective tissue proper is the fibroblast. Polysaccharides and proteins secreted by fibroblasts combine with extra-cellular fluids to produce a viscous ground substance that, with embedded fibrous proteins, forms the extra-cellular matrix. As you might expect, a fibrocyte, a less active form of fibroblast, is the second most common cell type in connective tissue proper.
Adipocytes are cells that store lipids as droplets that fill most of the cytoplasm. There are two basic types of adipocytes: white and brown. The brown adipocytes store lipids as many droplets, and have high metabolic activity. In contrast, white fat adipocytes store lipids as a single large drop and are metabolically less active. Their effectiveness at storing large amounts of fat is witnessed in obese individuals. The number and type of adipocytes depends on the tissue and location, and vary among individuals in the population.
The mesenchymal cell is a multipotent adult stem cell. These cells can differentiate into any type of connective tissue cells needed for repair and healing of damaged tissue.
The macrophage cell is a large cell derived from a monocyte, a type of blood cell, which enters the connective tissue matrix from the blood vessels. The macrophage cells are an essential component of the immune system, which is the body’s defence against potential pathogens and degraded host cells. When stimulated, macrophages release cytokines, small proteins that act as chemical messengers. Cytokines recruit other cells of the immune system to infected sites and stimulate their activities. Roaming, or free, macrophages move rapidly by amoeboid movement, engulfing infectious agents and cellular debris. In contrast, fixed macrophages are permanent residents of their tissues.
The mast cell, found in connective tissue proper, has many cytoplasmic granules. These granules contain the chemical signals histamine and heparin. When irritated or damaged, mast cells release histamine, an inflammatory mediator, which causes vasodilation and increased blood flow at a site of injury or infection, along with itching, swelling, and redness you recognise as an allergic response. Like blood cells, mast cells are derived from hematopoietic stem cells and are part of the immune system.
Connective Tissue Fibres and Ground Substance
Three main types of fibres are secreted by fibroblasts: collagen fibres, elastic fibres, and reticular fibres. Collagen fibres is made from fibrous protein subunits linked together to form a long and straight fibres. Collagen fibres, while flexible, have great tensile strength, resist stretching, and give ligaments and tendons their characteristic resilience and strength. These fibres hold connective tissues together, even during the movement of the body.
Elastic fibres contain the protein elastin along with lesser amounts of other proteins and glycoproteins. The main property of elastin is that after being stretched or compressed, it will return to its original shape. Elastic fibres are prominent in elastic tissues found in skin and the elastic ligaments of the vertebral column.
Reticular fibres are also formed from the same protein subunits as collagen fibres; however, these fibres remain narrow and are arrayed in a branching network. They are found throughout the body, but are most abundant in the reticular tissue of soft organs, such as liver and spleen, where they anchor and provide structural support to the parenchyma (the functional cells, blood vessels, and nerves of the organ).
All these fibres types are embedded in ground substance. Secreted by fibroblasts, ground substance is made of polysaccharides, specifically hyaluronic acid, and proteins. These combine to form a proteoglycan with a protein core and polysaccharide branches. The proteoglycan attracts and traps available moisture forming the clear, viscous, colourless matrix you now know as ground substance.
Loose Connective Tissue
Loose connective tissue is found between many organs where it acts both to absorb shock and bind tissues together. It allows water, salts, and various nutrients to diffuse through to adjacent or imbedded cells and tissues.
Adipose tissue consists mostly of fat storage cells, with little extracellular matrix (Figure 3.3.2). Many capillaries allow rapid storage and mobilisation of lipid molecules. White adipose tissue is most abundant. It can appear yellow and owes its colour to carotene and related pigments from plant food. White fat contributes mostly to lipid storage and can serve as insulation from cold temperatures and mechanical injuries. White adipose tissue can be found protecting the kidneys and cushioning the back of the eye. Brown adipose tissue is more common in infants, hence the term “baby fat.” In adults, there is a reduced amount of brown fat and it is found mainly in the neck and clavicular regions of the body. The many mitochondria in the cytoplasm of brown adipose tissue help explain its efficiency at metabolising stored fat. Brown adipose tissue is thermogenic, meaning that as it breaks down fats, it releases metabolic heat, rather than producing adenosine triphosphate (ATP), a key molecule used in metabolism.
Areolar tissue shows little specialisation. It contains all the cell types and fibres previously described and is distributed in a random, web-like fashion. It fills the spaces between muscle fibres, surrounds blood and lymph vessels, and supports organs in the abdominal cavity. Areolar tissue underlies most epithelia and represents the connective tissue component of epithelial membranes, which are described further in a later section.
Reticular tissue is a mesh-like, supportive framework for soft organs such as lymphatic tissue, the spleen, and the liver (Figure 3.3.3). Reticular cells produce the reticular fibres that form the network onto which other cells attach. It derives its name from the Latin reticulus, which means “little net.”
Dense connective tissue contains more collagen fibres than does loose connective tissue. Consequently, it displays greater resistance to stretching. There are two major categories of dense connective tissue: regular and irregular. Dense regular connective tissue fibres are parallel to each other, enhancing tensile strength and resistance to stretching in the direction of the fibre’s orientation. Ligaments and tendons are made of dense regular connective tissue, but in ligaments not all fibres are parallel. Dense regular elastic tissue contains elastin fibres in addition to collagen fibres, which allows the ligament to return to its original length after stretching. The ligaments in the vocal folds and between the vertebrae in the vertebral column are elastic.
In dense irregular connective tissue, the direction of fibres is random. This arrangement gives the tissue greater strength in all directions and less strength in one particular direction. In some tissues, fibres crisscross and form a mesh. In other tissues, stretching in several directions is achieved by alternating layers where fibres run in the same orientation in each layer, and it is the layers themselves that are stacked at an angle. The dermis of the skin is an example of dense irregular connective tissue rich in collagen fibres. Dense irregular elastic tissues give arterial walls the strength and the ability to regain original shape after stretching (Figure 3.3.4).
Your opponent stands ready as you prepare to hit the serve, but you are confident that you will smash the ball past your opponent. As you toss the ball high in the air, a burning pain shoots across your wrist and you drop the tennis racket. That dull ache in the wrist that you ignored through the summer is now an unbearable pain. The game is over for now.
After examining your swollen wrist, the doctor in the emergency room announces that you have developed wrist tendinitis. She recommends icing the tender area, taking non-steroidal anti-inflammatory medication to ease the pain and to reduce swelling, and complete rest for a few weeks. She interrupts your protests that you cannot stop playing. She issues a stern warning about the risk of aggravating the condition and the possibility of surgery. She consoles you by mentioning that well known tennis players such as Venus and Serena Williams and Rafael Nadal have also suffered from tendinitis related injuries.
What is tendinitis and how did it happen? Tendinitis is the inflammation of a tendon, the thick band of fibrous connective tissue that attaches a muscle to a bone. The condition causes pain and tenderness in the area around a joint. On rare occasions, a sudden serious injury will cause tendinitis. Most often, the condition results from repetitive motions over time that strain the tendons needed to perform the tasks.
Persons whose jobs and hobbies involve performing the same movements over and over again are often at the greatest risk of tendinitis. You hear of tennis and golfer’s elbow, jumper’s knee, and swimmer’s shoulder. In all cases, overuse of the joint causes a microtrauma that initiates the inflammatory response. Tendinitis is routinely diagnosed through a clinical examination. In case of severe pain, X-rays can be examined to rule out the possibility of a bone injury. Severe cases of tendinitis can even tear loose a tendon. Surgical repair of a tendon is painful. Connective tissue in the tendon does not have abundant blood supply and heals slowly.
While older adults are at risk for tendinitis because the elasticity of tendon tissue decreases with age, active people of all ages can develop tendinitis. Young athletes, dancers, and computer operators; anyone who performs the same movements constantly is at risk for tendinitis. Although repetitive motions are unavoidable in many activities and may lead to tendinitis, precautions can be taken that can lessen the probability of developing tendinitis. For active individuals, stretches before exercising and cross training or changing exercises are recommended. For the passionate athlete, it may be time to take some lessons to improve technique. All of the preventive measures aim to increase the strength of the tendon and decrease the stress put on it. With proper rest and managed care, you will be back on the court to hit that slice-spin serve over the net.
Supportive Connective Tissues
Two major forms of supportive connective tissue, cartilage and bone, allow the body to maintain its posture and protect internal organs.
Cartilage
The distinctive appearance of cartilage is due to polysaccharides called chondroitin sulphates, which bind with ground substance proteins to form proteoglycans. Embedded within the cartilage matrix are chondrocytes, or cartilage cells, and the space they occupy are called lacunae (singular = lacuna). A layer of dense irregular connective tissue, the perichondrium, encapsulates the cartilage. Cartilaginous tissue is avascular, thus all nutrients need to diffuse through the matrix to reach the chondrocytes. This is a factor contributing to the very slow healing of cartilaginous tissues.
The three main types of cartilage tissue are hyaline cartilage, fibrocartilage, and elastic cartilage (Figure 3.3.5). Hyaline cartilage, the most common type of cartilage in the body, consists of short and dispersed collagen fibres and contains large amounts of proteoglycans. Under the microscope, tissue samples appear clear. The surface of hyaline cartilage is smooth. Both strong and flexible, it is found in the rib cage and nose and covers bones where they meet to form moveable joints. It makes up a template of the embryonic skeleton before bone formation. A plate of hyaline cartilage at the ends of bone allows continued growth until adulthood. Fibrocartilage is tough because it has thick bundles of collagen fibres dispersed through its matrix. The knee and jaw joints and the intervertebral discs are examples of fibrocartilage. Elastic cartilage contains elastic fibres as well as collagen and proteoglycans. This tissue gives rigid support as well as elasticity. Tug gently at your ear lobes and notice that the lobes return to their initial shape. The external ear contains elastic cartilage.
Bone is the hardest connective tissue. It provides protection to internal organs and supports the body. Bone’s rigid extracellular matrix contains mostly collagen fibres embedded in a mineralised ground substance containing hydroxyapatite, a form of calcium phosphate. Both components of the matrix, organic and inorganic, contribute to the unusual properties of bone. Without collagen, bones would be brittle and shatter easily. Without mineral crystals, bones would flex and provide little support. Osteocytes, bone cells like chondrocytes, are located within lacunae. The histology of transverse tissue from long bone shows a typical arrangement of osteocytes in concentric circles around a central canal. Bone is a highly vascularised tissue. Unlike cartilage, bone tissue can recover from injuries in a relatively short time.
Cancellous bone looks like a sponge under the microscope and contains empty spaces between trabeculae, or arches of bone proper. It is lighter than compact bone and found in the interior of some bones and at the end of long bones. Compact bone is solid and has greater structural strength.
Fluid Connective Tissue
Blood and lymph are fluid connective tissues. Cells circulate in a liquid extracellular matrix. The formed elements circulating in blood are all derived from hematopoietic stem cells located in bone marrow (Figure 3.3.6). Erythrocytes, red blood cells, transport oxygen and some carbon dioxide. Leukocytes, white blood cells, are responsible for defending against potentially harmful microorganisms or molecules. Platelets are cell fragments involved in blood clotting. Some white blood cells can cross the endothelial layer that lines blood vessels and enter adjacent tissues. Nutrients, salts, and wastes are dissolved in the liquid matrix and transported through the body.
Lymph contains a liquid matrix and white blood cells. Lymphatic capillaries are extremely permeable, allowing larger molecules and excess fluid from interstitial spaces to enter the lymphatic vessels. Lymph drains into blood vessels, delivering molecules to the blood that could not otherwise directly enter the bloodstream. In this way, specialised lymphatic capillaries transport absorbed fats away from the intestine and deliver these molecules to the blood.
Connective tissue is a heterogeneous tissue with many cell shapes and tissue architecture. Structurally, all connective tissues contain cells that are embedded in an extracellular matrix stabilised by proteins. The chemical nature and physical layout of the extracellular matrix and proteins vary enormously among tissues, reflecting the variety of functions that connective tissue fulfills in the body. Connective tissues separate and cushion organs, protecting them from shifting or traumatic injury. Connect tissues provide support and assist movement, store and transport energy molecules, protect against infections, and contribute to temperature homeostasis.
Many different cells contribute to the formation of connective tissues. They originate in the mesodermal germ layer and differentiate from mesenchyme and hematopoietic tissue in the bone marrow. Fibroblasts are the most abundant and secrete many protein fibres, adipocytes specialise in fat storage, hematopoietic cells from the bone marrow give rise to all the blood cells, chondrocytes form cartilage, and osteocytes form bone. The extracellular matrix contains fluid, proteins, polysaccharide derivatives, and, in the case of bone, mineral crystals. Protein fibres fall into three major groups: collagen fibres that are thick, strong, flexible, and resist stretch; reticular fibres that are thin and form a supportive mesh; and elastin fibres that are thin and elastic.
The major types of connective tissue are connective tissue proper, supportive tissue, and fluid tissue. Loose connective tissue proper includes adipose tissue, areolar tissue, and reticular tissue. These serve to hold organs and other tissues in place and, in the case of adipose tissue, isolate and store energy reserves. The matrix is the most abundant feature for loose tissue although adipose tissue does not have much extracellular matrix. Dense connective tissue proper is richer in fibres and may be regular, with fibres oriented in parallel as in ligaments and tendons, or irregular, with fibres oriented in several directions. Organ capsules (collagenous type) and walls of arteries (elastic type) contain dense irregular connective tissue. Cartilage and bone are supportive tissue. Cartilage contains chondrocytes and is somewhat flexible. Hyaline cartilage is smooth and clear, covers joints, and is found in the growing portion of bones. Fibrocartilage is tough because of extra collagen fibres and forms, among other things, the intervertebral discs. Elastic cartilage can stretch and recoil to its original shape because of its high content of elastic fibres. The matrix contains very few blood vessels. Bones are made of a rigid, mineralised matrix containing calcium salts, crystals, and osteocytes lodged in lacunae. Bone tissue is highly vascularised. Cancellous bone is spongy and less solid than compact bone. Fluid tissue, for example blood and lymph, is characterised by a liquid matrix and no supporting fibres.
Compare and contrast the functions of each muscle tissue type
Explain how muscle tissue can enable motion
Muscle tissue is characterised by properties that allow movement. Muscle cells are excitable; they respond to a stimulus. They are contractile, meaning they can shorten and generate a pulling force. When attached between two movable objects, in other words, bones, contractions of the muscles cause the bones to move. Some muscle movement is voluntary, which means it is under conscious control. For example, a person decides to open a book and read a chapter on anatomy. Other movements are involuntary, meaning they are not under conscious control, such as the contraction of your pupil in bright light. Muscle tissue is classified into three types according to structure and function: skeletal, cardiac, and smooth (Table 3.4.1).
Table 3.4.1. Comparison of structure and properties of muscle tissue types
Tissue
Histology
Function
Location
Skeletal
Long cylindrical fibre, striated, many peripherally located nuclei
Attached to bones and around entrance points to body (e.g., mouth, anus)
Cardiac
Short, branched, striated, single central nucleus
Contracts to pump blood
Heart
Smooth
Short, spindle-shaped, no evident striation, single nucleus in each fibre
Involuntary movement, moves food, involuntary control of respiration, moves secretions, regulates flow of blood in arteries by contraction
Walls of major organs and passage ways
Skeletal muscle is attached to bones and its contraction makes possible locomotion, facial expressions, posture, and other voluntary movements of the body. Forty percent of your body mass is made up of skeletal muscle. Skeletal muscles generate heat as a by-product of their contraction and thus participate in thermal homeostasis. Shivering is an involuntary contraction of skeletal muscles in response to perceived lower than normal body temperature. The muscle cell, or myocyte, develops from myoblasts derived from the mesoderm. Myocytes and their numbers remain relatively constant throughout life. Skeletal muscle tissue is arranged in bundles surrounded by connective tissue. Under the light microscope, muscle cells appear striated with many nuclei squeezed along the membranes. The striation is due to the regular alternation of the contractile proteins, actin and myosin, along with the structural proteins that couple the contractile proteins to connective tissues. The cells are multinucleated as a result of the fusion of the many myoblasts that fuse to form each long muscle fibre.
Cardiac muscle forms the contractile walls of the heart. The cells of cardiac muscle, known as cardiomyocytes, also appear striated under the microscope. Unlike skeletal muscle fibres, cardiomyocytes are single cells typically with a single centrally located nucleus. A principal characteristic of cardiomyocytes is that they contract on their own intrinsic rhythms without any external stimulation. Cardiomyocyte attach to one another with specialised cell junctions called intercalated discs. Intercalated discs have both anchoring junctions and gap junctions. Attached cells form long, branching cardiac muscle fibres that are, essentially, a mechanical and electrochemical syncytium allowing the cells to synchronise their actions. The cardiac muscle pumps blood through the body and is under involuntary control. The attachment junctions hold adjacent cells together across the dynamic pressures changes of the cardiac cycle.
Smooth muscle tissue contraction is responsible for involuntary movements in the internal organs. It forms the contractile component of the digestive, urinary, and reproductive systems as well as the airways and arteries. Each cell is spindle shaped with a single nucleus and no visible striations (Figure 3.4.1).
The three types of muscle cells are skeletal, cardiac, and smooth. Their morphologies match their specific functions in the body. Skeletal muscle is voluntary and responds to conscious stimuli. The cells are striated and multinucleated appearing as long, unbranched cylinders. Cardiac muscle is involuntary and found only in the heart. Each cell is striated with a single nucleus and they attach to one another to form long fibres. Cells are attached to one another at intercalated disks. The cells are interconnected physically and electrochemically to act as a syncytium. Cardiac muscle cells contract autonomously and involuntarily. Smooth muscle is involuntary. Each cell is a spindle-shaped fibre and contains a single nucleus. No striations are evident because the actin and myosin filaments do not align in the cytoplasm.
3.5 Nervous Tissue Mediates Perception and Response
Learning Objectives
By the end of this section, you will be able to:
Identify the classes of cells that make up nervous tissue
Discuss how nervous tissue mediates perception and response
Nervous tissue is characterised as being excitable and capable of sending and receiving electrochemical signals that provide the body with information. Two main classes of cells make up nervous tissue: the neuron and neuroglia (Figure 3.5.1). Neurons propagate information via electrochemical impulses, called action potentials, which are biochemically linked to the release of chemical signals. Neuroglia play an essential role in supporting neurons and modulating their information propagation.
Neurons display distinctive morphology, well suited to their role as conducting cells, with three main parts. The cell body includes most of the cytoplasm, the organelles, and the nucleus. Dendrites branch off the cell body and appear as thin extensions. A long “tail,” the axon, extends from the neuron body and can be wrapped in an insulating layer known as myelin, which is formed by accessory cells. The synapse is the gap between nerve cells, or between a nerve cell and its target, for example, a muscle or a gland, across which the impulse is transmitted by chemical compounds known as neurotransmitters. Neurons categorised as multipolar neurons have several dendrites and a single prominent axon. Bipolar neurons possess a single dendrite and axon with the cell body, while unipolar neurons have only a single process extending out from the cell body, which divides into a functional dendrite and into a functional axon. When a neuron is sufficiently stimulated, it generates an action potential that propagates down the axon towards the synapse. If enough neurotransmitters are released at the synapse to stimulate the next neuron or target, a response is generated.
The second class of neural cells comprises the neuroglia or glial cells, which have been characterised as having a simple support role. The word “glia” comes from the Greek word for glue. Recent research is shedding light on the more complex role of neuroglia in the function of the brain and nervous system. Astrocyte cells, named for their distinctive star shape, are abundant in the central nervous system. The astrocytes have many functions, including regulation of ion concentration in the intercellular space, uptake and/or breakdown of some neurotransmitters, and formation of the blood-brain barrier, the membrane that separates the circulatory system from the brain. Microglia protect the nervous system against infection but are not nervous tissue because they are related to macrophages. Oligodendrocyte cells produce myelin in the central nervous system (brain and spinal cord) while the Schwann cell produces myelin in the peripheral nervous system (Figure 3.5.2).
The most prominent cell of the nervous tissue, the neuron, is characterised mainly by its ability to receive stimuli and respond by generating an electrical signal, known as an action potential, which can travel rapidly over great distances in the body. A typical neuron displays a distinctive morphology: a large cell body branches out into short extensions called dendrites, which receive chemical signals from other neurons, and a long tail called an axon, which relays signals away from the cell to other neurons, muscles, or glands. Many axons are wrapped by a myelin sheath, a lipid derivative that acts as an insulator and speeds up the transmission of the action potential. Other cells in the nervous tissue, the neuroglia, include the astrocytes, microglia, oligodendrocytes, and Schwann cells.
Discuss the progressive impact of ageing on tissue
Describe cancerous mutations’ effect on tissue
Tissues of all types are vulnerable to injury and, inevitably, ageing. In the former case, understanding how tissues respond to damage can guide strategies to aid repair. In the latter case, understanding the impact of ageing can help in the search for ways to diminish its effects.
Tissue Injury and Repair
Inflammation is the standard, initial response of the body to injury. Whether biological, chemical, physical, or radiation burns, all injuries lead to the same sequence of physiological events. Inflammation limits the extent of injury, partially or fully eliminates the cause of injury, and initiates repair and regeneration of damaged tissue. Necrosis, or accidental cell death, causes inflammation. Apoptosis is programmed cell death, a normal step-by-step process that destroys cells no longer needed by the body. By mechanisms still under investigation, apoptosis does not initiate the inflammatory response. Acute inflammation resolves over time by the healing of tissue. If inflammation persists, it becomes chronic and leads to diseased conditions. Arthritis and tuberculosis are examples of chronic inflammation. The suffix “-itis” denotes inflammation of a specific organ or type, for example, peritonitis is the inflammation of the peritoneum, and meningitis refers to the inflammation of the meninges, the tough membranes that surround the central nervous system
The four cardinal signs of inflammation—redness, swelling, pain and local heat—were first recorded in antiquity. Cornelius Celsus is credited with documenting these signs during the days of the Roman Empire, as early as the first century AD. A fifth sign, loss of function, may also accompany inflammation.
Upon tissue injury, damaged cells release inflammatory chemical signals that evoke local vasodilation, the widening of the blood vessels. Increased blood flow results in apparent redness and heat. In response to injury, mast cells present in tissue degranulate, releasing the potent vasodilator histamine. Increased blood flow and inflammatory mediators recruit white blood cells to the site of inflammation. The endothelium lining the local blood vessel becomes “leaky” under the influence of histamine and other inflammatory mediators allowing neutrophils, macrophages, and fluid to move from the blood into the interstitial tissue spaces. The excess liquid in tissue causes swelling, more properly called oedema. The swollen tissues squeezing pain receptors cause the sensation of pain. Prostaglandins released from injured cells also activate pain neurons. Non-steroidal anti-inflammatory drugs (NSAIDs) reduce pain because they inhibit the synthesis of prostaglandins. High levels of NSAIDs reduce inflammation. Antihistamines decrease allergies by blocking histamine receptors and as a result the histamine response.
After containment of an injury, the tissue repair phase starts with removal of toxins and waste products. Clotting (coagulation) reduces blood loss from damaged blood vessels and forms a network of fibrin proteins that trap blood cells and bind the edges of the wound together. A scab forms when the clot dries, reducing the risk of infection. Sometimes a mixture of dead leukocytes and fluid called pus accumulates in the wound. As healing progresses, fibroblasts from the surrounding connective tissues replace the collagen and extracellular material lost by the injury. Angiogenesis, the growth of new blood vessels, results in vascularisation of the new tissue known as granulation tissue. The clot retracts pulling the edges of the wound together, and it slowly dissolves as the tissue is repaired. When a large amount of granulation tissue forms and capillaries disappear, a pale scar is often visible in the healed area. A primary union describes the healing of a wound where the edges are close together. When there is a gaping wound, it takes longer to refill the area with cells and collagen. The process called secondary union occurs as the edges of the wound are pulled together by what is called wound contraction. When a wound is more than one quarter of an inch deep, sutures (stitches) are recommended to promote a primary union and avoid the formation of a disfiguring scar. Regeneration is the addition of new cells of the same type as the ones that were injured (Figure 3.6.1).
Figure 3.6.1. Tissue healing. During wound repair, collagen fibres are laid down randomly by fibroblasts that move into repair the area.
Tissue and Ageing
According to poet Ralph Waldo Emerson, “The surest poison is time.” In fact, biology confirms that many functions of the body decline with age. All the cells, tissues, and organs are affected by senescence, with noticeable variability between individuals owing to different genetic makeup and lifestyles. The outward signs of ageing are easily recognisable. The skin and other tissues become thinner and drier, reducing their elasticity, contributing to wrinkles and high blood pressure. Hair turns grey because follicles produce less melanin, the brown pigment of hair and the iris of the eye. The face looks flabby because elastic and collagen fibres decrease in connective tissue and muscle tone is lost. Glasses and hearing aids may become parts of life as the senses slowly deteriorate, all due to reduced elasticity. Overall height decreases as the bones lose calcium and other minerals. With age, fluid decreases in the fibrous cartilage disks intercalated between the vertebrae in the spine. Joints lose cartilage and stiffen. Many tissues, including those in muscles, lose mass through a process called atrophy. Lumps and rigidity become more widespread. As a consequence, the passageways, blood vessels, and airways become more rigid. The brain and spinal cord lose mass. Nerves do not transmit impulses with the same speed and frequency as in the past. Some loss of thought clarity and memory can accompany ageing. More severe problems are not necessarily associated with the ageing process and may be symptoms of underlying illness.
As exterior signs of ageing increase, so do the interior signs, which are not as noticeable. The incidence of heart diseases, respiratory syndromes, and type 2 diabetes increases with age, though these are not necessarily age-dependent effects. Wound healing is slower in the elderly, accompanied by a higher frequency of infection as the capacity of the immune system to fend off pathogen declines.
Ageing is also apparent at the cellular level because all cells experience changes with ageing. Telomeres, regions of the chromosomes necessary for cell division, shorten each time cells divide. As they do, cells are less able to divide and regenerate. Because of alterations in cell membranes, transport of oxygen and nutrients into the cell and removal of carbon dioxide and waste products from the cell are not as efficient in the elderly. Cells may begin to function abnormally, which may lead to diseases associated with ageing, including arthritis, memory issues, and some cancers.
The progressive impact of ageing on the body varies considerably among individuals, with some studies indicating that exercise and healthy lifestyle choices may slow down the deterioration of the body that comes with old age.
Homeostatic Imbalances
Tissues and Cancer
Cancer is a generic term for many diseases in which cells escape regulatory signals. Uncontrolled growth, invasion into adjacent tissues, and colonisation of other organs, if not treated early enough, are its hallmarks. Health suffers when tumours “rob” blood supply from the “normal” organs.
A mutation is defined as a permanent change in the DNA of a cell. Epigenetic modifications, changes that do not affect the code of the DNA but alter how the DNA is decoded, are also known to generate abnormal cells. Alterations in the genetic material may be caused by environmental agents, infectious agents, or errors in the replication of DNA that accumulate with age. Many mutations do not cause any noticeable change in the functions of a cell. However, if the modification affects key proteins that have an impact on the cell’s ability to proliferate in an orderly fashion, the cell starts to divide abnormally. As changes in cells accumulate, they lose their ability to form regular tissues. A tumour, a mass of cells displaying abnormal architecture, forms in the tissue. Many tumours are benign, meaning they do not metastasise nor cause disease. A tumour becomes malignant, or cancerous, when it breaches the confines of its tissue, promotes angiogenesis, attracts the growth of capillaries, and metastasises to other organs (Figure 3.6.2). The specific names of cancers reflect the tissue of origin. Cancers derived from epithelial cells are referred to as carcinomas. Cancer in myeloid tissue or blood cells form myelomas. Leukemias are cancers of white blood cells, whereas sarcomas derive from connective tissue. Cells in tumours differ both in structure and function. Some cells, called cancer stem cells, appear to be a subtype of cell responsible for uncontrolled growth. Recent research shows that contrary to what was previously assumed, tumours are not disorganised masses of cells, but have their own structures.
Figure 3.6.2. Development of cancer. Note the change in cell size, nucleus size and organisation in the tissue.
Cancer treatments vary depending on the disease’s type and stage. Traditional approaches, including surgery, radiation, chemotherapy, and hormonal therapy, aim to remove or kill rapidly dividing cancer cells, but these strategies have their limitations. Depending on a tumour’s location, for example, cancer surgeons may be unable to remove it. Radiation and chemotherapy are difficult, and it is often impossible to target only the cancer cells. The treatments inevitably destroy healthy tissue as well. To address this, researchers are working on pharmaceuticals that can target specific proteins implicated in cancer-associated molecular pathways.
Section Review
Inflammation is the classic response of the body to injury and follows a common sequence of events. The area is red, feels warm to the touch, swells, and is painful. Injured cells, mast cells, and resident macrophages release chemical signals that cause vasodilation and fluid leakage in the surrounding tissue. The repair phase includes blood clotting, followed by regeneration of tissue as fibroblasts deposit collagen. Some tissues regenerate more readily than others. Epithelial and connective tissues replace damaged or dead cells from a supply of adult stem cells. Muscle and nervous tissues undergo either slow regeneration or do not repair at all.
Age affects all the tissues and organs of the body. Damaged cells do not regenerate as rapidly as in younger people. Perception of sensation and effectiveness of response are lost in the nervous system. Muscles atrophy, and bones lose mass and become brittle. Collagen decreases in some connective tissue, and joints stiffen.
Identify the components of the integumentary system
Describe the layers of the skin and the functions of each layer
Identify and describe the hypodermis and deep fascia
Describe the role of keratinocytes and their life cycle
Describe the role of melanocytes in skin pigmentation
Although you may not typically think of the skin as an organ, it is in fact made of tissues that work together as a single structure to perform unique and critical functions. The skin and its accessory structures make up the integumentary system, which provides the body with overall protection. The skin is made of multiple layers of cells and tissues, which are held to underlying structures by connective tissue (Figure 4.1.1). The deeper layer of skin is well vascularised (has numerous blood vessels). It also has numerous sensory, and autonomic and sympathetic nerve fibres ensuring communication to and from the brain.
Figure 4.1.1. Layers of skin. The skin is composed of two main layers: the epidermis, made of closely packed epithelial cells, and the dermis, made of dense, irregular connective tissue that houses blood vessels, hair follicles, sweat glands, and other structures. Beneath the dermis lies the hypodermis, which is composed mainly of loose connective and fatty tissues.
The Epidermis
The epidermis is composed of keratinised, stratified squamous epithelium. It is made of four or five layers of epithelial cells, depending on its location in the body. It does not have any blood vessels within it (i.e., it is avascular). Skin that has four layers of cells is referred to as “thin skin.” From deep to superficial, these layers are the stratum basale, stratum spinosum, stratum granulosum, and stratum corneum. Most of the skin can be classified as thin skin. “Thick skin” is found only on the palms of the hands and the soles of the feet. It has a fifth layer, called the stratum lucidum, located between the stratum corneum and the stratum granulosum (Figure 4.1.2).
The cells in all of the layers except the stratum basale are called keratinocytes. A keratinocyte is a cell that manufactures and stores the protein keratin. Keratin is an intracellular fibrous protein that gives hair, nails, and skin their hardness and water-resistant properties. The keratinocytes in the stratum corneum are dead and regularly slough away, being replaced by cells from the deeper layers (Figure 4.1.3).
The stratum basale (also called the stratum germinativum) is the deepest epidermal layer and attaches the epidermis to the basal lamina, below which lie the layers of the dermis. The cells in the stratum basale bond to the dermis via intertwining collagen fibres, referred to as the basement membrane. A finger-like projection, or fold, known as the dermal papilla (plural = dermal papillae) is found in the superficial portion of the dermis. Dermal papillae increase the strength of the connection between the epidermis and dermis; the greater the folding, the stronger the connections made (Figure 4.1.4).
Figure 4.1.4. Layers of the epidermis. The epidermis of thick skin has five layers: stratum basale, stratum spinosum, stratum granulosum, stratum lucidum, and stratum corneum.
The stratum basale is a single layer of cells primarily made of basal cells. A basal cell is a cuboidal-shaped stem cell that is a precursor of the keratinocytes of the epidermis. All of the keratinocytes are produced from this single layer of cells, which are constantly going through mitosis to produce new cells. As new cells are formed, the existing cells are pushed superficially away from the stratum basale. Two other cell types are found dispersed among the basal cells in the stratum basale. The first is a Merkel cell, which functions as a receptor and is responsible for stimulating sensory nerves that the brain perceives as touch. These cells are especially abundant on the surfaces of the hands and feet. The second is a melanocyte, a cell that produces the pigment melanin. Melanin gives hair and skin its colour, and also helps protect the living cells of the epidermis from ultraviolet (UV) radiation damage.
In a growing foetus, fingerprints form where the cells of the stratum basale meet the papillae of the underlying dermal layer (papillary layer), resulting in the formation of the ridges on your fingers that you recognise as fingerprints. Fingerprints are unique to each individual and are used for forensic analyses because the patterns do not change with the growth and ageing processes.
Stratum Spinosum
As the name suggests, the stratum spinosum is spiny in appearance due to the protruding cell processes that join the cells via a structure called a desmosome. The desmosomes interlock with each other and strengthen the bond between the cells. It is interesting to note that the “spiny” nature of this layer is an artifact of the staining process. Unstained epidermis samples do not exhibit this characteristic appearance. The stratum spinosum is composed of eight to 10 layers of keratinocytes, formed as a result of cell division in the stratum basale (Figure 4.1.5). Interspersed among the keratinocytes of this layer is a type of dendritic cell called the Langerhans cell, which functions as a macrophage by engulfing bacteria, foreign particles, and damaged cells that occur in this layer.
The keratinocytes in the stratum spinosum begin the synthesis of keratin and release a water-repelling glycolipid that helps prevent water loss from the body, making the skin relatively waterproof. As new keratinocytes are produced atop the stratum basale, the keratinocytes of the stratum spinosum are pushed into the stratum granulosum.
Stratum Granulosum
The stratum granulosum has a grainy appearance due to further changes to the keratinocytes as they are pushed from the stratum spinosum. The cells (three to five layers deep) become flatter, their cell membranes thicken, and they generate large amounts of the proteins keratin, which is fibrous, and keratohyalin, which accumulates as lamellar granules within the cells (see Figure 4.1.4 and Figure 4.1.5). These two proteins make up the bulk of the keratinocyte mass in the stratum granulosum and give the layer its grainy appearance. The nuclei and other cell organelles disintegrate as the cells die, leaving behind the keratin, keratohyalin, and cell membranes that will form the stratum lucidum, the stratum corneum, and the accessory structures of hair and nails.
Stratum Lucidum
The stratum lucidum is a smooth, seemingly translucent layer of the epidermis located just above the stratum granulosum and below the stratum corneum. This thin layer of cells is found only in the thick skin of the palms, soles, and digits. The keratinocytes that compose the stratum lucidum are dead and flattened (see Figure 4.1.4 and Figure 4.1.5). These cells are densely packed with eleidin, a clear protein rich in lipids, derived from keratohyalin, which gives these cells their transparent (i.e., lucid) appearance and provides a barrier to water.
Stratum Corneum
The stratum corneum is the most superficial layer of the epidermis and is the layer exposed to the outside environment (see Figure 4.1.4 and Figure 4.1.5). The increased keratinisation (also called cornification) of the cells in this layer gives it its name. There are usually 15 to 30 layers of cells in the stratum corneum. This dry, dead layer helps prevent the penetration of microbes and the dehydration of underlying tissues and provides a mechanical protection against abrasion for the more delicate, underlying layers. Cells in this layer are shed periodically and are replaced by cells pushed up from the stratum granulosum (or stratum lucidum in the case of the palms and soles of feet). The entire layer is replaced during a period of about 4 weeks. Cosmetic procedures, such as microdermabrasion, help remove some of the dry, upper layer and aim to keep the skin looking “fresh” and healthy.
Dermis
The dermis might be considered the “core” of the integumentary system (derma- = “skin”), as distinct from the epidermis (epi- = “upon” or “over”) and hypodermis (hypo- = “below”). It contains blood and lymph vessels, nerves, and other structures, such as hair follicles and sweat glands. The dermis is made of two layers of connective tissue that compose an interconnected mesh of elastin and collagenous fibres, produced by fibroblasts (Figure 4.1.6).
Figure 4.1.6. Layers of the dermis. This stained slide shows the two components of the dermis—the papillary layer and the reticular layer. Both are made of connective tissue with fibres of collagen extending from one to the other, making the border between the two somewhat indistinct. The dermal papillae extending into the epidermis belong to the papillary layer, whereas the dense collagen fibre bundles below belong to the reticular layer. LM × 10. (credit: modification of work by “kilbad”/Wikimedia Commons).
Papillary Layer
The papillary layer is made of loose, areolar connective tissue, which means the collagen and elastin fibres of this layer form a loose mesh. This superficial layer of the dermis projects into the stratum basale of the epidermis to form finger-like dermal papillae (see Figure 4.1.6). Within the papillary layer are fibroblasts, a small number of fat cells (adipocytes), and an abundance of small blood vessels. In addition, the papillary layer contains phagocytes, defensive cells that help fight bacteria or other infections that have breached the skin. This layer also contains lymphatic capillaries, nerve fibres, and touch receptors called the Meissner corpuscles.
Reticular Layer
Underlying the papillary layer is the much thicker reticular layer, composed of dense, irregular connective tissue. This layer is well vascularised and has a rich sensory and sympathetic nerve supply. The reticular layer appears reticulated (net-like) due to a tight meshwork of fibres. Elastin fibres provide some elasticity to the skin, enabling movement. Collagen fibres provide structure and tensile strength, with strands of collagen extending into both the papillary layer and the hypodermis. In addition, collagen binds water to keep the skin hydrated. Collagen injections and Retin-A creams help restore skin turgor by either introducing collagen externally or stimulating blood flow and repair of the dermis, respectively.
Hypodermis
The hypodermis (also called the subcutaneous layer or superficial fascia) is a layer directly below the dermis and serves to connect the skin to the underlying fascia (fibrous tissue) of the bones and muscles. It is not strictly a part of the skin, although the border between the hypodermis and dermis can be difficult to distinguish. The hypodermis consists of well-vascularised, loose, areolar connective tissue and adipose tissue, which functions as a mode of fat storage and provides insulation and cushioning for the integument.
Everyday Connection
Lipid Storage
The hypodermis is home to most of the fat that concerns people when they are trying to keep their weight under control. Adipose tissue present in the hypodermis consists of fat-storing cells called adipocytes. This stored fat can serve as an energy reserve, insulate the body to prevent heat loss, and act as a cushion to protect underlying structures from trauma.
Where the fat is deposited and accumulates within the hypodermis depends on hormones (testosterone, oestrogen, insulin, glucagon, leptin and others), as well as genetic factors. Fat distribution changes as our bodies mature and age. Men tend to accumulate fat in different areas (neck, arms, lower back, and abdomen) than do women (breasts, hips, thighs, and buttocks). The body mass index (BMI) is often used as a measure of fat, although this measure is, in fact, derived from a mathematical formula that compares body weight (mass) to height. Therefore, its accuracy as a health indicator can be called into question in individuals who are extremely physically fit.
In many animals, there is a pattern of storing excess calories as fat to be used in times when food is not readily available. In much of the developed world, insufficient exercise coupled with the ready availability and consumption of high-calorie foods have resulted in unwanted accumulations of adipose tissue in many people. Although periodic accumulation of excess fat may have provided an evolutionary advantage to our ancestors, who experienced unpredictable bouts of famine, it is now becoming chronic and considered a major health threat. Recent studies indicate that a distressing percentage of our population is overweight and/or clinically obese. Not only is this a problem for the individuals affected, but it also has a severe impact on our healthcare system. Changes in lifestyle, specifically in diet and exercise, are the best ways to control body fat accumulation, especially when it reaches levels that increase the risk of heart disease and diabetes.
Pigmentation
The colour of skin is influenced by a number of pigments, including melanin, carotene and haemoglobin. Recall that melanin is produced by cells called melanocytes, which are found scattered throughout the stratum basale of the epidermis. The melanin is transferred into the keratinocytes via a cellular vesicle called a melanosome (Figure 4.1.7).
Figure 4.1.7. Skin pigmentation. The relative colouration of the skin depends on the amount of melanin produced by melanocytes in the stratum basale and taken up by keratinocytes.
Melanin occurs in two primary forms. Eumelanin exists as black and brown, whereas pheomelanin provides a red colour. Dark-skinned individuals produce more melanin than those with pale skin. Exposure to the UV rays of the sun or a tanning salon causes melanin to be manufactured and built up in keratinocytes, as sun exposure stimulates keratinocytes to secrete chemicals that stimulate melanocytes. The accumulation of melanin in keratinocytes results in the darkening of the skin, or a tan. This increased melanin accumulation protects the DNA of epidermal cells from UV ray damage and the breakdown of folic acid, a nutrient necessary for our health and well-being. In contrast, too much melanin can interfere with the production of vitamin D, an important nutrient involved in calcium absorption. Thus, the amount of melanin present in our skin is dependent on a balance between available sunlight and folic acid destruction, and protection from UV radiation and vitamin D production.
It requires about 10 days after initial sun exposure for melanin synthesis to peak, which is why pale-skinned individuals tend to suffer sunburns of the epidermis initially. Dark-skinned individuals can also get sunburned but are more protected than are pale-skinned individuals. Melanosomes are temporary structures that are eventually destroyed by fusion with lysosomes; this fact, along with melanin-filled keratinocytes in the stratum corneum sloughing off, makes tanning impermanent.
Too much sun exposure can eventually lead to wrinkling due to the destruction of the cellular structure of the skin, and in severe cases, can cause sufficient DNA damage to result in skin cancer. When there is an irregular accumulation of melanocytes in the skin, freckles appear. Moles are larger masses of melanocytes, and although most are benign, they should be monitored for changes that might indicate the presence of cancer (Figure 4.1.8).
Figure 4.1.8. Moles. Moles range from benign accumulations of melanocytes to melanomas. These structures populate the landscape of our skin. (credit: the National Cancer Institute).
Disorders of the Integumentary System
Figure 4.1.9. Vitiligo. Individuals with vitiligo experience depigmentation that results in lighter coloured patches of skin. The condition is especially noticeable on darker skin. (credit: Klaus D. Peter).
The first thing a clinician sees is the skin, and so the examination of the skin should be part of any thorough physical examination. Most skin disorders are relatively benign, but a few, including melanomas, can be fatal if untreated. A couple of the more noticeable disorders, albinism and vitiligo, affect the appearance of the skin and its accessory organs. Although neither is fatal, it would be hard to claim that they are benign, at least to the individuals so afflicted.
Albinism is a genetic disorder that affects (completely or partially) the colouring of skin, hair, and eyes. The defect is primarily due to the inability of melanocytes to produce melanin. Individuals with albinism tend to appear white or very pale due to the lack of melanin in their skin and hair. Recall that melanin helps protect the skin from the harmful effects of UV radiation. Individuals with albinism tend to need more protection from UV radiation, as they are more prone to sunburns and skin cancer. They also tend to be more sensitive to light and have vision problems due to the lack of pigmentation on the retinal wall. Treatment of this disorder usually involves addressing the symptoms, such as limiting UV light exposure to the skin and eyes. In vitiligo, the melanocytes in certain areas lose their ability to produce melanin, possibly due to an autoimmune reaction. This leads to a loss of colour in patches (Figure 4.1.9). Neither albinism nor vitiligo directly affects the lifespan of an individual.
Section Review
The skin is composed of two major layers: a superficial epidermis and a deeper dermis. The epidermis consists of several layers beginning with the innermost (deepest) stratum basale (germinatum), followed by the stratum spinosum, stratum granulosum, stratum lucidum (when present), and ending with the outermost layer, the stratum corneum. The topmost layer, the stratum corneum, consists of dead cells that shed periodically and is progressively replaced by cells formed from the basal layer. The stratum basale also contains melanocytes, cells that produce melanin, the pigment primarily responsible for giving skin its colour. Melanin is transferred to keratinocytes in the stratum spinosum to protect cells from UV rays.
The dermis connects the epidermis to the hypodermis and provides strength and elasticity due to the presence of collagen and elastin fibres. It has only two layers: the papillary layer with papillae that extend into the epidermis and the lower, reticular layer composed of loose connective tissue. The hypodermis, deep to the dermis of skin, is the connective tissue that connects the dermis to underlying structures; it also harbors adipose tissue for fat storage and protection.
Describe the structure and function of hair and nails
Describe the structure and function of sweat glands and sebaceous glands
Accessory structures of the skin include hair, nails, sweat glands, and sebaceous glands. These structures embryologically originate from the epidermis and can extend down through the dermis into the hypodermis.
Hair
Hair is a keratinous filament growing out of the epidermis. It is primarily made of dead, keratinised cells. Strands of hair originate in an epidermal penetration of the dermis called the hair follicle. The hair shaft is the part of the hair not anchored to the follicle, and much of this is exposed at the skin’s surface. The rest of the hair, which is anchored in the follicle, lies below the surface of the skin and is referred to as the hair root. The hair root ends deep in the dermis at the hair bulb and includes a layer of mitotically active basal cells called the hair matrix. The hair bulb surrounds the hair papilla, which is made of connective tissue and contains blood capillaries and nerve endings from the dermis (Figure 4.2.1).
Figure 4.2.1. Hair. Hair follicles originate in the epidermis and have many different parts.
Just as the basal layer of the epidermis forms the layers of epidermis that get pushed to the surface as the dead skin on the surface sheds, the basal cells of the hair bulb divide and push cells outward in the hair root and shaft as the hair grows. The medulla forms the central core of the hair, which is surrounded by the cortex, a layer of compressed, keratinised cells that is covered by an outer layer of very hard, keratinised cells known as the cuticle. These layers are depicted in a longitudinal cross-section of the hair follicle (Figure 4.2.2), although not all hair has a medullary layer. Hair texture (straight, curly) is determined by the shape and structure of the cortex, and to the extent that it is present, the medulla. The shape and structure of these layers are, in turn, determined by the shape of the hair follicle. Hair growth begins with the production of keratinocytes by the basal cells of the hair bulb. As new cells are deposited at the hair bulb, the hair shaft is pushed through the follicle toward the surface. Keratinisation is completed as the cells are pushed to the skin surface to form the shaft of hair that is externally visible. The external hair is completely dead and composed entirely of keratin. For this reason, our hair does not have sensation. Furthermore, you can cut your hair or shave without damaging the hair structure because the cut is superficial. Most chemical hair removers also act superficially; however, electrolysis and yanking both attempt to destroy the hair bulb so hair cannot grow.
Figure 4.2.2. Hair Follicle. The slide shows a cross-section of a hair follicle. Basal cells of the hair matrix in the centre differentiate into cells of the inner root sheath. Basal cells at the base of the hair root form the outer root sheath. LM × 4. (credit: modification of work by “kilbad”/Wikimedia Commons).
The wall of the hair follicle is made of three concentric layers of cells. The cells of the internal root sheath surround the root of the growing hair and extend just up to the hair shaft. They are derived from the basal cells of the hair matrix. The external root sheath, which is an extension of the epidermis, encloses the hair root. It is made of basal cells at the base of the hair root and tends to be more keratinous in the upper regions. The glassy membrane is a thick, clear connective tissue sheath covering the hair root, connecting it to the tissue of the dermis.
Hair serves a variety of functions, including protection, sensory input, thermoregulation and communication, for example, hair on the head protects the skull from the sun. The hair in the nose and ears, and around the eyes (eyelashes) defends the body by trapping and excluding dust particles that may contain allergens and microbes. Hair of the eyebrows prevents sweat and other particles from dripping into and bothering the eyes. Hair also has a sensory function due to sensory innervation by a hair root plexus surrounding the base of each hair follicle. Hair is extremely sensitive to air movement or other disturbances in the environment, much more so than the skin surface. This feature is also useful for the detection of the presence of insects or other potentially damaging substances on the skin surface. Each hair root is connected to a smooth muscle called the arrector pili that contracts in response to nerve signals from the sympathetic nervous system, making the external hair shaft “stand up.” The primary purpose for this is to trap a layer of air to add insulation. This is visible in humans as goose bumps and even more obvious in animals, such as when a frightened cat raises its fur. Of course, this is much more obvious in organisms with a heavier coat than most humans, such as dogs and cats.
Hair Growth
Hair grows and is eventually shed and replaced by new hair. This occurs in three phases. The first is the anagen phase, during which cells divide rapidly at the root of the hair, pushing the hair shaft up and out. The length of this phase is measured in years, typically from 2 to 7 years. The catagen phase lasts only 2 to 3 weeks and marks a transition from the hair follicle’s active growth. Finally, during the telogen phase, the hair follicle is at rest and no new growth occurs. At the end of this phase, which lasts about 2 to 4 months, another anagen phase begins. The basal cells in the hair matrix then produce a new hair follicle, which pushes the old hair out as the growth cycle repeats itself. Hair typically grows at the rate of 0.3 mm per day during the anagen phase. On average, 50 hairs are lost and replaced per day. Hair loss occurs if there is more hair shed than what is replaced and can happen due to hormonal or dietary changes. Hair loss can also result from the ageing process, or the influence of hormones.
Hair Colour
Similar to the skin, hair gets its colour from the pigment melanin, produced by melanocytes in the hair papilla. Different hair colour results from differences in the type of melanin, which is genetically determined. As a person ages, the melanin production decreases, and hair tends to lose its colour and becomes grey and/or white.
Nails
The nail bed is a specialised structure of the epidermis that is found at the tips of our fingers and toes. The nail body is formed on the nail bed, and protects the tips of our fingers and toes as they are the farthest extremities and the parts of the body that experience the maximum mechanical stress (Figure 4.2.3). In addition, the nail body forms a back-support for picking up small objects with the fingers. The nail body is composed of densely packed dead keratinocytes. The epidermis in this part of the body has evolved a specialised structure upon which nails can form. The nail body forms at the nail root, which has a matrix of proliferating cells from the stratum basale that enables the nail to grow continuously. The lateral nail fold overlaps the nail on the sides, helping to anchor the nail body. The nail fold that meets the proximal end of the nail body forms the nail cuticle, also called the eponychium. The nail bed is rich in blood vessels, making it appear pink, except at the base, where a thick layer of epithelium over the nail matrix forms a crescent-shaped region called the lunula (the “little moon”). The area beneath the free edge of the nail, furthest from the cuticle, is called the hyponychium. It consists of a thickened layer of stratum corneum.
Figure 4.2.3. Nails. The nail is an accessory structure of the integumentary system.
Sweat Glands
When the body becomes warm, sudoriferous glands produce sweat to cool the body. Sweat glands develop from epidermal projections into the dermis and are classified as merocrine glands; that is, the secretions are excreted by exocytosis through a duct without affecting the cells of the gland. There are two types of sweat glands, each secreting slightly different products.
An eccrine sweat gland is type of gland that produces a hypotonic sweat for thermoregulation. These glands are found all over the skin’s surface, but are especially abundant on the palms of the hand, the soles of the feet, and the forehead (Figure 4.2.4). They are coiled glands lying deep in the dermis, with the duct rising up to a pore on the skin surface, where the sweat is released. This type of sweat, released by exocytosis, is hypotonic and composed mostly of water, with some salt, antibodies, traces of metabolic waste, and dermicidin, an antimicrobial peptide. Eccrine glands are a primary component of thermoregulation in humans and thus help to maintain homeostasis.
Figure 4.2.4. Eccrine Gland. Eccrine glands are coiled glands in the dermis that release sweat that is mostly water.
An apocrine sweat gland is usually associated with hair follicles in densely hairy areas, such as armpits and genital regions. Apocrine sweat glands are larger than eccrine sweat glands and lie deeper in the dermis, sometimes even reaching the hypodermis, with the duct normally emptying into the hair follicle. In addition to water and salts, apocrine sweat includes organic compounds that make the sweat thicker and subject to bacterial decomposition and subsequent smell. The release of this sweat is under both nervous and hormonal control and plays a role in the poorly understood human pheromone response. Most commercial antiperspirants use an aluminium-based compound as their primary active ingredient to stop sweat. When the antiperspirant enters the sweat gland duct, the aluminium-based compounds precipitate due to a change in pH and form a physical block in the duct, which prevents sweat from coming out of the pore.
Sebaceous Glands
A sebaceous gland is a type of oil gland that is found all over the body and helps to lubricate and waterproof the skin and hair. Most sebaceous glands are associated with hair follicles. They generate and excrete sebum, a mixture of lipids, onto the skin surface, thereby naturally lubricating the dry and dead layer of keratinised cells of the stratum corneum, keeping it pliable. The fatty acids of sebum also have antibacterial properties and prevent water loss from the skin in low-humidity environments. The secretion of sebum is stimulated by hormones, many of which do not become active until puberty. Thus, sebaceous glands are relatively inactive during childhood.
Section Review
Accessory structures of the skin include hair, nails, sweat glands, and sebaceous glands. Hair is made of dead keratinised cells and gets its colour from melanin pigments. Nails, also made of dead keratinised cells, protect the extremities of our fingers and toes from mechanical damage. Sweat glands and sebaceous glands produce sweat and sebum, respectively. Each of these fluids has a role to play in maintaining homeostasis. Sweat cools the body surface when it gets overheated and helps excrete small amounts of metabolic waste. Sebum acts as a natural moisturiser and keeps the dead, flaky, outer keratin layer healthy.
Describe the different functions of the skin and the structures that enable them
Explain how the skin helps maintain body temperature
The skin and accessory structures perform a variety of essential functions, such as protecting the body from invasion by microorganisms, chemicals, and other environmental factors; preventing dehydration; acting as a sensory organ; modulating body temperature and electrolyte balance; and synthesising vitamin D. The underlying hypodermis has important roles in storing fats, forming a “cushion” over underlying structures, and providing insulation from cold temperatures.
Protection
The skin protects the rest of the body from the basic elements of nature such as wind, water, and UV sunlight. It acts as a protective barrier against water loss, due to the presence of layers of keratin and glycolipids in the stratum corneum. It also is the first line of defence against abrasive activity due to contact with grit, microbes, or harmful chemicals. Sweat excreted from sweat glands deters microbes from over-colonising the skin surface by generating dermicidin, which has antibiotic properties.
Everyday Connection
Tattoos and Piercings
The word “armour” evokes several images. You might think of a Roman centurion or a medieval knight in a suit of armour. The skin, in its own way, functions as a form of armour—body armour. It provides a barrier between your vital, life-sustaining organs and the influence of outside elements that could potentially damage them.
For any form of armour, a breach in the protective barrier poses a danger. The skin can be breached when a child skins a knee, or an adult has blood drawn—one is accidental and the other medically necessary. However, you also breach this barrier when you choose to “accessorise” your skin with a tattoo or body piercing. Because the needles involved in producing body art and piercings must penetrate the skin, there are dangers associated with the practice. These include allergic reactions; skin infections; blood-borne diseases, such as tetanus, hepatitis C and hepatitis D; and the growth of scar tissue. Despite the risk, the practice of piercing the skin for decorative purposes has become increasingly popular. According to the American Academy of Dermatology, 24 percent of people from ages 18 to 50 have a tattoo.
Sensory Function
The fact that you can feel an ant crawling on your skin, allowing you to flick it off before it bites, is because the skin, and especially the hairs projecting from hair follicles in the skin, can sense changes in the environment. The hair root plexus surrounding the base of the hair follicle senses a disturbance, and then transmits the information to the central nervous system (brain and spinal cord), which can then respond by activating the skeletal muscles of your eyes to see the ant and the skeletal muscles of the body to act against the ant.
The skin acts as a sense organ because the epidermis, dermis, and the hypodermis contain specialised sensory nerve structures that detect touch, surface temperature, and pain. These receptors are more concentrated on the tips of the fingers, which are most sensitive to touch, especially the Meissner corpuscle (tactile corpuscle) (Figure 4.3.1), which responds to light touch, and the Pacinian corpuscle (lamellated corpuscle), which responds to vibration. Merkel cells, seen scattered in the stratum basale, are also touch receptors. In addition to these specialised receptors, there are sensory nerves connected to each hair follicle, pain and temperature receptors scattered throughout the skin, and motor nerves innervate the arrector pili muscles and glands. This rich innervation helps us sense our environment and react accordingly.
Figure 4.3.1. Light micrograph of a Meissner corpuscle. In this micrograph of a skin cross-section, you can see a Meissner corpuscle (arrow), a type of touch receptor located in a dermal papilla adjacent to the basement membrane and stratum basale of the overlying epidermis. LM × 100. (credit: “Wbensmith”/Wikimedia Commons).
Thermoregulation
The integumentary system helps regulate body temperature through its tight association with the sympathetic nervous system, the division of the nervous system involved in our fight-or-flight responses. The sympathetic nervous system is continuously monitoring body temperature and initiating appropriate motor responses. Recall that sweat glands, accessory structures to the skin, secrete water, salt, and other substances to cool the body when it becomes warm. Even when the body does not appear to be noticeably sweating, approximately 500 mL of sweat (insensible perspiration) are secreted a day. If the body becomes excessively warm due to high temperatures, vigorous activity (Figure 4.3.2ac), or a combination of the two, sweat glands will be stimulated by the sympathetic nervous system to produce large amounts of sweat, as much as 0.7 to 1.5 L per hour for an active person. When the sweat evaporates from the skin surface, the body is cooled as body heat is dissipated.
In addition to sweating, arterioles in the dermis dilate so that excess heat carried by the blood can dissipate through the skin and into the surrounding environment (Figure 4.3.2b). This accounts for the skin redness that many people experience when exercising.
Figure 4.3.2. Thermoregulation. During strenuous physical activities, such as skiing (a) or running (c), the dermal blood vessels dilate and sweat secretion increases (b). These mechanisms prevent the body from overheating. In contrast, the dermal blood vessels constrict to minimise heat loss in response to low temperatures (b). (credit a: “Trysil”/flickr; credit c: Ralph Daily).
When body temperatures drop, the arterioles constrict to minimise heat loss, particularly in the ends of the digits and tip of the nose. This reduced circulation can result in the skin taking on a whitish hue. Although the temperature of the skin drops as a result, passive heat loss is prevented, and internal organs and structures remain warm. If the temperature of the skin drops too much (such as environmental temperatures below freezing), the conservation of body core heat can result in the skin actually freezing, a condition called frostbite.
Ageing and the Integumentary System
All systems in the body accumulate subtle and some not-so-subtle changes as a person ages. Among these changes are reductions in cell division, metabolic activity, blood circulation, hormonal levels, and muscle strength (Figure 4.3.3). In the skin, these changes are reflected in decreased mitosis in the stratum basale, leading to a thinner epidermis. The dermis, which is responsible for the elasticity and resilience of the skin, exhibits a reduced ability to regenerate, which leads to slower wound healing. The hypodermis, with its fat stores, loses structure due to the reduction and redistribution of fat, which in turn contributes to the thinning and sagging of skin.
Figure 4.3.3. Ageing. Generally, skin, especially on the face and hands, starts to display the first noticeable signs of ageing, as it loses its elasticity over time. (credit: Janet Ramsden).
The accessory structures also have lowered activity, generating thinner hair and nails, and reduced amounts of sebum and sweat. A reduced sweating ability can cause some elderly to be intolerant to extreme heat. Other cells in the skin, such as melanocytes and dendritic cells, also become less active, leading to a paler skin tone and lowered immunity. Wrinkling of the skin occurs due to breakdown of its structure, which results from decreased collagen and elastin production in the dermis, weakening of muscles lying under the skin, and the inability of the skin to retain adequate moisture.
Many anti-ageing products can be found in stores today. In general, these products try to rehydrate the skin and thereby fill out the wrinkles, and some stimulate skin growth using hormones and growth factors. Additionally, invasive techniques include collagen injections to plump the tissue and injections of BOTOX® (the name brand of the botulinum neurotoxin) that paralyse the muscles that crease the skin and cause wrinkling.
Vitamin D Synthesis
The epidermal layer of human skin synthesises vitamin D when exposed to UV radiation. In the presence of sunlight, a form of vitamin D3 called cholecalciferol is synthesised from a derivative of the steroid cholesterol in the skin. The liver converts cholecalciferol to calcidiol, which is then converted to calcitriol (the active chemical form of the vitamin) in the kidneys. Vitamin D is essential for normal absorption of calcium and phosphorous, which are required for healthy bones. The absence of sun exposure can lead to a lack of vitamin D in the body, leading to a condition called rickets, a painful condition in children where the bones are misshapen due to a lack of calcium, causing bowleggedness. Elderly individuals who suffer from vitamin D deficiency can develop a condition called osteomalacia, a softening of the bones. In present day society, vitamin D is added as a supplement to many foods, including milk and orange juice, compensating for the need for sun exposure.
In addition to its essential role in bone health, vitamin D is essential for general immunity against bacterial, viral, and fungal infections. Recent studies are also finding a link between insufficient vitamin D and cancer.
Section Review
The skin plays important roles in protection, sensing stimuli, thermoregulation, and vitamin D synthesis. It is the first layer of defence to prevent dehydration, infection, and injury to the rest of the body. Sweat glands in the skin allow the skin surface to cool when the body gets overheated. Thermoregulation is also accomplished by the dilation or constriction of heat-carrying blood vessels in the skin. Immune cells present among the skin layers patrol the areas to keep them free of foreign materials. Fat stores in the hypodermis aid in both thermoregulation and protection. Finally, the skin plays a role in the synthesis of vitamin D, which is necessary for our well-being but not easily available in natural foods.
4.4 Diseases, Disorders and Injuries of the Integumentary System
Learning Objectives
By the end of this section, you will be able to:
Describe several different diseases and disorders of the skin
Describe the effect of injury to the skin and the process of healing
The integumentary system is susceptible to a variety of diseases, disorders and injuries. These range from annoying but relatively benign bacterial or fungal infections that are categorised as disorders, to skin cancer and severe burns, which can be fatal. In this section, you will learn several of the most common skin conditions.
Diseases
One of the most talked about diseases is skin cancer. Cancer is a broad term that describes diseases caused by abnormal cells in the body dividing uncontrollably. Most cancers are identified by the organ or tissue in which the cancer originates. One common form of cancer is skin cancer. The Skin Cancer Foundation reports that one in five Americans will experience some type of skin cancer in their lifetime. The degradation of the ozone layer in the atmosphere and the resulting increase in exposure to UV radiation has contributed to its rise. Overexposure to UV radiation damages DNA, which can lead to the formation of cancerous lesions. Although melanin offers some protection against DNA damage from the sun, often it is not enough. The fact that cancers can also occur on areas of the body that are normally not exposed to UV radiation suggests that there are additional factors that can lead to cancerous lesions.
In general, cancers result from an accumulation of DNA mutations. These mutations can result in cell populations that do not die when they should and uncontrolled cell proliferation that leads to tumours. Although many tumours are benign (harmless), some produce cells that can mobilise and establish tumours in other organs of the body; this process is referred to as metastasis. Cancers are characterised by their ability to metastasise.
Basal Cell Carcinoma
Figure 4.4.1. Basal cell carcinoma. Basal cell carcinoma can take several different forms. Similar to other forms of skin cancer, it is readily cured if caught early and treated. (credit: John Hendrix, MD).
Basal cell carcinoma is a form of cancer that affects the mitotically active stem cells in the stratum basale of the epidermis. It is the most common of all cancers that occur in the United States and is frequently found on the head, neck, arms, and back, which are areas that are most susceptible to long-term sun exposure. Although UV rays are the main culprit, exposure to other agents, such as radiation and arsenic, can also lead to this type of cancer. Wounds on the skin due to open sores, tattoos, burns, etc. may be predisposing factors as well. Basal cell carcinomas start in the stratum basale and usually spread along this boundary. At some point, they begin to grow toward the surface and become an uneven patch, bump, growth, or scar on the skin surface (Figure 4.4.1). Like most cancers, basal cell carcinomas respond best to treatment when caught early. Treatment options include surgery, freezing (cryosurgery), and topical ointments (Mayo Clinic 2012).
Squamous Cell Carcinoma
Figure 4.4.2. Squamous cell carcinoma. Squamous cell carcinoma presents here as a lesion on an individual’s nose. (credit: the National Cancer Institute).
Squamous cell carcinoma is a cancer that affects the keratinocytes of the stratum spinosum and presents as lesions commonly found on the scalp, ears, and hands (Figure 4.4.2). It is the second most common skin cancer. The American Cancer Society reports that two of 10 skin cancers are squamous cell carcinomas, and it is more aggressive than basal cell carcinoma. If not removed, these carcinomas can metastasise. Surgery and radiation are used to cure squamous cell carcinoma.
Melanoma
A melanoma is a cancer characterised by the uncontrolled growth of melanocytes, the pigment-producing cells in the epidermis. Typically, a melanoma develops from a mole. It is the most fatal of all skin cancers, as it is highly metastatic and can be difficult to detect before it has spread to other organs. Melanomas usually appear as asymmetrical brown and black patches with uneven borders and a raised surface (Figure 4.4.3). Treatment typically involves surgical excision and immunotherapy.
Figure 4.4.3. Melanoma. Melanomas typically present as large brown or black patches with uneven borders and a raised surface. (credit: the National Cancer Institute).
Doctors often give their patients the following ABCDE mnemonic to help with the diagnosis of early-stage melanoma. If you observe a mole on your body displaying these signs, consult a doctor.
Asymmetry – the two sides are not symmetrical
Borders – the edges are irregular in shape
Colour – the colour is varied shades of brown or black
Diameter – it is larger than 6 mm (0.24 in)
Evolving – its shape has changed
Some specialists cite the following additional signs for the most serious form, nodular melanoma:
Elevated – it is raised on the skin surface
Firm – it feels hard to the touch
Growing – it is getting larger
Skin Disorders
Two common skin disorders are eczema and acne. Eczema is an inflammatory condition and occurs in individuals of all ages. Acne involves the clogging of pores, which can lead to infection and inflammation, and is often seen in adolescents. Other disorders, not discussed here, include seborrheic dermatitis (on the scalp), psoriasis, cold sores, impetigo, scabies, hives, and warts.
Eczema
Figure 4.4.4. Eczema. Eczema is a common skin disorder that presents as a red, flaky rash. (credit: “Jambula”/Wikimedia Commons).
Eczema is an allergic reaction that manifests as dry, itchy patches of skin that resemble rashes (Figure 4.4.4). It may be accompanied by swelling of the skin, flaking, and in severe cases, bleeding. Many who suffer from eczema have antibodies against dust mites in their blood, but the link between eczema and allergy to dust mites has not been proven. Symptoms are usually managed with moisturisers, corticosteroid creams and immunosuppressants.
Acne
Acne is a skin disturbance that typically occurs on areas of the skin that are rich in sebaceous glands (face and back). It is most common along with the onset of puberty due to associated hormonal changes but can also occur in infants and continue into adulthood. Hormones, such as androgens, stimulate the release of sebum. An overproduction and accumulation of sebum along with keratin can block hair follicles. This plug is initially white. The sebum, when oxidised by exposure to air, turns black. Acne results from infection by acne-causing bacteria (Propionibacterium and Staphylococcus), which can lead to redness and potential scarring due to the natural wound healing process (Figure 4.4.5).
Figure 4.4.5. Acne. Acne is a result of over-productive sebaceous glands, which leads to formation of blackheads and inflammation of the skin.
Career Connection
Dermatologist
Have you ever had a skin rash that did not respond to over-the-counter creams, or a mole that you were concerned about? Dermatologists help patients with these types of problems and more, on a daily basis. Dermatologists are medical doctors who specialise in diagnosing and treating skin disorders. Like all medical doctors, dermatologists earn a medical degree and then complete several years of residency training. In addition, dermatologists may then participate in a dermatology fellowship or complete additional, specialised training in a dermatology practice. In Australia, the pathway of becoming a dermatologist has four stages. Firstly, a completion of a medical school degree (medical doctor) which takes roughly four to six years. Second step is a completion of a yearlong in-hospital internship. Third step requires a minimum of one year residency at a hospital. The fourth and final stage requires further vocational training at the Fellowship of the Australasian College of Dermatologists (FACD) for approximately four years.
Most dermatologists work in a medical office or private practice setting. They diagnose skin conditions and rashes, prescribe oral and topical medications to treat skin conditions, and may perform simple procedures, such as mole or wart removal. In addition, they may refer patients to an oncologist if skin cancer that has metastasised is suspected. Recently, cosmetic procedures have also become a prominent part of dermatology. Botox injections, laser treatments, and collagen and dermal filler injections are popular among patients, hoping to reduce the appearance of skin ageing.
Dermatology is a competitive specialty in medicine. Limited openings in dermatology residency programs mean that many medical students compete for a few select spots. Dermatology is an appealing specialty to many prospective doctors, because unlike emergency room physicians or surgeons, dermatologists generally do not have to work excessive hours or be “on-call” weekends and holidays. Moreover, the popularity of cosmetic dermatology has made it a growing field with many lucrative opportunities. It is not unusual for dermatology clinics to market themselves exclusively as cosmetic dermatology centres, and for dermatologists to specialise exclusively in these procedures.
Injuries
Because the skin is the part of our bodies that meets the world most directly, it is especially vulnerable to injury. Injuries include burns and wounds, as well as scars and calluses. They can be caused by sharp objects, heat, or excessive pressure or friction to the skin.
Skin injuries set off a healing process that occurs in several overlapping stages. The first step to repairing damaged skin is the formation of a blood clot that helps stop the flow of blood and scabs over with time. Many different types of cells are involved in wound repair, especially if the surface area that needs repair is extensive. Before the basal stem cells of the stratum basale can recreate the epidermis, fibroblasts mobilise and divide rapidly to repair the damaged tissue by collagen deposition, forming granulation tissue. Blood capillaries follow the fibroblasts and help increase blood circulation and oxygen supply to the area. Immune cells, such as macrophages, roam the area and engulf any foreign matter to reduce the chance of infection.
Burns
A burn results when the skin is damaged by intense heat, radiation, electricity, or chemicals. The damage results in the death of skin cells, which can lead to a massive loss of fluid. Dehydration, electrolyte imbalance, and renal and circulatory failure follow, which can be fatal. Burn patients are treated with intravenous fluids to offset dehydration, as well as intravenous nutrients that enable the body to repair tissues and replace lost proteins. Another serious threat to the lives of burn patients is infection. Burned skin is extremely susceptible to bacteria and other pathogens, due to the loss of protection by intact layers of skin.
Figure 4.4.6. Calculating the size of a burn. The size of a burn will guide decisions made about the need for specialised treatment. Specific parts of the body are associated with a percentage of body area.
Burns are sometimes measured in terms of the size of the total surface area affected. This is referred to as the “rule of nines”, which associates specific anatomical areas with a percentage that is a factor of nine (Figure 4.4.6). Burns are also classified by the degree of their severity. In Australia, burns are divided into three main categories: superficial, partial thickness, and full thickness. In other countries, such as United States, these are classified by first, second, third and fourth degree burns. Superficial burns (first-degree) appear pink or red in colour with no visible blisters, and only affect the epidermis and dermal papillae. There are no texture changes to the epidermis, however they tend to be hypersensitive to touch. Although the skin may be painful and swollen, these burns typically heal on their own within a few days. Mild sunburn fits into the category of a superficial burn. Partial thicknessburns (second-degree), sometimes referred to as mid or deep dermal burns, refer to burns in which the entire epidermal layer had been destroyed and the injury reaches the dermis. The depth to which dermal layer is injured determines whether it is classified as a mid or deep dermal burn. Generally, partial thickness burns are distinguished by a creamy coloured base. In a mid-partial burn, sweat glands and hair follicles remain undamaged, however they are affected in a deep partial thickness burn. These burns result in swelling and a painful blistering of the skin. It is important to keep the burn site clean and sterile to prevent infection. If this is done, the burn will heal within several weeks. Full thickness burns (third-degree) are classified as those in which all layers of the epidermis, epithelial elements, dermal appendages, and nerve endings are injured. These burns are distinguished by a leathery appearance which may range from a white, brown, deep red or black. Additionally, the texture of these layers appears leathery and tough. Full-thickness burns cannot be repaired by the body, because the local tissues used for repair are damaged and require excision (debridement), or amputation in severe cases, followed by grafting of the skin from an unaffected part of the body, or from skin grown in tissue culture for grafting purposes. Oddly, full thickness burns are usually not as painful because the nerve endings themselves are damaged. A fourth-degree burn is even more severe, affecting the underlying muscle and bone.
Scars and Keloids
Most cuts or wounds, with the exception of ones that only scratch the surface (the epidermis), lead to scar formation. A scar is collagen-rich skin formed after the process of wound healing that differs from normal skin. Scarring occurs in cases in which there is repair of skin damage, but the skin fails to regenerate the original skin structure. Fibroblasts generate scar tissue in the form of collagen, and the bulk of repair is due to the basket-weave pattern generated by collagen fibres and does not result in regeneration of the typical cellular structure of skin. Instead, the tissue is fibrous in nature and does not allow for the regeneration of accessory structures, such as hair follicles, sweat glands, or sebaceous glands.
Sometimes, there is an overproduction of scar tissue, because the process of collagen formation does not stop when the wound is healed; this results in the formation of a raised or hypertrophic scar called a keloid. In contrast, scars that result from acne and chickenpox have a sunken appearance and are called atrophic scars.
Scarring of skin after wound healing is a natural process and does not need to be treated further. Application of mineral oil and lotions may reduce the formation of scar tissue. However, modern cosmetic procedures, such as dermabrasion, laser treatments, and filler injections have been invented as remedies for severe scarring. All of these procedures try to reorganise the structure of the epidermis and underlying collagen tissue to make it look more natural.
Bedsores and Stretch Marks
Skin and its underlying tissue can be affected by excessive pressure. One example of this is called a bedsore. Bedsores, also called decubitis ulcers, are caused by constant, long-term, unrelieved pressure on certain body parts that are bony, reducing blood flow to the area and leading to necrosis (tissue death). Bedsores are most common in elderly patients who have debilitating conditions that cause them to be immobile. Most hospitals and long-term care facilities have the practice of turning the patients every few hours to prevent the incidence of bedsores. If left untreated by removal of necrotised tissue, bedsores can be fatal if they become infected.
The skin can also be affected by pressure associated with rapid growth. A stretch mark results when the dermis is stretched beyond its limits of elasticity, as the skin stretches to accommodate the excess pressure. Stretch marks usually accompany rapid weight gain during puberty and pregnancy. They initially have a reddish hue but lighten over time. Other than for cosmetic reasons, treatment of stretch marks is not required. They occur most commonly over the hips and abdomen.
Calluses
When you wear shoes that do not fit well and are a constant source of abrasion on your toes, you tend to form a callus at the point of contact. This occurs because the basal stem cells in the stratum basale are triggered to divide more often to increase the thickness of the skin at the point of abrasion to protect the rest of the body from further damage. This is an example of a minor or local injury, and the skin manages to react and treat the problem independent of the rest of the body. Calluses can also form on your fingers if they are subject to constant mechanical stress, such as long periods of writing, playing string instruments, or video games. A corn is a specialised form of callus. Corns form from abrasions on the skin that result from an elliptical-type motion.
Section Review
Skin cancer is a result of damage to the DNA of skin cells, often due to excessive exposure to UV radiation. Basal cell carcinoma and squamous cell carcinoma are highly curable and arise from cells in the stratum basale and stratum spinosum, respectively. Melanoma is the most dangerous form of skin cancer, affecting melanocytes, which can spread/metastasise to other organs. Burns are an injury to the skin that occur as a result of exposure to extreme heat, radiation, or chemicals. Epidermal, superficial dermal and mid dermal usually heal quickly, but deep dermal and full thickness burns can be fatal because they penetrate the full thickness of the skin. Scars occur when there is repair of skin damage. Fibroblasts generate scar tissue in the form of collagen, which forms a basket-weave pattern that looks different from normal skin.
Bedsores and stretch marks are the result of excessive pressure on the skin and underlying tissue. Bedsores are characterised by necrosis of tissue due to immobility, whereas stretch marks result from rapid growth. Eczema is an allergic reaction that manifests as a rash, and acne results from clogged sebaceous glands. Eczema and acne are usually long-term skin conditions that may be treated successfully in mild cases. Calluses and corns are the result of abrasive pressure on the skin.
Identify the primary functions of blood in transportation, defence and maintenance of homeostasis
Name the fluid component of blood and the three major types of cellular components (formed elements), and identify their relative proportions in a blood sample
Discuss the unique physical characteristics of blood
Identify the composition of blood plasma, including its most important solutes and plasma proteins
Recall that blood is a connective tissue. Like all connective tissues, it is made up of cellular elements and an extracellular matrix. The cellular elements—referred to as the cellular components or formed elements—include red blood cells (RBCs), white blood cells (WBCs), and cell fragments called platelets. The extracellular matrix, called plasma, makes blood unique among connective tissues because it is fluid. This fluid, which is mostly water, perpetually suspends the cellular components and enables them to circulate throughout the body within the cardiovascular system.
Function of Blood
The primary function of blood is to deliver oxygen and nutrients to, and remove wastes from, body tissues and cells, but that is only the beginning of the story. The specific functions of blood also include defence, distribution of heat, and maintenance of homeostasis.
Transportation
Nutrients from the foods we eat are absorbed in the digestive tract. Most of these travel in the bloodstream directly to the liver, where they are processed and released back into the bloodstream for delivery to body cells. Oxygen from the air we breathe diffuses into the blood, which moves from the lungs to the heart, which then pumps it out to the rest of the body. Moreover, endocrine glands scattered throughout the body release their products, called hormones, into the bloodstream, which carries them to distant target cells. Blood also picks up cellular wastes and by-products and transports them to various organs for removal. For instance, blood moves carbon dioxide to the lungs for exhalation from the body, and various other waste products are transported to the kidneys and liver for excretion from the body in the form of urine or bile.
Defence
Many types of WBCs protect the body from external threats, such as disease-causing bacteria that have entered the tissues. Other WBCs seek out and destroy internal threats, such as cells with mutated DNA that could multiply to become cancerous, or body cells infected with viruses.
When damage to the blood vessels results in bleeding, blood platelets and certain proteins dissolved in the plasma, the fluid portion of the blood, interact to patch the ruptured areas of the blood vessels involved. This helps to protect the body from further blood loss and initiates the healing process.
Maintenance of Homeostasis
Recall that body temperature is regulated via a classic negative-feedback loop. If you were exercising on a warm day, your rising core body temperature would trigger several homeostatic mechanisms, including increased transport of blood from your core to your body periphery, which is typically cooler. As blood passes through the vessels of the skin, heat would be dissipated to the environment, and the blood returning to your body core would be cooler. In contrast, on a cold day, blood is diverted away from the skin to maintain a warmer body core. In extreme cases, this may result in frostbite.
Blood also helps to maintain the chemical balance of the body. For example, proteins and other compounds in blood act as pH buffers, which thereby help to regulate the pH of body fluids and cells. Blood also helps to regulate the water content of body cells and tissues.
Composition of Blood
You may have had blood drawn from a superficial vein in your arm, which was then sent to a lab for analysis. Some of the most common blood tests—for instance, those measuring lipid or glucose levels in plasma—determine which substances are present within blood and in what concentrations. Other blood tests check for the composition of the blood itself, including the quantities and types of cellular components.
One such test, called a haematocrit, measures the percentage of RBCs, clinically known as erythrocytes, in a blood sample. It is typically performed by automated analysers but can be performed by spinning the blood sample in a specialised centrifuge, a process that causes the heavier elements suspended within the blood sample to separate from the lightweight, liquid plasma (Figure 5.1.1). Because the heaviest elements in blood are the erythrocytes, these settle at the very bottom of the haematocrit tube. Located above the erythrocytes is a pale, thin layer composed of the remaining cellular components of blood. These are the WBCs, clinically known as leukocytes, and the platelets, cell fragments also called thrombocytes. This layer is referred to as the buffy coat because of its buff colour; it normally constitutes less than 1 percent of a blood sample. Above the buffy coat is the blood plasma, normally a pale, straw-coloured fluid, which constitutes the remainder of the sample.
The volume of erythrocytes after centrifugation is also commonly referred to as packed cell volume (PCV). In normal blood, about 45 percent of a sample is erythrocytes. The haematocrit of any one sample can vary significantly, between about 36 and 50 percent, according to gender and other factors. Normal haematocrit values for females range from 37 to 47, with a mean value of 41; for males, haematocrit ranges from 42 to 52, with a mean of 47. The percentage of other cellular components, the WBCs and platelets, is extremely small so it is not normally considered with the haematocrit. The mean plasma percentage is the percent of blood that is not erythrocytes: for females, it is approximately 59 (or 100 minus 41), and for males, it is approximately 53 (or 100 minus 47).
Figure 5.1.1. Composition of blood. The cellular elements of blood include a vast number of erythrocytes and comparatively fewer leukocytes and platelets. Plasma is the fluid in which the cellular components are suspended. A sample of blood spun in a centrifuge reveals that plasma is the lightest component. It floats at the top of the tube separated from the heaviest elements, the erythrocytes, by a buffy coat of leukocytes (WBCs) and platelets. Haematocrit is the percentage of the total sample that is comprised of erythrocytes. Depressed and elevated haematocrit levels are shown for comparison.
Characteristics of Blood
When you think about blood, the first characteristic that probably comes to mind is its colour. Blood that has just taken up oxygen in the lungs is bright red, and blood that has released oxygen in the tissues is a darker red colour. This is because the major component of red blood cells, a protein called haemoglobin that binds oxygen, is a pigment that changes colour depending upon the degree of oxygen saturation. Blood is viscous and somewhat sticky to the touch. It has a viscosity approximately five times greater than water. Viscosity is a measure of a fluid’s thickness or resistance to flow and is influenced by the presence of the plasma proteins and cellular components within the blood. The viscosity of blood has a dramatic impact on blood pressure and flow. Consider the difference in flow between water and honey. The more viscous honey would demonstrate a greater resistance to flow than the less viscous water. The same principle applies to blood.
The normal temperature of blood is slightly higher than normal body temperature—about 38°C (or 100.4°F), compared to 37°C (or 98.6°F) for an internal body temperature reading, although daily variations of 0.5 °C are normal. Although the surface of blood vessels is relatively smooth, as blood flows through them, it experiences some friction and resistance, especially as vessels age and lose their elasticity, thereby producing heat. This accounts for its slightly higher temperature.
The pH of blood averages about 7.4; however, it can range from 7.35 to 7.45 in a healthy person. Blood is therefore somewhat more basic (alkaline) on a chemical scale than pure water, which has a pH of 7.0. Blood contains numerous buffers, including bicarbonate, haemoglobin and proteins, that help to regulate pH.
Blood constitutes approximately eight percent of adult body weight. Adult males typically average about five to six litres of blood. Adult females average four to five litres.
Blood Plasma
Like other fluids in the body, plasma is composed primarily of water. In fact, it is about 92 percent water. Dissolved or suspended within this water is a mixture of substances, most of which are proteins. Although there are hundreds of substances dissolved or suspended in the plasma, many of them are present only in very small quantities.
Plasma Proteins
About seven percent of the volume of plasma—nearly all that is not water—is made up of proteins. These include several plasma proteins (proteins that are unique to the plasma), plus a much smaller number of regulatory proteins, including enzymes and some hormones. The major components of plasma are summarised in Figure 5.1.2.
The three major groups of plasma proteins are as follows:
Albumin is the most abundant of the plasma proteins. Manufactured by the liver, albumin molecules serve as binding proteins—transport vehicles for fatty acids, calcium and steroid hormones. Recall that lipids are hydrophobic; however, their binding to albumin enables their transport in the watery plasma. Albumin is also the most significant contributor to the osmotic pressure of blood; that is, its presence holds water inside the blood vessels and draws water from the tissues, across blood vessel walls, and into the bloodstream. This in turn helps to maintain both blood volume and blood pressure. Albumin normally accounts for approximately 54 percent of the total plasma protein content, in clinical levels of 35–50 g/L blood.
The second most common plasma proteins are the globulins. A heterogeneous group, there are three main subgroups known as alpha, beta, and gamma globulins. The alpha and beta globulins transport iron, lipids, and the fat-soluble vitamins A, D, E, and K to the cells; like albumin, they also contribute to osmotic pressure. The gamma globulins are proteins involved in immunity and are better known as an antibodies or immunoglobulins. Although other plasma proteins are produced by the liver, immunoglobulins are produced by specialised leukocytes known as plasma cells. Antibodies assist in protection against infection by binding to invading microorganisms and marking them for destruction by specialised WBCs. Globulins make up approximately 38 percent of the total plasma protein volume, in clinical levels of 10–15 g/L blood.
The least abundant plasma protein is fibrinogen. Like albumin and the alpha and beta globulins, fibrinogen is produced by the liver. It is essential for blood clotting, a process described later in this chapter. Fibrinogen accounts for about 7 percent of the total plasma protein volume, in clinical levels of 2–4.5 g/L blood.
Other Plasma Solutes
In addition to proteins, plasma contains a wide variety of other substances. These include various electrolytes, such as sodium, potassium, and calcium ions; dissolved gases, such as oxygen, carbon dioxide, and nitrogen; various organic nutrients, such as vitamins, lipids, glucose, and amino acids; and metabolic wastes. All these nonprotein solutes combined contribute approximately 1 percent to the total volume of plasma.
Figure 5.1.2. Major blood components.
Career Connection
Phlebotomy and Medical Laboratory Science
Phlebotomists are medical professionals trained to draw blood (phleb- = “a blood vessel”; -tomy = “to cut”). When more than a few drops of blood are required, phlebotomists perform a venepuncture, typically of a surface vein in the arm. They perform a capillary stick on a finger, an earlobe, or the heel of an infant when only a small quantity of blood is required. An arterial stick is collected from an artery and used to analyse blood gases. After collection, the blood may be analysed by medical laboratories or perhaps used for transfusions, donations, or research. While many allied health professionals practice phlebotomy, the American Society of Phlebotomy Technicians issues certificates to individuals passing a national examination, and some large labs and hospitals hire individuals expressly for their skill in phlebotomy. In Australia, individuals are required to complete a Certificate III in Pathology Collection or undertake on-the-job training by experienced phlebotomists.
Medical or clinical laboratories employ a variety of individuals in technical positions:
Medical laboratory scientists, also known as clinical laboratory scientists, typically hold a bachelor degree. They perform a wide variety of tests on various body fluids, including blood. The information they provide is essential to the primary care providers in screening for disease and increased risk of disease, determining a diagnosis and in monitoring the course of a disease and response to treatment.
Medical laboratory technicians typically have an associate degree and perform duties similar to those of a medical laboratory scientist but with some restrictions on career advancement.
Most pathology laboratories also employ pathology assistants who are responsible for processing patient samples and carrying out routine tasks within the lab. Although clinical training is required, this is usually provided by pathology staff.
Section Review
Blood is a fluid connective tissue critical to the transportation of nutrients, gases, and wastes throughout the body; to defend the body against infection and other threats; and to the homeostatic regulation of pH, temperature, and other internal conditions. Blood is composed of cellular components—erythrocytes, leukocytes, and cell fragments called platelets—and a fluid extracellular matrix called plasma. More than 90 percent of plasma is water. The remainder is mostly plasma proteins—mainly albumin, globulins and fibrinogen—and other dissolved solutes such as glucose, lipids, electrolytes, hormones, metabolic waste products and dissolved gases. Because of the cellular components and the plasma proteins and other solutes, blood is sticky and more viscous than water. It is also slightly alkaline and its temperature is slightly higher than normal body temperature.
Trace the generation of the formed elements (cellular components) of blood from bone marrow stem cells
Discuss the role of haemopoietic growth factors in promoting the production of the formed elements
The lifespan of the formed elements is very brief. Although one type of leukocyte called memory cells can survive for years, most erythrocytes, leukocytes, and platelets normally live only a few hours to a few months. Thus, the body must form new blood cells and platelets quickly and continuously. When you donate a unit of blood during a blood drive (approximately 475 mL), your body typically replaces the donated plasma within 24 hours, but it takes about 4 to 6 weeks to replace the blood cells. This restricts the frequency with which donors can contribute their blood. The process by which this replacement occurs is called haemopoiesis, or haematopoiesis (from the Greek root haima- = “blood”; -poiesis = “production”).
Sites of Haemopoiesis
Prior to birth, haemopoiesis occurs in a number of tissues, beginning with the yolk sac of the developing embryo and continuing in the foetal liver, spleen, lymphatic tissue and eventually the red bone marrow. Following birth, most haemopoiesis occurs in the red marrow, a connective tissue within the spaces of spongy (cancellous) bone tissue. In children, haemopoiesis can occur in all bones including the medullary cavity of long bones; in adults, the process is largely restricted to the cranial and pelvic bones, the vertebrae, the sternum, and the proximal epiphyses of the femur and humerus.
Throughout adulthood, the liver and spleen maintain their ability to generate the formed elements. This process is referred to as extramedullary haemopoiesis (meaning haemopoiesis outside the medullary cavity of adult bones). When a disease such as bone cancer destroys the bone marrow, causing medullary haemopoiesis to fail, extramedullary haemopoiesis may be initiated.
Differentiation of Formed Elements from Stem Cells
All formed elements arise from stem cells of the red bone marrow. Recall that stem cells undergo mitosis plus cytokinesis (cellular division) to give rise to new daughter cells: One of these remains a stem cell and the other differentiates into one of any number of diverse cell types. Stem cells may be viewed as occupying a hierarchal system, with some loss of the ability to diversify at each step. The totipotent stem cell is the zygote, or fertilised egg. The totipotent (toti- = “all”) stem cell gives rise to all cells of the human body. The next level is the pluripotent stem cell, which gives rise to multiple types of cells of the body and some of the supporting foetal membranes. Beneath this level, the mesenchymal cell is a stem cell that develops only into types of connective tissue, including fibrous connective tissue, bone, cartilage, and blood, but not epithelium, muscle, and nervous tissue. One step lower on the hierarchy of stem cells is the haemopoietic stem cell, or haemocytoblast. All the formed elements of blood originate from this specific type of cell.
Haemopoiesis begins when the haemopoietic stem cell is exposed to appropriate chemical stimuli collectively called haemopoietic growth factors, which prompt it to divide and differentiate. One daughter cell remains a haemopoietic stem cell, allowing haemopoiesis to continue. The other daughter cell becomes either of two types of more specialised stem cells (Figure 5.2.1):
Lymphoid Stem Cells, also called Common Lymphoid Progenitor Cells, give rise to a class of leukocytes known as lymphocytes, which include the various T cells, B cells, and natural killer (NK) cells, all which function in immunity. However, haemopoiesis of lymphocytes progresses somewhat differently from the process for the other formed elements. In brief, lymphoid stem cells quickly migrate from the bone marrow to lymphatic tissues, including the lymph nodes, spleen, and thymus, where their production and differentiation continues. B cells are so named since they mature in the bone marrow, while T cells mature in the thymus.
Myeloid Stem Cells, also called Common Myeloid Progenitor Cells, give rise to all the other formed elements, including the erythrocytes; megakaryocytes that produce platelets; and a myeloblast lineage that gives rise to monocytes and three forms of granular leukocytes: neutrophils, eosinophils, and basophils.
Figure 5.2.1. Haematopoietic system of bone marrow. Haemopoiesis is the proliferation and differentiation of the formed elements of blood.
Lymphoid and myeloid stem cells do not immediately divide and differentiate into mature formed elements. There are several intermediate stages of precursor cells, many of which can be recognised by their names, which have the suffix -blast. For instance, megakaryoblasts are the precursors of megakaryocytes, and proerythroblasts become reticulocytes, having lost their nucleus and other organelles yet retaining residual ribosomal RNA, before maturing into erythrocytes as illustrated in Figure 5.2.1.
Haemopoietic Growth Factors
Development from stem cells to precursor cells to mature cells is again initiated and controlled by haemopoietic growth factors. These include the following:
Erythropoietin (EPO) is a glycoprotein hormone secreted by the interstitial fibroblast cells of the kidneys in response to low oxygen levels. It prompts the production of haemoglobin and erythrocytes. Some athletes use synthetic EPO as a performance-enhancing drug (called blood doping) to increase RBC counts haemoglobin concentration and subsequently increase oxygen delivery to tissues throughout the body. EPO is a banned substance in most organised sports, but it is also used medically in the treatment of certain types of anaemia, specifically those triggered by certain types of cancer, and other disorders in which increased erythrocyte counts and oxygen levels are desirable.
Thrombopoietin, another glycoprotein hormone, is produced by the liver and kidneys. It triggers the production of megakaryoblasts and the development of megakaryocytes into platelets.
Cytokines are glycoproteins secreted by a wide variety of cells, including red bone marrow, leukocytes, macrophages, fibroblasts, and endothelial cells. They act locally as autocrine or paracrine factors, stimulating the proliferation of progenitor cells and helping to stimulate both nonspecific and specific resistance to disease. There are two major subtypes of cytokines known as colony-stimulating factors and interleukins.
Colony-stimulating factors (CSFs) are glycoproteins that act locally, as autocrine or paracrine factors. Some trigger the differentiation of myeloblasts into granular leukocytes, namely, neutrophils, eosinophils, and basophils. These are referred to as granulocyte CSFs. A different CSF induces the production of monocytes, called monocyte CSF. Both granulocytes and monocytes are stimulated by GM-CSF; granulocytes, monocytes, platelets, and erythrocytes are stimulated by multi-CSF. Synthetic forms of these hormones are often administered to patients, with various forms of cancer who are receiving chemotherapy, to revive their WBC counts.
Interleukins are another class of cytokine signalling molecules important in haemopoiesis. They were initially thought to be secreted uniquely by leukocytes and to communicate only with other leukocytes, and were named accordingly, but are now known to be produced by a variety of cells including bone marrow and endothelium. Researchers now suspect that interleukins may play other roles in body functioning, including differentiation and maturation of cells, producing immunity and inflammation. To date, more than a dozen interleukins have been identified, with others likely to follow. They are generally numbered IL-1, IL-2, IL-3, etc.
Everyday Connection
Blood Doping
In its original intent, the term blood doping was used to describe the practice of injecting by transfusion supplemental RBCs into an individual, typically to enhance performance in a sport. Additional RBCs would deliver more oxygen to the tissues, providing extra aerobic capacity, clinically referred to as VO2 max. The source of the cells was either from the recipient (autologous) or from a donor with compatible blood (homologous). This practice was aided by the well-developed techniques of harvesting, concentrating, and freezing of the RBCs that could be later thawed and injected, yet still retain their functionality. These practices are considered illegal in virtually all sports and run the risk of causing infection, significantly increasing the viscosity of the blood and the potential for transmission of blood-borne pathogens if the blood was collected from another individual.
With the development of synthetic EPO in the 1980s, it became possible to provide additional RBCs by artificially stimulating RBC production in the bone marrow. Originally developed to treat patients suffering from anaemia due to renal failure or cancer treatment, large quantities of EPO can be generated by recombinant DNA technology. Synthetic EPO is injected under the skin and can increase haematocrit for many weeks. It may also induce polycythaemia and raise haematocrit to 70 or greater. This increased viscosity raises the resistance of the blood and forces the heart to pump more powerfully; in extreme cases, it has resulted in death. Other drugs such as cobalt II chloride have been shown to increase natural EPO gene expression. Blood doping has become problematic in many sports, especially cycling. Lance Armstrong, winner of seven Tour de France and many other cycling titles, was stripped of his victories and admitted to blood doping in 2013.
Bone Marrow Sampling and Transplants
Sometimes, a healthcare provider will order a bone marrow biopsy, a diagnostic test of a sample of red bone marrow. This involves inserting a large-bore needle into the region near the posterior iliac crest of the pelvic bones (os coxae) and collecting an aspirate of bone marrow fluid and a core biopsy of bone marrow matrix. These samples are required for the diagnosis and differentiation of various types of leukaemia and other diseases of bone marrow.
A bone marrow (haemopoietic stem cell) transplant is a treatment in which a donor’s healthy bone marrow—and its stem cells—replaces the faulty bone marrow of a patient suffering from diseases such as aplastic anaemia, thalassaemia, sickle cell anaemia and leukaemia. This type of transplant is called an allogeneic transplant. In many cases of leukaemia, the patient’s own haemopoietic stem cells can be harvested when the patient is in remission and transferred back to the patient if the leukaemia reoccurs. This type of transplant is called an autologous transplant.
In the past, when haemopoietic stem cells were collected from a bone marrow donor, the procedure involved harvesting the stem cells directly from the marrow as described above for bone marrow biopsy: a very invasive and painful procedure. These days, bone marrow donors are pre-treated with CSFs, which mobilise the stem cells from the bone marrow to the peripheral blood where they are easily collected via simple venepuncture. The stem cells are isolated from the other cellular components of the blood and then frozen for later use.
For an individual requiring a transplant, a donor with a matching tissue type is essential to prevent the immune system from destroying the transplanted donor cells—a phenomenon known as tissue rejection. To treat patients with haemopoietic stem cells, it is first necessary to destroy the patient’s own diseased marrow through radiation and/or chemotherapy. Donor bone marrow stem cells are then intravenously infused into the transplant recipient. From the bloodstream, they establish themselves in the recipient’s bone marrow and repopulate the marrow and the blood with healthy blood cells.
Section Review
Through the process of haemopoiesis, the formed elements of blood are continually produced, replacing the relatively short-lived erythrocytes, leukocytes, and platelets. Haemopoiesis begins in the red bone marrow, with haemopoietic stem cells that differentiate into myeloid and lymphoid lineages. Myeloid stem cells give rise to most of the formed elements. Lymphoid stem cells give rise only to the various lymphocytes designated as B and T cells, and NK cells. Haemopoietic growth factors, including erythropoietin, thrombopoietin, colony-stimulating factors, and interleukins, promote the proliferation and differentiation of formed elements.
Discuss the various steps in the lifecycle of an erythrocyte
Explain the composition and function of haemoglobin
The erythrocyte, commonly known as a red blood cell (or RBC), is by far the most common formed element: A single drop of blood contains millions of erythrocytes and just thousands of leukocytes. Specifically, males have about 5.4 million erythrocytes per microlitre (µL) of blood, and females have approximately 4.8 million per µL. In fact, erythrocytes are estimated to make up about 25 percent of the total cells in the body. As you can imagine, they are quite small cells, with a mean diameter of only about 7–8 micrometres (µm) (Figure 5.3.1). The primary functions of erythrocytes are to pick up inhaled oxygen from the lungs and transport it to the body’s tissues, and to pick up some (about 24 percent) carbon dioxide waste at the tissues and transport it to the lungs for exhalation. Erythrocytes remain within the vascular network. Although leukocytes typically leave the blood vessels to perform their defensive functions, movement of erythrocytes from the blood vessels is abnormal.
Figure 5.3.1. Summary of formed elements in blood.
Shape and Structure of Erythrocytes
As an erythrocyte matures in the red bone marrow, it extrudes its nucleus and most of its other organelles. During the first day or two that it is in the circulation, an immature erythrocyte, known as a reticulocyte, will still typically contain remnants of organelles. Reticulocytes should comprise approximately 1–2 percent of the erythrocyte count and provide a rough estimate of the rate of RBC production, with abnormally low or high rates indicating deviations in the production of these cells. These remnants, primarily of networks (reticulum) of ribosomes, are quickly shed, however, and mature, circulating erythrocytes have few internal cellular structural components. Lacking mitochondria, for example, they rely on anaerobic respiration. This means that they do not utilise any of the oxygen they are transporting, so they can deliver it all to the tissues. They also lack endoplasmic reticula and do not synthesise proteins. Erythrocytes do, however, contain some structural proteins that help the blood cells maintain their unique structure and enable them to change their shape to squeeze through capillaries. This includes the protein spectrin, a cytoskeletal protein element. They also contain enzymes that protect haemoglobin from oxidisation.
Erythrocytes are biconcave disks; that is, they are plump at their periphery and very thin in the centre (Figure 5.3.2). Since they lack most organelles, there is more interior space for the presence of the haemoglobin molecules that, as you will see shortly, transport gases. The biconcave shape also provides a greater surface area across which gas exchange can occur, relative to its volume; a sphere of a similar diameter would have a lower surface area-to-volume ratio. In the capillaries, the oxygen carried by the erythrocytes can diffuse into the plasma and then through the capillary walls to reach the cells, whereas some of the carbon dioxide produced by the cells as a waste product diffuses into the capillaries to be picked up by the erythrocytes. Capillary beds are extremely narrow, slowing the passage of the erythrocytes and providing an extended opportunity for gas exchange to occur. However, the space within capillaries can be so minute that, despite their own small size, erythrocytes may have to fold in on themselves if they are to make their way through. Fortunately, their structural proteins like spectrin are flexible, allowing them to bend over themselves to a surprising degree, then spring back again when they enter a wider vessel. In wider vessels, erythrocytes may stack up much like a roll of coins, forming a rouleaux, from the French word for “roll.”
Figure 5.3.2. Shape of red blood cells. a) Erythrocytes are biconcave discs with very shallow centres. This shape optimises the ratio of surface area to volume, facilitating gas exchange. It also enables them to fold up as they move through narrow blood vessels; b) An image from a peripheral blood smear showing normochromic, normocytic red blood cells. A small lymphocyte is present from comparison. 100x oil immersion. From MLS Collection, University of Alberta, https://doi.org/10.7939/R3W669Q69.
Haemoglobin
Haemoglobin is a large molecule made up of proteins and iron. It consists of four folded chains of a protein called globin, designated alpha 1 and 2, and beta 1 and 2 (Figure 5.3.3a). Each of these globin molecules is bound to a red pigment molecule called haem, which contains an ion of iron (Fe2+) (Figure 5.3.3b).
Figure 5.3.3. Haemoglobin. (a) A molecule of haemoglobin contains four globin proteins, each of which is bound to one molecule of the iron-containing pigment haem. (b) A single erythrocyte can contain 300 million haemoglobin molecules, and thus more than 1 billion oxygen molecules.
Each iron ion in the haem can bind to one oxygen molecule; therefore, each haemoglobin molecule can transport four oxygen molecules. An individual erythrocyte may contain about 300 million haemoglobin molecules and therefore can bind to and transport up to 1.2 billion oxygen molecules (see Figure 5.3.3b).
In the lungs, haemoglobin picks up oxygen, which binds to the iron ions, forming oxyhaemoglobin. The bright red, oxygenated haemoglobin travels to the body tissues, where it releases some of the oxygen molecules, becoming darker red deoxyhemoglobin, sometimes referred to as reduced haemoglobin. Oxygen release depends on the need for oxygen in the surrounding tissues, so haemoglobin rarely if ever leaves all its oxygen behind. In the capillaries, carbon dioxide enters the bloodstream. About 76 percent dissolves in the plasma, some of it remaining as dissolved CO2, and the remainder forming bicarbonate ion. About 23–24 percent of it binds to the amino acids in haemoglobin, forming a molecule known as carbaminohemoglobin. From the capillaries, the haemoglobin carries carbon dioxide back to the lungs, where it releases it for exchange of oxygen.
Changes in the levels of RBCs can have significant effects on the body’s ability to effectively deliver oxygen to the tissues. Ineffective haematopoiesis results in insufficient numbers of RBCs and results in one of several forms of anaemia. An overproduction of RBCs produces a condition called polycythaemia. The primary drawback with polycythaemia is not a failure to directly deliver enough oxygen to the tissues, but rather the increased viscosity of the blood, which makes it more difficult for the heart to circulate the blood and increases the risk of thrombosis (excessive blood clot formation).
In patients with insufficient haemoglobin, the tissues may not receive sufficient oxygen, resulting in another form of anaemia. In determining oxygenation of tissues, the value of greatest interest in healthcare is the percent saturation; that is, the percentage of haemoglobin sites occupied by oxygen in a patient’s blood. Clinically this value is commonly referred to simply as “percent sat.”
Percent saturation is normally monitored using a device known as a pulse oximeter, which is applied to a thin part of the body, typically the tip of the patient’s finger. The device works by sending two different wavelengths of light (one red, the other infrared) through the finger and measuring the light with a photodetector as it exits. Haemoglobin absorbs light differentially depending upon its saturation with oxygen. The machine calibrates the amount of light received by the photodetector against the amount absorbed by the partially oxygenated haemoglobin and presents the data as percent saturation. Normal pulse oximeter readings range from 95–100 percent. Lower percentages reflect hypoxaemia, or low blood oxygen. The term hypoxia is more generic and simply refers to low oxygen levels. Oxygen levels are also directly monitored from free oxygen in the plasma typically following an arterial stick. When this method is applied, the amount of oxygen present is expressed in terms of partial pressure of oxygen or simply pO2 and is typically recorded in units of millimetres of mercury, mm Hg.
The kidneys filter about 180 litres (~380 pints) of blood in an average adult each day and thus serve as ideal sites for receptors that determine oxygen saturation. In response to hypoxaemia, less oxygen will exit the vessels supplying the kidney, resulting in hypoxia (low oxygen concentration) in the tissue fluid of the kidney where oxygen concentration is monitored. Interstitial fibroblasts within the kidney secrete EPO, thereby increasing erythrocyte and haemoglobin production and restoring oxygen levels. In a classic negative-feedback loop, as oxygen saturation rises, EPO secretion falls, and vice versa, thereby maintaining homeostasis. Populations dwelling at high altitudes, with inherently lower levels of oxygen in the atmosphere, naturally maintain a haematocrit higher than people living at sea level. Consequently, people traveling to high altitudes may experience symptoms of hypoxaemia, such as fatigue, headache, and shortness of breath, for a few days after their arrival. In response to the hypoxaemia, the kidneys secrete EPO to step up the production of erythrocytes until homeostasis is achieved once again. To avoid the symptoms of hypoxaemia, or altitude sickness, mountain climbers typically rest for several days to a week or more at a series of camps situated at increasing elevations to allow EPO levels and, consequently, erythrocyte counts to rise. When climbing the tallest peaks, such as Mt. Everest and K2 in the Himalayas, many mountain climbers rely upon bottled oxygen as they near the summit.
Lifecycle of Erythrocytes
Production of erythrocytes in the marrow occurs at the staggering rate of more than 2 million cells per second. For this production to occur, several raw materials must be present in adequate amounts. These include the same nutrients that are essential to the production and maintenance of any cell, such as glucose, lipids, and amino acids. However, erythrocyte production also requires several micronutrients including trace elements and vitamins:
Iron. We have said that each haem group in a haemoglobin molecule contains an ion of the trace mineral iron. On average, less than 20 percent of the iron we consume is absorbed. Haem iron, from animal foods such as meat, poultry, and fish, is absorbed more efficiently than non-haem iron from plant foods. Upon absorption, iron becomes part of the body’s total iron pool. The bone marrow, liver, and spleen can store iron in the protein compounds ferritin and haemosiderin. Ferroprotein transports the iron across the intestinal cell plasma membranes and from its storage sites into tissue fluid where it enters the blood. When EPO stimulates the production of erythrocytes, iron is released from storage, bound to transferrin, and carried to the red marrow where it attaches to erythrocyte precursors.
Copper. A trace mineral, copper is a component of two plasma proteins, hephaestin and ceruloplasmin. Without these, haemoglobin could not be adequately produced. Located in intestinal villi, hephaestin enables iron to be absorbed by intestinal cells. Ceruloplasmin transports copper. Both enable the oxidation of iron from Fe2+to Fe3+, a form in which it can be bound to its transport protein, transferrin, for transport to body cells. In a state of copper deficiency, the transport of iron for haem synthesis decreases, and iron can accumulate in tissues, where it can eventually lead to organ damage.
Zinc. The trace mineral zinc functions as a co-enzyme that facilitates the synthesis of the haem portion of haemoglobin.
B vitamins. The B vitamins folate and vitamin B12 function as co-enzymes that facilitate DNA synthesis. Thus, both are critical for the synthesis of new cells, including erythrocytes.
Erythrocytes live up to 120 days in the circulation, after which the worn-out cells are removed by a type of myeloid phagocytic cell called a macrophage, located primarily within the bone marrow, liver, and spleen. The components of the degraded erythrocytes’ haemoglobin are further processed as follows:
Globin, the protein portion of haemoglobin, is broken down into amino acids, which can be sent back to the liver for construction of new proteins.
The iron contained in the haem portion of haemoglobin may be stored in the liver or spleen, primarily in the form of ferritin or hemosiderin, or carried through the bloodstream by transferrin to the red bone marrow for recycling into new erythrocytes.
The non-iron portion of haem is degraded into the waste product biliverdin, a green pigment, and then into another waste product, bilirubin, a yellow pigment. Bilirubin binds to albumin and travels in the blood to the liver, which excretes it in bile, a fluid released into the intestines to help emulsify dietary fats. In the large intestine, bacteria break the bilirubin apart from the bile and convert it to urobilinogen and then into stercobilin. It is then eliminated from the body in the faeces. Broad-spectrum antibiotics typically eliminate these “good” bacteria as well and this may alter the colour of faeces. The kidneys also remove any circulating bilirubin and other related metabolic by-products such as urobilins and secrete them into the urine.
The breakdown pigments formed from the destruction of haemoglobin can be seen in a variety of situations. At the site of an injury, biliverdin from damaged RBCs produces some of the dramatic colours associated with bruising. With a failing liver, bilirubin cannot be removed effectively from circulation and causes the body to assume a yellowish tinge associated with jaundice. Stercobilins within the faeces produce the typical brown colour associated with this waste and the yellow of urine is associated with the urobilins.
The erythrocyte lifecycle is summarised in Figure 5.3.4.
Figure 5.3.4. Erythrocyte lifecycle. Erythrocytes are produced in the bone marrow and sent into the circulation. At the end of their lifecycle, they are destroyed by macrophages in the spleen, and their components are recycled.
Disorders of Erythrocytes
The size, shape, and number of erythrocytes, and the number of haemoglobin molecules can have a major impact on a person’s health. When the number of RBCs or amount of haemoglobin is deficient, the general condition is called anaemia. There are more than 400 types of anaemia and approximately 4.5% of Australian adults are at risk of anaemia. Anaemia can be broken down into three major groups: those caused by blood loss, those caused by faulty or decreased RBC production, and those caused by excessive destruction of RBCs. Clinicians often use two groupings in diagnosis: The kinetic approach focuses on evaluating the production, destruction, and removal of RBCs, whereas the morphological approach examines the RBCs themselves, paying particular emphasis to their size. A common test is the mean cell volume (MCV), which measures RBC size. Normal-sized cells are referred to as normocytic, smaller-than-normal cells are referred to as microcytic, and larger-than-normal cells are referred to as macrocytic. Reticulocyte counts are also important and may reveal inadequate production of RBCs. The effects of the various anaemias are widespread, because reduced numbers of RBCs or haemoglobin will result in lower levels of oxygen being delivered to body tissues. Since oxygen is required for tissue functioning, anaemia produces fatigue, lethargy, and an increased risk for infection. An oxygen deficit in the brain impairs the ability to think clearly and may prompt headaches and irritability. Lack of oxygen leaves the patient short of breath, even as the heart and lungs work harder in response to the deficit.
Anaemias resulting from acute blood loss are usually easy to identify. In addition to bleeding from external wounds or other visible lesions, blood loss anaemias may be due to chronic bleeding from ulcers, haemorrhoids, inflammation of the stomach (gastritis) and some cancers of the gastrointestinal tract. The excessive use of aspirin or other nonsteroidal anti-inflammatory drugs such as ibuprofen can trigger ulceration and gastritis leading to chronic blood loss. Excessive menstruation and loss of blood during childbirth are also potential causes of anaemia.
Anaemias caused by faulty or decreased RBC production include sickle cell anaemia, iron deficiency, vitamin deficiency anaemia and diseases of the bone marrow and stem cells.
Figure 5.3.5. Sickle cells. Sickle cell anaemia is caused by a mutation in one of the haemoglobin genes. Erythrocytes produce an abnormal type of haemoglobin, which causes the cell to take on a sickle or crescent shape. (credit: Janice Haney Carr).
A characteristic change in the shape of erythrocytes is seen in sickle cell disease (also referred to as sickle cell anaemia). Sickle cell anaemia is caused by production of an abnormal type of haemoglobin, called haemoglobin S, which delivers less oxygen to tissues and causes erythrocytes to assume a sickle (or crescent) shape, especially at low oxygen concentrations (Figure 5.3.5). These abnormally shaped cells can then become lodged in narrow capillaries because they are unable to fold in on themselves to squeeze through, blocking blood flow to tissues and causing a variety of serious problems from painful joints to delayed growth and even blindness and cerebrovascular accidents (strokes). Sickle cell anaemia is a genetic or inherited blood disorder more commonly found in individuals of African, Middle Eastern, Asian, Indian and Mediterranean descent.
Iron deficiency anaemia is the most common type of anaemia and results when the amount of available iron is insufficient to allow production of sufficient haemoglobin. This condition can occur in individuals with a deficiency of iron in the diet and is especially common in teens and children as well as in vegans and vegetarians. Additionally, iron deficiency anaemia may be caused by either an inability to absorb and transport iron, or slow chronic bleeding usually from the gastrointestinal tract or through heavy menstruation.
Vitamin-deficient anaemias generally involve insufficient vitamin B12 and folate.
Megaloblastic anaemia is the result of a deficiency of vitamin B12 and/or folate, and often involves diets deficient in these essential nutrients. Lack of animal products or viable alternate sources in the diet can lead to a lack of vitamin B12 and overcooking or eating insufficient amounts of vegetables may lead to a lack of folate.
Pernicious anaemia is a type of megaloblastic anaemia caused by poor absorption of vitamin B12 and is often seen in patients with Crohn’s disease (a severe intestinal disorder often treated by surgery), surgical removal of the intestines or stomach (common in some weight loss surgeries), the presence of intestinal parasites and AIDS.
Pregnancies, some medications, excessive alcohol consumption, and some diseases such as coeliac disease are also associated with vitamin deficiencies. It is essential to provide sufficient folic acid during the early stages of pregnancy to reduce the risk of neurological defects, including spina bifida, a failure of the neural tube to close.
Assorted disease processes can also interfere with the production and formation of RBCs and haemoglobin. If myeloid stem cells are defective or replaced by cancer cells, there will be insufficient quantities of RBCs produced.
Aplastic anaemia is the condition in which there are deficient numbers of RBC stem cells. Aplastic anaemia is often inherited, or it may be triggered by radiation, medication, chemotherapy, or infection, particularly by Parvo virus B19.
Thalassaemia is an inherited condition typically occurring in individuals from the Middle East, the Mediterranean, Africa, and Southeast Asia, in which the lifespan of RBCs is shortened due to unbalanced production of globin chains.
Hereditary spherocytosis is another inherited form of anaemia associated with shortened RBC survival. In this case the spectrin in the RBC membrane is defective resulting in loss of membrane without loss of cell contents as the cells circulate through small capillaries. This change in the surface area to volume ratio causes the RBCs to lose their bi-concave disc shape and become spherical (spherocytes). Spherocytes are less flexible than normal RBC and become trapped in the spleen, where they are removed by splenic macrophages.
Lead exposure from industrial sources or even dust from paint chips of iron-containing paints or pottery that has not been properly glazed may also lead to destruction of the red marrow.
Various disease processes can also lead to anaemias. These include chronic kidney diseases often associated with decreased production of EPO, hypothyroidism, some forms of cancer, especially leukaemia and bone cancers, and autoimmune diseases such as lupus, and rheumatoid arthritis.
In contrast to anaemia, an elevated RBC count is called polycythaemia and is detected in a patient’s elevated haematocrit. It can occur transiently in a person who is dehydrated; when water intake is inadequate or water losses are excessive, the plasma volume falls. As a result, the haematocrit rises. For reasons mentioned earlier, a mild form of polycythaemia is chronic but normal in people living at high altitudes. Some elite athletes train at high altitudes specifically to induce this phenomenon. Finally, a type of bone marrow disease called polycythaemia vera (from the Greek vera = “true”) causes an excessive production of immature erythrocytes. Polycythaemia vera can dangerously elevate the viscosity of blood, raising blood pressure and making it more difficult for the heart to pump blood throughout the body. It is a relatively rare disease that occurs more often in men than women and is more likely to be present in people over 60 years of age.
Section Review
The most abundant formed elements in blood, erythrocytes, are red, biconcave disks packed with an oxygen-carrying compound called haemoglobin. The haemoglobin molecule contains four globin chains each bound to a pigment molecule called haem, which contains an ion of iron. In the bloodstream, iron picks up oxygen in the lungs and drops it off in the tissues; the amino acids in haemoglobin then transport carbon dioxide from the tissues back to the lungs. Erythrocytes live only 120 days on average, and thus must be continually replaced. Worn-out erythrocytes are phagocytosed by macrophages and their haemoglobin is broken down. The breakdown products are recycled or removed as wastes: Globin is broken down into amino acids for synthesis of new proteins; iron is stored in the liver or spleen or used by the bone marrow for production of new erythrocytes; and the remnants of haem are converted into bilirubin, or other waste products that are taken up by the liver and excreted in the bile or removed by the kidneys. Anaemia is a deficiency of RBCs and/or haemoglobin, whereas polycythaemia is an excess of RBCs.
Describe the general characteristics of leukocytes
Classify leukocytes according to their lineage, their main structural features and their primary functions
Discuss the most common malignancies involving leukocytes
Identify the lineage, basic structure and function of platelets
The leukocyte, commonly known as a white blood cell (or WBC), is a major component of the body’s defences against disease. Leukocytes protect the body against invading microorganisms and body cells with mutated DNA, and they clean up debris. Platelets are essential for the repair of damaged blood vessels; they also provide growth factors for healing and repair.
Characteristics of Leukocytes
Although leukocytes and erythrocytes both originate from hematopoietic stem cells in the bone marrow, they are very different from each other in many significant ways. For instance, leukocytes are far less numerous than erythrocytes: Typically, there are only 5000 to 10,000 per µL. They are also larger than erythrocytes and are the only formed elements that are complete cells, possessing a nucleus and organelles. And although there is just one type of erythrocyte, there are many types of leukocytes. Most of these types have a much shorter lifespan than that of erythrocytes, some as short as a few hours or even a few minutes in the case of cells responding to acute infection.
One of the most distinctive characteristics of leukocytes is their movement. Whereas erythrocytes spend their days circulating within the blood vessels, leukocytes routinely leave the bloodstream to perform their defensive functions in the body’s tissues. For leukocytes, the vascular network is simply a highway they travel and soon exit to reach their true destination. When they arrive, they are often given distinct names, such as macrophage or microglia, depending on their function. As shown in Figure 5.4.1, they leave the capillaries—the smallest blood vessels—or other small vessels through a process known as emigration (from the Latin for “removal”) or diapedesis (dia- = “through”; -pedan = “to leap”) or extravasation (extra = “outside”; vas = “vessel”) in which they squeeze through adjacent cells in a blood vessel wall.
Once they have exited the capillaries, some leukocytes will take up fixed positions in lymphatic tissue, bone marrow, the spleen, the thymus, liver or other organs. Others will move about through the tissue spaces very much like amoebas, continuously extending their plasma membranes, sometimes wandering freely, and sometimes moving toward the direction in which they are drawn by chemical signals. This attracting of leukocytes occurs because of positive chemotaxis (movement in response to chemicals), a phenomenon in which injured or infected cells and nearby leukocytes emit the equivalent of a chemical “000” (or “911” in U.S.A) call, attracting more leukocytes to the site.
Figure 5.4.1. Emigration. Leukocytes exit the blood vessel and then move through the connective tissue of the dermis toward the site of a wound. Some leukocytes, such as the eosinophil and neutrophil, are characterised as granular leukocytes. They release chemicals from their granules that destroy pathogens; they are also capable of phagocytosis. The monocyte, sometimes referred to as an agranular leukocyte, differentiates into a macrophage that then phagocytises the pathogens.
Classification of Leukocytes
When scientists first began to observe stained blood slides, it quickly became evident that leukocytes could be divided into two groups, according to whether their cytoplasm contained highly visible granules:
Granular leukocytes, or granulocytes, contain abundant granules within the cytoplasm. They include neutrophils, eosinophils, and basophils.
While granules are not totally lacking in agranular leukocytes, they are far fewer and less obvious on stained blood slides. Agranular leukocytes include monocytes, which mature into macrophages that are phagocytic, and lymphocytes, which arise from the lymphoid stem cell line.
Granular Leukocytes
We will consider the granular leukocytes in order from most common to least common. All of these are produced in the red bone marrow and have a short lifespan of hours to days. They typically have a lobed nucleus and are classified according to which type of stain best highlights their granules (Figure 5.4.2).
Figure 5.4.2. Granular leukocytes. A neutrophil has small granules that stain light lilac and a nucleus with two to five lobes. An eosinophil’s granules are slightly larger and stain reddish-orange, and its nucleus has two to three lobes. A basophil has large granules that stain dark blue to purple and a two-lobed nucleus that is often obscured by the granules.
The most common of all the leukocytes, neutrophils will normally comprise 50–70 percent of the total leukocyte count. They are 10–12 µm in diameter, significantly larger than erythrocytes. They are called neutrophils because their granules show up most clearly with stains that are chemically neutral (neither acidic nor basic). The granules are numerous but quite fine and normally appear light lilac. The nucleus has a distinct lobed appearance and may have two to five lobes, the number increasing with the age of the cell. Older neutrophils have increasing numbers of lobes and are often referred to as polymorphonuclear (a nucleus with many forms), or simply “polys.” Younger and immature neutrophils begin to develop lobes and are known as “bands.”
Neutrophils are rapid responders to the site of infection and are efficient phagocytes with a preference for bacteria. Their granules contain lysozyme, an enzyme capable of lysing, or breaking down, bacterial cell walls; oxidants such as hydrogen peroxide; and defensins, proteins that bind to and puncture bacterial and fungal plasma membranes, so that the cell contents leak out. Abnormally high counts of neutrophils indicate infection and/or inflammation, particularly triggered by bacteria, but are also found in burn patients and others experiencing unusual stress. A burn injury increases the proliferation of neutrophils in order to fight off infection that can result from the destruction of the barrier of the skin. Low counts may be caused by drug toxicity and other disorders, including leukaemia and bone marrow failure, and may increase an individual’s susceptibility to infection.
Eosinophils typically represent 2–4 percent of the total leukocyte count. They are also 10–12 µm in diameter. The granules of eosinophils stain best with an acidic stain known as eosin. The nucleus of the eosinophil will typically have two to three lobes and, if stained properly, the granules will have a distinct red to orange colour.
The granules of eosinophils contain antihistamine molecules, which counteract the activities of histamines, inflammatory chemicals produced by basophils and mast cells. Some eosinophil granules contain molecules toxic to parasitic worms, which can enter the body through the integument, or when an individual consumes raw or undercooked fish or meat. Eosinophils are also capable of phagocytosis and are particularly effective when antibodies bind to the target and form an antigen-antibody complex. High counts of eosinophils are associated with allergies, parasitic worm infestations, and some autoimmune diseases. Low counts may be due to drug toxicity and stress.
Basophils are the least common leukocytes, typically comprising less than one percent of the total leukocyte count. They are slightly smaller than neutrophils and eosinophils at 8–10 µm in diameter. The granules of basophils stain best with basic (alkaline) stains. Basophils contain large granules that pick up a dark blue stain and are so dark and numerous they may make it difficult to see the two-lobed nucleus.
In general, basophils intensify the inflammatory response. They share this trait with mast cells. In the past, mast cells were considered to be basophils that had left the circulation. However, this appears not to be the case, as the two cell types develop from different lineages.
The granules of basophils release histamines, which contribute to inflammation, and heparin, which opposes blood clotting. High counts of basophils are associated with allergies, parasitic infections, and hypothyroidism. Low counts are associated with pregnancy, stress, and hyperthyroidism.
Agranular Leukocytes
Agranular leukocytes contain smaller, less-visible granules in their cytoplasm than do granular leukocytes. The nucleus is simple in shape, sometimes with an indentation but without distinct lobes. There are two major types of agranulocytes: lymphocytes and monocytes.
Lymphocytes are the only formed element of blood that arises from lymphoid stem cells. Although they form initially in the bone marrow, much of their subsequent development and proliferation occurs in the lymphatic tissues. Lymphocytes are the second most common type of leukocyte, accounting for about 20–30 percent of all leukocytes, and are essential for the immune response. The size range of lymphocytes is quite extensive, with some authorities recognising two size classes and others three. Typically, the large cells are 10–14 µm and have a smaller nucleus-to-cytoplasm ratio and more obvious granules. The smaller cells, referred to as “small resting lymphocytes” or “naive lymphocytes” are typically 6–9 µm with a larger volume of nucleus to cytoplasm. A few cells may fall outside these ranges, at 14–17 µm. This finding has led to the three size range classification.
The three major groups of lymphocytes include natural killer cells, B cells, and T cells. Natural killer (NK) cells are capable of recognising cells that do not express “self” proteins on their plasma membrane or that contain foreign or abnormal markers. These “non-self” cells include cancer cells, cells infected with a virus, and other cells with atypical surface proteins. Thus, they provide generalised, nonspecific immunity. The larger more granular lymphocytes are typically NK cells.
B cells and T cells, also called B lymphocytes and T lymphocytes, play prominent roles in defending the body against specific pathogens (disease-causing microorganisms) and are involved in specific immunity. One form of B cells (plasma cells) produces the antibodies or immunoglobulins that bind to specific foreign or abnormal components of plasma membranes. These B cells and the antibodies they produce form part of the immune response referred to as humoral (body fluid) immunity. T cells provide cell-mediated immunity by physically attacking foreign or diseased cells. A memory cell is a variety of both B and T cells that develops after exposure to a pathogen and mounts rapid responses upon subsequent exposures to the same pathogen. Unlike other leukocytes, memory cells live for many years. B cells undergo a maturation process in the bone marrow, whereas T cells undergo maturation in the thymus. This site of the maturation process gives rise to the name B and T cells. The functions of lymphocytes are complex and will be covered in detail in the chapter covering the lymphatic system and immunity. Smaller lymphocytes are either B or T cells that have not yet encountered a foreign invader, although they cannot be differentiated in a normal blood smear.
Abnormally high lymphocyte counts are characteristic of viral infections and Bordetella pertussis (Whooping cough) as well as some types of cancer, particularly lymphomas. Abnormally low lymphocyte counts are characteristic of prolonged (chronic) illness or immunosuppression, including that caused by HIV infection and drug therapies that often involve steroids.
Monocytes originate from myeloid stem cells. They normally represent 2–8 percent of the total leukocyte count. They are typically easily recognised by their large size of 12–20 µm and indented or horseshoe-shaped nuclei. Macrophages are monocytes that have left the circulation and they phagocytose debris, foreign pathogens, aged or damaged erythrocytes, and many other dead, worn out, or damaged cells. Macrophages also release antimicrobial defensins and chemotactic chemicals that attract other leukocytes to the site of an infection or tissue damage. Some macrophages occupy fixed locations, whereas others wander through the tissue fluid.
Abnormally high counts of monocytes are associated with viral or fungal infections, tuberculosis, and some forms of leukaemia and other chronic diseases. Abnormally low counts are typically caused by suppression of the bone marrow. Figure 5.4.4 illustrates the five main types of circulating leukocytes. In clinical medicine, the differential counts of the types and percentages of leukocytes present in a blood sample are often key indicators in making a diagnosis and selecting a treatment.
Most leukocytes have a relatively short lifespan, typically measured in hours or days. Production of all leukocytes begins in the bone marrow under the influence of CSFs and interleukins. Secondary production and maturation of lymphocytes occurs in specific regions of lymphatic tissue known as germinal centres. Lymphocytes are fully capable of mitosis and may produce clones of cells with identical properties. This capacity enables an individual to maintain immunity throughout life to many threats that have been encountered in the past.
Disorders of Leukocytes
Leukopenia is a condition in which too few leukocytes are produced. Leukopenic individuals may be unable to ward off infectious diseases. Excessive leukocyte proliferation is known as leukocytosis. Leukocytosis is often an indicator of the presence of infection or inflammation.
Leukaemia is a cancer involving over production of abnormal leukocytes. It usually involves only one specific type of leukocyte from either the myeloid line (myeloid leukaemia) or the lymphoid line (lymphocytic leukaemia). In chronic leukaemia, mature leukocytes accumulate and fail to die. In acute leukaemia, there is an overproduction of abnormal immature leukocytes. Both conditions arise from a genetic mutation in a single cell and in both conditions the cells do not function properly.
Lymphoma is a form of cancer in which masses of malignant T and/or B lymphocytes collect in lymph nodes, the spleen, the liver and other tissues. As in leukaemia, the malignant leukocytes do not function properly, and the patient is vulnerable to infection. Some forms of lymphoma tend to progress slowly and respond well to treatment. Others tend to progress quickly and require aggressive treatment, without which they are rapidly fatal.
Platelets
You may occasionally see platelets referred to as thrombocytes, but because this name suggests they are a type of cell, it is not accurate. A platelet is not a cell but rather a fragment of the cytoplasm of a cell called a megakaryocyte that is surrounded by a plasma membrane. Megakaryocytes are descended from myeloid stem cells and are large, typically 50–100 µm in diameter, and contain an enlarged, multi-lobed nucleus. As noted earlier, thrombopoietin, a glycoprotein secreted by the kidneys and liver, stimulates the proliferation of megakaryoblasts, which mature into megakaryocytes. These remain within bone marrow tissue and ultimately form platelet-precursor extensions that extend through the walls of bone marrow capillaries to release into the circulation thousands of cytoplasmic fragments, each enclosed in a plasma membrane. These enclosed fragments are platelets. Each megakaryocyte releases 2000–3000 platelets during its lifespan. Following platelet release, megakaryocyte remnants, which are little more than a cell nucleus, are consumed by macrophages. The process of platelet production, thrombopoiesis, is summarised in figure 5.4.4.
Platelets are relatively small, 2–4 µm in diameter, but numerous, with typically 150,000–160,000 per µL of blood. After entering the circulation, approximately one-third migrate to the spleen for storage for later release in response to any rupture in a blood vessel. When a blood vessel is damaged, platelets become activated to perform their primary function, which is to limit blood loss. Platelets remain in the circulation for only about 10 days and are then phagocytosed by macrophages.
In addition to their critical role in haemostasis, the stoppage of blood loss following damage to a blood vessel. Platelets also secrete a variety of growth factors essential for growth and repair of tissue, particularly connective tissue. Infusions of concentrated platelets are now being used in some therapies to stimulate healing. Platelet rich plasma injections are also used by beauty therapists to “reinvigorate” the skin of their clients.
Disorders of Platelets
Thrombocytosis is a condition in which there are too many platelets. This may trigger formation of unwanted blood clots (thrombosis), a potentially fatal disorder. If there is an insufficient number of platelets, called thrombocytopenia, blood may not clot properly, and excessive bleeding may result. Inherited and acquired disorders of platelet function can also lead to excessive bleeding despite normal platelet numbers.
Figure 5.4.4. Platelets. Platelets are derived from cells called megakaryocytes.
Section Review
Leukocytes function in body defences. They squeeze out of the walls of blood vessels through emigration or diapedesis, then may move through connective tissue, lymphatic fluid or become residents in various organs where they fight against pathogenic organisms, diseased cells, or other threats to health. Granular leukocytes or granulocytes, which include neutrophils, eosinophils, and basophils, originate from myeloid stem cells, as do monocytes. The other so called agranular leukocytes, NK cells, B cells, and T cells, arise from the lymphoid stem cell line. The most abundant leukocytes are the neutrophils, which are first responders to infections, especially bacterial infections. About 20–30 percent of all leukocytes are lymphocytes, which are critical to the body’s defences against specific threats. Leukaemia and lymphoma are malignancies involving leukocytes. Platelets are fragments of cells known as megakaryocytes that dwell within the bone marrow. While many platelets are stored in the spleen, others enter the circulation and are essential for haemostasis; they also produce several growth factors important for repair and healing.
Describe the three mechanisms involved in haemostasis
Explain how the extrinsic and intrinsic coagulation pathways lead to the common pathway and the coagulation factors involved in each
Discuss disorders affecting haemostasis
Platelets are key players in haemostasis, the process by which the body seals a ruptured blood vessel and prevents further loss of blood. Although rupture of larger vessels usually requires medical intervention, haemostasis is quite effective in dealing with small, simple wounds and small internal bleeds. There are three steps to the process: vascular spasm, the formation of a platelet plug, and coagulation (blood clotting). Failure of any of these steps will result in haemorrhage—excessive bleeding. The vascular response and platelet plug formation are referred to as primary haemostasis and coagulation is referred to a secondary haemostasis.
Vascular Spasm
When a blood vessel is severed or punctured, or when the wall of a vessel is damaged, vascular spasm occurs. In vascular spasm, the smooth muscle in the walls of the vessel contracts dramatically. This smooth muscle has both circular layers; larger vessels also have longitudinal layers. The circular layers tend to constrict, which narrows the vessel lumen, and slows the flow of blood. The longitudinal layers, when present, draw the vessel back into the surrounding tissue, often making it more difficult for a surgeon to locate, clamp, and tie off a severed vessel. The vascular spasm response is believed to be triggered by several chemicals called endothelins that are released by vessel-lining (endothelial) cells and by pain receptors in response to vessel injury. This phenomenon typically lasts for up to 30 minutes, although it can last for hours.
Formation of the Platelet Plug
In the second step, platelets, which normally float free in the plasma, encounter exposed underlying connective tissue and collagenous fibres at the site of blood vessel damage. The platelets bind (adhesion) to the exposed collagen and become activated. Activated platelets are spiked and sticky and bind to other activated platelets (aggregation) and the endothelial lining. Platelet adhesion is assisted by a glycoprotein released from neighbouring endothelial cells called von Willebrand factor, which helps stabilise the growing platelet plug. Platelet aggregation is aided by the plasma protein, fibrinogen, which forms bridges between adjacent platelets at the site of bleed vessel damage. As platelets collect, they simultaneously release chemicals from their granules into the plasma that further contribute to haemostasis. Among the substances released by the platelets are:
adenosine diphosphate (ADP), which helps additional platelets to adhere to the injury site, reinforcing and expanding the platelet plug
serotonin, which maintains vasoconstriction
prostaglandins and phospholipids, which also maintain vasoconstriction and help to activate further clotting chemicals, as discussed next
Von Willebrand factor which aids in the adhesion of platelets to exposed collagen
A platelet plug can temporarily seal a small opening in a blood vessel. Plug formation, in essence, buys the body time while more sophisticated and durable repairs are being made. In a similar manner, even modern naval warships still carry an assortment of wooden plugs to temporarily repair small breaches in their hulls until permanent repairs can be made.
Coagulation
Those more sophisticated and more durable repairs are collectively called coagulation, the formation of a blood clot. The process is sometimes characterised as a cascade, because one event prompts the next as in a multi-level waterfall. The result is the production of a gelatinous but robust clot made up of a mesh of fibrin—an insoluble filamentous protein derived from fibrinogen, the plasma protein introduced earlier—in which platelets and blood cells are trapped. Figure 5.5.1 summarises the three steps of haemostasis.
Figure 5.5.1. Haemostasis. (a) An injury to a blood vessel initiates the process of haemostasis. Blood clotting involves three steps. First, vascular spasm slows the flow of blood. Next, a platelet plug forms to temporarily seal small openings in the vessel. Coagulation then enables the repair of the vessel wall once the leakage of blood has stopped. (b) The synthesis of fibrin in blood clots involves both an intrinsic pathway and an extrinsic pathway, both of which lead to a common pathway. (credit a: Kevin MacKenzie).
Clotting Factors Involved in Coagulation
In the coagulation cascade, plasma proteins called clotting factors (or coagulation factors) prompt reactions that activate still more coagulation factors. The process is complex, but is initiated along two basic pathways:
The extrinsic pathway, which is triggered when clotting factors come into contact with substances outside of the blood vessel.
The intrinsic pathway, which is triggered when clotting factors come into contact with substances inside the blood vessel.
Both of these pathways merge into a third pathway, referred to as the common pathway (see Figure 5.5.1b). All three pathways are dependent upon the 12 known clotting factors, including Ca2+ and vitamin K (Table 5.5.1). Clotting factors are secreted as inactive enzymes primarily by the liver and the platelets. The liver requires the fat-soluble vitamin K to produce many of them. Vitamin K (along with biotin and folate) is somewhat unusual among vitamins in that it is not only consumed in the diet but is also synthesised by bacteria residing in the large intestine. The calcium ion, considered factor IV, is derived from the diet and from the breakdown of bone. Some recent evidence indicates that activation of various clotting factors occurs on specific receptor sites on the surfaces of activated platelets.
The 12 clotting factors are numbered I through XIII according to the order of their discovery. Factor VI was once believed to be a distinct clotting factor, but is now thought to be identical to factor V. Rather than renumber the other factors, factor VI was allowed to remain as a placeholder and also a reminder that knowledge changes over time. Activated factors are denoted by the additions of the letter “a” after the factor number. For example, factor Va denotes activated factor five.
Table 5.5.1. Clotting factors
*Vitamin K Required
Factor number
Name
Type of molecule
Source
Pathway(s)
I
Fibrinogen
Plasma protein
Liver
Common; converted into fibrin
II
Prothrombin
Plasma protein
Liver*
Common; converted into thrombin
III
Tissue thromboplastin or tissue factor
Lipoprotein mixture
Damaged cells and platelets
Extrinsic
IV
Calcium ions
Inorganic ions in plasma
Diet, platelets, bone matrix
Entire process
V
Proaccelerin
Plasma protein
Liver, platelets
Extrinsic and intrinsic
VI
Not used
Not used
Not used
Not used
VII
Proconvertin
Plasma protein
Liver*
Extrinsic
VIII
Antihemophilic factor A
Plasma protein factor
Platelets and endothelial cells
Intrinsic; deficiency results in haemophilia A
IX
Antihemophilic factor B (plasma thromboplastin component)
Plasma protein
Liver*
Intrinsic; deficiency results in haemophilia B
X
Stuart-Power factor (thrombokinase)
Protein
Liver*
Extrinsic and intrinsic
XI
Plasma thromboplastin antecedent
Plasma protein
Liver
Intrinsic; deficiency results in haemophilia C
XII
Hageman factor
Plasma protein
Liver
Intrinsic; initiates clotting in vitro also activates plasmin
XIII
Fibrin-stabilising factor
Plasma protein
Liver, platelets
Stabilises fibrin; slows fibrinolysis
Extrinsic Pathway
The quicker responding and more direct extrinsic pathway (also known as the tissue factor pathway) begins when damage occurs to the surrounding tissues, such as in a traumatic injury. Damaged extravascular cells, which are extrinsic to the bloodstream, express factor III, also known as tissue factor or thromboplastin. When factor VII in the plasma comes into contact with exposed factor III, it becomes activated to factor VIIa. Factor VIII, Factor VIIa and Ca2+ then form an active enzyme complex. This enzyme complex leads to activation of factor X (Stuart–Prower Factor), which activates the common pathway discussed below. The events in the extrinsic pathway are completed in a matter of seconds.
Intrinsic Pathway
The intrinsic pathway (also known as the contact activation pathway) is longer and more complex. In this case, the factors involved are intrinsic to (present within) the bloodstream. The pathway can be prompted by damage to the tissues, resulting from internal factors such as arterial disease; however, it is most often initiated when factor XII (Hageman factor) comes into contact with negatively charged molecules, such as phospholipids expressed on the surface of activated platelets. Factor XII can also be activated by contact with foreign materials such as artificial heart valves and the surface of some bacteria. Factor XII is also activated when blood comes into contact with glass, in a test tube for example. Factor XIIa sets off a series of reactions that in turn activates factor XI (plasma thromboplastin antecedent) then factor IX (antihemophilic factor B or plasma thromboplasmin). In the meantime, substances released by the platelets increase the rate of these activation reactions. Finally, factor IXa binds to its cofactor, factor VIII (antihemophilic factor A) to form an enzyme complex that activates factor X (Stuart-Prower Factor or thrombokinase), leading to the common pathway. The events in the intrinsic pathway are completed in a few minutes.
Common Pathway
Both the intrinsic and extrinsic pathways lead to the common pathway, in which fibrin is produced to seal off the vessel. Once factor X has been activated to become the enzyme prothrombinase (factor Xa) by either the intrinsic or extrinsic pathway, it converts factor II, the inactive enzyme prothrombin, into the active enzyme thrombin. (Note that if the enzyme thrombin were not normally in an inactive form, clots would form spontaneously, a condition not consistent with life.) Then, thrombin converts factor I, the soluble plasma protein fibrinogen, into the insoluble protein fibrin protein strands. Fibrin molecules spontaneously join together to form a mesh, which is stabilised by cross-linking reactions catalysed by Factor XIIIa.
Fibrinolysis
The stabilised clot undergoes contraction via the action of contractile proteins within platelets. As these proteins contract, they pull on the fibrin threads, bringing the edges of the clot more tightly together, somewhat as we do when tightening loose shoelaces. This process also wrings out of the clot a small amount of fluid called serum, which is blood plasma without its clotting factors.
To restore normal blood flow as the vessel heals, the clot must eventually be removed. The process by which the clot is gradually degraded is called fibrinolysis. Like coagulation, fibrinolysis involves a fairly complicated series of protein catabolising reactions. During this process, the inactive protein plasminogen, released by endothelial cells around the site of blood vessel damage, is converted into the active enzyme plasmin, which gradually breaks down the fibrin of the clot. Additionally, bradykinin, a vasodilator, released from damaged tissues as a pain signal, reverses the effects of the serotonin and prostaglandins secreted by the platelets. This allows the smooth muscle in the walls of the vessels to relax and helps to restore the circulation.
Plasma Anticoagulants
An anticoagulant is any substance that opposes coagulation. Several circulating plasma anticoagulants play a role in limiting the coagulation process to the region of injury and restoring a normal, clot-free condition of blood. For instance, a cluster of proteins collectively referred to as the protein C system inactivates clotting factors involved in the intrinsic pathway. TFPI (tissue factor pathway inhibitor) inhibits the conversion of the inactive factor VII to the active form in the extrinsic pathway. Antithrombin inactivates factor X and opposes the conversion of prothrombin (factor II) to thrombin in the common pathway. And as noted earlier, basophils release heparin, a short-acting anticoagulant that also opposes prothrombin. Heparin is also found on the surfaces of cells lining the blood vessels. A pharmaceutical form of heparin is often administered therapeutically, for example, in surgical patients at risk for blood clots.
Disorders of Clotting
Either an insufficient or an excessive production of platelets can lead to severe bleeding on the hand or excessive platelet plug formation on the other. Both of these situations can be life-threatening. As discussed earlier, an insufficient number of platelets, called thrombocytopenia, typically results in the inability of blood to form clots. This can lead to excessive bleeding, even from minor wounds.
Another reason for failure of the blood to clot is the inadequate production of functional amounts of one or more clotting factors. This is the case in the genetic disorder haemophilia, which is actually a group of related disorders, the most common of which is haemophilia A, accounting for approximately 80 percent of cases. This disorder results in the inability to synthesise enough factor VIII. Haemophilia B is the second most common form, accounting for approximately 20 percent of cases. In this case, there is a deficiency of factor IX. Both of these defects are linked to the X chromosome and are typically passed from a healthy (carrier) mother to her male offspring, since males are XY. Females would need to inherit a defective gene from each parent to manifest the disease, since they are XX. Patients with haemophilia bleed from even minor internal and external wounds, and leak blood into joint spaces after exercise and into urine and stool. Haemophilia C is a rare condition that is triggered by an autosomal (not sex) chromosome defect that renders factor XI non-functional. It is not a true recessive condition, since even individuals with a single copy of the mutant gene show a tendency to bleed. Regular infusions of clotting factors isolated from healthy donors, or manufactured as recombinant proteins, can help prevent bleeding in haemophiliac patients. At some point, genetic therapy will become a viable option.
In contrast to the disorders characterised by thrombocytopenia is thrombocytosis, also mentioned earlier, a condition characterised by excessive numbers of platelets that increases the risk for excessive clot formation, a condition known as thrombosis. Inherited or acquired failures in the control or regulation of coagulation can also result in a tendency towards thrombosis: a condition referred to as thrombophilia or hypercoagulation. Deficiencies of protein C or antithrombin, for example, can predispose an individual to thrombosis. Acquired forms of thrombophilia include the autoimmune disease lupus, immune reactions to heparin, polycythaemia vera, sickle cell disease, pregnancy, and even obesity. A thrombus (plural = thrombi) is an aggregation of platelets, erythrocytes, and even WBCs typically trapped within a mass of fibrin strands. While the formation of a clot is normal following the haemostatic response to blood vessel damage just described, thrombi can form within an intact or only slightly damaged blood vessel. In a large vessel, a thrombus will adhere to the vessel wall and decrease the flow of blood and is referred to as a mural thrombus. In a small vessel, it may totally block the flow of blood and is termed an occlusive thrombus. Thrombi are most caused by vessel damage to the endothelial lining, which activates the clotting mechanism. These may include venous stasis, when blood in the veins, particularly in the legs, remains stationary for long periods. This is one of the dangers of long airplane flights in crowded conditions and may lead to deep vein thrombosis or atherosclerosis, an accumulation of fats, macrophages and cell debris in the walls of arteries. A thrombus can seriously impede blood flow to or from a region and will cause a local increase in blood pressure. If flow is to be maintained, the heart will need to generate a greater pressure to overcome the resistance.
When a portion of a thrombus breaks free from the vessel wall and enters the circulation, it is referred to as an embolus. An embolus that is carried through the bloodstream can be large enough to block a vessel critical to a major organ. When it becomes trapped, an embolus is called an embolism. In the heart, brain, or lungs, an embolism may accordingly cause a heart attack, a stroke, or a pulmonary embolism. These are medical emergencies.
Among the many known biochemical activities of aspirin is its role as an anticoagulant. Aspirin (acetylsalicylic acid) is very effective at inhibiting the aggregation of platelets. Physicians sometimes recommend that patients at risk for cardiovascular disease, which can lead to heart attack or stroke, take a low dose of aspirin on a daily basis as a preventive measure. However, aspirin can also lead to serious side effects, including increasing the risk of gastrointestinal ulcers. A patient is well advised to consult a physician before beginning any aspirin regimen.
A class of drugs collectively known as thrombolytic agents can help speed up the degradation of an abnormal clot. If a thrombolytic agent is administered to a patient within 3 hours following a thrombotic stroke, the patient’s prognosis improves significantly. However, some strokes are not caused by thrombi, but by haemorrhage. Thus, the cause must be determined before treatment begins. Tissue plasminogen activator is an enzyme that catalyses the conversion of plasminogen to plasmin, the primary enzyme that breaks down clots. It is released naturally by endothelial cells but is also used in clinical medicine. New research is progressing using compounds isolated from the venom of some species of snakes, particularly vipers and cobras, which may eventually have therapeutic value as thrombolytic agents. Streptokinase, an enzyme produced by some species of streptococci, has long been used to help degrade the thrombus in heart attack cases.
Section Review
Haemostasis is the physiological process by which bleeding ceases. Haemostasis involves three basic steps: vascular spasm, the formation of a platelet plug, and coagulation, by which clotting factors promote the formation of a fibrin clot. Fibrinolysis is the process in which a clot is degraded in a healing vessel. Anticoagulants are substances that oppose coagulation. They are important in limiting the extent and duration of clotting. Inadequate clotting can result from too few platelets, or inadequate production of clotting factors, for instance, in the genetic disorder haemophilia. Excessive clotting, called thrombosis, can be caused by excessive numbers of platelets or deficiencies in coagulation control factors. A thrombus is a collection of fibrin, platelets, and erythrocytes that has accumulated along the lining of a blood vessel, whereas an embolus is a thrombus that has broken free from the vessel wall and is circulating in the bloodstream.
Describe the two basic physiological consequences of transfusion of incompatible blood
Compare and contrast ABO and Rh blood groups
Identify which blood groups may be safely transfused into patients with different ABO types
Discuss the pathophysiology of haemolytic disease of the newborn
Blood transfusions in humans were risky procedures until the discovery of the major human blood groups by Karl Landsteiner, an Austrian biologist and physician, in 1900. Until that point, physicians did not understand why rapid death often followed blood transfusions. They assumed that all human blood was the same. Landsteiner discovered that the plasma of some individuals caused the red blood cells of some other individuals to clump together (agglutinate), and he surmised that fatal transfusion reaction occurred when the type of donor blood infused into the patient was incompatible with the patient’s own blood. Blood groups are determined by the presence or absence of specific marker molecules on the plasma membranes of erythrocytes. With the discovery of these inherited markers, it became possible for the first time to match patient-donor blood types and prevent severe haemolytic transfusion reactions and deaths.
Antigens, Antibodies and Transfusion Reactions
Antigens are substances that the body does not recognise as belonging to the “self” and that therefore trigger a defensive response from the leukocytes of the immune system. (Seek more content for additional information on immunity.) Here, we will focus on the role of antibody-mediated (humoral) immunity in blood transfusion reactions. With RBCs in particular, you may see the antigens referred to as isoantigens or agglutinogens (surface antigens) and the antibodies referred to as isoantibodies or agglutinins. In this chapter, we will use the more common terms antigens and antibodies.
Antigens are generally large proteins, but may include other classes of organic molecules, including carbohydrates, lipids, and nucleic acids. Following an infusion of incompatible blood, erythrocytes with foreign antigens appear in the bloodstream and trigger an immune response. Proteins called antibodies (immunoglobulins), which are produced by certain B lymphocytes called plasma cells, attach to the antigens on the plasma membranes of the infused erythrocytes. Large antibodies, called IgM antibodies, can cause direct destruction of incompatible RBC via activation of group of plasma proteins called complement. This type of RBC destruction is called intravascular haemolysis and the reaction is referred to as an acute haemolytic transfusion reaction, which can be fatal after infusion of as little as 10mL of incompatible RBC. Smaller antibodies, called IgG antibodies, bind to the incompatible RBC and mark them for destruction by macrophages in the spleen. This type of RBC destruction is called extravascular haemolysis and the reaction is called a delayed haemolytic transfusion reaction, which is not often fatal but leads to removal of the transfused RBC, effectively making the transfusion a waste of time.
During an acute haemolytic transfusion reaction haemoglobin is released into the bloodstream as a result of intravascular haemolysis. This haemoglobin travels to the kidneys, which are responsible for filtration of the blood. However, haemoglobin is toxic to endothelial cells in the kidney and the patient can quickly develop kidney failure. In addition, damage to these endothelial cells can trigger coagulation and acute inflammatory response that can lead to shock and death within minutes.
More than 50 antigens have been identified on erythrocyte membranes, but by far the most significant in terms of their potential harm to patients are the A and B antigens of the ABO blood group system discovered by Landsteiner. The next most clinically significant antigen is the D antigen of the Rhesus or Rh blood group system.
The ABO Blood Group
Although the ABO blood group name consists of three letters, ABO blood typing designates the presence or absence of just two antigens, A and B. Both are carbohydrate or saccharide antigens and their presence or absence on the RBC membrane is genetically determined. People who have A antigens only on their erythrocyte membrane surfaces are designated blood type A, and those whose erythrocytes have B antigens only are blood type B. People can also have both A and B antigens on their erythrocytes, in which case they are blood type AB. People with neither A nor B antigens are designated blood type O.
Normally the body must be exposed to a foreign antigen before an antibody can be produced. This is not the case for the ABO blood group. Individuals with type A blood—without any prior exposure to incompatible blood—have pre-formed or naturally occurring antibodies to the B antigen circulating in their blood plasma. These antibodies, referred to as anti-B antibodies, are IgM type antibodies and will cause agglutination and haemolysis if they ever encounter erythrocytes with B antigens. Similarly, an individual with type B blood has pre-formed anti-A antibodies. Individuals with type AB blood, which has both antigens, do not have pre-formed antibodies to either of these. People with type O blood lack antigens A and B on their erythrocytes, but both anti-A and anti-B antibodies circulate in their blood plasma. Anti-A and anti-B antibodies are not present at birth but develop during the first six months of life in response to A and B-like substances in the environment, especially of the surface of bacteria.
Rh Blood Groups
The Rhesus or Rh blood group is classified according to the presence or absence of a second erythrocyte antigen identified as Rh. (It was first discovered in a type of primate known as a rhesus macaque, which is often used in research, because its blood is like that of humans.) Although dozens of Rh antigens have been identified, the antigen, designated D, is the most clinically important. Those who have the Rh D antigen present on their erythrocytes—about 85 percent of Americans—are described as Rh positive (Rh+) and those who lack it are Rh negative (Rh−). Note that the Rh group is distinct from the ABO group, so any individual, no matter their ABO blood type, may or may not have Rh antigen on their RBCs. When identifying a patient’s blood type, the Rh group is designated by adding the word positive or negative to the ABO type. For example, A positive (A+) means ABO group A blood with the Rh(D) antigen present, and AB negative (AB−) means ABO group AB blood without the Rh(D) antigen.
Explore ABO and Rh blood type distribution by country and dependency (population averages) by looking at Table 5.6.2.
In contrast to the ABO group antibodies, which are preformed, antibodies to the Rh antigen are produced only in Rh− individuals after exposure to the antigen. This process, called sensitisation, occurs following a transfusion with Rh-incompatible blood or, more commonly, with the birth of an Rh+ baby to an Rh− mother. Problems are rare in a first pregnancy, since the baby’s Rh+ cells rarely cross the placenta (the organ of gas and nutrient exchange between the baby and the mother). However, during or immediately after birth, the Rh− mother can be exposed to the baby’s Rh+ cells (Figure 5.6.1). Research has shown that this occurs in about 13−14 percent of such pregnancies. After exposure, the mother’s immune system begins to generate anti-Rh (anti-D) antibodies. If the mother should then conceive another Rh+ baby, the Rh antibodies she has produced, which are small IgG antibodies, can cross the placenta into the foetal bloodstream and mark the foetal RBCs fir destruction by macrophages in the foetal spleen. This condition, known as haemolytic disease of the foetus and newborn (HDFN) or erythroblastosis foetalis, may cause anaemia in mild cases, but the haemolysis can be so severe that without treatment the foetus may die in the uterus or shortly after birth. An Rh- mother can also produce Rh antibodies after a miscarriage, termination of pregnancy or abdominal trauma during pregnancy with an Rh+ foetus.
Figure 5.6.1. Erythroblastosis foetalis. The first exposure of an Rh− mother to Rh+ erythrocytes during pregnancy induces sensitisation. Anti-Rh antibodies begin to circulate in the mother’s bloodstream. A second exposure occurs with a subsequent pregnancy with an Rh+ foetus in the uterus. Maternal anti-Rh antibodies may cross the placenta and enter the foetal bloodstream, causing agglutination and haemolysis of foetal erythrocytes.
Rh (D) immune globulin (Rh(D)Ig), derived from the plasma of women who have developed Rh antibodies, can temporarily prevent the development of Rh antibodies in the Rh− mother, thereby averting this potentially serious disease for the foetus. The anti-Rh antibodies, injected into the Rh- mother destroy any foetal Rh+ erythrocytes that may cross the placental barrier and enter the mother’s circulation. This effectively prevents sensitisation and the mother’s immune system does not begin to produce Rh antibodies. Rh(D)Ig is normally administered to Rh− mothers during weeks 26−28 of pregnancy, within 72 hours following birth of an Rh+ baby and following any potentially sensitive event. It has proven remarkably effective in decreasing the incidence of HDFN. Earlier we noted that the incidence of HDFN in an Rh+ subsequent pregnancy to an Rh− mother is about 13–14 percent without preventive treatment.
Determining ABO Blood Types
Laboratory technicians and scientists are able to determine a patient’s blood type quickly and easily using commercially prepared antibodies. A drop of the patient’s blood sample is placed into separate wells on a plastic tile or into separate glass test tubes. Into one well a drop of commercially prepared anti-A antibody is added and to another a drop of anti-B antibody is added. If the antigen is present, the antibodies will cause visible agglutination of the cells (Figure 5.6.2). The patient’s blood should also be tested for the Rh(D) antigen using commercially prepared anti-Rh(D) antibodies.
Figure 5.6.2. Determining blood types. This sample of a commercially produced “bedside” card enables quick typing of both a recipient’s and donor’s blood before transfusion. The card contains three reaction sites or wells. One is coated with an anti-A antibody, one with an anti-B antibody, and one with an anti-D antibody. Mixing a drop of blood and saline into each well enables the blood to interact with a preparation of type-specific antibodies, also called anti-sera. Agglutination of RBCs in a given site indicates a positive identification of the blood antigens, in this case A and Rh(D) antigens for blood type A+. For the purpose of transfusion, the donor’s and recipient’s blood types must be compatible: the donor’s RBC must not express antigens that will react with antibodies in the patient’s plasma.
ABO Transfusion Protocols
To avoid transfusion reactions, it is best to transfuse only matching blood types; that is, a type B+ recipient should ideally receive blood only from a type B+ donor and so on. That said, in emergency situations, when acute haemorrhage threatens the patient’s life, there may not be time for blood typing and cross matching. In these cases, blood from a universal RBC donor—an individual with type O− blood—may be transfused. Recall that type O erythrocytes do not display A or B antigens. Thus, anti-A or anti-B antibodies that might be circulating in the patient’s blood plasma will not encounter any erythrocyte surface antigens on the donated blood and therefore will not be provoked into a response. A patient with blood type AB+ is known as the universal RBC recipient. This patient can theoretically receive RBC of any type of blood, because the patient’s own plasma – having both A and B antigens on the erythrocyte surface – does not contain anti-A or anti-B antibodies. Figure 5.6.3 summarises the ABO blood types and their associated antigen and antibody profiles.
Note that type A RBC can be given to recipients with type A or type AB blood, because both cases, the recipient’s plasma does not contain anti-A antibodies. However, type AB RBC cannot be given to recipients with type A blood because in this case the patient’s plasma will contain anti-B antibodies, which will react with the B antigens on the donor blood.
While most transfusions involve infusion of RBC only, some patients require plasma transfusions. In the case of plasma transfusion, type AB is the universal donor because type AB plasma does not contain either anti-A or anti-B. The Rh(D) antigen also plays a role in blood transfusion. If Rh− individuals receiving blood have had prior exposure to Rh antigen, antibodies for this antigen may be present in the plasma and will trigger delayed haemolytic transfusion reaction and extravascular haemolysis. Although it is always preferable to type and cross match a patient’s blood before transfusing, in a true life-threatening emergency, this is not always possible, and type O- RBC or type AB plasma can be given until the patient’s blood type is established.
At the scene of multiple-vehicle accidents, military engagements, and natural or human-caused disasters, many victims may suffer simultaneously from acute haemorrhage, yet type O blood may not be immediately available. In these circumstances, medics may at least try to replace some of the volume of blood that has been lost. This is done by intravenous administration of a saline solution that provides fluids and electrolytes in proportions equivalent to those of normal blood plasma. Research is ongoing to develop a safe and effective artificial blood that would perform the oxygen-carrying function of blood without the RBCs, enabling transfusions in the field without concern for incompatibility.
Figure 5.6.3. ABO blood group. This chart summarises the characteristics of the blood types in the ABO blood group system. See the text for more on the concept of a universal donor or recipient.
Section Review
Antigens are non-self molecules, usually large proteins, which provoke an immune response. In transfusion reactions, antibodies attach to antigens on the surfaces of erythrocytes and cause agglutination and haemolysis. ABO blood group antigens are designated A and B. People with type A blood have A antigens on their erythrocytes, whereas those with type B blood have B antigens. Those with AB blood have both A and B antigens, and those with type O blood have neither A nor B antigens. The blood plasma contains preformed antibodies against the antigens not present on a person’s erythrocytes.
A second group of clinically important blood antigens is the Rh group, the most important of which is Rh D. People with Rh− blood do not have this antigen on their erythrocytes, whereas those who are Rh+ do. About 85 percent of Americans are Rh+. When a woman who is Rh− becomes pregnant with an Rh+ foetus, her body may begin to produce anti-Rh antibodies. If she subsequently becomes pregnant with a second Rh+ foetus and has not been treated preventively with Rh(D)Ig, the foetus will be at risk for haemolytic disease of the foetus and newborn.
Agglutination testing to determine blood type is necessary before transfusing blood, unless the patient is experiencing haemorrhage that is an immediate threat to life, in which case type O− red blood cells or type AB plasma may be transfused.
Describe the location and position of the heart within the body cavity
Describe the internal and external anatomy of the heart
Identify the tissue layers of the heart
Relate the structure of the heart to its function as a pump
Compare systemic circulation to pulmonary circulation
Identify the veins and arteries of the coronary circulation system
Trace the pathway of oxygenated and deoxygenated blood through the chambers of the heart
The vital importance of the heart is obvious. If one assumes an average rate of contraction of 75 contractions per minute, a human heart would contract approximately 108,000 times in one day, more than 39 million times in one year and nearly 3 billion times during a 75-year lifespan! Each of the major pumping chambers of the heart ejects approximately 70 mL blood per contraction in a resting adult. This would be equal to 5.25 litres of fluid per minute and approximately 14,000 litres per day. Over one year, that would equal 10,000,000 litres (2.6 million gallons) of blood sent through roughly 97,000 kilometres (60,000 miles) of vessels. In order to understand how that happens, it is necessary to understand the anatomy and physiology of the heart.
Location of the Heart
The human heart is located within the thoracic cavity, medially between the lungs in the space known as the mediastinum. Figure 6.1.1 shows the position of the heart within the thoracic cavity. Within the mediastinum, the heart is separated from the other mediastinal structures by a tough membrane known as the pericardium, or pericardial sac, and sits in its own space called the pericardial cavity. The dorsal surface of the heart lies near the bodies of the vertebrae, and its anterior surface sits deep to the sternum and costal cartilages. The great veins, the superior and inferior venae cavae, and the great arteries, the aorta and pulmonary trunk, are attached to the superior surface of the heart, called the base. The base of the heart is located at the level of the third costal cartilage, as seen in Figure 6.1.1. The inferior tip of the heart, the apex, lies just to the left of the sternum between the junction of the fourth and fifth ribs near their articulation with the costal cartilages. The right side of the heart is deflected anteriorly, and the left side is deflected posteriorly. It is important to remember the position and orientation of the heart when placing a stethoscope on the chest of a patient and listening for heart sounds, and when looking at images taken from a midsagittal perspective. The slight deviation of the apex to the left is reflected in a depression in the medial surface of the inferior lobe of the left lung, called the cardiac notch.
Figure 6.1.1. Position of the heart in the thorax. The heart is located within the thoracic cavity, medially between the lungs in the mediastinum. It is about the size of a fist, is broad at the top, and tapers toward the base.
Everyday Connection
CPR
The position of the heart in the torso between the vertebrae and sternum (see Figure 6.1.1 for the position of the heart within the thorax) allows for individuals to apply an emergency technique known as cardiopulmonary resuscitation (CPR) if the heart of a patient should stop. In adults, pressure is applied with the flat portion of one hand on the sternum in the area between the line at T4 and T9 (Figure 6.1.2), it is possible to manually compress the blood within the heart enough to push some of the blood within it into the pulmonary and systemic circuits. This is particularly critical for the brain, as irreversible damage and death of neurons occur within minutes of loss of blood flow. Current standards call for compression of the chest at least 5 cm deep and at a rate of 100-120 compressions per minute, a rate equal to the beat in “Staying Alive,” recorded in 1977 by the Bee Gees. If you are unfamiliar with this song, a version is available on www.youtube.com. At this stage, the emphasis is on performing high-quality chest compressions, rather than providing artificial respiration, but typically compression to ventilation ratio is 30:2. CPR is generally performed until the patient regains spontaneous contraction or is declared dead by an experienced healthcare professional.
When performed by untrained or overzealous individuals, CPR can result in broken ribs or a broken sternum and can inflict additional severe damage on the patient. It is also possible, if the hands are placed too low on the sternum, to manually drive the xiphoid process into the liver, a consequence that may prove fatal for the patient. Proper training is essential. This proven life-sustaining technique is so valuable that virtually all medical personnel as well as concerned members of the public should be certified and routinely recertified in its application. CPR courses are offered at a variety of locations, including colleges, hospitals, the Australian Red Cross and some commercial companies. They normally include practice of the compression technique on a mannequin.
Figure 6.1.2. CPR technique. If the heart should stop, CPR can maintain the flow of blood until the heart resumes beating. By applying pressure to the sternum, the blood within the heart will be squeezed out of the heart and into the circulation. Proper positioning of the hands on the sternum to perform CPR would be between the lines at T4 and T9.
Shape and Size of the Heart
The shape of the heart is similar to a pinecone, rather broad at the superior surface and tapering to the apex (see Figure 6.1.1). A typical heart is approximately the size of your fist: 12 cm (5 in) in length, 8 cm (3.5 in) wide, and 6 cm (2.5 in) in thickness. Given the size difference between most members of the sexes, the weight of a female heart is approximately 250–300 grams (9 to 11 ounces), and the weight of a male heart is approximately 300–350 grams (11 to 12 ounces). The heart of a well-trained athlete, especially one specialising in aerobic sports, can be considerably larger than this. Cardiac muscle responds to exercise in a manner like that of skeletal muscle. That is, exercise results in the addition of protein myofilaments that increase the size of the individual cells without increasing their numbers, a concept called hypertrophy. Hearts of athletes can pump blood more effectively at lower rates than those of nonathletes. Enlarged hearts are not always a result of exercise; they can result from pathologies, such as hypertrophic cardiomyopathy. The cause of an abnormally enlarged heart muscle is unknown, but the condition is often undiagnosed and can cause sudden death in apparently otherwise healthy young people.
Chambers and Circulation Through the Heart
The human heart consists of four chambers: The left side and the right side each have one atrium and one ventricle. Each of the upper chambers, the right atrium (plural = atria) and the left atrium, acts as a receiving chamber and contracts to push blood into the lower chambers, the right ventricle and the left ventricle. The ventricles serve as the primary pumping chambers of the heart, propelling blood to the lungs or to the rest of the body.
There are two distinct but linked circuits in the human circulation called the pulmonary and systemic circuits. Although both circuits transport blood and everything it carries, we can initially view the circuits from the point of view of gases. The pulmonary circuit transports blood to and from the lungs, where it picks up oxygen and delivers carbon dioxide for exhalation. The systemic circuit transports oxygenated blood to virtually all the tissues of the body and returns relatively deoxygenated blood and carbon dioxide to the heart to be sent back to the pulmonary circulation.
The right ventricle pumps deoxygenated blood into the pulmonary trunk, which leads toward the lungs and bifurcates into the left and right pulmonary arteries. These vessels in turn branch many times before reaching the pulmonary capillaries, where gas exchange occurs: Carbon dioxide exits the blood and oxygen enters. The pulmonary trunk arteries and their branches are the only arteries in the post-natal body that carry relatively deoxygenated blood. Highly oxygenated blood returning from the pulmonary capillaries in the lungs passes through a series of vessels that join together to form the pulmonary veins—the only post-natal veins in the body that carry highly oxygenated blood. The pulmonary veins conduct blood into the left atrium, which pumps the blood into the left ventricle, which in turn pumps oxygenated blood into the aorta and on to the many branches of the systemic circuit. Eventually, these vessels will lead to the systemic capillaries, where exchange with the tissue fluid and cells of the body occurs. In this case, oxygen and nutrients exit the systemic capillaries to be used by the cells in their metabolic processes, and carbon dioxide and waste products will enter the blood.
The blood exiting the systemic capillaries is lower in oxygen concentration than when it entered. The capillaries will ultimately unite to form venules, joining to form ever-larger veins, eventually flowing into the two major systemic veins, the superior vena cava and the inferior vena cava, which return blood to the right atrium. The blood in the superior and inferior venae cavae flows into the right atrium, which pumps blood into the right ventricle. This process of blood circulation continues as long as the individual remains alive. Understanding the flow of blood through the pulmonary and systemic circuits is critical to all health professions (Figure 6.1.3).
Figure 6.1.3. Dual system of the human blood circulation. Blood flows from the right atrium to the right ventricle, where it is pumped into the pulmonary circuit. The blood in the pulmonary artery branches is low in oxygen but relatively high in carbon dioxide. Gas exchange occurs in the pulmonary capillaries (oxygen into the blood, carbon dioxide out), and blood high in oxygen and low in carbon dioxide is returned to the left atrium. From here, blood enters the left ventricle, which pumps it into the systemic circuit. Following exchange in the systemic capillaries (oxygen and nutrients out of the capillaries and carbon dioxide and wastes in), blood returns to the right atrium and the cycle is repeated.
Membranes, Surface Features and Layers
Our exploration of more in-depth heart structures begins by examining the membrane that surrounds the heart, the prominent surface features of the heart and the layers that form the wall of the heart. Each of these components plays its own unique role in terms of function.
Membranes
The membrane that directly surrounds the heart and defines the pericardial cavity is called the pericardium or pericardial sac. It also surrounds the “roots” of the major vessels, or the areas of closest proximity to the heart. The pericardium, which literally translates as “around the heart,” consists of two distinct sublayers: the sturdy outer fibrous pericardium and the inner serous pericardium. The fibrous pericardium is made of tough, dense connective tissue that protects the heart and maintains its position in the thorax. The more delicate serous pericardium consists of two layers: the parietal pericardium, which is fused to the fibrous pericardium, and an inner visceral pericardium, or epicardium, which is fused to the heart and is part of the heart wall. The pericardial cavity, filled with lubricating serous fluid, lies between the epicardium and the pericardium.
In most organs within the body, visceral serous membranes such as the epicardium are microscopic. However, in the case of the heart, it is not a microscopic layer but rather a macroscopic layer, consisting of a simple squamous epithelium called a mesothelium, reinforced with loose, irregular, or areolar connective tissue that attaches to the pericardium. This mesothelium secretes the lubricating serous fluid that fills the pericardial cavity and reduces friction as the heart contracts. Figure 6.1.4 illustrates the pericardial membranes and the layers of the heart.
Figure 6.1.4. Pericardial membranes and layers of the heart wall. The pericardial membranes that surrounds the heart consists of three layers and the pericardial cavity. The heart wall also consists of three layers. The pericardial membrane and the heart wall share the epicardium.
Disorders of the Heart: Cardiac Tamponade
If excess fluid builds within the pericardial space, it can lead to a condition called cardiac tamponade, or pericardial tamponade. With each contraction of the heart, more fluid—in most instances, blood—accumulates within the pericardial cavity. In order to fill with blood for the next contraction, the heart must relax. However, the excess fluid in the pericardial cavity puts pressure on the heart and prevents full relaxation, so the chambers within the heart contain slightly less blood as they begin each heart cycle. Over time, less and less blood is ejected from the heart. If the fluid builds up slowly, as in hypothyroidism, the pericardial cavity may be able to expand gradually to accommodate this extra volume. Some cases of fluid in excess of one litre within the pericardial cavity have been reported. Rapid accumulation of as little as 100 mL of fluid following trauma may trigger cardiac tamponade. Other common causes include myocardial rupture, pericarditis, cancer, or even cardiac surgery. Removal of this excess fluid requires insertion of drainage tubes into the pericardial cavity. Premature removal of these drainage tubes, for example, following cardiac surgery, or clot formation within these tubes are causes of this condition. Untreated, cardiac tamponade can lead to death.
Surface Features of the Heart
Inside the pericardium, the surface features of the heart are visible, including the four chambers. There is a superficial leaf-like extension of the atria near the superior surface of the heart, one on each side, called an auricle—a name that means “ear like”—because its shape resembles the external ear of a human (Figure 6.1.5). Auricles are relatively thin-walled structures that can fill with blood and empty into the atria or upper chambers of the heart. You may also hear them referred to as atrial appendages. Also prominent is a series of fat-filled grooves, each of which is known as a sulcus (plural = sulci), along the superior surfaces of the heart. Major coronary blood vessels are located in these sulci. The deep coronary sulcus is located between the atria and ventricles. Located between the left and right ventricles are two additional sulci that are not as deep as the coronary sulcus. The anterior interventricular sulcus is visible on the anterior surface of the heart, whereas the posterior interventricular sulcus is visible on the posterior surface of the heart. Figure 6.1.5 illustrates anterior and posterior views of the surface of the heart.
Figure 6.1.5. External anatomy of the heart. Inside the pericardium, the surface features of the heart are visible.
Layers
The wall of the heart is composed of three layers of unequal thickness. From superficial to deep, these are the epicardium, the myocardium and the endocardium (see Figure 6.1.4). The outermost layer of the wall of the heart is also the innermost layer of the pericardium, the epicardium, or the visceral pericardium discussed earlier.
The middle and thickest layer is the myocardium, made largely of cardiac muscle cells. It is built upon a framework of collagenous fibres, plus the blood vessels that supply the myocardium and the nerve fibres that help regulate the heart. It is the contraction of the myocardium that pumps blood through the heart and into the major arteries. The muscle pattern is elegant and complex, as the muscle cells swirl and spiral around the chambers of the heart. They form a figure 8 pattern around the atria and around the bases of the great vessels. Deeper ventricular muscles also form a figure 8 around the two ventricles and proceed toward the apex. More superficial layers of ventricular muscle wrap around both ventricles. This complex swirling pattern allows the heart to pump blood more effectively than a simple linear pattern would. Figure 6.1.6 illustrates the arrangement of muscle cells.
Figure 6.1.6. Heart musculature. The swirling pattern of cardiac muscle tissue contributes significantly to the heart’s ability to pump blood effectively.
Although the ventricles on the right and left sides pump the same amount of blood per contraction, the muscle of the left ventricle is much thicker and better developed than that of the right ventricle. In order to overcome the high resistance required to pump blood into the long systemic circuit, the left ventricle must generate a great amount of pressure. The right ventricle does not need to generate as much pressure, since the pulmonary circuit is shorter and provides less resistance. Figure 6.1.7 illustrates the differences in muscular thickness needed for each of the ventricles.
Figure 6.1.7. Differences in ventricular muscle thickness. The myocardium in the left ventricle is significantly thicker than that of the right ventricle. Both ventricles pump the same amount of blood, but the left ventricle must generate a much greater pressure to overcome greater resistance in the systemic circuit. The ventricles are shown in both relaxed and contracting states. Note the differences in the relative size of the lumens, the region inside each ventricle where the blood is contained.
The innermost layer of the heart wall, the endocardium, is joined to the myocardium with a thin layer of connective tissue. The endocardium lines the chambers where the blood circulates and covers the heart valves. It is made of simple squamous epithelium called endothelium, which is continuous with the endothelial lining of the blood vessels (see Figure 6.1.4).
Once regarded as a simple lining layer, recent evidence indicates that the endothelium of the endocardium and the coronary capillaries may play active roles in regulating the contraction of the muscle within the myocardium. The endothelium may also regulate the growth patterns of the cardiac muscle cells throughout life, and the endothelins it secretes create an environment in the surrounding tissue fluids that regulates ionic concentrations and states of contractility. Endothelins are potent vasoconstrictors and, in a normal individual, establish a homeostatic balance with other vasoconstrictors and vasodilators.
Internal Structure of the Heart
Recall that the heart’s contraction cycle follows a dual pattern of circulation—the pulmonary and systemic circuits—because of the pairs of chambers that pump blood into the circulation. In order to develop a more precise understanding of cardiac function, it is first necessary to explore the internal anatomical structures in more detail.
Septa of the Heart
The word septum is derived from the Latin for “something that encloses;” in this case, a septum (plural = septa) refers to a wall or partition that divides the heart into chambers. The septa are physical extensions of the myocardium lined with endocardium. Located between the two atria is the interatrial septum. Normally in an adult heart, the interatrial septum bears an oval-shaped depression known as the fossa ovalis, a remnant of an opening in the foetal heart known as the foramen ovale. The foramen ovale allowed blood in the foetal heart to pass directly from the right atrium to the left atrium, allowing some blood to bypass the pulmonary circuit. Within seconds after birth, a flap of tissue known as the septum primum that previously acted as a valve closes the foramen ovale and establishes the typical cardiac circulation pattern.
Between the two ventricles is a second septum known as the interventricular septum. Unlike the interatrial septum, the interventricular septum is normally intact after its formation during foetal development. It is substantially thicker than the interatrial septum, since the ventricles generate far greater pressure when they contract.
The septum between the atria and ventricles is known as the atrioventricular septum. It is marked by the presence of four openings that allow blood to move from the atria into the ventricles and from the ventricles into the pulmonary trunk and aorta. Located in each of these openings between the atria and ventricles is a valve, a specialised structure that ensures one-way flow of blood. The valves between the atria and ventricles are known generically as atrioventricular valves. The valves at the openings that lead to the pulmonary trunk and aorta are known generically as semilunar valves. The interventricular septum is visible in Figure 6.1.8. In this figure, the atrioventricular septum has been removed to better show the bicuspid and tricuspid valves; the interatrial septum is not visible, since its location is covered by the aorta and pulmonary trunk. Since these openings and valves structurally weaken the atrioventricular septum, the remaining tissue is heavily reinforced with dense connective tissue called the cardiac skeleton, or skeleton of the heart. It includes four rings that surround the openings between the atria and ventricles, and the openings to the pulmonary trunk and aorta, and serve as the point of attachment for the heart valves. The cardiac skeleton also provides an important boundary in the heart electrical conduction system.
Figure 6.1.8. Internal structures of the heart. This anterior view of the heart shows the four chambers, the major vessels and their early branches, as well as the valves. The presence of the pulmonary trunk and aorta covers the interatrial septum, and the atrioventricular septum is cut away to show the atrioventricular valves.
Disorders of the Heart: Heart Defects
One very common form of interatrial septum pathology is patent foramen ovale, which occurs when the septum primum does not close at birth, and the fossa ovalis is unable to fuse. The word patent is from the Latin root patens for “open.” It may be benign or asymptomatic, perhaps never being diagnosed, or in extreme cases, it may require surgical repair to close the opening permanently. As much as 20–25 percent of the general population may have a patent foramen ovale, but fortunately, most have the benign, asymptomatic version. Patent foramen ovale is normally detected by auscultation of a heart murmur (an abnormal heart sound) and confirmed by imaging with an echocardiogram. Despite its prevalence in the general population, the causes of patent ovale are unknown, and there are no known risk factors. In nonlife-threatening cases, it is better to monitor the condition than to risk heart surgery to repair and seal the opening.
Coarctation of the aorta is a congenital abnormal narrowing of the aorta that is normally located at the insertion of the ligamentum arteriosum, the remnant of the foetal shunt called the ductus arteriosus. If severe, this condition drastically restricts blood flow through the primary systemic artery, which is life threatening. In some individuals, the condition may be benign and not detected until later in life. Detectable symptoms in an infant include difficulty breathing, poor appetite, trouble feeding, or failure to thrive. In older individuals, symptoms include dizziness, fainting, shortness of breath, chest pain, fatigue, headache, and nosebleeds. Treatment involves surgery to resect (remove) the affected region or angioplasty to open the abnormally narrow passageway. Studies have shown that the earlier the surgery is performed, the better the chance of survival.
A patent ductus arteriosus is a congenital condition in which the ductus arteriosus fails to close. The condition may range from severe to benign. Failure of the ductus arteriosus to close results in blood flowing from the higher-pressure aorta into the lower pressure pulmonary trunk. This additional fluid moving toward the lungs increases pulmonary pressure and makes respiration difficult. Symptoms include shortness of breath (dyspnoea), tachycardia, enlarged heart, a widened pulse pressure, and poor weight gain in infants. Treatments include surgical closure (ligation), manual closure using platinum coils or specialised mesh inserted via the femoral artery or vein, or nonsteroidal anti-inflammatory drugs to block the synthesis of prostaglandin E2, which maintains the vessel in an open position. If untreated, the condition can result in congestive heart failure.
Septal defects are not uncommon in individuals and may be congenital or caused by various disease processes. Tetralogy of Fallot is a congenital condition that may also occur from exposure to unknown environmental factors; it occurs when there is an opening in the interventricular septum caused by blockage of the pulmonary trunk, normally at the pulmonary semilunar valve. This allows blood that is relatively low in oxygen from the right ventricle to flow into the left ventricle and mix with the blood that is relatively high in oxygen. Symptoms include a distinct heart murmur, low blood oxygen percent saturation, dyspnoea or difficulty in breathing, polycythaemia, broadening (clubbing) of the fingers and toes, and in children, difficulty in feeding or failure to grow and develop. It is the most common cause of cyanosis following birth. The term “tetralogy” is derived from the four components of the condition, although only three may be present in an individual patient: pulmonary infundibular stenosis (rigidity of the pulmonary valve), overriding aorta (the aorta is shifted above both ventricles), ventricular septal defect (opening), and right ventricular hypertrophy (enlargement of the right ventricle). Other heart defects may also accompany this condition, which is typically confirmed by echocardiography imaging. Tetralogy of Fallot occurs in approximately 400 out of one million live births. Normal treatment involves extensive surgical repair, including the use of stents to redirect blood flow and replacement of valves and patches to repair the septal defect, but the condition has a relatively high mortality. Survival rates are currently 75 percent during the first year of life; 60 percent by four years of age; 30 percent by 10 years; and five percent by 40 years.
In the case of severe septal defects, including both tetralogy of Fallot and patent foramen ovale, failure of the heart to develop properly can lead to a condition commonly known as a “blue baby.” Regardless of normal skin pigmentation, individuals with this condition have an insufficient supply of oxygenated blood, which leads to cyanosis, a blue or purple coloration of the skin, especially when active.
Septal defects are commonly first detected through auscultation, listening to the chest using a stethoscope. In this case, instead of hearing normal heart sounds attributed to the flow of blood and closing of heart valves, unusual heart sounds may be detected. This is often followed by medical imaging to confirm or rule out a diagnosis. In many cases, treatment may not be needed. Some common congenital heart defects are illustrated in Figure 6.1.9.
Figure 6.1.9. Congenital heart defects. (a) A patent foramen ovale defect is an abnormal opening in the interatrial septum, or more commonly, a failure of the foramen ovale to close. (b) Coarctation of the aorta is an abnormal narrowing of the aorta. (c) A patent ductus arteriosus is the failure of the ductus arteriosus to close. (d) Tetralogy of Fallot includes an abnormal opening in the interventricular septum.
Right Atrium
The right atrium serves as the receiving chamber for blood returning to the heart from the systemic circulation. The two major systemic veins, the superior and inferior venae cavae, and the large coronary vein called the coronary sinus that drains the heart myocardium empty into the right atrium. The superior vena cava drains blood from regions superior to the diaphragm: the head, neck, upper limbs, and the thoracic region. It empties into the superior and posterior portions of the right atrium. The inferior vena cava drains blood from areas inferior to the diaphragm: the lower limbs and abdominopelvic region of the body. It, too, empties into the posterior portion of the atria, but inferior to the opening of the superior vena cava. Immediately superior and slightly medial to the opening of the inferior vena cava on the posterior surface of the atrium is the opening of the coronary sinus. This thin-walled vessel drains most of the coronary veins that return systemic blood from the heart. The majority of the internal heart structures discussed in this and subsequent sections are illustrated in Figure 6.1.8.
While the bulk of the internal surface of the right atrium is smooth, the depression of the fossa ovalis is medial, and the anterior surface demonstrates prominent ridges of muscle called the pectinate muscles. The right auricle also has pectinate muscles. The left atrium does not have pectinate muscles except in the auricle.
The atria receive venous blood on a nearly continuous basis, preventing venous flow from stopping while the ventricles are contracting. While most ventricular filling occurs while the atria are relaxed, they do demonstrate a contractile phase and actively pump blood into the ventricles just prior to ventricular contraction. The opening between the atrium and ventricle is guarded by the tricuspid valve.
Right Ventricle
The right ventricle receives blood from the right atrium through the tricuspid valve. Each flap of the valve is attached to strong strands of connective tissue, the chordae tendineae, literally “tendinous cords,” or sometimes more poetically referred to as “heart strings.” There are several chordae tendineae associated with each of the flaps. They are composed of approximately 80 percent collagenous fibres with the remainder consisting of elastic fibres and endothelium. They connect each of the flaps to a papillary muscle that extends from the inferior ventricular surface. There are three papillary muscles in the right ventricle, called the anterior, posterior, and septal muscles, which correspond to the three sections of the valves.
Figure 6.1.10. Chordae tendineae and papillary muscles. In this frontal section, you can see papillary muscles attached to the tricuspid valve on the right as well as the mitral valve on the left via chordae tendineae. (credit: modification of work by “PV KS”/flickr.com).
When the myocardium of the ventricle contracts, pressure within the ventricular chamber rises. Blood, like any fluid, flows from higher pressure to lower pressure areas, in this case, toward the pulmonary trunk and the atrium. To prevent any potential backflow, the papillary muscles also contract, generating tension on the chordae tendineae. This prevents the flaps of the valves from being forced into the atria and regurgitation of the blood back into the atria during ventricular contraction. Figure 6.1.10 shows papillary muscles and chordae tendineae attached to the tricuspid valve.
The walls of the ventricle are lined with trabeculae carneae, ridges of cardiac muscle covered by endocardium. In addition to these muscular ridges, a band of cardiac muscle, also covered by endocardium, known as the moderator band (see Figure 6.1.8) reinforces the thin walls of the right ventricle and plays a crucial role in cardiac conduction. It arises from the inferior portion of the interventricular septum and crosses the interior space of the right ventricle to connect with the inferior papillary muscle.
When the right ventricle contracts, it ejects blood into the pulmonary trunk, which branches into the left and right pulmonary arteries that carry it to each lung. The superior surface of the right ventricle begins to taper as it approaches the pulmonary trunk. At the base of the pulmonary trunk is the pulmonary semilunar valve that prevents backflow from the pulmonary trunk.
Left Atrium
After exchange of gases in the pulmonary capillaries, blood returns to the left atrium high in oxygen via one of the four pulmonary veins. While the left atrium does not contain pectinate muscles, it does have an auricle that includes these pectinate ridges. Blood flows nearly continuously from the pulmonary veins back into the atrium, which acts as the receiving chamber, and from here through an opening into the left ventricle. Most blood flows passively into the heart while both the atria and ventricles are relaxed, but toward the end of the ventricular relaxation period, the left atrium will contract, pumping blood into the ventricle. This atrial contraction accounts for approximately 20 percent of ventricular filling. The opening between the left atrium and ventricle is guarded by the mitral valve.
Left Ventricle
Recall that, although both sides of the heart will pump the same amount of blood, the muscular layer is much thicker in the left ventricle compared to the right (see Figure 6.1.7). Like the right ventricle, the left also has trabeculae carneae, but there is no moderator band. The mitral valve is connected to papillary muscles via chordae tendineae. There are two papillary muscles on the left—the anterior and posterior—as opposed to three on the right.
The left ventricle is the major pumping chamber for the systemic circuit; it ejects blood into the aorta through the aortic semilunar valve.
Heart Valve Structure and Function
A transverse section through the heart slightly above the level of the atrioventricular septum reveals all four heart valves along the same plane (Figure 6.1.11). The valves ensure unidirectional blood flow through the heart. Between the right atrium and the right ventricle is the right atrioventricular valve, or tricuspid valve. It typically consists of three flaps, or leaflets, made of endocardium reinforced with additional connective tissue. The flaps are connected by chordae tendineae to the papillary muscles, which control the opening and closing of the valves.
Figure 6.1.11. Heart valves. With the atria and major vessels removed, all four valves are clearly visible, although it is difficult to distinguish the three separate cusps of the tricuspid valve.
Emerging from the right ventricle at the base of the pulmonary trunk is the pulmonary semilunar valve, or the pulmonary valve; it is also known as the pulmonic valve or the right semilunar valve. The pulmonary valve is comprised of three small flaps of endothelium reinforced with connective tissue. When the ventricle relaxes, the pressure differential causes blood to flow back into the ventricle from the pulmonary trunk. This flow of blood fills the pocket-like flaps of the pulmonary valve, causing the valve to close and producing an audible sound. Unlike the atrioventricular valves, there are no papillary muscles or chordae tendineae associated with the pulmonary valve.
Located at the opening between the left atrium and left ventricle is the mitral valve, also called the bicuspid valve or the left atrioventricular valve. Structurally, this valve consists of two cusps, known as the anterior medial cusp and the posterior medial cusp, compared to the three cusps of the tricuspid valve. In a clinical setting, the valve is referred to as the mitral valve, rather than the bicuspid valve. The two cusps of the mitral valve are attached by chordae tendineae to two papillary muscles that project from the wall of the ventricle.
At the base of the aorta is the aortic semilunar valve, or the aortic valve, which prevents backflow from the aorta. It normally is composed of three flaps. When the ventricle relaxes and blood attempts to flow back into the ventricle from the aorta, blood will fill the cusps of the valve, causing it to close and producing an audible sound.
In Figure 6.1.12a, the two atrioventricular valves are open and the two semilunar valves are closed. This occurs when both atria and ventricles are relaxed and when the atria contract to pump blood into the ventricles. Figure 6.1.12b shows a frontal view. Although only the left side of the heart is illustrated, the process is virtually identical on the right.
Figure 6.1.12. Blood flow from the left atrium to the left ventricle. (a) A transverse section through the heart illustrates the four heart valves. The two atrioventricular valves are open; the two semilunar valves are closed. The atria and vessels have been removed. (b) A frontal section through the heart illustrates blood flow through the mitral valve. When the mitral valve is open, it allows blood to move from the left atrium to the left ventricle. The aortic semilunar valve is closed to prevent backflow of blood from the aorta to the left ventricle.
Figure 6.1.13a shows the atrioventricular valves closed while the two semilunar valves are open. This occurs when the ventricles contract to eject blood into the pulmonary trunk and aorta. Closure of the two atrioventricular valves prevents blood from being forced back into the atria. This stage can be seen from a frontal view in Figure 6.1.13b.
Figure 6.1.13. Blood flow from the left ventricle into the great vessels. (a) A transverse section through the heart illustrates the four heart valves during ventricular contraction. The two atrioventricular valves are closed, but the two semilunar valves are open. The atria and vessels have been removed. (b) A frontal view shows the closed mitral (bicuspid) valve that prevents backflow of blood into the left atrium. The aortic semilunar valve is open to allow blood to be ejected into the aorta.
When the ventricles begin to contract, pressure within the ventricles rises and blood flows toward the area of lowest pressure, which is initially in the atria. This backflow causes the cusps of the tricuspid and mitral (bicuspid) valves to close. These valves are tied down to the papillary muscles by chordae tendineae. During the relaxation phase of the cardiac cycle, the papillary muscles are also relaxed and the tension on the chordae tendineae is slight (see Figure 6.1.12b). However, as the myocardium of the ventricle contracts, so do the papillary muscles. This creates tension on the chordae tendineae (see Figure 6.1.13b), helping to hold the cusps of the atrioventricular valves in place and preventing them from being blown back into the atria.
The aortic and pulmonary semilunar valves lack the chordae tendineae and papillary muscles associated with the atrioventricular valves. Instead, they consist of pocket-like folds of endocardium reinforced with additional connective tissue. When the ventricles relax and the change in pressure forces the blood toward the ventricles, the blood presses against these cusps and seals the openings.
Disorders of the Heart Valves
When heart valves do not function properly, they are often described as incompetent and result in valvular heart disease, which can range from benign to lethal. Some of these conditions are congenital, that is, the individual was born with the defect, whereas others may be attributed to disease processes or trauma. Some malfunctions are treated with medications, others require surgery, and still others may be mild enough that the condition is merely monitored since treatment might trigger more serious consequences.
Valvular disorders are often caused by carditis, or inflammation of the heart. One common trigger for this inflammation is rheumatic fever, or scarlet fever, an autoimmune response to the presence of a bacterium, Streptococcus pyogenes, normally a disease of childhood.
While any of the heart valves may be involved in valve disorders, mitral regurgitation is the most common, detected in approximately 2 percent of the population, and the pulmonary semilunar valve is the least frequently involved. When a valve malfunctions, the flow of blood to a region will often be disrupted. The resulting inadequate flow of blood to this region will be described in general terms as an insufficiency. The specific type of insufficiency is named for the valve involved: aortic insufficiency, mitral insufficiency, tricuspid insufficiency, or pulmonary insufficiency.
If one of the cusps of the valve is forced backward by the force of the blood, the condition is referred to as a prolapsed valve. Prolapse may occur if the chordae tendineae are damaged or broken, causing the closure mechanism to fail. The failure of the valve to close properly disrupts the normal one-way flow of blood and results in regurgitation, when the blood flows backward from its normal path. Using a stethoscope, the disruption to the normal flow of blood produces a heart murmur.
Stenosis is a condition in which the heart valves become rigid and may calcify over time. The loss of flexibility of the valve interferes with normal function and may cause the heart to work harder to propel blood through the valve, which eventually weakens the heart. Aortic stenosis affects approximately two percent of the population over 65 years of age, and the percentage increases to approximately four percent in individuals over 85 years. Occasionally, one or more of the chordae tendineae will tear or the papillary muscle itself may die as a component of a myocardial infarction (heart attack). In this case, the patient’s condition will deteriorate dramatically and rapidly, and immediate surgical intervention may be required.
Auscultation, or listening to a patient’s heart sounds, is one of the most useful diagnostic tools, since it is proven, safe, and inexpensive. The term auscultation is derived from the Latin for “to listen,” and the technique has been used for diagnostic purposes as far back as the ancient Egyptians. Valve and septal disorders will trigger abnormal heart sounds. If a valvular disorder is detected or suspected, a test called an echocardiogram, or simply an “echo,” may be ordered. Echocardiograms are sonograms of the heart and can help in the diagnosis of valve disorders as well as a wide variety of heart pathologies.
Career Connection
Cardiologist
Cardiologists are medical doctors that specialise in the diagnosis and treatment of diseases of the heart. The pathway to become a certified cardiologist in Australia consists of four basic steps. Firstly, a completion of an undergraduate degree (3 years full time) in either a health science or biomedical science field with provisional entry into medicine. Second step is completing either a Bachelor of Medicine/Bachelor of Surgery or a Doctor of Medicine (MD) degree which generally takes four years full time. The third step is to complete an internship/residency in a public hospital system (two years). The final stage requires further vocational training and a Fellowship in a specialised training program (e.g. Fellowship of the Royal Australasian College of Physicians (FRACP) which takes approximately three to five years to complete. The annual salary of a Cardiologist in Australia can range between $60k up to $630k.
Cardiovascular Technologist/Technician
Cardiovascular technologists/technicians are trained professionals who perform a variety of imaging techniques, such as sonograms or echocardiograms, used by physicians to diagnose and treat diseases of the heart. Nearly all of these positions require an associate degree, and include, but are not limited to, Grade 1 Cardiac Technician (~$48k – $80k per annum), radiographer (~$55k – $104k per annum), cardiac sonographer (~$62k – $135k per annum).
Coronary Circulation
You will recall that the heart is a remarkable pump composed largely of cardiac muscle cells that are incredibly active throughout life. Like all other cells, a cardiomyocyte requires a reliable supply of oxygen and nutrients, and a way to remove wastes, so it needs a dedicated, complex, and extensive coronary circulation. And because of the critical and nearly ceaseless activity of the heart throughout life, this need for a blood supply is even greater than for a typical cell. However, coronary circulation is not continuous; rather, it cycles, reaching a peak when the heart muscle is relaxed and nearly ceasing while it is contracting.
Coronary Arteries
Coronary arteries supply blood to the myocardium and other components of the heart. The first portion of the aorta after it arises from the left ventricle gives rise to the coronary arteries. There are three dilations in the wall of the aorta just superior to the aortic semilunar valve. Two of these, the left posterior aortic sinus and anterior aortic sinus, give rise to the left and right coronary arteries, respectively. The third sinus, the right posterior aortic sinus, typically does not give rise to a vessel. Coronary vessel branches that remain on the surface of the artery and follow the sulci are called epicardial coronary arteries.
The left coronary artery distributes blood to the left side of the heart, the left atrium and ventricle, and the interventricular septum. The circumflex artery arises from the left coronary artery and follows the coronary sulcus to the left. Eventually, it will fuse with the small branches of the right coronary artery. The larger anterior interventricular artery, also known as the left anterior descending artery (LAD), is the second major branch arising from the left coronary artery. It follows the anterior interventricular sulcus around the pulmonary trunk. Along the way it gives rise to numerous smaller branches that interconnect with the branches of the posterior interventricular artery, forming anastomoses. An anastomosis is an area where vessels unite to form interconnections that normally allow blood to circulate to a region even if there may be partial blockage in another branch. The anastomoses in the heart are very small. Therefore, this ability is somewhat restricted in the heart, so a coronary artery blockage often results in death of the cells (myocardial infarction) supplied by the particular vessel.
The right coronary artery proceeds along the coronary sulcus and distributes blood to the right atrium, portions of both ventricles, and the heart conduction system. Normally, one or more marginal arteries arise from the right coronary artery inferior to the right atrium. The marginal arteries supply blood to the superficial portions of the right ventricle. On the posterior surface of the heart, the right coronary artery gives rise to the posterior interventricular artery, also known as the posterior descending artery. It runs along the posterior portion of the interventricular sulcus toward the apex of the heart, giving rise to branches that supply the interventricular septum and portions of both ventricles. Figure 6.1.14 presents views of the coronary circulation from both the anterior and posterior views.
Figure 6.1.14. Coronary circulation. The anterior view of the heart shows the prominent coronary surface vessels. The posterior view of the heart shows the prominent coronary surface vessels.
Diseases of the Heart: Myocardial Infarction
Myocardial infarction (MI) is the formal term for what is commonly referred to as a heart attack. It normally results from a lack of blood flow (ischaemia) and oxygen (hypoxia) to a region of the heart, resulting in death of the cardiac muscle cells. An MI often occurs when a coronary artery is blocked by the build-up of atherosclerotic plaque consisting of lipids, cholesterol and fatty acids, and white blood cells, primarily macrophages. It can also occur when a portion of an unstable atherosclerotic plaque travels through the coronary arterial system and lodges in one of the smaller vessels. The resulting blockage restricts the flow of blood and oxygen to the myocardium and causes death of the tissue. MIs may be triggered by excessive exercise, in which the partially occluded artery is no longer able to pump sufficient quantities of blood, or severe stress, which may induce spasm of the smooth muscle in the walls of the vessel.
In the case of acute MI, there is often sudden pain beneath the sternum (retrosternal pain) called angina pectoris, often radiating down the left arm in males but not in female patients. Until this anomaly between the sexes was discovered, many female patients suffering MIs were misdiagnosed and sent home. In addition, patients typically present with difficulty breathing and shortness of breath (dyspnoea), irregular heartbeat (palpations), nausea and vomiting, sweating (diaphoresis), anxiety and fainting (syncope), although not all of these symptoms may be present. Many of the symptoms are shared with other medical conditions, including anxiety attacks and simple indigestion, so differential diagnosis is critical. It is estimated that between 22 and 64 percent of MIs present without any symptoms.
An MI can be confirmed by examining the patient’s ECG, which frequently reveals alterations in the ST and Q components. Some classification schemes of MI are referred to as ST-elevated MI (STEMI) and non-elevated MI (non-STEMI). In addition, echocardiography or cardiac magnetic resonance imaging may be employed. Common blood tests indicating an MI include elevated levels of creatine kinase MB (an enzyme that catalyses the conversion of creatine to phosphocreatine, consuming ATP) and cardiac troponin (the regulatory protein for muscle contraction), both of which are released by damaged cardiac muscle cells.
Immediate treatments for MI are essential and include administering supplemental oxygen, aspirin that helps to break up clots, and nitroglycerine administered sublingually (under the tongue) to facilitate its absorption. Despite its unquestioned success in treatments and use since the 1880s, the mechanism of nitroglycerine is still incompletely understood but is believed to involve the release of nitric oxide, a known vasodilator, and endothelium-derived releasing factor, which also relaxes the smooth muscle in the tunica media of coronary vessels. Longer-term treatments include injections of thrombolytic agents such as streptokinase that dissolve the clot, the anticoagulant heparin, balloon angioplasty and stents to open blocked vessels, and bypass surgery to allow blood to pass around the site of blockage. If the damage is extensive, coronary replacement with a donor heart or coronary assist device, a sophisticated mechanical device that supplements the pumping activity of the heart, may be employed. Despite the attention, development of artificial hearts to augment the severely limited supply of heart donors has proven less than satisfactory but will likely improve in the future.
MIs may trigger cardiac arrest, but the two are not synonymous. Important risk factors for MI include cardiovascular disease, age, smoking, high blood levels of the low-density lipoprotein (LDL, often referred to as “bad” cholesterol), low levels of high-density lipoprotein (HDL, or “good” cholesterol), hypertension, diabetes mellitus, obesity, lack of physical exercise, chronic kidney disease, excessive alcohol consumption, and use of illegal drugs.
Coronary Veins
Coronary veins drain the heart and generally parallel the large surface arteries (see Figure 6.1.14). The great cardiac vein can be seen initially on the surface of the heart following the interventricular sulcus, but it eventually flows along the coronary sulcus into the coronary sinus on the posterior surface. The great cardiac vein initially parallels the anterior interventricular artery and drains the areas supplied by this vessel. It receives several major branches, including the posterior cardiac vein, the middle cardiac vein, and the small cardiac vein. The posterior cardiac vein parallels and drains the areas supplied by the marginal artery branch of the circumflex artery. The middle cardiac vein parallels and drains the areas supplied by the posterior interventricular artery. The small cardiac vein parallels the right coronary artery and drains the blood from the posterior surfaces of the right atrium and ventricle. The coronary sinus is a large, thin-walled vein on the posterior surface of the heart lying within the atrioventricular sulcus and emptying directly into the right atrium. The anterior cardiac veins parallel the small cardiac arteries and drain the anterior surface of the right ventricle. Unlike these other cardiac veins, it bypasses the coronary sinus and drains directly into the right atrium.
Diseases of the Heart: Coronary Artery Disease
Figure 6.1.15. Atherosclerotic coronary arteries. In this coronary angiogram (X-ray), the dye makes visible two occluded coronary arteries. Such blockages can lead to decreased blood flow (ischaemia) and insufficient oxygen (hypoxia) delivered to the cardiac tissues. If uncorrected, this can lead to cardiac muscle death (myocardial infarction).
Coronary artery disease is the leading cause of death worldwide. It occurs when the build-up of plaque—a fatty material including cholesterol, connective tissue, white blood cells, and some smooth muscle cells—within the walls of the arteries obstructs the flow of blood and decreases the flexibility or compliance of the vessels. This condition is called atherosclerosis, a hardening of the arteries that involves the accumulation of plaque. As the coronary blood vessels become occluded, the flow of blood to the tissues will be restricted, a condition called ischaemia that causes the cells to receive insufficient amounts of oxygen, called hypoxia. Figure 6.1.15 shows the blockage of coronary arteries highlighted by the injection of dye. Some individuals with coronary artery disease report pain radiating from the chest called angina pectoris, but others remain asymptomatic. If untreated, coronary artery disease can lead to MI or a heart attack.
The disease progresses slowly and often begins in children and can be seen as fatty “streaks” in the vessels. It then gradually progresses throughout life. Well-documented risk factors include smoking, family history, hypertension, obesity, diabetes, high alcohol consumption, lack of exercise, stress, and hyperlipidaemia or high circulating levels of lipids in the blood. Treatments may include medication, changes to diet and exercise, angioplasty with a balloon catheter, insertion of a stent, or coronary bypass procedure.
Angioplasty is a procedure in which the occlusion is mechanically widened with a balloon. A specialised catheter with an expandable tip is inserted into a superficial vessel, normally in the leg, and then directed to the site of the occlusion. At this point, the balloon is inflated to compress the plaque material and to open the vessel to increase blood flow. Then, the balloon is deflated and retracted. A stent consisting of a specialised mesh is typically inserted at the site of occlusion to reinforce the weakened and damaged walls. Stent insertions have been routine in cardiology for more than 40 years.
Coronary bypass surgery may also be performed. This surgical procedure grafts a replacement vessel obtained from another, less vital portion of the body to bypass the occluded area. This procedure is clearly effective in treating patients experiencing a MI, but overall does not increase longevity. Nor does it seem advisable in patients with stable although diminished cardiac capacity since frequently loss of mental acuity occurs following the procedure. Long-term changes to behaviour, emphasising diet and exercise plus a medicine regime tailored to lower blood pressure, lower cholesterol and lipids, and reduce clotting are equally as effective.
Section Review
The heart resides within the pericardial sac and is located in the mediastinal space within the thoracic cavity. The pericardial sac consists of two fused layers: an outer fibrous capsule and an inner parietal pericardium lined with a serous membrane. Between the pericardial sac and the heart is the pericardial cavity, which is filled with lubricating serous fluid. The walls of the heart are composed of an outer epicardium, a thick myocardium, and an inner lining layer of endocardium. The human heart consists of a pair of atria, which receive blood and pump it into a pair of ventricles, which pump blood into the vessels. The right atrium receives systemic blood relatively low in oxygen and pumps it into the right ventricle, which pumps it into the pulmonary circuit. Exchange of oxygen and carbon dioxide occurs in the lungs, and blood high in oxygen returns to the left atrium, which pumps blood into the left ventricle, which in turn pumps blood into the aorta and the remainder of the systemic circuit. The septa are the partitions that separate the chambers of the heart. They include the interatrial septum, the interventricular septum, and the atrioventricular septum. Two of these openings are guarded by the atrioventricular valves, the right tricuspid valve and the left mitral valve, which prevent the backflow of blood. Each is attached to chordae tendineae that extend to the papillary muscles, which are extensions of the myocardium, to prevent the valves from being blown back into the atria. The pulmonary valve is located at the base of the pulmonary trunk, and the left semilunar valve is located at the base of the aorta. The right and left coronary arteries are the first to branch off the aorta and arise from two of the three sinuses located near the base of the aorta and are generally located in the sulci. Cardiac veins parallel the small cardiac arteries and generally drain into the coronary sinus.
Identify and describe the components of the conducting system that distributes electrical impulses through the heart
Compare the effect of ion movement on membrane potential of cardiac conductive and contractile cells
Relate characteristics of an electrocardiogram (ECG) to events in the cardiac cycle
Identify blocks that can interrupt the cardiac cycle
Recall that cardiac muscle shares a few characteristics with both skeletal muscle and smooth muscle, but it has some unique properties of its own. Not the least of these exceptional properties is its ability to initiate an electrical potential at a fixed rate that spreads rapidly from cell to cell to trigger the contractile mechanism. This property is known as autorhythmicity. Neither smooth nor skeletal muscle can do this. Even though cardiac muscle has autorhythmicity, heart rate is modulated by the endocrine and nervous systems.
There are two major types of cardiac muscle cells: myocardial contractile cells and myocardial conducting cells. The myocardial contractile cells constitute the bulk (99 percent) of the cells in the atria and ventricles. Contractile cells conduct impulses and are responsible for contractions that pump blood through the body. The myocardial conducting cells (1 percent of the cells) form the conduction system of the heart. Except for Purkinje cells, they are generally much smaller than the contractile cells and have few of the myofibrils or filaments needed for contraction. Their function is similar in many respects to neurons, although they are specialised muscle cells. Myocardial conduction cells initiate and propagate the action potential (the electrical impulse) that travels throughout the heart and triggers the contractions that propel the blood.
Structure of Cardiac Muscle
Compared to the giant cylinders of skeletal muscle, cardiac muscle cells, or cardiomyocytes, are considerably shorter with much smaller diameters. Cardiac muscle also demonstrates striations, the alternating pattern of dark A bands and light I bands attributed to the precise arrangement of the myofilaments and fibrils that are organised in sarcomeres along the length of the cell (Figure 6.2.1a). These contractile elements are virtually identical to skeletal muscle. T (transverse) tubules penetrate from the surface plasma membrane, the sarcolemma, to the interior of the cell, allowing the electrical impulse to reach the interior. The T tubules are only found at the Z discs, whereas in skeletal muscle, they are found at the junction of the A and I bands. Therefore, there are one-half as many T tubules in cardiac muscle as in skeletal muscle. In addition, the sarcoplasmic reticulum stores few calcium ions, so most of the calcium ions must come from outside the cells. The result is a slower onset of contraction. Mitochondria are plentiful, providing energy for the contractions of the heart. Typically, cardiomyocytes have a single, central nucleus, but two or more nuclei may be found in some cells.
Cardiac muscle cells branch freely. A junction between two adjoining cells is marked by a critical structure called an intercalated disc, which helps support the synchronised contraction of the muscle (Figure 6.2.1b). The sarcolemmas from adjacent cells bind together at the intercalated discs. They consist of desmosomes, specialised linking proteoglycans, tight junctions, and large numbers of gap junctions that allow the passage of ions between the cells and help to synchronise the contraction (Figure 6.2.1c). Intercellular connective tissue also helps to bind the cells together. The importance of strongly binding these cells together is necessitated by the forces exerted by contraction.
Cardiac muscle undergoes aerobic respiration patterns, primarily metabolising lipids and carbohydrates. Myoglobin, lipids, and glycogen are all stored within the cytoplasm. Cardiac muscle cells undergo twitch-type contractions with long refractory periods followed by brief relaxation periods. The relaxation is essential so the heart can fill with blood for the next cycle. The refractory period is very long to prevent the possibility of tetany, a condition in which muscle remains involuntarily contracted. In the heart, tetany is not compatible with life, since it would prevent the heart from pumping blood.
Everyday Connection
Repair and Replacement
Damaged cardiac muscle cells have extremely limited abilities to repair themselves or to replace dead cells via mitosis. Recent evidence indicates that at least some stem cells remain within the heart that continue to divide and at least potentially replace these dead cells. However, newly formed or repaired cells are rarely as functional as the original cells, and cardiac function is reduced. In the event of a heart attack or MI, dead cells are often replaced by patches of scar tissue. Autopsies performed on individuals who had successfully received heart transplants show some proliferation of original cells. If researchers can unlock the mechanism that generates new cells and restore full mitotic capabilities to heart muscle, the prognosis for heart attack survivors will be greatly enhanced. To date, myocardial cells produced within the patient (in situ) by cardiac stem cells seem to be non-functional, although those grown in Petri dishes (in vitro) do beat. Perhaps soon this mystery will be solved, and new advances in treatment will be commonplace.
Conduction System of the Heart
If embryonic heart cells are separated into a Petri dish and kept alive, each can generate its own electrical impulse followed by contraction. When two independently beating embryonic cardiac muscle cells are placed together, the cell with the higher inherent rate sets the pace, and the impulse spreads from the faster to the slower cell to trigger a contraction. As more cells are joined together, the fastest cell continues to assume control of the rate. A fully developed adult heart maintains the capability of generating its own electrical impulse, triggered by the fastest cells, as part of the cardiac conduction system. The components of the cardiac conduction system include the sinoatrial node, the atrioventricular node, the atrioventricular bundle, the atrioventricular bundle branches, and the Purkinje cells (Figure 6.2.2).
Figure 6.2.2. Conduction system of the heart. Specialised conducting components of the heart include the sinoatrial node, the internodal pathways, the atrioventricular node, the atrioventricular bundle, the right and left bundle branches, and the Purkinje fibres.
Sinoatrial (SA) Node
Normal cardiac rhythm is established by the sinoatrial (SA) node, a specialised clump of myocardial conducting cells located in the superior and posterior walls of the right atrium in close proximity to the orifice of the superior vena cava. The SA node has the highest inherent rate of depolarisation and is known as the pacemaker of the heart. It initiates the sinus rhythm, or normal electrical pattern followed by contraction of the heart.
This impulse spreads from its initiation in the SA node throughout the atria through specialised internodal pathways, to the atrial myocardial contractile cells and the atrioventricular node. The internodal pathways consist of three bands (anterior, middle, and posterior) that lead directly from the SA node to the next node in the conduction system, the atrioventricular node (see Figure 6.2.2). The impulse takes approximately 50 ms (milliseconds) to travel between these two nodes. The relative importance of this pathway has been debated since the impulse would reach the atrioventricular node simply following the cell-by-cell pathway through the contractile cells of the myocardium in the atria. In addition, there is a specialised pathway called Bachmann’s bundle or the interatrial band that conducts the impulse directly from the right atrium to the left atrium. Regardless of the pathway, as the impulse reaches the atrioventricular septum, the connective tissue of the cardiac skeleton prevents the impulse from spreading into the myocardial cells in the ventricles except at the atrioventricular node. Figure 6.2.3 illustrates the initiation of the impulse in the SA node that then spreads the impulse throughout the atria to the atrioventricular node.
Figure 6.2.3. Cardiac conduction. (1) The sinoatrial (SA) node and the remainder of the conduction system are at rest. (2) The SA node initiates the action potential, which sweeps across the atria. (3) After reaching the atrioventricular node, there is a delay of approximately 100 ms that allows the atria to complete pumping blood before the impulse is transmitted to the atrioventricular bundle. (4) Following the delay, the impulse travels through the atrioventricular bundle and bundle branches to the Purkinje fibres, and reaches the right papillary muscle via the moderator band. (5) The impulse spreads to the contractile fibres of the ventricle. (6) Ventricular contraction begins.
The electrical event, the wave of depolarisation, is the trigger for muscular contraction. The wave of depolarisation begins in the right atrium, and the impulse spreads across the superior portions of both atria and then down through the contractile cells. The contractile cells then begin contraction from the superior to the inferior portions of the atria, efficiently pumping blood into the ventricles.
Atrioventricular (AV) Node
The atrioventricular (AV) node is a second clump of specialised myocardial conductive cells, located in the inferior portion of the right atrium within the atrioventricular septum. The septum prevents the impulse from spreading directly to the ventricles without passing through the AV node. There is a critical pause before the AV node depolarises and transmits the impulse to the atrioventricular bundle (see Figure 6.2.3, step 3). This delay in transmission is partially attributable to the small diameter of the cells of the node, which slow the impulse. Also, conduction between nodal cells is less efficient than between conducting cells. These factors mean that it takes the impulse approximately 100 ms to pass through the node. This pause is critical to heart function, as it allows the atrial cardiomyocytes to complete their contraction that pumps blood into the ventricles before the impulse is transmitted to the cells of the ventricle itself. With extreme stimulation by the SA node, the AV node can transmit impulses maximally at 220 per minute. This establishes the typical maximum heart rate in a healthy young individual. Damaged hearts or those stimulated by drugs can contract at higher rates, but at these rates, the heart can no longer effectively pump blood.
Atrioventricular Bundle (Bundle of His), Bundle Branches and Purkinje fibres
Arising from the AV node, the atrioventricular bundle, or bundle of His, proceeds through the interventricular septum before dividing into two atrioventricular bundle branches, commonly called the left and right bundle branches. The left bundle branch has two fascicles. The left bundle branch supplies the left ventricle, and the right bundle branch the right ventricle. Since the left ventricle is much larger than the right, the left bundle branch is also considerably larger than the right. Portions of the right bundle branch are found in the moderator band and supply the right papillary muscles. Because of this connection, each papillary muscle receives the impulse at approximately the same time, so they begin to contract simultaneously just prior to the remainder of the myocardial contractile cells of the ventricles. This is believed to allow tension to develop on the chordae tendineae prior to right ventricular contraction. There is no corresponding moderator band on the left. Both bundle branches descend and reach the apex of the heart where they connect with the Purkinje fibres (see Figure 6.2.3, step 4). This passage takes approximately 25 ms.
The Purkinje fibres are additional myocardial conductive fibres that spread the impulse to the myocardial contractile cells in the ventricles. They extend throughout the myocardium from the apex of the heart toward the atrioventricular septum and the base of the heart. The Purkinje fibres have a fast inherent conduction rate, and the electrical impulse reaches all the ventricular muscle cells in about 75 ms (see Figure 6.2.3, step 5). Since the electrical stimulus begins at the apex, the contraction also begins at the apex and travels toward the base of the heart, like squeezing a tube of toothpaste from the bottom. This allows the blood to be pumped out of the ventricles and into the aorta and pulmonary trunk. The total time elapsed from the initiation of the impulse in the SA node until depolarisation of the ventricles is approximately 225 ms.
Membrane Potentials and Ion Movement in Cardiac Conductive Cells
Action potentials are considerably different between cardiac conductive cells and cardiac contractive cells. While Na+ and K+ play essential roles, Ca2+ is also critical for both types of cells. Unlike skeletal muscles and neurons, cardiac conductive cells do not have a stable resting potential. Conductive cells contain a series of sodium ion channels that allow a normal and slow influx of sodium ions that causes the membrane potential to rise slowly from an initial value of −60 mV up to about –40 mV. The resulting movement of sodium ions creates spontaneous depolarisation (or prepotential depolarisation). At this point, calcium ion channels open and Ca2+ enters the cell, further depolarising it at a more rapid rate until it reaches a value of approximately +5 mV. At this point, the calcium ion channels close and K+ channels open, allowing outflux of K+ and resulting in repolarisation. When the membrane potential reaches approximately −60 mV, the K+ channels close and Na+ channels open, and the prepotential phase begins again. This phenomenon explains the autorhythmicity properties of cardiac muscle (Figure 6.2.4).
Figure 6.2.4. Action potential at the SA node. The prepotential is due to a slow influx of sodium ions until the threshold is reached followed by a rapid depolarisation and repolarisation. The prepotential accounts for the membrane reaching threshold and initiates the spontaneous depolarisation and contraction of the cell. Note the lack of a resting potential.
Membrane Potentials and Ion Movement in Cardiac Contractile Cells
There is a distinctly different electrical pattern involving the contractile cells. In this case, there is a rapid depolarisation, followed by a plateau phase and then repolarisation. This phenomenon accounts for the long refractory periods required for the cardiac muscle cells to pump blood effectively before they are capable of firing for a second time. These cardiac myocytes normally do not initiate their own electrical potential, although they can do so, but rather wait for an impulse to reach them.
Contractile cells demonstrate a much more stable resting phase than conductive cells at approximately −80 mV for cells in the atria and −90 mV for cells in the ventricles. Despite this initial difference, the other components of their action potentials are virtually identical. In both cases, when stimulated by an action potential, voltage-gated channels rapidly open, beginning the positive-feedback mechanism of depolarisation. This rapid influx of positively charged ions raises the membrane potential to approximately +30 mV, at which point the sodium channels close. The rapid depolarisation period typically lasts 3–5 ms. Depolarisation is followed by the plateau phase, in which membrane potential declines relatively slowly. This is due in large part to the opening of the slow Ca2+ channels, allowing Ca2+ to enter the cell while few K+ channels are open, allowing K+ to exit the cell. The relatively long plateau phase lasts approximately 175 ms. Once the membrane potential reaches approximately zero, the Ca2+ channels close and K+ channels open, allowing K+ to exit the cell. The repolarisation lasts approximately 75 ms. At this point, membrane potential drops until it reaches resting levels once more and the cycle repeats. The entire event lasts between 250 and 300 ms (Figure 6.2.5).
The absolute refractory period for cardiac contractile muscle lasts approximately 200 ms, and the relative refractory period lasts approximately 50 ms, for a total of 250 ms. This extended period is critical, since the heart muscle must contract to pump blood effectively and the contraction must follow the electrical events. Without extended refractory periods, premature contractions would occur in the heart and would not be compatible with life.
Figure 6.2.5. Action potential in cardiac contractile cells. (a) Note the long plateau phase due to the influx of calcium ions. The extended refractory period allows the cell to fully contract before another electrical event can occur. (b) The action potential for heart muscle is compared to that of skeletal muscle.
Calcium Ions
Calcium ions play two critical roles in the physiology of cardiac muscle. Their influx through slow calcium channels accounts for the prolonged plateau phase and absolute refractory period that enable cardiac muscle to function properly. Calcium ions also combine with the regulatory protein troponin in the troponin-tropomyosin complex; this complex removes the inhibition that prevents the heads of the myosin molecules from forming cross bridges with the active sites on actin that provide the power stroke of contraction. This mechanism is virtually identical to that of skeletal muscle. Approximately 20 percent of the calcium required for contraction is supplied by the influx of Ca2+ during the plateau phase. The remaining Ca2+ for contraction is released from storage in the sarcoplasmic reticulum.
Comparative Rates of Conduction System Firing
The pattern of prepotential or spontaneous depolarisation, followed by rapid depolarisation and repolarisation just described, are seen in the SA node and a few other conductive cells in the heart. Since the SA node is the pacemaker, it reaches threshold faster than any other component of the conduction system. It will initiate the impulses spreading to the other conducting cells. The SA node, without nervous or endocrine control, would initiate a heart impulse approximately 80–100 times per minute. Although each component of the conduction system can generate its own impulse, the rate progressively slows as you proceed from the SA node to the Purkinje fibres. Without the SA node, the AV node would generate a heart rate of 40–60 beats per minute. If the AV node were blocked, the atrioventricular bundle would fire at a rate of approximately 30–40 impulses per minute. The bundle branches would have an inherent rate of 20–30 impulses per minute, and the Purkinje fibres would fire at 15–20 impulses per minute. While a few exceptionally trained aerobic athletes demonstrate resting heart rates in the range of 30–40 beats per minute (the lowest recorded figure is 28 beats per minute for Miguel Indurain, a cyclist), for most individuals, rates lower than 50 beats per minute would indicate a condition called bradycardia. Depending upon the specific individual, as rates fall much below this level, the heart would be unable to maintain adequate flow of blood to vital tissues, initially resulting in decreasing loss of function across the systems, unconsciousness, and ultimately death.
Electrocardiogram
By careful placement of surface electrodes on the body, it is possible to record the complex, compound electrical signal of the heart. This tracing of the electrical signal is the electrocardiogram (ECG), also commonly abbreviated EKG (K coming kardiology, from the German term for cardiology). Careful analysis of the ECG reveals a detailed picture of both normal and abnormal heart function and is an indispensable clinical diagnostic tool. The standard electrocardiograph (the instrument that generates an ECG) uses 3, 5, or 12 leads. The greater the number of leads an electrocardiograph uses, the more information the ECG provides. The term “lead” may be used to refer to the cable from the electrode to the electrical recorder, but it typically describes the voltage difference between two of the electrodes. The 12-lead electrocardiograph uses 10 electrodes placed in standard locations on the patient’s skin (Figure 6.2.6). In continuous ambulatory electrocardiographs, the patient wears a small, portable, battery-operated device known as a Holter monitor, or simply a Holter, that continuously monitors heart electrical activity, typically for a period of 24 hours during the patient’s normal routine.
Figure 6.2.6. Standard placement of ECG leads. In a 12-lead ECG, six electrodes are placed on the chest, and four electrodes are placed on the limbs.
A normal ECG tracing is presented in Figure 6.2.7. Each component, segment and interval is labelled and corresponds to important electrical events, demonstrating the relationship between these events and contraction in the heart.
There are five prominent points on the ECG: the P wave, the QRS complex and the T wave. The small P wave represents the depolarisation of the atria. The atria begin contracting approximately 25 ms after the start of the P wave. The large QRS complex represents the depolarisation of the ventricles, which requires a much stronger electrical signal because of the larger size of the ventricular cardiac muscle. The ventricles begin to contract as the QRS reaches the peak of the R wave. Lastly, the T wave represents the repolarisation of the ventricles. The repolarisation of the atria occurs during the QRS complex, which masks it on an ECG.
The major segments and intervals of an ECG tracing are indicated in Figure 6.2.7. Segments are defined as the regions between two waves. Intervals include one segment plus one or more waves. For example, the PR segment begins at the end of the P wave and ends at the beginning of the QRS complex. The PR interval starts at the beginning of the P wave and ends with the beginning of the QRS complex. The PR interval is more clinically relevant, as it measures the duration from the beginning of atrial depolarisation (the P wave) to the initiation of the QRS complex. Since the Q wave may be difficult to view in some tracings, the measurement is often extended to the R that is more easily visible. Should there be a delay in passage of the impulse from the SA node to the AV node, it would be visible in the PR interval. Figure 6.2.8 correlates events of heart contraction to the corresponding segments and intervals of an ECG.
Figure 6.2.7. Electrocardiogram. A normal tracing shows the P wave, QRS complex, and T wave. Also indicated are the PR, QT, QRS and ST intervals, plus the P-R and S-T segments.Figure 6.2.8. ECG tracing correlated to the cardiac cycle. This diagram correlates an ECG tracing with the electrical and mechanical events of a heart contraction. Each segment of an ECG tracing corresponds to one event in the cardiac cycle.
Everyday Connection
ECG Abnormalities
Occasionally, an area of the heart other than the SA node will initiate an impulse that will be followed by a premature contraction. Such an area, which may be a component of the conduction system or some other contractile cells, is known as an ectopic focus or ectopic pacemaker. An ectopic focus may be stimulated by localised ischaemia; exposure to certain drugs, including caffeine, digitalis, or acetylcholine; elevated stimulation by both sympathetic or parasympathetic divisions of the autonomic nervous system; or several disease or pathological conditions. Occasional occurrences are generally transitory and nonlife threatening, but if the condition becomes chronic, it may lead to either an arrhythmia, a deviation from the normal pattern of impulse conduction and contraction, or to fibrillation, an uncoordinated beating of the heart.
While interpretation of an ECG is possible and extremely valuable after some training, a full understanding of the complexities and intricacies generally requires several years of experience. In general, the size of the electrical variations, the duration of the events, and detailed vector analysis provide the most comprehensive picture of cardiac function. For example, an amplified P wave may indicate enlargement of the atria, an enlarged Q wave may indicate a MI, and an enlarged suppressed or inverted Q wave often indicates enlarged ventricles. T waves often appear flatter when insufficient oxygen is being delivered to the myocardium. An elevation of the ST segment above baseline is often seen in patients with an acute MI and may appear depressed below the baseline when hypoxia is occurring.
As useful as analysing these electrical recordings may be, there are limitations. For example, not all areas suffering a MI may be obvious on the ECG. Additionally, it will not reveal the effectiveness of the pumping, which requires further testing, such as an ultrasound test called an echocardiogram or nuclear medicine imaging. It is also possible for there to be pulseless electrical activity, which will show up on an ECG tracing, although there is no corresponding pumping action. Common abnormalities that may be detected by the ECGs are shown in Figure 6.2.9.
Figure 6.2.9. Common ECG abnormalities. (a) In a second-degree or partial block, one-half of the P waves are not followed by the QRS complex and T waves while the other half are. (b) In atrial fibrillation, the electrical pattern is abnormal prior to the QRS complex, and the frequency between the QRS complexes has increased. (c) In ventricular tachycardia, the shape of the QRS complex is abnormal. (d) In ventricular fibrillation, there is no normal electrical activity. (e) In a third-degree block, there is no correlation between atrial activity (the P wave) and ventricular activity (the QRS complex).
Everyday Connection
External Automated Defibrillators
If the electrical activity of the heart is severely disrupted, cessation of electrical activity or fibrillation may occur. In fibrillation, the heart beats in a wild, uncontrolled manner, which prevents it from being able to pump effectively. Atrial fibrillation (see Figure 6.2.9b) is a serious condition, but as long as the ventricles continue to pump blood, the patient’s life may not be in immediate danger. Ventricular fibrillation (see Figure 6.2.9d) is a medical emergency that requires life support, because the ventricles are not effectively pumping blood. In a hospital setting, it is often described as “code blue.” If untreated for as little as a few minutes, ventricular fibrillation may lead to brain death. The most common treatment is defibrillation, which uses special paddles to apply a charge to the heart from an external electrical source in an attempt to establish a normal sinus rhythm (Figure 6.2.10). A defibrillator effectively stops the heart so that the SA node can trigger a normal conduction cycle. Because of their effectiveness in re-establishing a normal sinus rhythm, external automated defibrillators (EADs) are being placed in areas frequented by large numbers of people, such as schools, restaurants, and airports. These devices contain simple and direct verbal instructions that can be followed by nonmedical personnel to save a life.
Figure 6.2.10. Defibrillators. (a) An external automatic defibrillator can be used by nonmedical personnel to re-establish a normal sinus rhythm in a person with fibrillation. (b) Defibrillator paddles are more commonly used in hospital settings. (credit b: “widerider107”/flickr.com).
A heart block refers to an interruption in the normal conduction pathway. The nomenclature for these is very straightforward. SA nodal blocks occur within the SA node. AV nodal blocks occur within the AV node. Infra-Hisian blocks involve the bundle of His. Bundle branch blocks occur within either the left or right atrioventricular bundle branches. Hemiblocks are partial and occur within one or more fascicles of the atrioventricular bundle branch. Clinically, the most common types are the AV nodal and infra-Hisian blocks.
AV blocks are often described by degrees. A first-degree or partial block indicates a delay in conduction between the SA and AV nodes. This can be recognised on the ECG as an abnormally long PR interval. A second-degree or incomplete block occurs when some impulses from the SA node reach the AV node and continue, while others do not. In this instance, the ECG would reveal some P waves not followed by a QRS complex, while others would appear normal. In the third-degree or complete block, there is no correlation between atrial activity (the P wave) and ventricular activity (the QRS complex). Even in the event of a total SA block, the AV node will assume the role of pacemaker and continue initiating contractions at 40–60 contractions per minute, which is adequate to maintain consciousness.
When arrhythmias become a chronic problem, the heart maintains a junctional rhythm, which originates in the AV node. In order to speed up the heart rate and restore full sinus rhythm, a cardiologist can implant an artificial pacemaker, which delivers electrical impulses to the heart muscle to ensure that the heart continues to contract and pump blood effectively. These artificial pacemakers are programmable by the cardiologists and can either provide stimulation temporarily upon demand or on a continuous basis. Some devices also contain built-in defibrillators.
Cardiac Muscle Metabolism
Normally, cardiac muscle metabolism is entirely aerobic. Oxygen from the lungs is brought to the heart, and every other organ, attached to the haemoglobin molecules within the erythrocytes. Heart cells also store appreciable amounts of oxygen in myoglobin. Normally, these two mechanisms, circulating oxygen and oxygen attached to myoglobin, can supply sufficient oxygen to the heart, even during peak performance.
Fatty acids and glucose from the circulation are broken down within the mitochondria to release energy in the form of ATP. Both fatty acid droplets and glycogen are stored within the sarcoplasm and provide additional nutrient supply.
Section Review
The heart is regulated by both neural and endocrine control, yet it can initiate its own action potential followed by muscular contraction. The conductive cells within the heart establish the heart rate and transmit it through the myocardium. The contractile cells contract and propel the blood. The normal path of transmission for the conductive cells is the sinoatrial (SA) node, internodal pathways, atrioventricular (AV) node, atrioventricular (AV) bundle of His, bundle branches, and Purkinje fibres. The action potential for the conductive cells consists of a prepotential phase with a slow influx of Na+ followed by a rapid influx of Ca2+ and outflux of K+. Contractile cells have an action potential with an extended plateau phase that results in an extended refractory period to allow complete contraction for the heart to pump blood effectively. Recognisable points on the ECG include the P wave that corresponds to atrial depolarisation, the QRS complex that corresponds to ventricular depolarisation and the T wave that corresponds to ventricular repolarisation.
Describe the relationship between blood pressure and blood flow
Summarise the events of the cardiac cycle
Compare atrial and ventricular systole and diastole
Relate heart sounds detected by auscultation to action of heart’s valves
The period of time that begins with contraction of the atria and ends with ventricular relaxation is known as the cardiac cycle (Figure 6.3.1). The period of contraction that the heart undergoes while it pumps blood into circulation is called systole. The period of relaxation that occurs as the chambers fill with blood is called diastole. Both the atria and ventricles undergo systole and diastole, and it is essential that these components be carefully regulated and coordinated to ensure blood is pumped efficiently to the body.
Figure 6.3.1. Overview of the cardiac cycle. The cardiac cycle begins with atrial systole and progresses to ventricular systole, atrial diastole, and ventricular diastole, when the cycle begins again. Correlations to the ECG are highlighted.
Pressures and Flow
Fluids, whether gases or liquids, are materials that flow according to pressure gradients—that is, they move from regions that are higher in pressure to regions that are lower in pressure. Accordingly, when the heart chambers are relaxed (diastole), blood will flow into the atria from the veins, which are higher in pressure. As blood flows into the atria, the pressure will rise, so the blood will initially move passively from the atria into the ventricles. When the action potential triggers the muscles in the atria to contract (atrial systole), the pressure within the atria rises further, pumping blood into the ventricles. During ventricular systole, pressure rises in the ventricles, pumping blood into the pulmonary trunk from the right ventricle and into the aorta from the left ventricle. Again, as you consider this flow and relate it to the conduction pathway, the elegance of the system should become apparent.
Phases of the Cardiac Cycle
At the beginning of the cardiac cycle, both the atria and ventricles are relaxed (diastole). Blood is flowing into the right atrium from the superior and inferior venae cavae and the coronary sinus. Blood flows into the left atrium from the four pulmonary veins. The two atrioventricular valves, the tricuspid and mitral valves, are both open, so blood flows unimpeded from the atria and into the ventricles. Approximately 70–80 percent of ventricular filling occurs by this method. The two semilunar valves, the pulmonary and aortic valves, are closed, preventing backflow of blood into the right and left ventricles from the pulmonary trunk on the right and the aorta on the left.
Atrial Systole and Diastole
Contraction of the atria follows depolarisation, represented by the P wave of the ECG. As the atrial muscles contract from the superior portion of the atria toward the atrioventricular septum, pressure rises within the atria and blood is pumped into the ventricles through the open atrioventricular (tricuspid, and mitral or bicuspid) valves. At the start of atrial systole, the ventricles are normally filled with approximately 70–80 percent of their capacity due to inflow during diastole. Atrial contraction, also referred to as the “atrial kick,” contributes the remaining 20–30 percent of filling (see Figure 6.3.1). Atrial systole lasts approximately 100 ms and ends prior to ventricular systole, as the atrial muscle returns to diastole.
Ventricular Systole
Ventricular systole (see Figure 6.3.1) follows the depolarisation of the ventricles and is represented by the QRS complex in the ECG. It may be conveniently divided into two phases, lasting a total of 270 ms. At the end of atrial systole and just prior to atrial contraction, the ventricles contain approximately 130 mL blood in a resting adult in a standing position. This volume is known as the end diastolic volume (EDV) or preload.
Initially, as the muscles in the ventricle contract, the pressure of the blood within the chamber rises, but it is not yet high enough to open the semilunar (pulmonary and aortic) valves and be ejected from the heart. However, blood pressure quickly rises above that of the atria that are now relaxed and in diastole. This increase in pressure causes blood to flow back toward the atria, closing the tricuspid and mitral valves. Since blood is not being ejected from the ventricles at this early stage, the volume of blood within the chamber remains constant. Consequently, this initial phase of ventricular systole is known as isovolumic contraction, also called isovolumetric contraction (see Figure 6.3.1).
In the second phase of ventricular systole, the ventricular ejection phase, the contraction of the ventricular muscle has raised the pressure within the ventricle to the point that it is greater than the pressures in the pulmonary trunk and the aorta. Blood is pumped from the heart, pushing open the pulmonary and aortic semilunar valves. Pressure generated by the left ventricle will be appreciably greater than the pressure generated by the right ventricle, since the existing pressure in the aorta will be so much higher. Nevertheless, both ventricles pump the same amount of blood. This quantity is referred to as stroke volume. Stroke volume will normally be in the range of 70–80 mL. Since ventricular systole began with an EDV of approximately 130 mL of blood, this means that there is still 50–60 mL of blood remaining in the ventricle following contraction. This volume of blood is known as the end systolic volume (ESV).
Ventricular Diastole
Ventricular relaxation, or diastole, follows repolarisation of the ventricles and is represented by the T wave of the ECG. It too is divided into two distinct phases and lasts approximately 430 ms.
During the early phase of ventricular diastole, as the ventricular muscle relaxes, pressure on the remaining blood within the ventricle begins to fall. When pressure within the ventricles drops below pressure in both the pulmonary trunk and aorta, blood flows back toward the heart, producing the dicrotic notch (small dip) seen in blood pressure tracings. The semilunar valves close to prevent backflow into the heart. Since the atrioventricular valves remain closed at this point, there is no change in the volume of blood in the ventricle, so the early phase of ventricular diastole is called the isovolumic ventricular relaxation phase, also called isovolumetric ventricular relaxation phase (see Figure 6.3.1).
In the second phase of ventricular diastole, called late ventricular diastole, as the ventricular muscle relaxes, pressure on the blood within the ventricles drops even further. Eventually, it drops below the pressure in the atria. When this occurs, blood flows from the atria into the ventricles, pushing open the tricuspid and mitral valves. As pressure drops within the ventricles, blood flows from the major veins into the relaxed atria and from there into the ventricles. Both chambers are in diastole, the atrioventricular valves are open, and the semilunar valves remain closed (see Figure 6.3.1). The cardiac cycle is complete.
Figure 6.3.2 illustrates the relationship between the cardiac cycle and the ECG.
Figure 6.3.2. Relationship between the cardiac cycle and ECG. Initially, both the atria and ventricles are relaxed (diastole). The P wave represents depolarisation of the atria and is followed by atrial contraction (systole). Atrial systole extends until the QRS complex, at which point, the atria relax. The QRS complex represents depolarisation of the ventricles and is followed by ventricular contraction. The T wave represents the repolarisation of the ventricles and marks the beginning of ventricular relaxation.
Heart Sounds
One of the simplest, yet effective, diagnostic techniques applied to assess the state of a patient’s heart is auscultation using a stethoscope.
In a normal, healthy heart, there are only two audible heart sounds: S1 and S2. S1 is the sound created by the closing of the atrioventricular valves during ventricular contraction and is normally described as a “lub,” or first heart sound. The second heart sound, S2, is the sound of the closing of the semilunar valves during ventricular diastole and is described as a “dub” (Figure 6.3.3). In both cases, as the valves close, the openings within the atrioventricular septum guarded by the valves will become reduced, and blood flow through the opening will become more turbulent until the valves are fully closed. There is a third heart sound, S3, but it is rarely heard in healthy individuals. It may be the sound of blood flowing into the atria, or blood sloshing back and forth in the ventricle, or even tensing of the chordae tendineae. S3 may be heard in youth, some athletes, and pregnant women. If the sound is heard later in life, it may indicate congestive heart failure, warranting further tests. Some cardiologists refer to the collective S1, S2, and S3 sounds as the “Kentucky gallop,” because they mimic those produced by a galloping horse. The fourth heart sound, S4, results from the contraction of the atria pushing blood into a stiff or hypertrophic ventricle, indicating failure of the left ventricle. S4 occurs prior to S1 and the collective sounds S4, S1, and S2 are referred to by some cardiologists as the “Tennessee gallop,” because of their similarity to the sound produced by a galloping horse with a different gait. A few individuals may have both S3 and S4, and this combined sound is referred to as S7.
Figure 6.3.3. Heart sounds and the cardiac cycle. In this illustration, the x-axis reflects time with a recording of the heart sounds. The y-axis represents pressure.
The term murmur is used to describe an unusual sound coming from the heart that is caused by the turbulent flow of blood. Murmurs are graded on a scale of 1 to 6, with 1 being the most common, the most difficult sound to detect, and the least serious. The most severe is a 6. Phonocardiograms or auscultograms can be used to record both normal and abnormal sounds using specialised electronic stethoscopes.
During auscultation, it is common practice for the clinician to ask the patient to breathe deeply. This procedure not only allows for listening to airflow, but it may also amplify heart murmurs. Inhalation increases blood flow into the right side of the heart and may increase the amplitude of right-sided heart murmurs. Expiration partially restricts blood flow into the left side of the heart and may amplify left-sided heart murmurs. Figure 6.3.4 indicates proper placement of the bell of the stethoscope to facilitate auscultation.
Figure 6.3.4. Stethoscope placement for auscultation. Proper placement of the bell of the stethoscope facilitates auscultation. At each of the four locations on the chest, a different valve can be heard.
Section Review
The cardiac cycle comprises a complete relaxation and contraction of both the atria and ventricles and lasts approximately 0.8 seconds. Beginning with all chambers in diastole, blood flows passively from the veins into the atria and past the atrioventricular valves into the ventricles. The atria begin to contract (atrial systole), following depolarisation of the atria, and pump blood into the ventricles. The ventricles begin to contract (ventricular systole), raising pressure within the ventricles. When ventricular pressure rises above the pressure in the atria, blood flows toward the atria, producing the first heart sound, S1 or lub. As pressure in the ventricles rises above two major arteries, blood pushes open the two semilunar valves and moves into the pulmonary trunk and aorta in the ventricular ejection phase. Following ventricular repolarisation, the ventricles begin to relax (ventricular diastole), and pressure within the ventricles drops. As ventricular pressure drops, there is a tendency for blood to flow back into the atria from the major arteries, producing the dicrotic notch in the ECG and closing the two semilunar valves. The second heart sound, S2 or dub, occurs when the semilunar valves close. When the pressure falls below that of the atria, blood moves from the atria into the ventricles, opening the atrioventricular valves and marking one complete heart cycle. The valves prevent backflow of blood. Failure of the valves to operate properly produces turbulent blood flow within the heart; the resulting heart murmur can often be heard with a stethoscope.
Identify cardiovascular centres and cardiac reflexes that regulate heart function
Describe factors affecting heart rate
Distinguish between positive and negative factors that affect heart contractility
Summarise factors affecting stroke volume and cardiac output
Describe the cardiac response to variations in blood flow and pressure
The autorhythmicity inherent in cardiac cells keeps the heart beating at a regular pace; however, the heart is regulated by and responds to outside influences as well. Neural and endocrine controls are vital to the regulation of cardiac function. In addition, the heart is sensitive to several environmental factors, including electrolytes.
Resting Cardiac Output
Cardiac output (CO) is a measurement of the amount of blood pumped by each ventricle in one minute. To calculate this value, multiply stroke volume (SV), the amount of blood pumped by each ventricle, by heart rate (HR), in contractions per minute (or beats per minute, bpm). It can be represented mathematically by the following equation:
CO = HR × SV
SV is normally measured using an echocardiogram to record EDV and ESV and calculating the difference: SV = EDV – ESV. SV can also be measured using a specialised catheter, but this is an invasive procedure and far more dangerous to the patient. A mean SV for a resting 70-kg (150-lb) individual would be approximately 70 mL. There are several important variables, including size of the heart, physical and mental condition of the individual, sex, contractility, duration of contraction, preload or EDV, and afterload or resistance. Normal range for SV would be 55–100 mL. An average resting HR would be approximately 75 bpm but could range from 60–100 in some individuals.
Using these numbers, the mean CO is 5.25 L/min, with a range of 4.0–8.0 L/min. Remember, however, that these numbers refer to CO from each ventricle separately, not the total for the heart. Factors influencing CO are summarised in Figure 6.4.1.
Figure 6.4.1. Major factors influencing cardiac output. Cardiac output is influenced by heart rate and stroke volume, both of which are also variable.
SVs are also used to calculate ejection fraction, which is the portion of the blood that is pumped or ejected from the heart with each contraction. To calculate ejection fraction, SV is divided by EDV. Despite the name, the ejection fraction is normally expressed as a percentage. Ejection fractions range from approximately 55–70 percent, with a mean of 58 percent.
Exercise and Maximum Cardiac Output
In healthy young individuals, HR may increase to 150 bpm during exercise. SV can also increase from 70 to approximately 130 mL due to increased strength of contraction. This would increase CO to approximately 19.5 L/min, 4–5 times the resting rate. Top cardiovascular athletes can achieve even higher levels. At their peak performance, they may increase resting CO by 7–8 times.
Since the heart is a muscle, exercising it increases its efficiency. The difference between maximum and resting CO is known as the cardiac reserve. It measures the residual capacity of the heart to pump blood.
Heart Rates
HRs vary considerably, not only with exercise and fitness levels, but also with age. Newborn resting HRs may be 120 bpm. HR gradually decreases until young adulthood and then gradually increases again with age.
Maximum HRs are normally in the range of 200–220 bpm, although there are some extreme cases in which they may reach higher levels. As one ages, the ability to generate maximum rates decreases. This may be estimated by taking the maximal value of 220 bpm and subtracting the individual’s age. So, a 40-year-old individual would be expected to hit a maximum rate of approximately 180, and a 60-year-old person would achieve a HR of 160.
Disorders of the Heart: Abnormal Heart Rates
For an adult, normal resting HR will be in the range of 60–100 bpm. Bradycardia is the condition in which resting rate drops below 60 bpm, and tachycardia is the condition in which the resting rate is above 100 bpm. Trained athletes typically have very low HRs. If the patient is not exhibiting other symptoms, such as weakness, fatigue, dizziness, fainting, chest discomfort, palpitations, or respiratory distress, bradycardia is not considered clinically significant. However, if any of these symptoms are present, they may indicate that the heart is not providing sufficient oxygenated blood to the tissues. The term relative bradycardia may be used with a patient who has a HR in the normal range but is still suffering from these symptoms. Most patients remain asymptomatic if the HR remains above 50 bpm.
Bradycardia may be caused by either inherent factors or causes external to the heart. While the condition may be inherited, typically it is acquired in older individuals. Inherent causes include abnormalities in either the SA or AV node. If the condition is serious, a pacemaker may be required. Other causes include ischaemia to the heart muscle or diseases of the heart vessels or valves. External causes include metabolic disorders, pathologies of the endocrine system often involving the thyroid, electrolyte imbalances, neurological disorders including inappropriate autonomic responses, autoimmune pathologies, over-prescription of beta blocker drugs that reduce HR, recreational drug use, or even prolonged bed rest. Treatment relies upon establishing the underlying cause of the disorder and may necessitate supplemental oxygen.
Tachycardia is not normal in a resting patient but may be detected in pregnant women or individuals experiencing extreme stress. In the latter case, it would likely be triggered by stimulation from the limbic system or disorders of the autonomic nervous system. In some cases, tachycardia may involve only the atria. Some individuals may remain asymptomatic, but when present, symptoms may include dizziness, shortness of breath, light-headedness, rapid pulse, heart palpations, chest pain, or fainting (syncope). While tachycardia is defined as a HR above 100 bpm, there is considerable variation among people. Further, the normal resting HRs of children are often above 100 bpm, but this is not considered to be tachycardia Many causes of tachycardia may be benign, but the condition may also be correlated with fever, anaemia, hypoxia, hyperthyroidism, hypersecretion of catecholamines, some cardiomyopathies, some disorders of the valves, and acute exposure to radiation. Elevated rates in an exercising or resting patient are normal and expected. Resting rate should always be taken after recovery from exercise. Treatment depends upon the underlying cause but may include medications, implantable cardioverter defibrillators, ablation or surgery.
Correlation Between Heart Rates and Cardiac Output
Initially, physiological conditions that cause HR to increase also trigger an increase in SV. During exercise, the rate of blood returning to the heart increases. However, as the HR rises, there is less time spent in diastole and consequently less time for the ventricles to fill with blood. Even though there is less filling time, SV will initially remain high. However, as HR continues to increase, SV gradually decreases due to decreased filling time. CO will initially stabilise as the increasing HR compensates for the decreasing SV, but at very high rates, CO will eventually decrease as increasing rates are no longer able to compensate for the decreasing SV. Consider this phenomenon in a healthy young individual. Initially, as HR increases from resting to approximately 120 bpm, CO will rise. As HR increases from 120 to 160 bpm, CO remains stable, since the increase in rate is offset by decreasing ventricular filling time and, consequently, SV. As HR continues to rise above 160 bpm, CO decreases as SV falls faster than HR increases. So, although aerobic exercises are critical to maintain the health of the heart, individuals are cautioned to monitor their HR to ensure they stay within the target heart rate range of between 120 and 160 bpm, so CO is maintained. The target HR is loosely defined as the range in which both the heart and lungs receive the maximum benefit from the aerobic workout and is dependent upon age.
Cardiovascular Centres
Nervous control over HR is centralised within the two paired cardiovascular centres of the medulla oblongata (Figure 6.4.2). The cardioaccelerator regions stimulate activity via sympathetic stimulation of the cardioaccelerator nerves, and the cardioinhibitory centres decrease heart activity via parasympathetic stimulation as one component of the vagus nerve, cranial nerve X. During rest, both centres provide slight stimulation to the heart, contributing to autonomic tone. This is a similar concept to tone in skeletal muscles. Normally, vagal stimulation predominates as, left unregulated, the SA node would initiate a sinus rhythm of approximately 100 bpm.
Both sympathetic and parasympathetic stimulations flow through a paired complex network of nerve fibres known as the cardiac plexus near the base of the heart. The cardioaccelerator centres also sends additional fibres, forming the cardiac nerves via sympathetic ganglia (the cervical ganglia plus superior thoracic ganglia T1–T4) to both the SA and AV nodes, plus additional fibres to the atria and ventricles. The ventricles are more richly innervated by sympathetic fibres than parasympathetic fibres. Sympathetic stimulation causes the release of the neurotransmitter noradrenaline (NA) at the neuromuscular junction of the cardiac nerves. NA shortens the repolarisation period, thus speeding the rate of depolarisation and contraction, which results in an increase in HR. It opens chemical- or ligand-gated sodium and calcium ion channels, allowing an influx of positively charged ions.
NA binds to the beta-1 receptor. Some cardiac medications (for example, beta blockers) work by blocking these receptors, thereby slowing HR and are one possible treatment for hypertension. Over prescription of these drugs may lead to bradycardia and even stoppage of the heart.
Figure 6.4.2. Autonomic innervation of the heart. Cardioaccelerator and cardioinhibitory areas are components of the paired cardiac centres located in the medulla oblongata of the brain. They innervate the heart via sympathetic cardiac nerves that increase cardiac activity and vagus (parasympathetic) nerves that slow cardiac activity.
Parasympathetic stimulation originates from the cardioinhibitory region with impulses traveling via the vagus nerve (cranial nerve X). The vagus nerve sends branches to both the SA and AV nodes, and to portions of both the atria and ventricles. Parasympathetic stimulation releases the neurotransmitter acetylcholine (ACh) at the neuromuscular junction. ACh slows HR by opening chemical- or ligand-gated potassium ion channels to slow the rate of spontaneous depolarisation, which extends repolarisation and increases the time before the next spontaneous depolarisation occurs. Without any nervous stimulation, the SA node would establish a sinus rhythm of approximately 100 bpm. Since resting rates are considerably less than this, it becomes evident that parasympathetic stimulation normally slows HR. This is like an individual driving a car with one foot on the brake pedal. To speed up, one need merely remove one’s foot from the break and let the engine increase speed. In the case of the heart, decreasing parasympathetic stimulation decreases the release of ACh, which allows HR to increase up to approximately 100 bpm. Any increases beyond this rate would require sympathetic stimulation. Figure 6.4.3 illustrates the effects of parasympathetic and sympathetic stimulation on the normal sinus rhythm.
Figure 6.4.3. Effects of parasympathetic and sympathetic stimulation on normal sinus rhythm. The wave of depolarisation in a normal sinus rhythm shows a stable resting HR. Following parasympathetic stimulation, HR slows. Following sympathetic stimulation, HR increases.
Input to the Cardiovascular Centre
The cardiovascular centre receives input from a series of visceral receptors with impulses traveling through visceral sensory fibres within the vagus and sympathetic nerves via the cardiac plexus. Among these receptors are various proprioreceptors, baroreceptors, and chemoreceptors, plus stimuli from the limbic system. Collectively, these inputs normally enable the cardiovascular centre to regulate heart function precisely, a process known as cardiac reflexes. Increased physical activity results in increased rates of firing by various proprioreceptors located in muscles, joint capsules, and tendons. Any such increase in physical activity would logically warrant increased blood flow. The cardiac centre monitor these increased rates of firing and suppress parasympathetic stimulation and increase sympathetic stimulation as needed in order to increase blood flow.
Similarly, baroreceptors are stretch receptors located in the aortic sinus, carotid bodies, the venae cavae, and other locations, including pulmonary vessels and the right side of the heart itself. Rates of firing from the baroreceptors represent blood pressure, level of physical activity, and the relative distribution of blood. The cardiac centre monitor baroreceptor firing to maintain cardiac homeostasis, a mechanism called the baroreceptor reflex. With increased pressure and stretch, the rate of baroreceptor firing increases, and the cardiac centre decrease sympathetic stimulation and increase parasympathetic stimulation. As pressure and stretch decrease, the rate of baroreceptor firing decreases, and the cardiac centre increase sympathetic stimulation and decrease parasympathetic stimulation.
There is a similar reflex, called the atrial reflex or Bainbridge reflex, associated with varying rates of blood flow to the atria. Increased venous return stretches the walls of the atria where specialised baroreceptors are located. However, as the atrial baroreceptors increase their rate of firing and as they stretch due to the increased blood pressure, the cardiac centre responds by increasing sympathetic stimulation and inhibiting parasympathetic stimulation to increase HR. The opposite is also true.
Increased metabolic by-products associated with increased activity, such as carbon dioxide, hydrogen ions, and lactic acid, plus falling oxygen levels, are detected by a suite of chemoreceptors innervated by the glossopharyngeal and vagus nerves. These chemoreceptors provide feedback to the cardiovascular centre about the need for increased or decreased blood flow, based on the relative levels of these substances.
The limbic system can also significantly impact HR related to emotional state. During periods of stress, it is not unusual to identify higher than normal HRs, often accompanied by a surge in the stress hormone cortisol. Individuals experiencing extreme anxiety may manifest panic attacks with symptoms that resemble those of heart attacks. These events are typically transient and treatable. Meditation techniques have been developed to ease anxiety and have been shown to lower HR effectively. Doing simple deep and slow breathing exercises with one’s eyes closed can also significantly reduce this anxiety and HR.
Disorders of the Heart: Broken Heart Syndrome
Extreme stress from such life events as the death of a loved one, an emotional break up, loss of income, or foreclosure of a home may lead to a condition commonly referred to as broken heart syndrome. This condition may also be called Takotsubo cardiomyopathy, transient apical ballooning syndrome, apical ballooning cardiomyopathy, stress-induced cardiomyopathy, Gebrochenes-Herz syndrome, and stress cardiomyopathy. The recognised effects on the heart include congestive heart failure due to a profound weakening of the myocardium not related to lack of oxygen. This may lead to acute heart failure, lethal arrhythmias, or even the rupture of a ventricle. The exact aetiology is not known, but several factors have been suggested, including transient vasospasm, dysfunction of the cardiac capillaries, or thickening of the myocardium—particularly in the left ventricle—that may lead to the critical circulation of blood to this region. While many patients survive the initial acute event with treatment to restore normal function, there is a strong correlation with death. Careful statistical analysis by the Cass Business School, a prestigious institution located in London, published in 2008, revealed that within one year of the death of a loved one, women are more than twice as likely to die and males are six times as likely to die as would otherwise be expected.
Other Factors Influencing Heart Rate
Using a combination of autorhythmicity and innervation, the cardiovascular centre can provide relatively precise control over HR. However, there are a number of other factors that have an impact on HR as well, including adrenaline, noradrenaline and thyroid hormones; levels of various ions including calcium, potassium, and sodium; body temperature; hypoxia; and pH balance (Table 6.4.1 and Table 6.4.2). After reading this section, the importance of maintaining homeostasis should become even more apparent.
Table 6.4.1. Major factors increasing heart rate and force of contraction
Factor
Effect
Cardioaccelerator nerves
Release of noradrenaline (NA)
Proprioceptors
Increased rates of firing during exercise
Chemoreceptors
Decreased levels of O2, increased levels of H+, CO2 and lactic acid
Baroreceptors
Decreased rates of firing, indicating falling blood volume/pressure
Limbic system
Anticipation of physical exercise or strong emotions
Catecholamines
Increased adrenaline and noradrenaline
Thyroid hormones
Increased T3 and T4
Calcium
Increased Ca2+
Potassium
Decreased K+
Sodium
Decreased Na+
Body temperature
Increased body temperature
Nicotine and caffeine
Stimulants, increased heart rate
Table 6.4.2. Factors decreasing heart rate and force of contraction
Factor
Effect
Cardioinhibitory nerves (vagus)
Release of acetylcholine
Proprioceptors
Decreased rates of firing following exercise
Chemoreceptors
Increased levels of O2; decreased levels of H+ and CO2
Baroreceptors
Increased rates of firing, indicating higher blood volume/pressure
Limbic System
Anticipation of relaxation
Catecholamines
Decreased adrenaline and noradrenaline
Thyroid hormones
Decreased T3 and T4
Calcium
Decreased Ca2+
Potassium
Increased K+
Sodium
Increased Na+
Body temperature
Decrease in body temperature
Adrenaline (A) and Noradrenaline (NA)
The catecholamines, adrenaline and noradrenaline, secreted by the adrenal medulla form one component of the extended fight-or-flight mechanism. The other component is sympathetic stimulation. Adrenaline and NA have similar effects: binding to the beta-1 receptors, and opening sodium and calcium ion chemical- or ligand-gated channels. The rate of depolarisation is increased by this additional influx of positively charged ions, so the threshold is reached more quickly and the period of repolarisation is shortened. However, massive releases of these hormones coupled with sympathetic stimulation may actually lead to arrhythmias. There is no parasympathetic stimulation to the adrenal medulla.
Thyroid Hormones
In general, increased levels of thyroid hormone, or thyroxine, increase cardiac rate and contractility. The impact of thyroid hormone is typically of a much longer duration than that of the catecholamines. The physiologically active form of thyroid hormone, T3 or triiodothyronine, has been shown to directly enter cardiomyocytes and alter activity at the level of the genome. It also impacts the beta adrenergic response similar to adrenaline and NA described above. Excessive levels of thyroxine may trigger tachycardia.
Calcium
Calcium ion levels have great impacts upon both HR and contractility; as the levels of calcium ions increase, so do HR and contractility. High levels of calcium ions (hypercalcaemia) may be implicated in a short QT interval and a widened T wave in the ECG. The QT interval represents the time from the start of depolarisation to repolarisation of the ventricles and includes the period of ventricular systole. Extremely high levels of calcium may induce cardiac arrest. Drugs known as calcium channel blockers slow HR by binding to these channels and blocking or slowing the inward movement of calcium ions.
Caffeine and Nicotine
Caffeine and nicotine are not found naturally within the body. Both of these nonregulated drugs have an excitatory effect on membranes of neurons in general and have a stimulatory effect on the cardiac centres specifically, causing an increase in HR. Caffeine works by increasing the rates of depolarisation at the SA node, whereas nicotine stimulates the activity of the sympathetic neurons that deliver impulses to the heart.
Although it is the world’s most widely consumed psychoactive drug, caffeine is legal and not regulated. While precise quantities have not been established, “normal” consumption is not considered harmful to most people, although it may cause disruptions to sleep and acts as a diuretic. Its consumption by pregnant women is cautioned against, although no evidence of negative effects has been confirmed. Tolerance and even physical and mental addiction to the drug result in individuals who routinely consume the substance.
Nicotine, too, is a stimulant and produces addiction. While legal and nonregulated, concerns about nicotine’s safety and documented links to respiratory and cardiac disease and cancer have resulted in warning labels on cigarette packages.
Factors Decreasing Heart Rate
HR can be slowed when a person experiences altered sodium and potassium levels, hypoxia, acidosis, alkalosis and hypothermia (see Table 6.4.2). The relationship between electrolytes and HR is complex but maintaining electrolyte balance is critical to the normal wave of depolarisation. Of the two ions, potassium has the greater clinical significance. Initially, both hyponatraemia (low sodium levels) and hypernatraemia (high sodium levels) may lead to tachycardia. Severely high hypernatraemia may lead to fibrillation, which may cause CO to cease. Severe hyponatraemia leads to both bradycardia and other arrhythmias. Hypokalaemia (low potassium levels) also leads to arrhythmias, whereas hyperkalaemia (high potassium levels) causes the heart to become weak and flaccid, and ultimately to fail.
Heart muscle relies exclusively on aerobic metabolism for energy. Hypoxia (an insufficient supply of oxygen) leads to decreasing HRs, since metabolic reactions fuelling heart contraction are restricted.
Acidosis is a condition in which excess hydrogen ions are present, and the patient’s blood expresses a low pH value. Alkalosis is a condition in which there are too few hydrogen ions, and the patient’s blood has an elevated pH. Normal blood pH falls in the range of 7.35–7.45, so a number lower than this range represents acidosis and a higher number represents alkalosis. Recall that enzymes are the regulators or catalysts of virtually all biochemical reactions; they are sensitive to pH and will change shape slightly with values outside their normal range. These variations in pH and accompanying slight physical changes to the active site on the enzyme decrease the rate of formation of the enzyme-substrate complex, subsequently decreasing the rate of many enzymatic reactions, which can have complex effects on HR. Severe changes in pH will lead to denaturation of the enzyme.
The last variable is body temperature. Elevated body temperature is called hyperthermia, and suppressed body temperature is called hypothermia. Slight hyperthermia results in increasing HR and strength of contraction. Hypothermia slows the rate and strength of heart contractions. This distinct slowing of the heart is one component of the larger diving reflex that diverts blood to essential organs while submerged. If sufficiently chilled, the heart will stop beating, a technique that may be employed during open heart surgery. In this case, the patient’s blood is normally diverted to an artificial heart-lung machine to maintain the body’s blood supply and gas exchange until the surgery is complete, and sinus rhythm can be restored. Excessive hyperthermia and hypothermia will both result in death, as enzymes drive the body systems to cease normal function, beginning with the central nervous system.
Stroke Volume
Many of the same factors that regulate HR also impact cardiac function by altering SV. While several variables are involved, SV is ultimately dependent upon the difference between EDV and ESV. The three primary factors to consider are preload, or the stretch on the ventricles prior to contraction; the contractility, or the force or strength of the contraction itself; and afterload, the force the ventricles must generate to pump blood against the resistance in the vessels. These factors are summarised in Table 6.4.1 and Table 6.4.2.
Preload
Preload is another way of expressing EDV. Therefore, the greater the EDV is, the greater the preload is. One of the primary factors to consider is filling time, or the duration of ventricular diastole during which filling occurs. The more rapidly the heart contracts, the shorter the filling time becomes, and the lower the EDV and preload are. This effect can be partially overcome by increasing the second variable, contractility, and raising SV, but over time, the heart is unable to compensate for decreased filling time, and preload also decreases.
With increasing ventricular filling, both EDV or preload increase, and the cardiac muscle itself is stretched to a greater degree. At rest, there is little stretch of the ventricular muscle, and the sarcomeres remain short. With increased ventricular filling, the ventricular muscle is increasingly stretched and the sarcomere length increases. As the sarcomeres reach their optimal lengths, they will contract more powerfully, because more of the myosin heads can bind to the actin on the thin filaments, forming cross bridges and increasing the strength of contraction and SV. If this process were to continue and the sarcomeres stretched beyond their optimal lengths, the force of contraction would decrease. However, due to the physical constraints of the location of the heart, this excessive stretch is not a concern.
The relationship between ventricular stretch and contraction has been stated in the well-known Frank-Starling mechanism or simply Starling’s Law of the Heart. This principle states that, within physiological limits, the force of heart contraction is directly proportional to the initial length of the muscle fibre. This means that the greater the stretch of the ventricular muscle (within limits), the more powerful the contraction is, which in turn increases SV. Therefore, by increasing preload, you increase the second variable, contractility.
Otto Frank (1865–1944) was a German physiologist; among his many published works are detailed studies of this important heart relationship. Ernest Starling (1866–1927) was an important English physiologist who also studied the heart. Although they worked largely independently, their combined efforts and similar conclusions have been recognised in the name “Frank-Starling mechanism.”
Any sympathetic stimulation to the venous system will increase venous return to the heart, which contributes to ventricular filling, and EDV and preload. While much of the ventricular filling occurs while both atria and ventricles are in diastole, the contraction of the atria, the atrial kick, plays a crucial role by providing the last 20–30 percent of ventricular filling.
Contractility
It is virtually impossible to consider preload or ESV without including an early mention of the concept of contractility. Indeed, the two parameters are intimately linked. Contractility refers to the force of the contraction of the heart muscle, which controls SV, and is the primary parameter for impacting ESV. The more forceful the contraction is, the greater the SV and smaller the ESV are. Less forceful contractions result in smaller SVs and larger ESVs. Factors that increase contractility are described as positive inotropic factors, and those that decrease contractility are described as negative inotropic factors (ino- = “fibre;” -tropic = “turning toward”).
Not surprisingly, sympathetic stimulation is a positive inotrope, whereas parasympathetic stimulation is a negative inotrope. Sympathetic stimulation triggers the release of NE at the neuromuscular junction from the cardiac nerves and stimulates the adrenal cortex to secrete adrenaline and NE. In addition to their stimulatory effects on HR, they also bind to both alpha and beta receptors on the cardiac muscle cell membrane to increase metabolic rate and the force of contraction. This combination of actions has the net effect of increasing SV and leaving a smaller residual ESV in the ventricles. In comparison, parasympathetic stimulation releases ACh at the neuromuscular junction from the vagus nerve. The membrane hyperpolarises and inhibits contraction to decrease the strength of contraction and SV, and to raise ESV. Since parasympathetic fibres are more widespread in the atria than in the ventricles, the primary site of action is in the upper chambers. Parasympathetic stimulation in the atria decreases the atrial kick and reduces EDV, which decreases ventricular stretch and preload, thereby further limiting the force of ventricular contraction. Stronger parasympathetic stimulation also directly decreases the force of contraction of the ventricles.
Several synthetic drugs, including dopamine and isoproterenol, have been developed that mimic the effects of adrenaline and NE by stimulating the influx of calcium ions from the extracellular fluid. Higher concentrations of intracellular calcium ions increase the strength of contraction. Excess calcium (hypercalcaemia) also acts as a positive inotropic agent. The drug digitalis lowers HR and increases the strength of the contraction, acting as a positive inotropic agent by blocking the sequestering of calcium ions into the sarcoplasmic reticulum. This leads to higher intracellular calcium levels and greater strength of contraction. In addition to the catecholamines from the adrenal medulla, other hormones also demonstrate positive inotropic effects. These include thyroid hormones and glucagon from the pancreas.
Negative inotropic agents include hypoxia, acidosis, hyperkalaemia, and a variety of synthetic drugs. These include numerous beta blockers and calcium channel blockers. Early beta blocker drugs include propranolol and pronethalol and are credited with revolutionising treatment of cardiac patients experiencing angina pectoris. There is also a large class of dihydropyridine, phenylalkylamine, and benzothiazepine calcium channel blockers that may be administered decreasing the strength of contraction and SV.
Afterload
Afterload refers to the tension that the ventricles must develop to pump blood effectively against the resistance in the vascular system. Any condition that increases resistance requires a greater afterload to force open the semilunar valves and pump the blood. Damage to the valves, such as stenosis, which makes them harder to open will also increase afterload. Any decrease in resistance decreases the afterload. Figure 6.4.4 summarises the major factors influencing SV, Figure 6.4.5 summarises the major factors influencing CO, and Table 6.4.3 and Table 6.4.4 summarise cardiac responses to increased and decreased blood flow and pressure in order to restore homeostasis.
Figure 6.4.4. Major factors influencing stroke volume. Multiple factors impact preload, afterload, and contractility, and are the major considerations influencing SV.Figure 6.4.5. Summary of major factors influencing cardiac output. The primary factors influencing HR include autonomic innervation plus endocrine control. Not shown are environmental factors, such as electrolytes, metabolic products, and temperature. The primary factors controlling SV include preload, contractility, and afterload. Other factors such as electrolytes may be classified as either positive or negative inotropic agents.
Table 6.4.3. Cardiac response to decreasing blood flow and pressure due to decreasing cardiac output
Baroreceptors (aorta, carotid arteries, venae cavae, and atria)
Chemoreceptors (both central nervous system and in proximity to baroreceptors)
Sensitive to
Decreasing stretch
Decreasing O2 and increasing CO2, H+ and lactic acid
Target
Parasympathetic stimulation suppressed
Sympathetic stimulation increased
Response of heart
Increasing heart rate and increasing stroke volume
Increasing heart rate and increasing stroke volume
Overall effect
Increasing blood flow and pressure due to increasing cardiac output; haemostasis restored
Increased blood flow and pressure due to increasing cardiac output; haemostasis restored
Table 6.4.4. Cardiac response to increasing blood flow and pressure due to increasing cardiac output
Baroreceptors (aorta, carotid arteries, venae cavae, and atria)
Chemoreceptors (both central nervous system and in proximity to baroreceptors)
Sensitive to
Increasing stretch
Increasing O2 and increasing CO2, H+ and lactic acid
Target
Parasympathetic stimulation increased
Sympathetic stimulation suppressed
Response of heart
Decreasing heart rate and decreasing stroke volume
Decreasing heart rate and decreasing stroke volume
Overall effect
Decreasing blood flow and pressure due to decreasing cardiac output; haemostasis restored
Decreasing blood flow and pressure due to decreasing cardiac output; haemostasis restored
Section Review
Many factors affect HR and SV, and together, they contribute to cardiac function. HR is largely determined and regulated by autonomic stimulation and hormones. There are several feedback loops that contribute to maintaining homeostasis dependent upon activity levels, such as the atrial reflex, which is determined by venous return.
SV is regulated by autonomic innervation and hormones, but also by filling time and venous return. Venous return is determined by activity of the skeletal muscles, blood volume, and changes in peripheral circulation. Venous return determines preload and the atrial reflex. Filling time directly related to HR also determines preload. Preload then impacts both EDV and ESV. Autonomic innervation and hormones largely regulate contractility. Contractility impacts EDV as does afterload. CO is the product of HR multiplied by SV. SV is the difference between EDV and ESV.
Describe the embryological development of heart structures
Identify five regions of the foetal heart
Relate foetal heart structures to adult counterparts
The human heart is the first functional organ to develop. It begins beating and pumping blood around day 21 or 22, a mere three weeks after fertilisation. This emphasises the critical nature of the heart in distributing blood through the vessels and the vital exchange of nutrients, oxygen, and wastes both to and from the developing baby. The critical early development of the heart is reflected by the prominent heart bulge that appears on the anterior surface of the embryo.
The heart forms from an embryonic tissue called mesoderm around 18 to 19 days after fertilisation. Mesoderm is one of the three primary germ layers that differentiates early in development that collectively gives rise to all subsequent tissues and organs. The heart begins to develop near the head of the embryo in a region known as the cardiogenic area. Following chemical signals called factors from the underlying endoderm (another of the three primary germ layers), the cardiogenic area begins to form two strands called the cardiogenic cords (Figure 6.5.1). As the cardiogenic cords develop, a lumen rapidly develops within them. At this point, they are referred to as endocardial tubes. The two tubes migrate together and fuse to form a single primitive heart tube. The primitive heart tube quickly forms five distinct regions. From head to tail, these include the truncus arteriosus, bulbus cordis, primitive ventricle, primitive atrium, and the sinus venosus. Initially, all venous blood flows into the sinus venosus, and contractions propel the blood from tail to head, or from the sinus venosus to the truncus arteriosus. This is a very different pattern from that of an adult.
Figure 6.5.1. Development of the human heart. This diagram outlines the embryological development of the human heart during the first eight weeks and the subsequent formation of the four heart chambers.
The five regions of the primitive heart tube develop into recognisable structures in a fully developed heart. The truncus arteriosus will eventually divide and give rise to the ascending aorta and pulmonary trunk. The bulbus cordis develops into the right ventricle. The primitive ventricle forms the left ventricle. The primitive atrium becomes the anterior portions of both the right and left atria, and the two auricles. The sinus venosus develops into the posterior portion of the right atrium, the SA node, and the coronary sinus.
As the primitive heart tube elongates, it begins to fold within the pericardium, eventually forming an S shape, which places the chambers and major vessels into an alignment like the adult heart. This process occurs between days 23 and 28. The remainder of the heart development pattern includes development of septa and valves and remodelling of the actual chambers. Partitioning of the atria and ventricles by the interatrial septum, interventricular septum, and atrioventricular septum is complete by the end of the fifth week, although the foetal blood shunts remain until birth or shortly after. The atrioventricular valves form between weeks five and eight, and the semilunar valves form between weeks five and nine.
Section Review
The heart is the first organ to form and become functional, emphasising the importance of transport of material to and from the developing infant. It originates about day 18 or 19 from the mesoderm and begins beating and pumping blood about day 21 or 22. It forms from the cardiogenic region near the head and is visible as a prominent heart bulge on the surface of the embryo. Originally, it consists of a pair of strands called cardiogenic cords that quickly form a hollow lumen and are referred to as endocardial tubes. These then fuse into a single heart tube and differentiate into the truncus arteriosus, bulbus cordis, primitive ventricle, primitive atrium, and sinus venosus, starting about day 22. The primitive heart begins to form an S shape within the pericardium between days 23 and 28. The internal septa begin to form about day 28, separating the heart into the atria and ventricles, although the foramen ovale persists until shortly after birth. Between weeks five and eight, the atrioventricular valves form. The semilunar valves form between weeks five and nine.
Compare and contrast the three tunics that make up the walls of most blood vessels
Distinguish between elastic arteries, muscular arteries, and arterioles on the basis of structure, location, and function
Describe the basic structure of a capillary bed, from the supplying metarteriole to the venule into which it drains
Explain the structure and function of venous valves in the large veins of the extremities
Blood is carried through the body via blood vessels. An artery is a blood vessel that carries blood away from the heart, where it branches into ever-smaller vessels. Eventually, the smallest arteries, vessels called arterioles, further branch into tiny capillaries, where nutrients and wastes are exchanged, and then combine with other vessels that exit capillaries to form venules, small blood vessels that carry blood to a vein, a larger blood vessel that returns blood to the heart.
Arteries and veins transport blood in two distinct circuits: the systemic circuit and the pulmonary circuit (Figure 6.6.1). Systemic arteries provide blood rich in oxygen to the body’s tissues. The blood returned to the heart through systemic veins has less oxygen, since much of the oxygen carried by the arteries has been delivered to the cells. In contrast, in the pulmonary circuit, arteries carry blood low in oxygen exclusively to the lungs for gas exchange. Pulmonary veins then return freshly oxygenated blood from the lungs to the heart to be pumped back out into systemic circulation. Although arteries and veins differ structurally and functionally, they share certain features.
Figure 6.6.1. Cardiovascular circulation. The pulmonary circuit moves blood from the right side of the heart to the lungs and back to the heart. The systemic circuit moves blood from the left side of the heart to the head and body and returns it to the right side of the heart to repeat the cycle. The arrows indicate the direction of blood flow, and the colours show the relative levels of oxygen concentration.
Shared Structures
Different types of blood vessels vary slightly in their structures, but they share the same general features. Arteries and arterioles have thicker walls than veins and venules because they are closer to the heart and receive blood that is surging at a far greater pressure (Figure 6.6.2). Each type of vessel has a lumen—a hollow passageway through which blood flows. Arteries have smaller lumens than veins, a characteristic that helps to maintain the pressure of blood moving through the system. Together, their thicker walls and smaller diameters give arterial lumens a more rounded appearance in cross section than the lumens of veins.
By the time blood has passed through capillaries and entered venules, the pressure initially exerted upon it by heart contractions has diminished. In other words, in comparison to arteries, venules and veins withstand a much lower pressure from the blood that flows through them. Their walls are considerably thinner and their lumens are correspondingly larger in diameter, allowing more blood to flow with less vessel resistance. In addition, many veins of the body, particularly those of the limbs, contain valves that assist the unidirectional flow of blood toward the heart. This is critical because blood flow becomes sluggish in the extremities, as a result of the lower pressure and the effects of gravity.
The walls of arteries and veins are largely composed of living cells and their products (including collagenous and elastic fibres); the cells require nourishment and produce waste. Since blood passes through the larger vessels relatively quickly, there is limited opportunity for blood in the lumen of the vessel to provide nourishment to or remove waste from the vessel’s cells. Further, the walls of the larger vessels are too thick for nutrients to diffuse through to all of the cells. Larger arteries and veins contain small blood vessels within their walls known as the vasa vasorum—literally “vessels of the vessel”—to provide them with this critical exchange. Since the pressure within arteries is relatively high, the vasa vasorum must function in the outer layers of the vessel (see Figure 6.2.2) or the pressure exerted by the blood passing through the vessel would collapse it, preventing any exchange from occurring. The lower pressure within veins allows the vasa vasorum to be located closer to the lumen. The restriction of the vasa vasorum to the outer layers of arteries is thought to be one reason that arterial diseases are more common than venous diseases, since its location makes it more difficult to nourish the cells of the arteries and remove waste products. There are also minute nerves within the walls of both types of vessels that control the contraction and dilation of smooth muscle. These minute nerves are known as the nervi vasorum.
Both arteries and veins have the same three distinct tissue layers, called tunics (from the Latin term tunica), for the garments first worn by ancient Romans; the term tunic is also used for some modern garments. From the most interior layer to the outer, these tunics are the tunica intima, the tunica media, and the tunica externa (see Figure 6.6.2). Table 6.6.1 compares and contrasts the tunics of the arteries and veins.
Table 6.6.1. Comparison of tunics in arteries and veins
Arteries
Veins
General appearance
Thick walls with small lumens
Thin walls with large lumens
Generally appear rounded
Generally appear flattened
Tunica intima
Endothelium usually appears wavy due to constriction of smooth muscle
Endothelium appears smooth
Internal elastic membrane present in larger vessels
Internal elastic membrane absent
Tunica media
Normally the thickest layer in arteries
Normally thinner than the tunica externa
Smooth muscle cells and elastic fibres predominate (the proportions of these vary with distance from the heart)
Smooth muscle cells and collagenous fibres predominate
External elastic membrane present in larger vessels
Nervi vasorum and vasa vasorum present
External elastic membrane absent
Tunica externa
Normally thinner than the tunica media in all but the largest arteries
Normally the thicket layer in veins
Collagenous and elastic fibres
Some smooth muscle fibres
Nervi vasorum and vasa vasorum present
Nervi vasorum and vasa vasorum present
Tunica Intima
The tunica intima (also called the tunica interna) is composed of epithelial and connective tissue layers. Lining the tunica intima is the specialised simple squamous epithelium called the endothelium, which is continuous throughout the entire vascular system, including the lining of the chambers of the heart. Damage to this endothelial lining and exposure of blood to the collagenous fibres beneath is one of the primary causes of clot formation. Until recently, the endothelium was viewed simply as the boundary between the blood in the lumen and the walls of the vessels. Recent studies, however, have shown that it is physiologically critical to such activities as helping to regulate capillary exchange and altering blood flow. The endothelium releases local chemicals called endothelins that can constrict the smooth muscle within the walls of the vessel to increase blood pressure. Uncompensated overproduction of endothelins may contribute to hypertension (high blood pressure) and cardiovascular disease.
Next to the endothelium is the basement membrane, or basal lamina, that effectively binds the endothelium to the connective tissue. The basement membrane provides strength while maintaining flexibility, and it is permeable, allowing materials to pass through it. The thin outer layer of the tunica intima contains a small amount of areolar connective tissue that consists primarily of elastic fibres to provide the vessel with additional flexibility; it also contains some collagenous fibres to provide additional strength.
In larger arteries, there is also a thick, distinct layer of elastic fibres known as the internal elastic membrane (also called the internal elastic lamina) at the boundary with the tunica media. Like the other components of the tunica intima, the internal elastic membrane provides structure while allowing the vessel to stretch. It is permeated with small openings that allow exchange of materials between the tunics. The internal elastic membrane is not apparent in veins. In addition, many veins, particularly in the lower limbs, contain valves formed by sections of thickened endothelium that are reinforced with connective tissue, extending into the lumen.
Under the microscope, the lumen and the entire tunica intima of a vein will appear smooth, whereas those of an artery will normally appear wavy because of the partial constriction of the smooth muscle in the tunica media, the next layer of blood vessel walls.
Tunica Media
The tunica media is the substantial middle layer of the vessel wall (see Figure 6.6.2). It is generally the thickest layer in arteries, and it is much thicker in arteries than it is in veins. The tunica media consists of layers of smooth muscle supported by connective tissue that is primarily made up of elastic fibres, most of which are arranged in circular sheets. Toward the outer portion of the tunic, there are also layers of longitudinal muscle. Contraction and relaxation of the circular muscles decrease and increase the diameter of the vessel lumen, respectively. Specifically, in arteries vasoconstriction decreases blood flow as the smooth muscle in the walls of the tunica media contracts, making the lumen narrower and increasing blood pressure. Similarly, vasodilation increases blood flow as the smooth muscle relaxes, allowing the lumen to widen and blood pressure to drop. Both vasoconstriction and vasodilation are regulated in part by small vascular nerves, known as nervi vasorum, or “nerves of the vessel,” that run within the walls of blood vessels. These are generally all sympathetic fibres, although some trigger vasodilation and others induce vasoconstriction, depending upon the nature of the neurotransmitter and receptors located on the target cell. Parasympathetic stimulation does trigger vasodilation as well as erection during sexual arousal in the external genitalia of both sexes. Nervous control over vessels tends to be more generalised than the specific targeting of individual blood vessels. Local controls, discussed later, account for this phenomenon. (Seek additional content for more information on these dynamic aspects of the autonomic nervous system.) Hormones and local chemicals also control blood vessels. Together, these neural and chemical mechanisms reduce or increase blood flow in response to changing body conditions, from exercise to hydration. Regulation of both blood flow and blood pressure is discussed in detail later in this chapter.
The smooth muscle layers of the tunica media are supported by a framework of collagenous fibres that also binds the tunica media to the inner and outer tunics. Along with the collagenous fibres are large numbers of elastic fibres that appear as wavy lines in prepared slides. Separating the tunica media from the outer tunica externa in larger arteries is the external elastic membrane (also called the external elastic lamina), which also appears wavy in slides. This structure is not usually seen in smaller arteries, nor is it seen in veins.
Tunica Externa
The outer tunic, the tunica externa (also called the tunica adventitia), is a substantial sheath of connective tissue composed primarily of collagenous fibres. Some bands of elastic fibres are found here as well. The tunica externa in veins also contains groups of smooth muscle fibres. This is normally the thickest tunic in veins and may be thicker than the tunica media in some larger arteries. The outer layers of the tunica externa are not distinct but rather blend with the surrounding connective tissue outside the vessel, helping to hold the vessel in relative position. If you are able to palpate some of the superficial veins on your upper limbs and try to move them, you will find that the tunica externa prevents this. If the tunica externa did not hold the vessel in place, any movement would likely result in disruption of blood flow.
Arteries
An artery is a blood vessel that conducts blood away from the heart. All arteries have relatively thick walls that can withstand the high pressure of blood ejected from the heart. However, those close to the heart have the thickest walls, containing a high percentage of elastic fibres in all three of their tunics. This type of artery is known as an elastic artery (Figure 6.6.3). Vessels larger than 10 mm in diameter are typically elastic. Their abundant elastic fibres allow them to expand, as blood pumped from the ventricles passes through them, and then to recoil after the surge has passed. If artery walls were rigid and unable to expand and recoil, their resistance to blood flow would greatly increase and blood pressure would rise to even higher levels, which would in turn require the heart to pump harder to increase the volume of blood expelled by each pump (the stroke volume) and maintain adequate pressure and flow. Artery walls would have to become even thicker in response to this increased pressure. The elastic recoil of the vascular wall helps to maintain the pressure gradient that drives the blood through the arterial system. An elastic artery is also known as a conducting artery, because the large diameter of the lumen enables it to accept a large volume of blood from the heart and conduct it to smaller branches.
Figure 6.6.3. Types of arteries and arterioles. a) Comparison of the walls of an elastic artery, a muscular artery, and an arteriole is shown. In terms of scale, the diameter of an arteriole is measured in micrometres compared to millimetres for elastic and muscular arteries; b) hyoid artery, human, 40x, transverse. Image from AnatomyTool used under CC-BY.
Farther from the heart, where the surge of blood has dampened, the percentage of elastic fibres in an artery’s tunica intima decreases and the amount of smooth muscle in its tunica media increases. The artery at this point is described as a muscular artery. The diameter of muscular arteries typically ranges from 0.1 mm to 10 mm. Their thick tunica media allows muscular arteries to play a leading role in vasoconstriction. In contrast, their decreased quantity of elastic fibres limits their ability to expand. Fortunately, because the blood pressure has eased by the time it reaches these more distant vessels, elasticity has become less important.
Notice that although the distinctions between elastic and muscular arteries are important, there is no “line of demarcation” where an elastic artery suddenly becomes muscular. Rather, there is a gradual transition as the vascular tree repeatedly branches. In turn, muscular arteries branch to distribute blood to the vast network of arterioles. For this reason, a muscular artery is also known as a distributing artery.
Arterioles
An arteriole is a very small artery that leads to a capillary. Arterioles have the same three tunics as the larger vessels, but the thickness of each is greatly diminished. The critical endothelial lining of the tunica intima is intact. The tunica media is restricted to one or two smooth muscle cell layers in thickness. The tunica externa remains but is very thin (see Figure 6.6.3).
With a lumen averaging 30 micrometres or less in diameter, arterioles are critical in slowing down—or resisting—blood flow and, thus, causing a substantial drop in blood pressure. Because of this, you may see them referred to as resistance vessels. The muscle fibres in arterioles are normally slightly contracted, causing arterioles to maintain a consistent muscle tone—in this case referred to as vascular tone—in a similar manner to the muscular tone of skeletal muscle. In reality, all blood vessels exhibit vascular tone due to the partial contraction of smooth muscle. The importance of the arterioles is that they will be the primary site of both resistance and regulation of blood pressure. The precise diameter of the lumen of an arteriole at any given moment is determined by neural and chemical controls, and vasoconstriction and vasodilation in the arterioles are the primary mechanisms for distribution of blood flow.
Capillaries
A capillary is a microscopic channel that supplies blood to the tissues themselves, a process called perfusion. Exchange of gases and other substances occurs in the capillaries between the blood and the surrounding cells and their tissue fluid (interstitial fluid). The diameter of a capillary lumen ranges from 5–10 micrometres; the smallest are just barely wide enough for an erythrocyte to squeeze through. Flow through capillaries is often described as microcirculation.
The wall of a capillary consists of the endothelial layer surrounded by a basement membrane with occasional smooth muscle fibres. There is some variation in wall structure: In a large capillary, several endothelial cells bordering each other may line the lumen; in a small capillary, there may be only a single cell layer that wraps around to contact itself.
For capillaries to function, their walls must be leaky, allowing substances to pass through. There are three major types of capillaries, which differ according to their degree of “leakiness:” continuous, fenestrated, and sinusoid capillaries (Figure 6.6.4).
Continuous Capillaries
The most common type of capillary, the continuous capillary, is found in almost all vascularised tissues. Continuous capillaries are characterised by a complete endothelial lining with tight junctions between endothelial cells. Although a tight junction is usually impermeable and only allows for the passage of water and ions, they are often incomplete in capillaries, leaving intercellular clefts that allow for exchange of water and other very small molecules between the blood plasma and the interstitial fluid. Substances that can pass between cells include metabolic products, such as glucose, water, and small hydrophobic molecules like gases and hormones, as well as various leukocytes. Continuous capillaries not associated with the brain are rich in transport vesicles, contributing to either endocytosis or exocytosis. Those in the brain are part of the blood-brain barrier. Here, there are tight junctions and no intercellular clefts, plus a thick basement membrane and astrocyte extensions called end feet; these structures combine to prevent the movement of nearly all substances.
Figure 6.6.4. Types of capillaries. The three major types of capillaries: continuous, fenestrated and sinusoid.
Fenestrated Capillaries
A fenestrated capillary is one that has pores (or fenestrations) in addition to tight junctions in the endothelial lining. These make the capillary permeable to larger molecules. The number of fenestrations and their degree of permeability vary, however, according to their location. Fenestrated capillaries are common in the small intestine, which is the primary site of nutrient absorption, as well as in the kidneys, which filter the blood. They are also found in the choroid plexus of the brain and many endocrine structures, including the hypothalamus, pituitary, pineal and thyroid glands.
Sinusoid Capillaries
A sinusoid capillary (or sinusoid) is the least common type of capillary. Sinusoid capillaries are flattened, and they have extensive intercellular gaps and incomplete basement membranes, in addition to intercellular clefts and fenestrations. This gives them an appearance not unlike Swiss cheese. These very large openings allow for the passage of the largest molecules, including plasma proteins and even cells. Blood flow through sinusoids is very slow, allowing more time for exchange of gases, nutrients, and wastes. Sinusoids are found in the liver and spleen, bone marrow, lymph nodes (where they carry lymph, not blood), and many endocrine glands including the pituitary and adrenal glands. Without these specialised capillaries, these organs would not be able to provide their myriad of functions. For example, when bone marrow forms new blood cells, the cells must enter the blood supply and can only do so through the large openings of a sinusoid capillary; they cannot pass through the small openings of continuous or fenestrated capillaries. The liver also requires extensive specialised sinusoid capillaries in order to process the materials brought to it by the hepatic portal vein from both the digestive tract and spleen, and to release plasma proteins into circulation.
Metarterioles and Capillary Beds
A metarteriole is a type of vessel that has structural characteristics of both an arteriole and a capillary. Slightly larger than the typical capillary, the smooth muscle of the tunica media of the metarteriole is not continuous but forms rings of smooth muscle (sphincters) prior to the entrance to the capillaries. Each metarteriole arises from a terminal arteriole and branches to supply blood to a capillary bed that may consist of 10–100 capillaries.
The precapillary sphincters, circular smooth muscle cells that surround the capillary at its origin with the metarteriole, tightly regulate the flow of blood from a metarteriole to the capillaries it supplies. Their function is critical: If all of the capillary beds in the body were to open simultaneously, they would collectively hold every drop of blood in the body and there would be none in the arteries, arterioles, venules, veins, or the heart itself. Normally, the precapillary sphincters are closed. When the surrounding tissues need oxygen and have excess waste products, the precapillary sphincters open, allowing blood to flow through and exchange to occur before closing once more (Figure 6.6.5). If all of the precapillary sphincters in a capillary bed are closed, blood will flow from the metarteriole directly into a thoroughfare channel and then into the venous circulation, bypassing the capillary bed entirely. This creates what is known as a vascular shunt. In addition, an arteriovenous anastomosis may bypass the capillary bed and lead directly to the venous system.
Although you might expect blood flow through a capillary bed to be smooth, in reality, it moves with an irregular, pulsating flow. This pattern is called vasomotion and is regulated by chemical signals that are triggered in response to changes in internal conditions, such as oxygen, carbon dioxide, hydrogen ion, and lactic acid levels. For example, during strenuous exercise when oxygen levels decrease and carbon dioxide, hydrogen ion, and lactic acid levels all increase, the capillary beds in skeletal muscle are open, as they would be in the digestive system when nutrients are present in the digestive tract. During sleep or rest periods, vessels in both areas are largely closed; they open only occasionally to allow oxygen and nutrient supplies to travel to the tissues to maintain basic life processes.
Figure 6.6.5. Capillary bed. In a capillary bed, arterioles give rise to metarterioles. Precapillary sphincters located at the junction of a metarteriole with a capillary regulate blood flow. A thoroughfare channel connects the metarteriole to a venule. An arteriovenous anastomosis, which directly connects the arteriole with the venule, is shown at the bottom.
Venules
A venule is an extremely small vein, generally 8–100 micrometres in diameter. Postcapillary venules join multiple capillaries exiting from a capillary bed. Multiple venules join to form veins. The walls of venules consist of endothelium, a thin middle layer with a few muscle cells and elastic fibres, plus an outer layer of connective tissue fibres that constitute a very thin tunica externa (Figure 6.6.6). Venules as well as capillaries are the primary sites of emigration or diapedesis, in which the white blood cells adhere to the endothelial lining of the vessels and then squeeze through adjacent cells to enter the tissue fluid.
Veins
A vein is a blood vessel that conducts blood toward the heart. Compared to arteries, veins are thin-walled vessels with large and irregular lumens (see Figure 6.6.6). Because they are low-pressure vessels, larger veins are commonly equipped with valves that promote the unidirectional flow of blood toward the heart and prevent backflow toward the capillaries caused by the inherent low blood pressure in veins as well as the pull of gravity. Table 2 compares the features of arteries and veins.
Figure 6.6.6. Comparison of veins and venules. Many veins have valves to prevent back flow of blood, whereas venules do not. In terms of scale, the diameter of a venule is measured in micrometres compared to millimetres for veins.
Table 6.6.2. Comparison of arteries and veins
Arteries
Veins
Direction of blood flow
Conducts blood away from heart
Conducts blood toward the heart
General appearance
Rounded
Irregular, often collapsed
Pressure
High
Low
Wall thickness
Thick
Thin
Relative oxygen concentration
Higher in systemic arteries
Lower in pulmonary arteries
Lower in systemic veins
Higher in pulmonary veins
Valves
Not present
Present most commonly in limbs and in veins inferior to the heart
Disorders of the Cardiovascular System: Oedema and Varicose Veins
Figure 6.6.7. Varicose veins. Varicose veins are commonly found in the lower limbs. (credit: Thomas Kriese).
Despite the presence of valves and the contributions of other anatomical and physiological adaptations we will cover shortly, over the course of a day, some blood will inevitably pool, especially in the lower limbs, due to the pull of gravity. Any blood that accumulates in a vein will increase the pressure within it, which can then be reflected back into the smaller veins, venules, and eventually even the capillaries. Increased pressure will promote the flow of fluids out of the capillaries and into the interstitial fluid. The presence of excess tissue fluid around the cells leads to a condition called oedema.
Most people experience a daily accumulation of tissue fluid, especially if they spend much of their work life on their feet (like most health professionals). However, clinical oedema goes beyond normal swelling and requires medical treatment. Oedema has many potential causes, including hypertension and heart failure, severe protein deficiency, renal failure, and many others. In order to treat oedema, which is a sign rather than a discrete disorder, the underlying cause must be diagnosed and alleviated.
Oedema may be accompanied by varicose veins, especially in the superficial veins of the legs (Figure 6.6.7). This disorder arises when defective valves allow blood to accumulate within the veins, causing them to distend, twist, and become visible on the surface of the integument. Varicose veins may occur in both sexes but are more common in women and are often related to pregnancy. More than simple cosmetic blemishes, varicose veins are often painful and sometimes itchy or throbbing. Without treatment, they tend to grow worse over time. The use of support hose, as well as elevating the feet and legs whenever possible, may be helpful in alleviating this condition. Laser surgery and interventional radiologic procedures can reduce the size and severity of varicose veins. Severe cases may require conventional surgery to remove the damaged vessels. As there are typically redundant circulation patterns, that is, anastomoses, for the smaller and more superficial veins, removal does not typically impair the circulation. There is evidence that patients with varicose veins suffer a greater risk of developing a thrombus or clot.
Veins as Blood Reservoirs
In addition to their primary function of returning blood to the heart, veins may be considered blood reservoirs, since systemic veins contain approximately 64 percent of the blood volume at any given time (Figure 6.6.8). Their ability to hold this much blood is due to their high capacitance, that is, their capacity to distend (expand) readily to store a high volume of blood, even at a low pressure. The large lumens and relatively thin walls of veins make them far more distensible than arteries; thus, they are said to be capacitance vessels.
Figure 6.6.8. Distribution of Blood Flow.
When blood flow needs to be redistributed to other portions of the body, the vasomotor centre located in the medulla oblongata sends sympathetic stimulation to the smooth muscles in the walls of the veins, causing constriction—or in this case, venoconstriction. Less dramatic than the vasoconstriction seen in smaller arteries and arterioles, venoconstriction may be likened to a “stiffening” of the vessel wall. This increases pressure on the blood within the veins, speeding its return to the heart. As you will note in Figure 6.6.8, approximately 21 percent of the venous blood is located in venous networks within the liver, bone marrow, and integument. This volume of blood is referred to as venous reserve. Through venoconstriction, this “reserve” volume of blood can get back to the heart more quickly for redistribution to other parts of the circulation.
Career Connection
Vascular Surgeons and Technicians
Vascular surgery is a specialty in which the physician deals primarily with diseases of the vascular portion of the cardiovascular system. This includes repair and replacement of diseased or damaged vessels, removal of plaque from vessels, minimally invasive procedures including the insertion of venous catheters, and traditional surgery. Following completion of medical school, the physician generally completes a five-year surgical residency followed by an additional one to two years of vascular specialty training. In Australia, most vascular surgeons are a member of the Vascular Surgeons in Australia and New Zealand (ANZSVS) which is the principal body for the education and training of vascular surgeons in Australia and New Zealand. This is similar to the Society of Vascular Surgery in the United States.
Vascular technicians are specialists in imaging technologies that provide information on the health of the vascular system. They may also assist physicians in treating disorders involving the arteries and veins. This profession often overlaps with cardiovascular technology, which would also include treatments involving the heart. In Australia, a cardiac surgeon may earn between $99k – $509k per annum.
Section Review
Blood pumped by the heart flows through a series of vessels known as arteries, arterioles, capillaries, venules, and veins before returning to the heart. Arteries transport blood away from the heart and branch into smaller vessels, forming arterioles. Arterioles distribute blood to capillary beds, the sites of exchange with the body tissues. Capillaries lead back to small vessels known as venules that flow into the larger veins and eventually back to the heart.
The arterial system is a relatively high-pressure system, so arteries have thick walls that appear round in cross section. The venous system is a lower-pressure system, containing veins that have larger lumens and thinner walls. They often appear flattened. Arteries, arterioles, venules, and veins are composed of three tunics known as the tunica intima, tunica media, and tunica externa. Capillaries have only a tunica intima layer. The tunica intima is a thin layer composed of a simple squamous epithelium known as endothelium and a small amount of connective tissue. The tunica media is a thicker area composed of variable amounts of smooth muscle and connective tissue. It is the thickest layer in all but the largest arteries. The tunica externa is primarily a layer of connective tissue, although in veins, it also contains some smooth muscle. Blood flow through vessels can be dramatically influenced by vasoconstriction and vasodilation in their walls.
Distinguish between systolic pressure, diastolic pressure, pulse pressure, and mean arterial pressure
Describe the clinical measurement of pulse and blood pressure
Identify and discuss five variables affecting arterial blood flow and blood pressure
Discuss several factors affecting blood flow in the venous system
Blood flow refers to the movement of blood through a vessel, tissue, or organ, and is usually expressed in terms of volume of blood per unit of time. It is initiated by the contraction of the ventricles of the heart. Ventricular contraction ejects blood into the major arteries, resulting in flow from regions of higher pressure to regions of lower pressure, as blood encounters smaller arteries and arterioles, then capillaries, then the venules and veins of the venous system. This section discusses a number of critical variables that contribute to blood flow throughout the body. It also discusses the factors that impede or slow blood flow, a phenomenon known as resistance.
As noted earlier, hydrostatic pressure is the force exerted by a fluid due to gravitational pull, usually against the wall of the container in which it is located. One form of hydrostatic pressure is blood pressure, the force exerted by blood upon the walls of the blood vessels or the chambers of the heart. Blood pressure may be measured in capillaries and veins, as well as the vessels of the pulmonary circulation; however, the term blood pressure without any specific descriptors typically refers to systemic arterial blood pressure—that is, the pressure of blood flowing in the arteries of the systemic circulation. In clinical practice, this pressure is measured in mm Hg and is usually obtained using the brachial artery of the arm.
Components of Arterial Blood Pressure
Arterial blood pressure in the larger vessels consists of several distinct components (Figure 6.7.1): systolic and diastolic pressures, pulse pressure, and mean arterial pressure.
Systolic and Diastolic Pressure
When systemic arterial blood pressure is measured, it is recorded as a ratio of two numbers (e.g., 120/80 is a normal adult blood pressure), expressed as systolic pressure over diastolic pressure. The systolic pressure is the higher value (typically around 120 mm Hg) and reflects the arterial pressure resulting from the ejection of blood during ventricular contraction, or systole. The diastolic pressure is the lower value (usually about 80 mm Hg) and represents the arterial pressure of blood during ventricular relaxation, or diastole.
Figure 6.7.1. Systemic blood pressure. The graph shows the components of blood pressure throughout the blood vessels, including systolic, diastolic, mean arterial, and pulse pressures.
Pulse Pressure
As shown in Figure 6.7.1, the difference between the systolic pressure and the diastolic pressure is the pulse pressure. For example, an individual with a systolic pressure of 120 mm Hg and a diastolic pressure of 80 mm Hg would have a pulse pressure of 40 mmHg.
Generally, a pulse pressure should be at least 25 percent of the systolic pressure. A pulse pressure below this level is described as low or narrow. This may occur, for example, in patients with a low stroke volume, which may be seen in congestive heart failure, stenosis of the aortic valve, or significant blood loss following trauma. In contrast, a high or wide pulse pressure is common in healthy people following strenuous exercise, when their resting pulse pressure of 30–40 mm Hg may increase temporarily to 100 mm Hg as stroke volume increases. A persistently high pulse pressure at or above 100 mm Hg may indicate excessive resistance in the arteries and can be caused by a variety of disorders. Chronic high resting pulse pressures can degrade the heart, brain, and kidneys, and warrant medical treatment.
Mean Arterial Pressure
Mean arterial pressure (MAP) represents the “average” pressure of blood in the arteries, that is, the average force driving blood into vessels that serve the tissues. Mean is a statistical concept and is calculated by taking the sum of the values divided by the number of values. Although complicated to measure directly and complicated to calculate, MAP can be approximated by adding the diastolic pressure to one-third of the pulse pressure or systolic pressure minus the diastolic pressure:
MAP = diastolic BP + ((systolic-diastolic BP) / 3)
In Figure 6.7.1, this value is approximately 80 + (120 − 80) / 3, or 93.33. Normally, the MAP falls within the range of 70–110 mm Hg. If the value falls below 60 mm Hg for an extended time, blood pressure will not be high enough to ensure circulation to and through the tissues, which results in ischaemia, or insufficient blood flow. A condition called hypoxia, inadequate oxygenation of tissues, commonly accompanies ischaemia. The term hypoxaemia refers to low levels of oxygen in systemic arterial blood. Neurons are especially sensitive to hypoxia and may die or be damaged if blood flow and oxygen supplies are not quickly restored.
Pulse
After blood is ejected from the heart, elastic fibres in the arteries help maintain a high-pressure gradient as they expand to accommodate the blood, then recoil. This expansion and recoiling effect, known as the pulse, can be palpated manually or measured electronically. Although the effect diminishes over distance from the heart, elements of the systolic and diastolic components of the pulse are still evident down to the level of the arterioles.
Because pulse indicates heart rate, it is measured clinically to provide clues to a patient’s state of health. It is recorded as beats per minute. Both the rate and the strength of the pulse are important clinically. A high or irregular pulse rate can be caused by physical activity or other temporary factors, but it may also indicate a heart condition. The pulse strength indicates the strength of ventricular contraction and cardiac output. If the pulse is strong, then systolic pressure is high. If it is weak, systolic pressure has fallen, and medical intervention may be warranted.
Pulse can be palpated manually by placing the tips of the fingers across an artery that runs close to the body surface and pressing lightly. While this procedure is normally performed using the radial artery in the wrist or the common carotid artery in the neck, any superficial artery that can be palpated may be used (Figure 6.7.2). Common sites to find a pulse include temporal and facial arteries in the head, brachial arteries in the upper arm, femoral arteries in the thigh, popliteal arteries behind the knees, posterior tibial arteries near the medial tarsal regions, and dorsalis pedis arteries in the feet. A variety of commercial electronic devices are also available to measure pulse.
Figure 6.7.2. Pulse sites. The pulse is most readily measured at the radial artery but can be measured at any of the pulse points shown.
Measurement of Blood Pressure
Blood pressure is one of the critical parameters measured on virtually every patient in every healthcare setting. The technique used today was developed more than 100 years ago by a pioneering Russian physician, Dr. Nikolai Korotkoff. Turbulent blood flow through the vessels can be heard as a soft ticking while measuring blood pressure; these sounds are known as Korotkoff sounds. The technique of measuring blood pressure requires the use of a sphygmomanometer (a blood pressure cuff attached to a measuring device) and a stethoscope. The technique is as follows:
The clinician wraps an inflatable cuff tightly around the patient’s arm at about the level of the heart.
The clinician squeezes a rubber pump to inject air into the cuff, raising pressure around the artery and temporarily cutting off blood flow into the patient’s arm.
The clinician places the stethoscope on the patient’s antecubital region and, while gradually allowing air within the cuff to escape, listens for the Korotkoff sounds.
Although there are five recognised Korotkoff sounds, only two are normally recorded. Initially, no sounds are heard since there is no blood flow through the vessels, but as air pressure drops, the cuff relaxes, and blood flow returns to the arm. As shown in Figure 6.7.3, the first sound heard through the stethoscope—the first Korotkoff sound—indicates systolic pressure. As more air is released from the cuff, blood is able to flow freely through the brachial artery and all sounds disappear. The point at which the last sound is heard is recorded as the patient’s diastolic pressure.
Figure 6.7.3. Blood pressure measurement. When pressure in a sphygmomanometer cuff is released, a clinician can hear the Korotkoff sounds. In this graph, a blood pressure tracing is aligned to a measurement of systolic and diastolic pressures.
The majority of hospitals and clinics have automated equipment for measuring blood pressure that work on the same principles. An even more recent innovation is a small instrument that wraps around a patient’s wrist. The patient then holds the wrist over the heart while the device measures blood flow and records pressure.
Variables Affecting Blood Flow and Blood Pressure
Five variables influence blood flow and blood pressure:
Cardiac output
Compliance
Volume of the blood
Viscosity of the blood
Blood vessel length and diameter
Recall that blood moves from higher pressure to lower pressure. It is pumped from the heart into the arteries at high pressure. If you increase pressure in the arteries (afterload), and cardiac function does not compensate, blood flow will actually decrease. In the venous system, the opposite relationship is true. Increased pressure in the veins does not decrease flow as it does in arteries, but actually increases flow. Since pressure in the veins is normally relatively low, for blood to flow back into the heart, the pressure in the atria during atrial diastole must be even lower. It normally approaches zero, except when the atria contract (see Figure 6.7.1).
Cardiac Output
Cardiac output is the measurement of blood flow from the heart through the ventricles and is usually measured in litres per minute. Any factor that causes cardiac output to increase, by elevating heart rate or stroke volume or both, will elevate blood pressure and promote blood flow. These factors include sympathetic stimulation, the catecholamines adrenaline and noradrenaline, thyroid hormones, and increased calcium ion levels. Conversely, any factor that decreases cardiac output, by decreasing heart rate or stroke volume or both, will decrease arterial pressure and blood flow. These factors include parasympathetic stimulation, elevated or decreased potassium ion levels, decreased calcium levels, anoxia, and acidosis.
Compliance
Compliance is the ability of any compartment to expand to accommodate increased content. A metal pipe, for example, is not compliant, whereas a balloon is. The greater the compliance of an artery, the more effectively it is able to expand to accommodate surges in blood flow without increased resistance or blood pressure. Veins are more compliant than arteries and can expand to hold more blood. When vascular disease causes stiffening of arteries, compliance is reduced and resistance to blood flow is increased. The result is more turbulence, higher pressure within the vessel, and reduced blood flow. This increases the work of the heart.
A Mathematical Approach to Factors Affecting Blood Flow
Jean Louis Marie Poiseuille was a French physician and physiologist who devised a mathematical equation describing blood flow and its relationship to known parameters. The same equation also applies to engineering studies of the flow of fluids. Although understanding the math behind the relationships among the factors affecting blood flow is not necessary to understand blood flow, it can help solidify an understanding of their relationships. Please note that even if the equation looks intimidating, breaking it down into its components and following the relationships will make these relationships clearer, even if you are weak in math. Focus on the three critical variables: radius (r), vessel length (λ), and viscosity (η).
Poiseuille’s equation:
Blood flow = π ΔP r48ηλ
π is the Greek letter pi, used to represent the mathematical constant that is the ratio of a circle’s circumference to its diameter. It may commonly be represented as 3.14, although the actual number extends to infinity.
ΔP represents the difference in pressure.
r4 is the radius (one-half of the diameter) of the vessel to the fourth power.
η is the Greek letter eta and represents the viscosity of the blood.
λ is the Greek letter lambda and represents the length of a blood vessel.
One of several things this equation allows us to do is calculate the resistance in the vascular system. Normally this value is extremely difficult to measure, but it can be calculated from this known relationship:
Blood flow = ΔP/Resistance
If we rearrange this slightly,
Resistance = ΔP/Blood flow
Then by substituting Poiseuille’s equation for blood flow:
Resistance =8ηλ/πr4
By examining this equation, you can see that there are only three variables: viscosity, vessel length, and radius, since 8 and π are both constants. The important thing to remember is this: Two of these variables, viscosity and vessel length, will change slowly in the body. Only one of these factors, the radius, can be changed rapidly by vasoconstriction and vasodilation, thus dramatically impacting resistance and flow. Further, small changes in the radius will greatly affect flow, since it is raised to the fourth power in the equation.
We have briefly considered how cardiac output and blood volume impact blood flow and pressure; the next step is to see how the other variables (contraction, vessel length, and viscosity) articulate with Poiseuille’s equation and what they can teach us about the impact on blood flow.
Blood Volume
The relationship between blood volume, blood pressure, and blood flow is intuitively obvious. Water may merely trickle along a creek bed in a dry season but rush quickly and under great pressure after a heavy rain. Similarly, as blood volume decreases, pressure and flow decrease. As blood volume increases, pressure and flow increase.
Under normal circumstances, blood volume varies little. Low blood volume, called hypovolaemia, may be caused by bleeding, dehydration, vomiting, severe burns, or some medications used to treat hypertension. It is important to recognise that other regulatory mechanisms in the body are so effective at maintaining blood pressure that an individual may be asymptomatic until 10–20 percent of the blood volume has been lost. Treatment typically includes intravenous fluid replacement.
Hypervolaemia, excessive fluid volume, may be caused by retention of water and sodium, as seen in patients with heart failure, liver cirrhosis, some forms of kidney disease, hyperaldosteronism, and some glucocorticoid steroid treatments. Restoring homeostasis in these patients depends upon reversing the condition that triggered the hypervolaemia.
Blood Viscosity
Viscosity is the thickness of fluids that affects their ability to flow. Clean water, for example, is less viscous than mud. The viscosity of blood is directly proportional to resistance and inversely proportional to flow; therefore, any condition that causes viscosity to increase will also increase resistance and decrease flow. For example, imagine sipping milk, then a milkshake, through the same size straw. You experience more resistance and therefore less flow from the milkshake. Conversely, any condition that causes viscosity to decrease (such as when the milkshake melts) will decrease resistance and increase flow.
Normally the viscosity of blood does not change over short periods of time. The two primary determinants of blood viscosity are the formed elements and plasma proteins. Since most formed elements are erythrocytes, any condition affecting erythropoiesis, such as polycythaemia or anaemia, can alter viscosity. Since most plasma proteins are produced by the liver, any condition affecting liver function can also change the viscosity slightly and therefore decrease blood flow. Liver abnormalities include hepatitis, cirrhosis, alcohol damage, and drug toxicities. While leukocytes and platelets are normally a small component of the formed elements, there are some rare conditions in which severe overproduction can impact viscosity as well.
Vessel Length and Diameter
The length of a vessel is directly proportional to its resistance: the longer the vessel, the greater the resistance and the lower the flow. As with blood volume, this makes intuitive sense, since the increased surface area of the vessel will impede the flow of blood. Likewise, if the vessel is shortened, the resistance will decrease and flow will increase.
The length of our blood vessels increases throughout childhood as we grow, of course, but is unchanging in adults under normal physiological circumstances. Further, the distribution of vessels is not the same in all tissues. Adipose tissue does not have an extensive vascular supply. One pound of adipose tissue contains approximately 200 miles of vessels, whereas skeletal muscle contains more than twice that. Overall, vessels decrease in length only during loss of mass or amputation. An individual weighing 68kg has approximately 96,560km of vessels in the body. Gaining about 4.5km adds from 3218 to 6437 kilometres of vessels, depending upon the nature of the gained tissue. One of the great benefits of weight reduction is the reduced stress to the heart, which does not have to overcome the resistance of as many miles of vessels.
In contrast to length, the diameter of blood vessels changes throughout the body, according to the type of vessel, as we discussed earlier. The diameter of any given vessel may also change frequently throughout the day in response to neural and chemical signals that trigger vasodilation and vasoconstriction. The vascular tone of the vessel is the contractile state of the smooth muscle and the primary determinant of diameter, and thus of resistance and flow. The effect of vessel diameter on resistance is inverse: Given the same volume of blood, an increased diameter means there is less blood contacting the vessel wall, thus lower friction and lower resistance, subsequently increasing flow. A decreased diameter means more of the blood contacts the vessel wall, and resistance increases, subsequently decreasing flow.
The influence of lumen diameter on resistance is dramatic: A slight increase or decrease in diameter causes a huge decrease or increase in resistance. This is because resistance is inversely proportional to the radius of the blood vessel (one-half of the vessel’s diameter) raised to the fourth power (R = 1/r4). This means, for example, that if an artery or arteriole constricts to one-half of its original radius, the resistance to flow will increase 16 times. And if an artery or arteriole dilates to twice its initial radius, then resistance in the vessel will decrease to 1/16 of its original value and flow will increase 16 times.
The Roles of Vessel Diameter and Total Area in Blood Flow and Blood Pressure
Recall that we classified arterioles as resistance vessels, because given their small lumen, they dramatically slow the flow of blood from arteries. In fact, arterioles are the site of greatest resistance in the entire vascular network. This may seem surprising, given that capillaries have a smaller size. How can this phenomenon be explained?
Figure 6.7.4 compares vessel diameter, total cross-sectional area, average blood pressure, and blood velocity through the systemic vessels. Notice in parts (a) and (b) that the total cross-sectional area of the body’s capillary beds is far greater than any other type of vessel. Although the diameter of an individual capillary is significantly smaller than the diameter of an arteriole, there are vastly more capillaries in the body than there are other types of blood vessels. Part (c) shows that blood pressure drops unevenly as blood travels from arteries to arterioles, capillaries, venules, and veins, and encounters greater resistance. However, the site of the most precipitous drop, and the site of greatest resistance, is the arterioles. This explains why vasodilation and vasoconstriction of arterioles play more significant roles in regulating blood pressure than do the vasodilation and vasoconstriction of other vessels.
Part (d) shows that the velocity (speed) of blood flow decreases dramatically as the blood moves from arteries to arterioles to capillaries. This slow flow rate allows more time for exchange processes to occur. As blood flows through the veins, the rate of velocity increases, as blood is returned to the heart.
Figure 6.7.4. Relationships among vessels in the systemic circuit. The relationships among blood vessels that can be compared include (a) vessel diameter, (b) total cross-sectional area, (c) average blood pressure, and (d) velocity of blood flow.
Disorders of the Cardiovascular System: Arteriosclerosis
Compliance allows an artery to expand when blood is pumped through it from the heart, and then to recoil after the surge has passed. This helps promote blood flow. In arteriosclerosis, compliance is reduced, and pressure and resistance within the vessel increase. This is a leading cause of hypertension and coronary heart disease, as it causes the heart to work harder to generate a pressure great enough to overcome the resistance.
Arteriosclerosis begins with injury to the endothelium of an artery, which may be caused by irritation from high blood glucose, infection, tobacco use, excessive blood lipids, and other factors. Artery walls that are constantly stressed by blood flowing at high pressure are also more likely to be injured—which means that hypertension can promote arteriosclerosis, as well as result from it.
Recall that tissue injury causes inflammation. As inflammation spreads into the artery wall, it weakens and scars it, leaving it stiff (sclerotic). As a result, compliance is reduced. Moreover, circulating triglycerides and cholesterol can seep between the damaged lining cells and become trapped within the artery wall, where they are frequently joined by leukocytes, calcium, and cellular debris. Eventually, this build-up, called plaque, can narrow arteries enough to impair blood flow. The term for this condition, atherosclerosis (athero- = “porridge”) describes the mealy deposits (Figure 6.7.5).
Sometimes a plaque can rupture, causing microscopic tears in the artery wall that allow blood to leak into the tissue on the other side. When this happens, platelets rush to the site to clot the blood. This clot can further obstruct the artery and—if it occurs in a coronary or cerebral artery—cause a sudden heart attack or stroke. Alternatively, plaque can break off and travel through the bloodstream as an embolus until it blocks a more distant, smaller artery.
Even without total blockage, vessel narrowing leads to ischaemia—reduced blood flow—to the tissue region “downstream” of the narrowed vessel. Ischaemia in turn leads to hypoxia—decreased supply of oxygen to the tissues. Hypoxia involving cardiac muscle or brain tissue can lead to cell death and severe impairment of brain or heart function.
A major risk factor for both arteriosclerosis and atherosclerosis is advanced age, as the conditions tend to progress over time. Arteriosclerosis is normally defined as the more generalised loss of compliance, “hardening of the arteries,” whereas atherosclerosis is a more specific term for the build-up of plaque in the walls of the vessel and is a specific type of arteriosclerosis. There is also a distinct genetic component, and pre-existing hypertension and/or diabetes also greatly increase the risk. However, obesity, poor nutrition, lack of physical activity, and tobacco use all are major risk factors.
Treatment includes lifestyle changes, such as weight loss, smoking cessation, regular exercise, and adoption of a diet low in sodium and saturated fats. Medications to reduce cholesterol and blood pressure may be prescribed. For blocked coronary arteries, surgery is warranted. In angioplasty, a catheter is inserted into the vessel at the point of narrowing, and a second catheter with a balloon-like tip is inflated to widen the opening. To prevent subsequent collapse of the vessel, a small mesh tube called a stent is often inserted. In an endarterectomy, plaque is surgically removed from the walls of a vessel. This operation is typically performed on the carotid arteries of the neck, which are a prime source of oxygenated blood for the brain. In a coronary bypass procedure, a non-vital superficial vessel from another part of the body (often the great saphenous vein) or a synthetic vessel is inserted to create a path around the blocked area of a coronary artery.
Venous System
The pumping action of the heart propels the blood into the arteries, from an area of higher pressure toward an area of lower pressure. If blood is to flow from the veins back into the heart, the pressure in the veins must be greater than the pressure in the atria of the heart. Two factors help maintain this pressure gradient between the veins and the heart. First, the pressure in the atria during diastole is very low, often approaching zero when the atria are relaxed (atrial diastole). Second, two physiologic “pumps” increase pressure in the venous system. The use of the term “pump” implies a physical device that speeds flow. These physiological pumps are less obvious.
Skeletal Muscle Pump
In many body regions, the pressure within the veins can be increased by the contraction of the surrounding skeletal muscle. This mechanism, known as the skeletal muscle pump (Figure 6.7.6), helps the lower-pressure veins counteract the force of gravity, increasing pressure to move blood back to the heart. As leg muscles contract, for example during walking or running, they exert pressure on nearby veins with their numerous one-way valves. This increased pressure causes blood to flow upward, opening valves superior to the contracting muscles so blood flows through. Simultaneously, valves inferior to the contracting muscles close; thus, blood should not seep back downward toward the feet. Military recruits are trained to flex their legs slightly while standing at attention for prolonged periods. Failure to do so may allow blood to pool in the lower limbs rather than returning to the heart. Consequently, the brain will not receive enough oxygenated blood, and the individual may lose consciousness.
Figure 6.7.6. Skeletal muscle pump. The contraction of skeletal muscles surrounding a vein compresses the blood and increases the pressure in that area. This action forces blood closer to the heart where venous pressure is lower. Note the importance of the one-way valves to assure that blood flows only in the proper direction.
Respiratory Pump
The respiratory pump aids blood flow through the veins of the thorax and abdomen. During inhalation, the volume of the thorax increases, largely through the contraction of the diaphragm, which moves downward and compresses the abdominal cavity. The elevation of the chest caused by the contraction of the external intercostal muscles also contributes to the increased volume of the thorax. The volume increase causes air pressure within the thorax to decrease, allowing us to inhale. Additionally, as air pressure within the thorax drops, blood pressure in the thoracic veins also decreases, falling below the pressure in the abdominal veins. This causes blood to flow along its pressure gradient from veins outside the thorax, where pressure is higher, into the thoracic region, where pressure is now lower. This in turn promotes the return of blood from the thoracic veins to the atria. During exhalation, when air pressure increases within the thoracic cavity, pressure in the thoracic veins increases, speeding blood flow into the heart while valves in the veins prevent blood from flowing backward from the thoracic and abdominal veins.
Pressure Relationship in the Venous System
Although vessel diameter increases from the smaller venules to the larger veins and eventually to the venae cavae (singular = vena cava), the total cross-sectional area actually decreases (see Figure 6.7.4a and 6.7.4b). The individual veins are larger in diameter than the venules, but their total number is much lower, so their total cross-sectional area is also lower.
Also notice that, as blood moves from venules to veins, the average blood pressure drops (see Figure 6.7.4c), but the blood velocity actually increases (see Figure 6.7.4d). This pressure gradient drives blood back toward the heart. Again, the presence of one-way valves and the skeletal muscle and respiratory pumps contribute to this increased flow. Since approximately 64 percent of the total blood volume resides in systemic veins, any action that increases the flow of blood through the veins will increase venous return to the heart. Maintaining vascular tone within the veins prevents the veins from merely distending, dampening the flow of blood, and as you will see, vasoconstriction actually enhances the flow.
The Role of Venoconstriction in Resistance, Blood Pressure and Flow
As previously discussed, vasoconstriction of an artery or arteriole decreases the radius, increasing resistance and pressure, but decreasing flow. Venoconstriction, on the other hand, has a very different outcome. The walls of veins are thin but irregular; thus, when the smooth muscle in those walls constricts, the lumen becomes more rounded. The more rounded the lumen, the less surface area the blood encounters, and the less resistance the vessel offers. Vasoconstriction increases pressure within a vein as it does in an artery, but in veins, the increased pressure increases flow. Recall that the pressure in the atria, into which the venous blood will flow, is very low, approaching zero for at least part of the relaxation phase of the cardiac cycle. Thus, venoconstriction increases the return of blood to the heart. Another way of stating this is that venoconstriction increases the preload or stretch of the cardiac muscle and increases contraction.
Section Review
Blood flow is the movement of blood through a vessel, tissue, or organ. The slowing or blocking of blood flow is called resistance. Blood pressure is the force that blood exerts upon the walls of the blood vessels or chambers of the heart. The components of blood pressure include systolic pressure, which results from ventricular contraction, and diastolic pressure, which results from ventricular relaxation. Pulse pressure is the difference between systolic and diastolic measures, and mean arterial pressure is the “average” pressure of blood in the arterial system, driving blood into the tissues. Pulse, the expansion and recoiling of an artery, reflects the heartbeat. The variables affecting blood flow and blood pressure in the systemic circulation are cardiac output, compliance, blood volume, blood viscosity, and the length and diameter of the blood vessels. In the arterial system, vasodilation and vasoconstriction of the arterioles is a significant factor in systemic blood pressure: Slight vasodilation greatly decreases resistance and increases flow, whereas slight vasoconstriction greatly increases resistance and decreases flow. In the arterial system, as resistance increases, blood pressure increases and flow decreases. In the venous system, constriction increases blood pressure as it does in arteries; the increasing pressure helps to return blood to the heart. In addition, constriction causes the vessel lumen to become more rounded, decreasing resistance and increasing blood flow. Venoconstriction, while less important than arterial vasoconstriction, works with the skeletal muscle pump, the respiratory pump, and their valves to promote venous return to the heart.
Identify the primary mechanisms of capillary exchange
Distinguish between capillary hydrostatic pressure and blood colloid osmotic pressure, explaining the contribution of each to net filtration pressure
Compare filtration and reabsorption
Explain the fate of fluid that is not reabsorbed from the tissues into the vascular capillaries
The primary purpose of the cardiovascular system is to circulate gases, nutrients, wastes, and other substances to and from the cells of the body. Small molecules, such as gases, lipids, and lipid-soluble molecules, can diffuse directly through the membranes of the endothelial cells of the capillary wall. Glucose, amino acids, and ions—including sodium, potassium, calcium, and chloride—use transporters to move through specific channels in the membrane by facilitated diffusion. Glucose, ions, and larger molecules may also leave the blood through intercellular clefts. Larger molecules can pass through the pores of fenestrated capillaries, and even large plasma proteins can pass through the great gaps in the sinusoids. Some large proteins in blood plasma can move into and out of the endothelial cells packaged within vesicles by endocytosis and exocytosis. Water moves by osmosis.
Bulk Flow
The mass movement of fluids into and out of capillary beds requires a transport mechanism far more efficient than mere diffusion. This movement, often referred to as bulk flow, involves two pressure-driven mechanisms: Volumes of fluid move from an area of higher pressure in a capillary bed to an area of lower pressure in the tissues via filtration. In contrast, the movement of fluid from an area of higher pressure in the tissues into an area of lower pressure in the capillaries is reabsorption. Two types of pressure interact to drive each of these movements: hydrostatic pressure and osmotic pressure.
Hydrostatic Pressure
The primary force driving fluid transport between the capillaries and tissues is hydrostatic pressure, which can be defined as the pressure of any fluid enclosed in a space. Blood hydrostatic pressure is the force exerted by the blood confined within blood vessels or heart chambers. Even more specifically, the pressure exerted by blood against the wall of a capillary is called capillary hydrostatic pressure (CHP), and is the same as capillary blood pressure. CHP is the force that drives fluid out of capillaries and into the tissues.
As fluid exits a capillary and moves into tissues, the hydrostatic pressure in the interstitial fluid correspondingly rises. This opposing hydrostatic pressure is called the interstitial fluid hydrostatic pressure (IFHP). Generally, the CHP originating from the arterial pathways is considerably higher than the IFHP, because lymphatic vessels are continually absorbing excess fluid from the tissues. Thus, fluid generally moves out of the capillary and into the interstitial fluid. This process is called filtration.
Osmotic Pressure
The net pressure that drives reabsorption—the movement of fluid from the interstitial fluid back into the capillaries—is called osmotic pressure (sometimes referred to as oncotic pressure). Whereas hydrostatic pressure forces fluid out of the capillary, osmotic pressure draws fluid back in. Osmotic pressure is determined by osmotic concentration gradients, that is, the difference in the solute-to-water concentrations in the blood and tissue fluid. A region higher in solute concentration (and lower in water concentration) draws water across a semipermeable membrane from a region higher in water concentration (and lower in solute concentration).
As we discuss osmotic pressure in blood and tissue fluid, it is important to recognise that the formed elements of blood do not contribute to osmotic concentration gradients. Rather, it is the plasma proteins that play the key role. Solutes also move across the capillary wall according to their concentration gradient, but overall, the concentrations should be similar and not have a significant impact on osmosis. Because of their large size and chemical structure, plasma proteins are not truly solutes, that is, they do not dissolve but are dispersed or suspended in their fluid medium, forming a colloid rather than a solution.
The pressure created by the concentration of colloidal proteins in the blood is called the blood colloidal osmotic pressure (BCOP). Its effect on capillary exchange accounts for the reabsorption of water. The plasma proteins suspended in blood cannot move across the semipermeable capillary cell membrane, and so they remain in the plasma. As a result, blood has a higher colloidal concentration and lower water concentration than tissue fluid. It therefore attracts water. We can also say that the BCOP is higher than the interstitial fluid colloidal osmotic pressure (IFCOP), which is always very low because interstitial fluid contains few proteins. Thus, water is drawn from the tissue fluid back into the capillary, carrying dissolved molecules with it. This difference in colloidal osmotic pressure accounts for reabsorption.
Interaction of Hydrostatic and Osmotic Pressures
The normal unit used to express pressures within the cardiovascular system is millimetres of mercury (mm Hg). When blood leaving an arteriole first enters a capillary bed, the CHP is quite high—about 35 mm Hg. Gradually, this initial CHP declines as the blood moves through the capillary so that by the time the blood has reached the venous end, the CHP has dropped to approximately 18 mm Hg. In comparison, the plasma proteins remain suspended in the blood, so the BCOP remains fairly constant at about 25 mm Hg throughout the length of the capillary and considerably below the osmotic pressure in the interstitial fluid.
The net filtration pressure (NFP) represents the interaction of the hydrostatic and osmotic pressures, driving fluid out of the capillary. It is equal to the difference between the CHP and the BCOP. Since filtration is, by definition, the movement of fluid out of the capillary, when reabsorption is occurring, the NFP is a negative number.
NFP changes at different points in a capillary bed (Figure 6.8.1). Close to the arterial end of the capillary, it is approximately 10 mm Hg, because the CHP of 35 mm Hg minus the BCOP of 25 mm Hg equals 10 mm Hg. Recall that the hydrostatic and osmotic pressures of the interstitial fluid are essentially negligible. Thus, the NFP of 10 mm Hg drives a net movement of fluid out of the capillary at the arterial end. At approximately the middle of the capillary, the CHP is about the same as the BCOP of 25 mm Hg, so the NFP drops to zero. At this point, there is no net change of volume: Fluid moves out of the capillary at the same rate as it moves into the capillary. Near the venous end of the capillary, the CHP has dwindled to about 18 mm Hg due to loss of fluid. Because the BCOP remains steady at 25 mm Hg, water is drawn into the capillary, that is, reabsorption occurs. Another way of expressing this is to say that at the venous end of the capillary, there is an NFP of −7 mm Hg.
Figure 6.8.1. Capillary exchange. Net filtration occurs near the arterial end of the capillary since capillary hydrostatic pressure (CHP) is greater than blood colloidal osmotic pressure (BCOP). There is no net movement of fluid near the midpoint since CHP = BCOP. Net reabsorption occurs near the venous end since BCOP is greater than CHP.
The Role of Lymphatic Capillaries
Since overall CHP is higher than BCOP, it is inevitable that more net fluid will exit the capillary through filtration at the arterial end than enters through reabsorption at the venous end. Considering all capillaries over the course of a day, this can be quite a substantial amount of fluid: Approximately 24 litres per day are filtered, whereas 20.4 litres are reabsorbed. This excess fluid is picked up by capillaries of the lymphatic system. These extremely thin-walled vessels have copious numbers of valves that ensure unidirectional flow through ever-larger lymphatic vessels that eventually drain into the subclavian veins in the neck. An important function of the lymphatic system is to return the fluid (lymph) to the blood. Lymph may be thought of as recycled blood plasma.
Section Review
Small molecules can cross into and out of capillaries via simple or facilitated diffusion. Some large molecules can cross in vesicles or through clefts, fenestrations, or gaps between cells in capillary walls. However, the bulk flow of capillary and tissue fluid occurs via filtration and reabsorption. Filtration, the movement of fluid out of the capillaries, is driven by the CHP. Reabsorption, the influx of tissue fluid into the capillaries, is driven by the BCOP. Filtration predominates in the arterial end of the capillary; in the middle section, the opposing pressures are virtually identical so there is no net exchange, whereas reabsorption predominates at the venule end of the capillary. The hydrostatic and colloid osmotic pressures in the interstitial fluid are negligible in healthy circumstances.
Discuss the mechanisms involved in the neural regulation of vascular homeostasis
Describe the contribution of a variety of hormones to the renal regulation of blood pressure
Identify the effects of exercise on vascular homeostasis
Discuss how hypertension, haemorrhage, and circulatory shock affect vascular health
In order to maintain homeostasis in the cardiovascular system and provide adequate blood to the tissues, blood flow must be redirected continually to the tissues as they become more active. In a very real sense, the cardiovascular system engages in resource allocation, because there is not enough blood flow to distribute blood equally to all tissues simultaneously. For example, when an individual is exercising, more blood will be directed to skeletal muscles, the heart, and the lungs. Following a meal, more blood is directed to the digestive system. Only the brain receives a more or less constant supply of blood whether you are active, resting, thinking, or engaged in any other activity.
Table 6.9.1 provides the distribution of systemic blood at rest and during exercise. Although most of the data appears logical, the values for the distribution of blood to the integument may seem surprising. During exercise, the body distributes more blood to the body surface where it can dissipate the excess heat generated by increased activity into the environment.
Table 6.9.1 Systemic blood flow during rest, mild exercise, and maximal exercise in a healthy young individual.
Organ
Resting (mL/min)
Mild exercise (mL/min)
Maximal exercise (mL/min)
Skeletal muscle
1200
4500
12,500
Heart
250
350
750
Brain
750
750
750
Integument
500
1500
1900
Kidney
1100
900
600
Gastrointestinal
1400
1100
600
Others
(i.e., liver, spleen)
600
400
400
Total
5800
9500
17,500
Three homeostatic mechanisms ensure adequate blood flow, blood pressure, distribution, and ultimately perfusion: neural, endocrine, and autoregulatory mechanisms. They are summarised in Figure 6.9.1.
Figure 6.9.1. Summary of factors maintaining vascular homeostasis. Adequate blood flow, blood pressure, distribution, and perfusion involve autoregulatory, neural, and endocrine mechanisms.
Neural Regulation
The nervous system plays a critical role in the regulation of vascular homeostasis. The primary regulatory sites include the cardiovascular centres in the brain that control both cardiac and vascular functions. In addition, more generalised neural responses from the limbic system and the autonomic nervous system are factors.
The Cardiovascular Centres in the Brain
Neurological regulation of blood pressure and flow depends on the cardiovascular centres located in the medulla oblongata. This cluster of neurons responds to changes in blood pressure as well as blood concentrations of oxygen, carbon dioxide, and hydrogen ions. The cardiovascular centre contains three distinct paired components:
The cardioaccelerator centres stimulate cardiac function by regulating heart rate and stroke volume via sympathetic stimulation from the cardiac accelerator nerve.
The cardioinhibitor centre slow cardiac function by decreasing heart rate and stroke volume via parasympathetic stimulation from the vagus nerve.
The vasomotor centre control vessel tone or contraction of the smooth muscle in the tunica media. Changes in diameter affect peripheral resistance, pressure, and flow, which affect cardiac output. The majority of these neurons act via the release of the neurotransmitter noradrenaline from sympathetic neurons.
Although each centre functions independently, they are not anatomically distinct.
There is also a small population of neurons that control vasodilation in the vessels of the brain and skeletal muscles by relaxing the smooth muscle fibres in the vessel tunics. Many of these are cholinergic neurons, that is, they release acetylcholine, which in turn stimulates the vessels’ endothelial cells to release nitric oxide (NO), which causes vasodilation. Others release noradrenaline that binds to β2 receptors. A few neurons release NO directly as a neurotransmitter.
Recall that mild stimulation of the skeletal muscles maintains muscle tone. A similar phenomenon occurs with vascular tone in vessels. As noted earlier, arterioles are normally partially constricted: With maximal stimulation, their radius may be reduced to one-half of the resting state. Full dilation of most arterioles requires that this sympathetic stimulation be suppressed. When it is, an arteriole can expand by as much as 150 percent. Such a significant increase can dramatically affect resistance, pressure, and flow.
Baroreceptor Reflexes
Baroreceptors are specialised stretch receptors located within thin areas of blood vessels and heart chambers that respond to the degree of stretch caused by the presence of blood. They send impulses to the cardiovascular centre to regulate blood pressure. Vascular baroreceptors are found primarily in sinuses (small cavities) within the aorta and carotid arteries: The aortic sinuses are found in the walls of the ascending aorta just superior to the aortic valve, whereas the carotid sinuses are in the base of the internal carotid arteries. There are also low-pressure baroreceptors located in the walls of the venae cavae and right atrium.
When blood pressure increases, the baroreceptors are stretched more tightly and initiate action potentials at a higher rate. At lower blood pressures, the degree of stretch is lower and the rate of firing is slower. When the cardiovascular centre in the medulla oblongata receives this input, it triggers a reflex that maintains homeostasis (Figure 6.9.2).
When blood pressure rises too high, the baroreceptors fire at a higher rate and trigger parasympathetic stimulation of the heart. As a result, cardiac output falls. Sympathetic stimulation of the peripheral arterioles will also decrease, resulting in vasodilation. Combined, these activities cause blood pressure to fall.
When blood pressure drops too low, the rate of baroreceptor firing decreases. This will trigger an increase in sympathetic stimulation of the heart, causing cardiac output to increase. It will also trigger sympathetic stimulation of the peripheral vessels, resulting in vasoconstriction. Combined, these activities cause blood pressure to rise.
Figure 6.9.2. Baroreceptor reflexes for maintaining vascular homeostasis. Increased blood pressure results in increased rates of baroreceptor firing, whereas decreased blood pressure results in slower rates of fire, both initiating the homeostatic mechanism to restore blood pressure.
The baroreceptors in the venae cavae and right atrium monitor blood pressure as the blood returns to the heart from the systemic circulation. Normally, blood flow into the aorta is the same as blood flow back into the right atrium. If blood is returning to the right atrium more rapidly than it is being ejected from the left ventricle, the atrial receptors will stimulate the cardiovascular centres to increase sympathetic firing and increase cardiac output until homeostasis is achieved. The opposite is also true. This mechanism is referred to as the atrial reflex.
Chemoreceptor Reflexes
In addition to the baroreceptors are chemoreceptors that monitor levels of oxygen, carbon dioxide, and hydrogen ions (pH), and thereby contribute to vascular homeostasis. Chemoreceptors monitoring the blood are located in close proximity to the baroreceptors in the aortic and carotid sinuses. They signal the cardiovascular centre as well as the respiratory centres in the medulla oblongata.
Since tissues consume oxygen and produce carbon dioxide and acids as waste products, when the body is more active, oxygen levels fall and carbon dioxide levels rise as cells undergo cellular respiration to meet the energy needs of activities. This causes more hydrogen ions to be produced, causing the blood pH to drop. When the body is resting, oxygen levels are higher, carbon dioxide levels are lower, more hydrogen is bound, and pH rises. (Seek additional content for more detail about pH.)
The chemoreceptors respond to increasing carbon dioxide and hydrogen ion levels (falling pH) by stimulating the cardioaccelerator and vasomotor centres, increasing cardiac output and constricting peripheral vessels. The cardioinhibitor centres are suppressed. With falling carbon dioxide and hydrogen ion levels (increasing pH), the cardioinhibitor centres are stimulated, and the cardioaccelerator and vasomotor centres are suppressed, decreasing cardiac output and causing peripheral vasodilation. In order to maintain adequate supplies of oxygen to the cells and remove waste products such as carbon dioxide, it is essential that the respiratory system respond to changing metabolic demands. In turn, the cardiovascular system will transport these gases to the lungs for exchange, again in accordance with metabolic demands. This interrelationship of cardiovascular and respiratory control cannot be overemphasised.
Other neural mechanisms can also have a significant impact on cardiovascular function. These include the limbic system that links physiological responses to psychological stimuli, as well as generalised sympathetic and parasympathetic stimulation.
Endocrine Regulation
Endocrine control over the cardiovascular system involves the catecholamines, adrenaline and noradrenaline, as well as several hormones that interact with the kidneys in the regulation of blood volume.
Adrenaline and Noradrenaline
The catecholamines adrenaline and noradrenaline are released by the adrenal medulla, and enhance and extend the body’s sympathetic or “fight-or-flight” response (see Figure 6.9.3). They increase heart rate and force of contraction, while temporarily constricting blood vessels to organs not essential for flight-or-fight responses and redirecting blood flow to the liver, muscles, and heart.
Antidiuretic Hormone (ADH)
Antidiuretic hormone (ADH), also known as vasopressin, is secreted by the cells in the hypothalamus and transported via the hypothalamic-hypophyseal tracts to the posterior pituitary where it is stored until released upon nervous stimulation. The primary trigger prompting the hypothalamus to release ADH is increasing osmolarity of tissue fluid, usually in response to significant loss of blood volume. ADH signals its target cells in the kidneys to reabsorb more water, thus preventing the loss of additional fluid in the urine. This will increase overall fluid levels and help restore blood volume and pressure. In addition, ADH constricts peripheral vessels.
Renin-Angiotensin-Aldosterone System (RAAS)
The renin-angiotensin-aldosterone mechanism has a major effect upon the cardiovascular system (Figure 6.9.3). Renin is an enzyme, although because of its importance in the renin-angiotensin-aldosterone pathway, some sources identify it as a hormone. Specialised cells in the kidneys found in the juxtaglomerular apparatus respond to decreased blood flow by secreting renin into the blood. Renin converts the plasma protein angiotensinogen, which is produced by the liver, into its active form—angiotensin I. Angiotensin I circulates in the blood and is then converted into angiotensin II in the lungs. This reaction is catalysed by the enzyme angiotensin-converting enzyme (ACE).
Angiotensin II is a powerful vasoconstrictor, greatly increasing blood pressure. It also stimulates the release of ADH and aldosterone, a hormone produced by the adrenal cortex. Aldosterone increases the reabsorption of sodium into the blood by the kidneys. Since water follows sodium, this increases the reabsorption of water. This in turn increases blood volume, raising blood pressure. Angiotensin II also stimulates the thirst centre in the hypothalamus, so an individual will likely consume more fluids, again increasing blood volume and pressure.
Figure 6.9.3. Hormones involved in renal control of blood pressure. In the renin-angiotensin-aldosterone mechanism, increasing angiotensin II will stimulate the production of antidiuretic hormone and aldosterone. In addition to renin, the kidneys produce erythropoietin, which stimulates the production of red blood cells, further increasing blood volume.
Erythropoietin (EPO)
Erythropoietin (EPO) is released by the kidneys when blood flow and/or oxygen levels decrease. EPO stimulates the production of erythrocytes within the bone marrow. Erythrocytes are the major formed element of the blood and may contribute 40 percent or more to blood volume, a significant factor of viscosity, resistance, pressure, and flow. In addition, EPO is a vasoconstrictor. Overproduction of EPO or excessive intake of synthetic EPO, often to enhance athletic performance, will increase viscosity, resistance, and pressure, and decrease flow in addition to its contribution as a vasoconstrictor.
Atrial Natriuretic Peptide (ANP)
Secreted by cells in the atria of the heart, atrial natriuretic hormone (ANH) or also known as atrial natriuretic peptide, is secreted when blood volume is high enough to cause extreme stretching of the cardiac cells. Cells in the ventricle produce a hormone with similar effects, called B-type natriuretic hormone. Natriuretic hormones are antagonists to angiotensin II. They promote loss of sodium and water from the kidneys, and suppress renin, aldosterone and ADH production and release. All of these actions promote loss of fluid from the body, so blood volume and blood pressure drop.
Autoregulation of Perfusion
As the name would suggest, autoregulation mechanisms require neither specialised nervous stimulation nor endocrine control. Rather, these are local, self-regulatory mechanisms that allow each region of tissue to adjust its blood flow—and thus its perfusion. These local mechanisms include chemical signals and myogenic controls.
Chemical Signals Involved in Autoregulation
Chemical signals work at the level of the precapillary sphincters to trigger either constriction or relaxation. As you know, opening a precapillary sphincter allows blood to flow into that particular capillary, whereas constricting a precapillary sphincter temporarily shuts off blood flow to that region. The factors involved in regulating the precapillary sphincters include the following:
Opening of the sphincter is triggered in response to decreased oxygen concentrations; increased carbon dioxide concentrations; increasing levels of lactic acid or other by-products of cellular metabolism; increasing concentrations of potassium ions or hydrogen ions (falling pH); inflammatory chemicals such as histamines; and increased body temperature. These conditions in turn stimulate the release of NO, a powerful vasodilator, from endothelial cells.
Contraction of the precapillary sphincter is triggered by the opposite levels of the regulators, which prompt the release of endothelins, powerful vasoconstricting peptides secreted by endothelial cells. Platelet secretions and certain prostaglandins may also trigger constriction.
Again, these factors alter tissue perfusion via their effects on the precapillary sphincter mechanism, which regulates blood flow to capillaries. Since the amount of blood is limited, not all capillaries can fill at once, so blood flow is allocated based upon the needs and metabolic state of the tissues as reflected in these parameters. Bear in mind, however, that dilation and constriction of the arterioles feeding the capillary beds is the primary control mechanism.
The Myogenic Response
The myogenic response is a reaction to the stretching of the smooth muscle in the walls of arterioles as changes in blood flow occur through the vessel. This may be viewed as a largely protective function against dramatic fluctuations in blood pressure and blood flow to maintain homeostasis. If perfusion of an organ is too low (ischaemia), the tissue will experience low levels of oxygen (hypoxia). In contrast, excessive perfusion could damage the organ’s smaller and more fragile vessels. The myogenic response is a localised process that serves to stabilise blood flow in the capillary network that follows that arteriole.
When blood flow is low, the vessel’s smooth muscle will be only minimally stretched. In response, it relaxes, allowing the vessel to dilate and thereby increase the movement of blood into the tissue. When blood flow is too high, the smooth muscle will contract in response to the increased stretch, prompting vasoconstriction that reduces blood flow.
Table 6.9.2 Summary of the effects of nervous, endocrine and local controls on arterioles.
Control
Factor
Vasoconstriction
Vasodilation
Neural
Sympathetic stimulation
Arterioles within integument, abdominal viscera, and mucosa membrane; skeletal muscle (at high levels); varied in veins and venules
Arterioles within heart; skeletal muscles at low to moderate levels
Parasympathetic
No known intervention for most
Arterioles in external genitalia, no known innervation for most other arterioles or veins
Endocrine
Adrenaline
Similar to sympathetic stimulation for extended flight-or-fight responses; at high levels, binds to specialised alpha (α) receptors
Similar to sympathetic stimulation for extended fight-or-flight responses; at low to moderate levels, binds to specialised beta (β) receptors
Noradrenaline
Similar to adrenaline
Similar to adrenaline
Angiotensin II
Powerful generalised vasoconstrictor; also stimulates release of aldosterone and ADH
n/a
ANH (peptide)
n/a
Powerful generalised vasodilator; also promotes loss of fluid volume from kidneys, hence reducing blood volume, pressure, and flow
ADH
Moderately strong generalised vasoconstrictor; also causes body to retain more fluid via kidneys, increasing blood volume and pressure
n/a
Other factors
Decreasing levels of oxygen
n/a
Vasodilation, also opens precapillary sphincters
Decreasing pH
n/a
Vasodilation, also opens precapillary sphincters
Increasing levels of carbon dioxide
n/a
Vasodilation, also opens precapillary sphincters
Increasing levels of potassium ion
n/a
Vasodilation, also opens precapillary sphincters
Increasing levels of prostaglandins
Vasoconstriction, closes precapillary sphincters for many
Vasodilation, opens precapillary sphincters for many
Increasing levels of adenosine
n/a
Vasodilation
Increasing levels of NO
n/a
Vasodilation, also opens precapillary sphincters
Increasing levels of lactic acid and other metabolites
n/a
Vasodilation, also opens precapillary sphincters
Increasing levels of endothelins
Vasoconstriction
n/a
Increasing levels of platelet secretion
Vasoconstriction
n/a
Increasing hyperthermia
n/a
Vasodilation
Stretching of vascular wall (myogenic)
Vasoconstriction
n/a
Increasing levels of histamines from basophils and mast cells
n/a
Vasodilation
Effect of Exercise on Vascular Homeostasis
The heart is a muscle and, like any muscle, it responds dramatically to exercise. For a healthy young adult, cardiac output (heart rate × stroke volume) increases in the nonathlete from approximately 5.0 litres per minute to a maximum of about 20 litres per minute. Accompanying this will be an increase in blood pressure from about 120/80 to 185/75. However, well-trained aerobic athletes can increase these values substantially. For these individuals, cardiac output soars from approximately 5.3 litres per minute resting to more than 30 litres per minute during maximal exercise. Along with this increase in cardiac output, blood pressure increases from 120/80 at rest to 200/90 at maximum values.
In addition to improved cardiac function, exercise increases the size and mass of the heart. The average weight of the heart for the nonathlete is about 300 g, whereas in an athlete it can increase to 500 g. This increase in size generally makes the heart stronger and more efficient at pumping blood, increasing both stroke volume and cardiac output.
Tissue perfusion also increases as the body transitions from a resting state to light exercise and eventually to heavy exercise. These changes result in selective vasodilation in the skeletal muscles, heart, lungs, liver, and integument. Simultaneously, vasoconstriction occurs in the vessels leading to the kidneys and most of the digestive and reproductive organs. The flow of blood to the brain remains largely unchanged whether at rest or exercising, since the vessels in the brain largely do not respond to regulatory stimuli, in most cases, because they lack the appropriate receptors.
As vasodilation occurs in selected vessels, resistance drops and more blood rushes into the organs they supply. This blood eventually returns to the venous system. Venous return is further enhanced by both the skeletal muscle and respiratory pumps. As blood returns to the heart more quickly, preload rises and the Frank-Starling principle tells us that contraction of the cardiac muscle in the atria and ventricles will be more forceful. Eventually, even the best-trained athletes will fatigue and must undergo a period of rest following exercise. Cardiac output and distribution of blood then return to normal.
Regular exercise promotes cardiovascular health in a variety of ways. Because an athlete’s heart is larger than a nonathlete’s, stroke volume increases, so the athletic heart can deliver the same amount of blood as the nonathletic heart but with a lower heart rate. This increased efficiency allows the athlete to exercise for longer periods of time before muscles fatigue and places less stress on the heart. Exercise also lowers overall cholesterol levels by removing from the circulation a complex form of cholesterol, triglycerides, and proteins known as low-density lipoproteins (LDLs), which are widely associated with increased risk of cardiovascular disease. Although there is no way to remove deposits of plaque from the walls of arteries other than specialised surgery, exercise does promote the health of vessels by decreasing the rate of plaque formation and reducing blood pressure, so the heart does not have to generate as much force to overcome resistance.
Generally as little as 30 minutes of noncontinuous exercise over the course of each day has beneficial effects and has been shown to lower the rate of heart attack by nearly 50 percent. While it is always advisable to follow a healthy diet, stop smoking, and lose weight, studies have clearly shown that fit, overweight people may actually be healthier overall than sedentary slender people. Thus, the benefits of moderate exercise are undeniable.
Clinical Considerations in Vascular Homeostasis
Any disorder that affects blood volume, vascular tone, or any other aspect of vascular functioning is likely to affect vascular homeostasis as well. That includes hypertension, haemorrhage, and shock.
Hypertension and Hypotension
Chronically elevated blood pressure is known clinically as hypertension. It is defined as chronic and persistent blood pressure measurements of 140/90 mm Hg or above. Pressures between 120/80 and 140/90 mm Hg are defined as prehypertension. The Australian Bureau of Statistics report that between 2017-2018, one in three Australian adults had high blood pressure, with hypertension making up over five percent of the total burden of disease in Australia. It has been estimated that over one billion people worldwide have hypertension, most of whom live in lower-income countries. Unfortunately, hypertension is typically a silent disorder; therefore, hypertensive patients may fail to recognise the seriousness of their condition and fail to follow their treatment plan. The result is often a heart attack or stroke. Hypertension may also lead to an aneurism (ballooning of a blood vessel caused by a weakening of the wall), peripheral arterial disease (obstruction of vessels in peripheral regions of the body), chronic kidney disease, or heart failure.
Figure 6.9.4. Aneurysm. A) Pseudoaneurysm of the left ventricle, four-chamber echocardiography view (image by Patrick J. Lynchused under CC-BY). B) Thoracic aortic aneurysm (image by BruceBlaus used under CC-BY).
Haemorrhage
Minor blood loss is managed by haemostasis and repair. Haemorrhage is a loss of blood that cannot be controlled by haemostatic mechanisms. Initially, the body responds to haemorrhage by initiating mechanisms aimed at increasing blood pressure and maintaining blood flow. Ultimately, however, blood volume will need to be restored, either through physiological processes or through medical intervention.
In response to blood loss, stimuli from the baroreceptors trigger the cardiovascular centres to stimulate sympathetic responses to increase cardiac output and vasoconstriction. This typically prompts the heart rate to increase to about 180–200 contractions per minute, restoring cardiac output to normal levels. Vasoconstriction of the arterioles increases vascular resistance, whereas constriction of the veins increases venous return to the heart. Both of these steps will help increase blood pressure. Sympathetic stimulation also triggers the release of adrenaline and noradrenaline, which enhance both cardiac output and vasoconstriction. If blood loss were less than 20 percent of total blood volume, these responses together would usually return blood pressure to normal and redirect the remaining blood to the tissues.
Additional endocrine involvement is necessary, however, to restore the lost blood volume. The angiotensin-renin-aldosterone mechanism stimulates the thirst centre in the hypothalamus, which increases fluid consumption to help restore the lost blood. More importantly, it increases renal reabsorption of sodium and water, reducing water loss in urine output. The kidneys also increase the production of EPO, stimulating the formation of erythrocytes that not only deliver oxygen to the tissues but also increase overall blood volume. Figure 6.9.5 summarises the responses to loss of blood volume.
Figure 6.9.5. Homeostatic responses to loss of blood volume.
Circulatory Shock
The loss of too much blood may lead to circulatory shock, a life-threatening condition in which the circulatory system is unable to maintain blood flow to adequately supply sufficient oxygen and other nutrients to the tissues to maintain cellular metabolism. It should not be confused with emotional or psychological shock. Typically, the patient in circulatory shock will demonstrate an increased heart rate but decreased blood pressure, but there are cases in which blood pressure will remain normal. Urine output will fall dramatically, and the patient may appear confused or lose consciousness. Urine output less than 1 mL/kg body weight/hour is cause for concern. Unfortunately, shock is an example of a positive-feedback loop that, if uncorrected, may lead to the death of the patient.
There are several recognised forms of shock:
Hypovolaemic shock in adults is typically caused by haemorrhage, although in children it may be caused by fluid losses related to severe vomiting or diarrhoea. Other causes for hypovolaemic shock include extensive burns, exposure to some toxins, and excessive urine loss related to diabetes insipidus or ketoacidosis. Typically, patients present with a rapid, almost tachycardic heart rate; a weak pulse often described as “thread;” cool, clammy skin, particularly in the extremities, due to restricted peripheral blood flow; rapid, shallow breathing; hypothermia; thirst; and dry mouth. Treatments generally involve providing intravenous fluids to restore the patient to normal function and various drugs such as dopamine, adrenaline, and noradrenaline to raise blood pressure.
Cardiogenic shock results from the inability of the heart to maintain cardiac output. Most often, it results from a myocardial infarction (heart attack), but it may also be caused by arrhythmias, valve disorders, cardiomyopathies, cardiac failure, or simply insufficient flow of blood through the cardiac vessels. Treatment involves repairing the damage to the heart or its vessels to resolve the underlying cause, rather than treating cardiogenic shock directly.
Vascular shock occurs when arterioles lose their normal muscular tone and dilate dramatically. It may arise from a variety of causes, and treatments almost always involve fluid replacement and medications, called inotropic or pressor agents, which restore tone to the muscles of the vessels. In addition, eliminating or at least alleviating the underlying cause of the condition is required. This might include antibiotics and antihistamines, or select steroids, which may aid in the repair of nerve damage. A common cause is sepsis (or septicaemia), also called “blood poisoning,” which is a widespread bacterial infection that results in an organismal-level inflammatory response known as septic shock. Neurogenic shock is a form of vascular shock that occurs with cranial or spinal injuries that damage the cardiovascular centres in the medulla oblongata or the nervous fibres originating from this region. Anaphylactic shock is a severe allergic response that causes the widespread release of histamines, triggering vasodilation throughout the body.
Obstructive shock, as the name would suggest, occurs when a significant portion of the vascular system is blocked. It is not always recognised as a distinct condition and may be grouped with cardiogenic shock, including pulmonary embolism and cardiac tamponade. Treatments depend upon the underlying cause and, in addition to administering fluids intravenously, often include the administration of anticoagulants, removal of fluid from the pericardial cavity, or air from the thoracic cavity, and surgery as required. The most common cause is a pulmonary embolism, a clot that lodges in the pulmonary vessels and interrupts blood flow. Other causes include stenosis of the aortic valve; cardiac tamponade, in which excess fluid in the pericardial cavity interferes with the ability of the heart to fully relax and fill with blood (resulting in decreased preload); and a pneumothorax, in which an excessive amount of air is present in the thoracic cavity, outside of the lungs, which interferes with venous return, pulmonary function, and delivery of oxygen to the tissues.
Section Review
Neural, endocrine, and autoregulatory mechanisms affect blood flow, blood pressure, and eventually perfusion of blood to body tissues. Neural mechanisms include the cardiovascular centres in the medulla oblongata, baroreceptors in the aorta and carotid arteries and right atrium, and associated chemoreceptors that monitor blood levels of oxygen, carbon dioxide, and hydrogen ions. Endocrine controls include adrenaline and noradrenaline, as well as ADH, the renin-angiotensin-aldosterone mechanism, ANP and EPO. Autoregulation is the local control of vasodilation and constriction by chemical signals and the myogenic response. Exercise greatly improves cardiovascular function and reduces the risk of cardiovascular diseases, including hypertension, a leading cause of heart attacks and strokes. Significant haemorrhage can lead to a form of circulatory shock known as hypovolemic shock. Sepsis, obstruction, and widespread inflammation can also cause circulatory shock.
Identify the vessels through which blood travels within the pulmonary circuit, beginning from the right ventricle of the heart and ending at the left atrium
Create a flow chart showing the major systemic arteries through which blood travels from the aorta and its major branches, to the most significant arteries feeding into the right and left upper and lower limbs
Create a flow chart showing the major systemic veins through which blood travels from the feet to the right atrium of the heart
Virtually every cell, tissue, organ, and system in the body is impacted by the circulatory system. This includes the generalised and more specialised functions of transport of materials, capillary exchange, maintaining health by transporting white blood cells and various immunoglobulins (antibodies), haemostasis, regulation of body temperature, and helping to maintain acid-base balance. In addition to these shared functions, many systems enjoy a unique relationship with the circulatory system. Figure 6.10.1 summarises these relationships.
Figure 6.10.1. Interaction of the circulatory system with other body systems.
As you learn about the vessels of the systemic and pulmonary circuits, notice that many arteries and veins share the same names, parallel one another throughout the body, and are very similar on the right and left sides of the body. These pairs of vessels will be traced through only one side of the body. Where differences occur in branching patterns or when vessels are singular, this will be indicated. For example, you will find a pair of femoral arteries and a pair of femoral veins, with one vessel on each side of the body. In contrast, some vessels closer to the midline of the body, such as the aorta, are unique. Moreover, some superficial veins, such as the great saphenous vein in the femoral region, have no arterial counterpart. Another phenomenon that can make the study of vessels challenging is that names of vessels can change with location. Like a street that changes name as it passes through an intersection, an artery or vein can change names as it passes an anatomical landmark. For example, the left subclavian artery becomes the axillary artery as it passes through the body wall and into the axillary region, and then becomes the brachial artery as it flows from the axillary region into the upper arm (or brachium). You will also find examples of anastomoses where two blood vessels that previously branched reconnect. Anastomoses are especially common in veins, where they help maintain blood flow even when one vessel is blocked or narrowed, although there are some important ones in the arteries supplying the brain.
As you read about circular pathways, notice that there is an occasional, very large artery referred to as a trunk, a term indicating that the vessel gives rise to several smaller arteries. For example, the celiac trunk gives rise to the left gastric, common hepatic, and splenic arteries.
As you study this section, imagine you are on a “Voyage of Discovery” similar to Lewis and Clark’s expedition in 1804–1806, which followed rivers and streams through unfamiliar territory, seeking a water route from the Atlantic to the Pacific Ocean. You might envision being inside a miniature boat, exploring the various branches of the circulatory system. This simple approach has proven effective for many students in mastering these major circulatory patterns. Another approach that works well for many students is to create simple line drawings similar to the ones provided, labelling each of the major vessels. It is beyond the scope of this text to name every vessel in the body. However, we will attempt to discuss the major pathways for blood and acquaint you with the major named arteries and veins in the body. Also, please keep in mind that individual variations in circulation patterns are not uncommon.
Pulmonary Circulation
Recall that blood returning from the systemic circuit enters the right atrium (Figure 6.10.2) via the superior and inferior venae cavae and the coronary sinus, which drains the blood supply of the heart muscle. These vessels will be described more fully later in this section. This blood is relatively low in oxygen and relatively high in carbon dioxide, since much of the oxygen has been extracted for use by the tissues and the waste gas carbon dioxide was picked up to be transported to the lungs for elimination. From the right atrium, blood moves into the right ventricle, which pumps it to the lungs for gas exchange. This system of vessels is referred to as the pulmonary circuit.
The single vessel exiting the right ventricle is the pulmonary trunk. At the base of the pulmonary trunk is the pulmonary semilunar valve, which prevents backflow of blood into the right ventricle during ventricular diastole. As the pulmonary trunk reaches the superior surface of the heart, it curves posteriorly and rapidly bifurcates (divides) into two branches, a left and a right pulmonary artery. To prevent confusion between these vessels, it is important to refer to the vessel exiting the heart as the pulmonary trunk, rather than also calling it a pulmonary artery. The pulmonary arteries in turn branch many times within the lung, forming a series of smaller arteries and arterioles that eventually lead to the pulmonary capillaries. The pulmonary capillaries surround lung structures known as alveoli that are the sites of oxygen and carbon dioxide exchange.
Once gas exchange is completed, oxygenated blood flows from the pulmonary capillaries into a series of pulmonary venules that eventually lead to a series of larger pulmonary veins. Four pulmonary veins, two on the left and two on the right, return blood to the left atrium. At this point, the pulmonary circuit is complete. Table 6.10.1 defines the major arteries and veins of the pulmonary circuit discussed in the text.
Figure 6.10.2. Pulmonary circuit. Blood exiting from the right ventricle flows into the pulmonary trunk, which bifurcates into the two pulmonary arteries. These vessels branch to supply blood to the pulmonary capillaries, where gas exchange occurs within the lung alveoli. Blood returns via the pulmonary veins to the left atrium.
Table 6.10.1. Pulmonary arteries and veins
Vessel
Description
Pulmonary trunk
Single large vessel exiting the right ventricle that divides to form the right and left pulmonary arteries
Pulmonary arteries
Left and right vessels that form from the pulmonary trunk and lead to smaller arterioles and eventually to the pulmonary capillaries
Pulmonary veins
Two sets of paired vessels – one pair on each side – that are formed from the small venules, leading away from the pulmonary capillaries to flow into the left atrium
Overview of Systemic Arteries
Blood relatively high in oxygen concentration is returned from the pulmonary circuit to the left atrium via the four pulmonary veins. From the left atrium, blood moves into the left ventricle, which pumps blood into the aorta. The aorta and its branches—the systemic arteries—send blood to virtually every organ of the body (Figure 6.10.3).
Figure 6.10.3. Systemic arteries. The major systemic arteries shown here deliver oxygenated blood throughout the body.
The Aorta
The aorta is the largest artery in the body (Figure 6.10.4). It arises from the left ventricle and eventually descends to the abdominal region, where it bifurcates at the level of the fourth lumbar vertebra into the two common iliac arteries. The aorta consists of the ascending aorta, the aortic arch, and the descending aorta, which passes through the diaphragm and a landmark that divides into the superior thoracic and inferior abdominal components. Arteries originating from the aorta ultimately distribute blood to virtually all tissues of the body. At the base of the aorta is the aortic semilunar valve that prevents backflow of blood into the left ventricle while the heart is relaxing. After exiting the heart, the ascending aorta moves in a superior direction for approximately 5 cm and ends at the sternal angle. Following this ascent, it reverses direction, forming a graceful arc to the left, called the aortic arch. The aortic arch descends toward the inferior portions of the body and ends at the level of the intervertebral disk between the fourth and fifth thoracic vertebrae. Beyond this point, the descending aorta continues close to the bodies of the vertebrae and passes through an opening in the diaphragm known as the aortic hiatus. Superior to the diaphragm, the aorta is called the thoracic aorta, and inferior to the diaphragm, it is called the abdominal aorta. The abdominal aorta terminates when it bifurcates into the two common iliac arteries at the level of the fourth lumbar vertebra. See Figure 6.10.4 for an illustration of the ascending aorta, the aortic arch, and the initial segment of the descending aorta plus major branches; Table 6.10.2 summarises the structures of the aorta.
Figure 6.10.4. Aorta. The aorta has distinct regions, including the ascending aorta, aortic arch, and the descending aorta, which includes the thoracic and abdominal regions.
Table 6.10.2. Components of the aorta
Vessel
Description
Aorta
Largest artery in the body, originating from the left ventricle and descending to the abdominal region, where it bifurcates into the common iliac arteries at the level of the fourth lumbar vertebra; arteries originating from the aorta distribute blood to virtually all tissues of the body
Ascending aorta
Initial portion of the aorta, rising superiorly from the left ventricle for distance of approximately 5cm
Aortic arch
Graceful arc to the left that connects the ascending aorta to the descending aorta; ends at the intervertebral disk between the fourth and fifth thoracic vertebrae
Descending aorta
Portion of the aorta that continues inferiorly past the end of the aortic arch; subdivided into the thoracic aorta and the abdominal aorta
Thoracic aorta
Portion of the descending aorta superior to the aortic hiatus
Abdominal aorta
Portion of the aorta inferior to the aortic hiatus and superior to the common iliac arteries
Coronary Circulation
The first vessels that branch from the ascending aorta are the paired coronary arteries (see Figure 6.10.4), which arise from two of the three sinuses in the ascending aorta just superior to the aortic semilunar valve. These sinuses contain the aortic baroreceptors and chemoreceptors critical to maintain cardiac function. The left coronary artery arises from the left posterior aortic sinus. The right coronary artery arises from the anterior aortic sinus. Normally, the right posterior aortic sinus does not give rise to a vessel.
The coronary arteries encircle the heart, forming a ring-like structure that divides into the next level of branches that supplies blood to the heart tissues.
Aortic Arch Branches
There are three major branches of the aortic arch: the brachiocephalic artery, the left common carotid artery, and the left subclavian (literally “under the clavicle”) artery. As you would expect based upon proximity to the heart, each of these vessels is classified as an elastic artery.
The brachiocephalic artery is located only on the right side of the body; there is no corresponding artery on the left. The brachiocephalic artery branches into the right subclavian artery and the right common carotid artery. The left subclavian and left common carotid arteries arise independently from the aortic arch but otherwise follow a similar pattern and distribution to the corresponding arteries on the right side (see Figure 6.10.2).
Each subclavian artery supplies blood to the arms, chest, shoulders, back, and central nervous system. It then gives rise to three major branches: the internal thoracic artery, the vertebral artery, and the thyrocervical artery. The internal thoracic artery, or mammary artery, supplies blood to the thymus, the pericardium of the heart, and the anterior chest wall. The vertebral artery passes through the vertebral foramen in the cervical vertebrae and then through the foramen magnum into the cranial cavity to supply blood to the brain and spinal cord. The paired vertebral arteries join together to form the large basilar artery at the base of the medulla oblongata. This is an example of an anastomosis. The subclavian artery also gives rise to the thyrocervical artery that provides blood to the thyroid, the cervical region of the neck, and the upper back and shoulder.
The common carotid artery divides into internal and external carotid arteries. The right common carotid artery arises from the brachiocephalic artery and the left common carotid artery arises directly from the aortic arch. The external carotid artery supplies blood to numerous structures within the face, lower jaw, neck, oesophagus, and larynx. These branches include the lingual, facial, occipital, maxillary, and superficial temporal arteries. The internal carotid artery initially forms an expansion known as the carotid sinus, containing the carotid baroreceptors and chemoreceptors. Like their counterparts in the aortic sinuses, the information provided by these receptors is critical to maintaining cardiovascular homeostasis (see Figure 6.10.2).
The internal carotid arteries along with the vertebral arteries are the two primary suppliers of blood to the human brain. Given the central role and vital importance of the brain to life, it is critical that blood supply to this organ remains uninterrupted. Recall that blood flow to the brain is remarkably constant, with approximately 20 percent of blood flow directed to this organ at any given time. When blood flow is interrupted, even for just a few seconds, a transient ischemic attack (TIA), or mini-stroke, may occur, resulting in loss of consciousness or temporary loss of neurological function. In some cases, the damage may be permanent. Loss of blood flow for longer periods, typically between 3 and 4 minutes, will likely produce irreversible brain damage or a stroke, also called a cerebrovascular accident (CVA). The locations of the arteries in the brain not only provide blood flow to the brain tissue but also prevent interruption in the flow of blood. Both the carotid and vertebral arteries branch once they enter the cranial cavity, and some of these branches form a structure known as the arterial circle (or circle of Willis), an anastomosis that is remarkably like a traffic circle that sends off branches (in this case, arterial branches to the brain). As a rule, branches to the anterior portion of the cerebrum are normally fed by the internal carotid arteries; the remainder of the brain receives blood flow from branches associated with the vertebral arteries.
The internal carotid artery continues through the carotid canal of the temporal bone and enters the base of the brain through the carotid foramen where it gives rise to several branches (Figure 6.10.5 and Figure 6.10.6). One of these branches is the anterior cerebral artery that supplies blood to the frontal lobe of the cerebrum. Another branch, the middle cerebral artery, supplies blood to the temporal and parietal lobes, which are the most common sites of CVAs. The ophthalmic artery, the third major branch, provides blood to the eyes.
The right and left anterior cerebral arteries join together to form an anastomosis called the anterior communicating artery. The initial segments of the anterior cerebral arteries and the anterior communicating artery form the anterior portion of the arterial circle. The posterior portion of the arterial circle is formed by a left and a right posterior communicating artery that branches from the posterior cerebral artery, which arises from the basilar artery. It provides blood to the posterior portion of the cerebrum and brain stem. The basilar artery is an anastomosis that begins at the junction of the two vertebral arteries and sends branches to the cerebellum and brain stem. It flows into the posterior cerebral arteries. Table 6.10.3 summarises the aortic arch branches, including the major branches supplying the brain.
Figure 6.10.5. Arteries supplying the head and neck. The common carotid artery gives rise to the external and internal carotid arteries. The external carotid artery remains superficial and gives rise to many arteries of the head. The internal carotid artery first forms the carotid sinus and then reaches the brain via the carotid canal and carotid foramen, emerging into the cranium via the foramen lacerum. The vertebral artery branches from the subclavian artery and passes through the transverse foramen in the cervical vertebrae, entering the base of the skull at the vertebral foramen. The subclavian artery continues toward the arm as the axillary artery.Figure 6.10.6. Arteries serving the brain. This inferior view shows the network of arteries serving the brain. The structure is referred to as the arterial circle or circle of Willis.
Table 6.10.3. Aortic arch branches and brain circulation
Vessel
Description
Brachiocephalic artery
Single vessel located on the right side of the body; the first vessel branching from the aortic arch; gives rise to the right subclavian artery and the right common carotid artery; supplies blood to the head, neck, upper limb, and wall of the thoracic region
Subclavian artery
The right subclavian artery arises from the brachiocephalic artery while the left subclavian artery arises from the aortic arch; gives rise to the internal thoracic, vertebral, and thyrocervical arteries; supplies blood to the arms, chest, shoulders, back, and central nervous system
Internal thoracic artery
Also called the mammary artery; arises from the subclavian artery; supplies blood to the thymus, pericardium of the heart, and anterior chest wall
Vertebral artery
Arises from the subclavian artery and passes through the vertebral foramen through the foramen magnum to the brain; joins with the internal carotid artery to form the arterial circle; supplies blood to the brain and spinal cord
Thyrocervical artery
Arises from the subclavian artery; supplies blood to the thyroid, the cervical region, the upper back, and shoulder
Common carotid artery
The right common carotid artery arises from the brachiocephalic artery and the left common carotid artery arises from the aortic arch; gives rise to the external and internal carotid arteries; supplies the respective sides of the head and neck
External carotid artery
Arises from the common carotid artery; supplies blood to the numerous structures within the face, lower jaw, neck, oesophagus, and larynx
Internal carotid artery
Arises from the common carotid artery and begins the carotid sinus; goes through the carotid canal of the temporal bone to the base of the brain; combines with the branches of the vertebral artery, forming the arterial circle; supplies blood to the brain
Arterial circle or circle of Willis
An anastomosis located at the base of the brain that ensures continual blood supply; formed from the branches of the internal carotid and vertebral arteries; supplies blood to the brain
Middle cerebral artery
Another branch of the internal carotid artery; supplies blood to the temporal and parietal lobes of the cerebrum
Ophthalmic artery
Branch of the internal carotid artery; supplies blood to the eyes
Anterior communicating artery
An anastomosis of the right and left internal carotid arteries; supplies blood to the brain
Posterior communicating artery
Branches of the posterior cerebral artery that form part of the posterior portion of the arterial circle; supplies blood to the brain
Posterior cerebral artery
Branch of the basilar artery that forms a portion of the posterior segment of the arterial circle of Willis; supplies blood to the posterior portion of the cerebrum and brain stem
Basilar artery
Formed from the fusion of the two vertebral arteries; sends branches to the cerebellum, brainstem, and the posterior cerebral arteries; the main blood supply to the brain stem
Thoracic Aorta and Major Branches
The thoracic aorta begins at the level of vertebra T5 and continues through to the diaphragm at the level of T12, initially traveling within the mediastinum to the left of the vertebral column. As it passes through the thoracic region, the thoracic aorta gives rise to several branches, which are collectively referred to as visceral branches and parietal branches (Figure 6.10.7). Those branches that supply blood primarily to visceral organs are known as the visceral branches and include the bronchial arteries, pericardial arteries, oesophageal arteries, and the mediastinal arteries, each named after the tissues it supplies. Each bronchial artery (typically two on the left and one on the right) supplies systemic blood to the lungs and visceral pleura, in addition to the blood pumped to the lungs for oxygenation via the pulmonary circuit. The bronchial arteries follow the same path as the respiratory branches, beginning with the bronchi and ending with the bronchioles. There is considerable, but not total, intermingling of the systemic and pulmonary blood at anastomoses in the smaller branches of the lungs. This may sound incongruous—that is, the mixing of systemic arterial blood high in oxygen with the pulmonary arterial blood lower in oxygen—but the systemic vessels also deliver nutrients to the lung tissue just as they do elsewhere in the body. The mixed blood drains into typical pulmonary veins, whereas the bronchial artery branches remain separate and drain into bronchial veins described later. Each pericardial artery supplies blood to the pericardium, the oesophageal artery provides blood to the oesophagus, and the mediastinal artery provides blood to the mediastinum. The remaining thoracic aorta branches are collectively referred to as parietal branches or somatic branches and include the intercostal and superior phrenic arteries. Each intercostal artery provides blood to the muscles of the thoracic cavity and vertebral column. The superior phrenic artery provides blood to the superior surface of the diaphragm. Table 6.10.4 lists the arteries of the thoracic region.
Figure 6.10.7. Arteries of the thoracic and abdominal regions. The thoracic aorta gives rise to the arteries of the visceral and parietal branches.
Table 6.10.4. Arteries of the thoracic region
Vessel
Description
Visceral branches
A group of arterial branches of the thoracic aorta; supplies blood to the viscera (i.e., organs) of the thorax
Bronchial artery
Systemic branch from the aorta that provides oxygenated blood to the lungs; this blood supply is in addition to the pulmonary circuit that brings blood for oxygenation
Pericardial artery
Branch of the thoracic aorta; supplies blood to the pericardium
Oesophageal artery
Branch of the thoracic aorta; supplies blood to the oesophagus
Mediastinal artery
Branch of the thoracic aorta; supplies blood to the mediastinum
Parietal branches
Also called somatic branches, a group of arterial branches of the thoracic aorta; include those that supply blood to the thoracic wall, vertebral column, and the superior surface of the diaphragm
Intercostal artery
Branch of the thoracic aorta; supplies blood to the muscles of the thoracic cavity and vertebral column
Superior phrenic artery
Branch of the thoracic aorta; supplies blood to the superior surface of the diaphragm
Abdominal Aorta and Major Branches
After crossing through the diaphragm at the aortic hiatus, the thoracic aorta is called the abdominal aorta (see Figure 6.10.7). This vessel remains to the left of the vertebral column and is embedded in adipose tissue behind the peritoneal cavity. It formally ends at approximately the level of vertebra L4, where it bifurcates to form the common iliac arteries. Before this division, the abdominal aorta gives rise to several important branches. A single celiac trunk (artery) emerges and divides into the left gastric artery to supply blood to the stomach and oesophagus, the splenic artery to supply blood to the spleen, and the common hepatic artery, which in turn gives rise to the hepatic artery proper to supply blood to the liver, the right gastric artery to supply blood to the stomach, the cystic artery to supply blood to the gall bladder, and several branches, one to supply blood to the duodenum and another to supply blood to the pancreas. Two additional single vessels arise from the abdominal aorta. These are the superior and inferior mesenteric arteries. The superior mesenteric artery arises approximately 2.5 cm after the celiac trunk and branches into several major vessels that supply blood to the small intestine (duodenum, jejunum, and ileum), the pancreas, and a majority of the large intestine. The inferior mesenteric artery supplies blood to the distal segment of the large intestine, including the rectum. It arises approximately 5 cm superior to the common iliac arteries.
In addition to these single branches, the abdominal aorta gives rise to several significant paired arteries along the way. These include the inferior phrenic arteries, the adrenal arteries, the renal arteries, the gonadal arteries, and the lumbar arteries. Each inferior phrenic artery is a counterpart of a superior phrenic artery and supplies blood to the inferior surface of the diaphragm. The adrenal artery supplies blood to the adrenal (suprarenal) glands and arises near the superior mesenteric artery. Each renal artery branches approximately 2.5 cm inferior to the superior mesenteric arteries and supplies a kidney. The right renal artery is longer than the left since the aorta lies to the left of the vertebral column and the vessel must travel a greater distance to reach its target. Renal arteries branch repeatedly to supply blood to the kidneys. Each gonadal artery supplies blood to the gonads, or reproductive organs, and is also described as either an ovarian artery or a testicular artery (internal spermatic), depending upon the sex of the individual. An ovarian artery supplies blood to an ovary, uterine (Fallopian) tube, and the uterus, and is located within the suspensory ligament of the uterus. It is considerably shorter than a testicular artery, which ultimately travels outside the body cavity to the testes, forming one component of the spermatic cord. The gonadal arteries arise inferior to the renal arteries and are generally retroperitoneal. The ovarian artery continues to the uterus where it forms an anastomosis with the uterine artery that supplies blood to the uterus. Both the uterine arteries and vaginal arteries, which distribute blood to the vagina, are branches of the internal iliac artery. The four paired lumbar arteries are the counterparts of the intercostal arteries and supply blood to the lumbar region, the abdominal wall, and the spinal cord. In some instances, a fifth pair of lumbar arteries emerges from the median sacral artery.
The aorta divides at approximately the level of vertebra L4 into a left and a right common iliac artery but continues as a small vessel, the median sacral artery, into the sacrum. The common iliac arteries provide blood to the pelvic region and ultimately to the lower limbs. They split into external and internal iliac arteries approximately at the level of the lumbar-sacral articulation. Each internal iliac artery sends branches to the urinary bladder, the walls of the pelvis, the external genitalia, and the medial portion of the femoral region. In females, they also provide blood to the uterus and vagina. The much larger external iliac artery supplies blood to each of the lower limbs. Figure 6.10.8 shows the distribution of the major branches of the aorta into the thoracic and abdominal regions. Figure 6.10.9 shows the distribution of the major branches of the common iliac arteries. Table 6.10.5 summarises the major branches of the abdominal aorta.
Figure 6.10.8. Major branches of the aorta. The flow chart summarises the distribution of the major branches of the aorta into the thoracic and abdominal regions.
Major Branches of the Iliac Arteries
The flow chart summarises the distribution of the major branches of the common iliac arteries into the pelvis and lower limbs. The left side follows a similar pattern to the right.
Figure 6.10.9. Major branches of the iliac arteries. The flow chart summarises the distribution of the major branches of the common iliac arteries into the pelvis and lower limbs. The left side follows a similar pattern to the right.
Table 6.10.5. Vessels of the abdominal aorta
Vessel
Description
Coeliac trunk
Also called the coeliac artery; a major branch of the abdominal aorta; gives rise to the left gastric artery, the splenic artery, and the common hepatic artery that forms the hepatic artery to the liver, the right gastric artery to the stomach and the cystic artery to the gall bladder
Left gastric artery
Branch of the coeliac trunk; supplies blood to the stomach
Splenic artery
Branch of the coeliac trunk; supplies blood to the spleen
Common hepatic artery
Branch of the coeliac trunk that forms the hepatic artery, the right gastric artery, and the cystic artery
Hepatic artery proper
Branch of the common hepatic artery; supplies systemic blood to the liver
Right gastric artery
Branch of the common hepatic artery; supplies blood to the stomach
Cystic artery
Branch of the common hepatic artery; supplies blood to the gall bladder
Superior mesenteric artery
Branches of the abdominal aorta; supplies blood to the small intestine (duodenum, jejunum and ileum), the pancreas, and a majority of the large intestine
Inferior mesenteric artery
Branch of the abdominal aorta; supplies blood to the distal segment of the large intestine and rectum
Inferior phrenic arteries
Branches of the abdominal aorta; supply blood to the inferior surface of the diaphragm
Adrenal artery
Branch of the abdominal aorta; supplies blood to the adrenal (suprarenal) glands
Renal artery
Branch of the abdominal aorta; supplies each kidney
Gonadal artery
Branch of the abdominal aorta; supplies blood to the gonads or reproductive organs; also described as ovarian arteries or testicular arteries, depending upon the sex of the individual
Ovarian artery
Branch of the abdominal aorta; supplies blood to ovary, uterine (Fallopian) tube, and uterus
Testicular artery
Branch of the abdominal aorta; ultimately travels outside the body cavity to the testes and forms one component of the spermatic cord
Common iliac artery
Branch of the aorta that leads to the internal and external uterus and vagina
External iliac artery
Branch of the common iliac artery that leaves the body cavity and becomes a femoral artery; supplies blood to the lower limbs
Arteries Serving the Upper Limbs
As the subclavian artery exits the thorax into the axillary region, it is renamed the axillary artery. Although it does branch and supply blood to the region near the head of the humerus (via the humeral circumflex arteries), the majority of the vessel continues into the upper arm, or brachium, and becomes the brachial artery (Figure 6.10.10). The brachial artery supplies blood to much of the brachial region and divides at the elbow into several smaller branches, including the deep brachial arteries, which provide blood to the posterior surface of the arm, and the ulnar collateral arteries, which supply blood to the region of the elbow. As the brachial artery approaches the coronoid fossa, it bifurcates into the radial and ulnar arteries, which continue into the forearm, or antebrachium. The radial artery and ulnar artery parallel their namesake bones, giving off smaller branches until they reach the wrist, or carpal region. At this level, they fuse to form the superficial and deep palmar arches that supply blood to the hand, as well as the digital arteries that supply blood to the digits. Figure 6.10.11 shows the distribution of systemic arteries from the heart into the upper limb. Table 6.10.6 summarises the arteries serving the upper limbs.
Figure 6.10.10. Major arteries serving the thorax and upper limb. The arteries that supply blood to the arms and hands are extensions of the subclavian arteries.Figure 6.10.11. Major arteries of the upper limb. The flow chart summarises the distribution of the major arteries from the heart into the upper limb.
Table 6.10.6. Arteries serving the upper limbs
Vessel
Description
Axillary artery
Continuation of the subclavian artery as it penetrates the body wall and enters the axillary region; supplies blood to the region near the head of the humerus (humeral circumflex arteries); the majority of the vessel continues into the brachium and becomes the brachial artery
Brachial artery
Continuation of the axillary artery in the brachium; supplies blood to much of the brachial region; gives off several smaller branches that provide blood to the posterior surface of the arm in the region of the elbow; bifurcates into the radial and ulnar arteries at the coronoid fossa
Radial artery
Formed at the bifurcation of the brachial artery; parallels the radius; gives off smaller branches until it reaches the carpal region where it fuses with the ulnar artery to form the superficial and deep palmar arches; supplies blood to the lower arm and carpal region
Ulnar artery
Formed at the bifurcation of the brachial artery; parallels the ulna; gives off smaller brunches until it reaches the carpal region where it fuses with the radial artery to form the superficial and deep palmar arches; supplies blood to the lower arm and carpal region
Palmar arches
(superficial and deep)
Formed from anastomosis of the radial and ulnar arteries; supply blood to the hand and digital arteries
Digital arteries
Formed from the superficial and deep palmar arches; supply blood to the digits
Arteries Serving the Lower Limbs
The external iliac artery exits the body cavity and enters the femoral region of the lower leg (Figure 6.10.12). As it passes through the body wall, it is renamed the femoral artery. It gives off several smaller branches as well as the lateral deep femoral artery that in turn gives rise to a lateral circumflex artery. These arteries supply blood to the deep muscles of the thigh as well as ventral and lateral regions of the integument. The femoral artery also gives rise to the genicular artery, which provides blood to the region of the knee. As the femoral artery passes posterior to the knee near the popliteal fossa, it is called the popliteal artery. The popliteal artery branches into the anterior and posterior tibial arteries.
The anterior tibial artery is located between the tibia and fibula and supplies blood to the muscles and integument of the anterior tibial region. Upon reaching the tarsal region, it becomes the dorsalis pedis artery, which branches repeatedly and provides blood to the tarsal and dorsal regions of the foot. The posterior tibial artery provides blood to the muscles and integument on the posterior surface of the tibial region. The fibular or peroneal artery branches from the posterior tibial artery. It bifurcates and becomes the medial plantar artery and lateral plantar artery, providing blood to the plantar surfaces. There is an anastomosis with the dorsalis pedis artery, and the medial and lateral plantar arteries form two arches called the dorsal arch (also called the arcuate arch) and the plantar arch, which provide blood to the remainder of the foot and toes. Figure 6.10.13 shows the distribution of the major systemic arteries in the lower limb. Table 6.10.7 summarises the major systemic arteries discussed in the text.
Figure 6.10.12. Major arteries serving the lower limb. Major arteries serving the lower limb are shown in anterior and posterior views.Figure 13. Systemic arteries of the lower limb. The flow chart summarises the distribution of the systemic arteries from the external iliac artery into the lower limb.
Table 6.10.7. Arteries serving the lower limbs
Vessel
Description
Femoral artery
Continuation of the external iliac artery after it passes through the body cavity; divides into several smaller branches, the lateral deep femoral artery, and the genicular artery; becomes the popliteal artery as it passes posterior to the knee
Deep femoral artery
Branch of the femoral artery; gives rise to the lateral circumflex arteries
Lateral circumflex artery
Branch of the deep femoral artery; supplies blood to the deep muscles of the thigh and the ventral and lateral regions of the integument
Genicular artery
Branch of the femoral artery; supplies blood to the region of the knee
Popliteal artery
Continuation of the femoral artery posterior to the knee; branches into the anterior and posterior tibial arteries
Anterior tibial artery
Branches from the popliteal artery; supplies blood to the anterior tibial region; becomes the dorsalis pedis artery
Dorsalis pedis artery
Forms the anterior tibial artery; branches repeatedly to supply blood to the tarsal and dorsal regions of the foot
Posterior tibial artery
Branches from the popliteal artery and gives rise to the fibular or peroneal artery; supplies blood to the posterior tibial region
Medial planar artery
Arises from the bifurcation of the posterior tibial arteries; supplies blood to the medial plantar surfaces of the foot
Lateral plantar artery
Arises from the bifurcation of the posterior tibial arteries; supplies blood to the lateral plantar surface of the foot
Dorsal or arcuate arch
Formed from the anastomosis of the dorsalis pedis artery and the medial and plantar arteries; branches supply the distal portions of the foot and digits
Plantar arch
Formed from the anastomosis of the dorsalis pedis artery and the medial and plantar arteries; branches supply the distal portions of the foot and digits
Overview of Systemic Veins
Systemic veins return blood to the right atrium. Since the blood has already passed through the systemic capillaries, it will be relatively low in oxygen concentration. In many cases, there will be veins draining organs and regions of the body with the same name as the arteries that supplied these regions and the two often parallel one another. This is often described as a “complementary” pattern. However, there is a great deal more variability in the venous circulation than normally occurs in the arteries. For the sake of brevity and clarity, this text will discuss only the most commonly encountered patterns. However, keep this variation in mind when you move from the classroom to clinical practice.
In both the neck and limb regions, there are often both superficial and deeper levels of veins. The deeper veins generally correspond to the complementary arteries. The superficial veins do not normally have direct arterial counterparts, but in addition to returning blood, they also make contributions to the maintenance of body temperature. When the ambient temperature is warm, more blood is diverted to the superficial veins where heat can be more easily dissipated to the environment. In colder weather, there is more constriction of the superficial veins and blood is diverted deeper where the body can retain more of the heat.
The “Voyage of Discovery” analogy and stick drawings mentioned earlier remain valid techniques for the study of systemic veins, but veins present a more difficult challenge because there are numerous anastomoses and multiple branches. It is like following a river with many tributaries and channels, several of which interconnect. Tracing blood flow through arteries follows the current in the direction of blood flow, so that we move from the heart through the large arteries and into the smaller arteries to the capillaries. From the capillaries, we move into the smallest veins and follow the direction of blood flow into larger veins and back to the heart. Figure 6.10.14 outlines the path of the major systemic veins.
Figure 6.10.14. Major systemic veins of the body. The major systemic veins of the body are shown here in an anterior view.
The right atrium receives all the systemic venous return. Most of the blood flows into either the superior vena cava or inferior vena cava. If you draw an imaginary line at the level of the diaphragm, systemic venous circulation from above that line will generally flow into the superior vena cava; this includes blood from the head, neck, chest, shoulders, and upper limbs. The exception to this is that most venous blood flow from the coronary veins flows directly into the coronary sinus and from there directly into the right atrium. Beneath the diaphragm, systemic venous flow enters the inferior vena cava, that is, blood from the abdominal and pelvic regions and the lower limbs.
The Superior Vena Cava
The superior vena cava drains most of the body superior to the diaphragm (Figure 6.10.15). On both the left and right sides, the subclavian vein forms when the axillary vein passes through the body wall from the axillary region. It fuses with the external and internal jugular veins from the head and neck to form the brachiocephalic vein. Each vertebral vein also flows into the brachiocephalic vein close to this fusion. These veins arise from the base of the brain and the cervical region of the spinal cord, and flow largely through the intervertebral foramina in the cervical vertebrae. They are the counterparts of the vertebral arteries. Each internal thoracic vein, also known as an internal mammary vein, drains the anterior surface of the chest wall and flows into the brachiocephalic vein.
The remainder of the blood supply from the thorax drains into the azygos vein. Each intercostal vein drains muscles of the thoracic wall, each oesophageal vein delivers blood from the inferior portions of the oesophagus, each bronchial vein drains the systemic circulation from the lungs, and several smaller veins drain the mediastinal region. Bronchial veins carry approximately 13 percent of the blood that flows into the bronchial arteries; the remainder intermingles with the pulmonary circulation and returns to the heart via the pulmonary veins. These veins flow into the azygos vein, and with the smaller hemiazygos vein (hemi- = “half”) on the left of the vertebral column, drain blood from the thoracic region. The hemiazygos vein does not drain directly into the superior vena cava but enters the brachiocephalic vein via the superior intercostal vein.
The azygos vein passes through the diaphragm from the thoracic cavity on the right side of the vertebral column and begins in the lumbar region of the thoracic cavity. It flows into the superior vena cava at approximately the level of T2, making a significant contribution to the flow of blood. It combines with the two large left and right brachiocephalic veins to form the superior vena cava.
Table 6.10.8 summarises the veins of the thoracic region that flow into the superior vena cava.
Figure 6.10.15. Veins of the thoracic and abdominal regions. Veins of the thoracic and abdominal regions drain blood from the area above the diaphragm, returning it to the right atrium via the superior vena cava.
Table 6.10.8. Veins of the thoracic region
Vessel
Description
Superior vena cava
Large systemic vein; drains blood from most areas superior to the diaphragm; empties into the right atrium
Subclavian vein
Located deep in the thoracic cavity; formed by the axillary vein as it enters the thoracic cavity from the axillary region; drains the axillary and smaller local veins near the scapular region and leads to the brachiocephalic vein
Brachiocephalic vein
Pair of veins that form from a fusion of the external and internal jugular veins and the subclavian vein; subclavian, external and internal jugulars, vertebral and internal thoracic veins flow into it; drain the upper thoracic region and lead to the superior vena cava
Vertebral vein
Arises from the base of the brain and the cervical region of the spinal cord; passes through the intervertebral foramina in the cervical vertebrae; drains smaller veins from the cranium, spinal cord, and vertebrae, and leads to the brachiocephalic vein; counterpart of the vertebral artery
Internal thoracic veins
Also called internal mammary veins; drain the anterior surface of the chest wall and lead to the brachiocephalic vein
Intercostal vein
Drains the inferior portions of the oesophagus and leads to the azygos vein
Bronchial vein
Drains the systemic circulation from the lungs and leads to the azygos vein
Azygos vein
Originates in the lumbar region and passes through the diaphragm into the thoracic cavity on the right side of the vertebral column; drains blood from the intercostal veins, oesophageal veins, bronchial veins, and other veins draining the mediastinal region, and leads to the superior vena cava
Hemiazygos vein
Smaller vein complementary to the azygos vein; drains the oesophageal veins from the oesophagus and the left intercostal veins, and leads to the brachiocephalic vein via the superior intercostal vein
Veins of the Head and Neck
Blood from the brain and the superficial facial vein flow into each internal jugular vein (Figure 6.10.16). Blood from the more superficial portions of the head, scalp, and cranial regions, including the temporal vein and maxillary vein, flow into each external jugular vein. Although the external and internal jugular veins are separate vessels, there are anastomoses between them close to the thoracic region. Blood from the external jugular vein empties into the subclavian vein. Table 6.10.9 summarises the major veins of the head and neck.
Table 6.10.9. Major veins of the head and neck
Vessel
Description
Internal jugular vein
Parallel to the common carotid artery, which is more or less its counterpart, and passes through the jugular foramen and canal; primarily drains blood from the brain, receives the superficial facial vein, and empties into the subclavian vein
Temporal vein
Drains blood from the temporal region and flows into the external jugular vein
Maxillary vein
Drains blood from the maxillary region and flows into the external jugular vein
External jugular vein
Drains blood from the more superficial portions of the head, scalp, and cranial regions, and leads to the subclavian vein
Venous Drainage of the Brain
Circulation to the brain is both critical and complex (see Table 6.10.13). Many smaller veins of the brain stem and the superficial veins of the cerebrum lead to larger vessels referred to as intracranial sinuses. These include the superior and inferior sagittal sinuses, straight sinus, cavernous sinuses, left and right sinuses, the petrosal sinuses, and the occipital sinuses. Ultimately, sinuses will lead back to either the inferior jugular vein or vertebral vein.
Most of the veins on the superior surface of the cerebrum flow into the largest of the sinuses, the superior sagittal sinus. It is located midsagittally between the meningeal and periosteal layers of the dura mater within the falx cerebri and, at first glance in images or models, can be mistaken for the subarachnoid space. Most reabsorption of cerebrospinal fluid occurs via the chorionic villi (arachnoid granulations) into the superior sagittal sinus. Blood from most of the smaller vessels originating from the inferior cerebral veins flows into the great cerebral vein and into the straight sinus. Other cerebral veins and those from the eye socket flow into the cavernous sinus, which flows into the petrosal sinus and then into the internal jugular vein. The occipital sinus, sagittal sinus, and straight sinuses all flow into the left and right transverse sinuses near the lambdoid suture. The transverse sinuses in turn flow into the sigmoid sinuses that pass through the jugular foramen and into the internal jugular vein. The internal jugular vein flows parallel to the common carotid artery and is more or less its counterpart. It empties into the brachiocephalic vein. The veins draining the cervical vertebrae and the posterior surface of the skull, including some blood from the occipital sinus, flow into the vertebral veins. These parallel the vertebral arteries and travel through the transverse foramina of the cervical vertebrae. The vertebral veins also flow into the brachiocephalic veins. Table 6.10.10 summarises the major veins of the brain.
Figure 6.10.16. Veins of the head and neck. This left lateral view shows the veins of the head and neck, including the intercranial sinuses.
Table 6.10.10. Major veins of the brain
Vessel
Description
Superior sagittal sinus
Enlarged vein located midsagittally between the meningeal and periosteal layers of the dura mater within the flax cerebri; receives most of the blood drained from the superior surface of the cerebrum and leads to the inferior jugular vein and the vertebral vein
Great cerebral vein
Receives most of the smaller vessels from the inferior cerebral veins and leads to the straight sinus
Straight sinus
Enlarged vein that drains blood from the brain; receives most of the blood from the great cerebral vein and leads to the left or right transverse sinus
Cavernous sinus
Enlarged vein that receives blood from most of the other cerebral veins and the eye socket, and leads to the petrosal sinus
Petrosal sinus
Enlarged vein that receives blood from the cavernous sinus and leads into the internal jugular veins
Occipital sinus
Enlarged vein that drains the occipital region near the falx cerebelli and leads to the left and right transverse sinuses, and also the vertebral veins
Transverse sinuses
Pair of enlarged veins near the lambdoid suture that drains the occipital, sagittal, and straight sinuses, and leads to the sigmoid sinuses
Sigmoid sinuses
Enlarged vein that receives blood from the transverse sinuses and leads through the jugular foramen to the internal jugular vein
Veins Draining the Upper Limbs
The digital veins in the fingers come together in the hand to form the palmar venous arches (Figure 6.10.17). From here, the veins come together to form the radial vein, the ulnar vein, and the median antebrachial vein. The radial vein and the ulnar vein parallel the bones of the forearm and join together at the antebrachium to form the brachial vein, a deep vein that flows into the axillary vein in the brachium.
The median antebrachial vein parallels the ulnar vein, is more medial in location, and joins the basilic vein in the forearm. As the basilic vein reaches the antecubital region, it gives off a branch called the median cubital vein that crosses at an angle to join the cephalic vein. The median cubital vein is the most common site for drawing venous blood in humans. The basilic vein continues through the arm medially and superficially to the axillary vein.
The cephalic vein begins in the antebrachium and drains blood from the superficial surface of the arm into the axillary vein. It is extremely superficial and easily seen along the surface of the biceps brachii muscle in individuals with good muscle tone and in those without excessive subcutaneous adipose tissue in the arms.
The subscapular vein drains blood from the subscapular region and joins the cephalic vein to form the axillary vein. As it passes through the body wall and enters the thorax, the axillary vein becomes the subclavian vein.
Many of the larger veins of the thoracic and abdominal region and upper limb are further represented in the flow chart in Figure 6.10.18. Table 6.10.11 summarises the veins of the upper limbs.
Figure 6.10.17. Veins of the upper limb. This anterior view shows the veins that drain the upper limb.Figure 6.10.18. Veins flowing into the superior vena cava. The flow chart summarises the distribution of the veins flowing into the superior vena cava.
Table 6.10.11. Veins of the upper limbs
Vessel
Description
Digital veins
Drain the digits and lead to the palmar arches of the hand and dorsal venous arch of the foot
Palmar venous arches
Drain the hand and digits, and lead to the radial vein, ulnar veins, and the median antebrachial vein
Radial vein
Vein that parallels the radius and radial artery; arises from the palmar venous arches and leads to the branchial vein
Ulnar vein
Vein that parallels the ulna and ulnar artery; arises from the palmar venous arches and leads to the brachial vein
Brachial vein
Deeper vein of the arm that forms from the radial and ulnar veins in the lower arm; leads to the axillary vein
Median antebrachial vein
Vein that parallels the ulnar vein but is more medial in location; intertwines with the palmar venous arches; leads to the basilic vein
Basilic vein
Superficial vein of the arm that arises from the median antebrachial vein, intersects with the median cubital vein, parallels the ulnar vein, and continues into the upper arm; along with the brachial vein, it leads to the axillary vein
Median cubital vein
Superficial vessel located in the antecubital region that links the cephalic vein to the basilic vein in the form of a v; a frequent site from which to draw blood
Cephalic vein
Superficial vessel in the upper arm; leads to the axillary vein
Subscapular vein
Drains blood from the subscapular region and leads to the axillary vein
Axillary vein
The major vein in the axillary region; drains the upper limb and becomes the subclavian vein
The Inferior Vena Cava
Other than the small amount of blood drained by the azygos and hemiazygos veins, most of the blood inferior to the diaphragm drains into the inferior vena cava before it is returned to the heart (see Figure 6.10.15). Lying just beneath the parietal peritoneum in the abdominal cavity, the inferior vena cava parallels the abdominal aorta, where it can receive blood from abdominal veins. The lumbar portions of the abdominal wall and spinal cord are drained by a series of lumbar veins, usually four on each side. The ascending lumbar veins drain into either the azygos vein on the right or the hemiazygos vein on the left and return to the superior vena cava. The remaining lumbar veins drain directly into the inferior vena cava.
Blood supply from the kidneys flows into each renal vein, normally the largest veins entering the inferior vena cava. A number of other, smaller veins empty into the left renal vein. Each adrenal vein drains the adrenal or suprarenal glands located immediately superior to the kidneys. The right adrenal vein enters the inferior vena cava directly, whereas the left adrenal vein enters the left renal vein.
From the male reproductive organs, each testicular vein flows from the scrotum, forming a portion of the spermatic cord. Each ovarian vein drains an ovary in females. Each of these veins is generically called a gonadal vein. The right gonadal vein empties directly into the inferior vena cava, and the left gonadal vein empties into the left renal vein.
Each side of the diaphragm drains into a phrenic vein; the right phrenic vein empties directly into the inferior vena cava, whereas the left phrenic vein empties into the left renal vein. Blood supply from the liver drains into each hepatic vein and directly into the inferior vena cava. Since the inferior vena cava lies primarily to the right of the vertebral column and aorta, the left renal vein is longer, as are the left phrenic, adrenal, and gonadal veins. The longer length of the left renal vein makes the left kidney the primary target of surgeons removing this organ for donation. Figure 6.10.19 provides a flow chart of the veins flowing into the inferior vena cava. Table 6.10.12 summarises the major veins of the abdominal region.
Figure 6.10.19. Venous flow into inferior vena cava. The flow chart summarises veins that deliver blood to the inferior vena cava.
Table 6.10.12. Major veins of the abdominal region
Vessel
Description
Inferior vena cava
Large systemic vein that drains blood from areas largely inferior to the diaphragm; empties into the right atrium
Lumbar veins
Series of veins that drain the lumbar portion of the abdominal wall and spinal cord; the ascending lumbar veins drain into the azygos vein on the right or the hemiazygos vein on the left; the remaining lumbar veins drain directly into the inferior vena cava
Renal vein
Largest vein entering the inferior vena cava; drains the kidneys and flows into the interior vena cava
Adrenal vein
Drains the adrenal or suprarenal; the right adrenal vein enters the inferior vena cava directly and the left adrenal vein enters the left renal vein
Testicular vein
Drains the testes and forms part of the spermatic cord; the right testicular vein empties directly into the inferior vena cava and the left testicular vein empties into the left renal vein
Ovarian vein
Drains the ovary; the right ovarian vein empties directly into the inferior vena cava and the left ovarian vein empties into the left renal vein
Gonadal vein
Generic term for a vein draining a reproductive organ; may be either an ovarian vein or a testicular vein, depending on the sex of the individual
Phrenic vein
Drains the diaphragm; the right phrenic vein flows into the inferior vena cava and the left phrenic vein empties into the left renal vein
Hepatic vein
Drains systemic blood from the liver and flows into the inferior vena cava
Veins Draining the Lower Limbs
The superior surface of the foot drains into the digital veins, and the inferior surface drains into the plantar veins, which flow into a complex series of anastomoses in the feet and ankles, including the dorsal venous arch and the plantar venous arch (Figure 6.10.20). From the dorsal venous arch, blood supply drains into the anterior and posterior tibial veins. The anterior tibial vein drains the area near the tibialis anterior muscle and combines with the posterior tibial vein and the fibular vein to form the popliteal vein. The posterior tibial vein drains the posterior surface of the tibia and joins the popliteal vein. The fibular vein drains the muscles and integument in proximity to the fibula and also joins the popliteal vein. The small saphenous vein located on the lateral surface of the leg drains blood from the superficial regions of the lower leg and foot and flows into to the popliteal vein. As the popliteal vein passes behind the knee in the popliteal region, it becomes the femoral vein. It is palpable in patients without excessive adipose tissue.
Close to the body wall, the great saphenous vein, the deep femoral vein, and the femoral circumflex vein drain into the femoral vein. The great saphenous vein is a prominent surface vessel located on the medial surface of the leg and thigh that collects blood from the superficial portions of these areas. The deep femoral vein, as the name suggests, drains blood from the deeper portions of the thigh. The femoral circumflex vein forms a loop around the femur just inferior to the trochanters and drains blood from the areas in proximity to the head and neck of the femur.
As the femoral vein penetrates the body wall from the femoral portion of the upper limb, it becomes the external iliac vein, a large vein that drains blood from the leg to the common iliac vein. The pelvic organs and integument drain into the internal iliac vein, which forms from several smaller veins in the region, including the umbilical veins that run on either side of the bladder. The external and internal iliac veins combine near the inferior portion of the sacroiliac joint to form the common iliac vein. In addition to blood supply from the external and internal iliac veins, the middle sacral vein drains the sacral region into the common iliac vein. Similar to the common iliac arteries, the common iliac veins come together at the level of L5 to form the inferior vena cava.
Figure 6.10.21 is a flow chart of veins flowing into the lower limb. Table 6.10.13 summarises the major veins of the lower limbs.
Figure 6.10.20. Major veins serving the lower limbs. Anterior and posterior views show the major veins that drain the lower limb into the inferior vena cava.Figure 6.10.21. Major veins of the lower limb. The flow chart summarises venous flow from the lower limb.
Table 6.10.13. Veins of the lower limbs
Vessel
Description
Plantar veins
Drains the foot and flow into the plantar venous arch
Dorsal venous arch
Drains blood from digital veins and vessels on the superior surface of the foot
Plantar venous arch
Formed from the plantar veins; flows into the anterior and posterior tibial veins through anastomoses
Anterior tibial vein
Formed from the dorsal venous arch; drains the area near the tibialis anterior muscle and flows into the popliteal vein
Posterior tibial vein
Formed from the dorsal venous arch; drains the area near the posterior surface of the tibia and flows into the popliteal vein
Fibular vein
Drains the muscles and integument near the fibula and flows into the popliteal vein
Small saphenous vein
Located on the lateral surface of the leg; drains blood from the superficial regions of the lower leg and foot, and flows into the popliteal vein
Popliteal vein
Drains the region behind the knee and forms from the fusion of the fibular, anterior and posterior tibial veins; flows into the femoral vein
Great saphenous vein
Prominent surface vessel located on the medial surface of the leg and thigh; drains the superficial portions of these areas and flows into the femoral vein
Deep femoral vein
Drains blood from the deeper portions of the thigh and flows into the femoral vein
Femoral circumflex vein
Forms a loop around the femur just inferior to the trochanters; drains blood from the areas around the head and neck of the femur; flows into the femoral vein
External iliac vein
Formed when the femoral vein passes into the body cavity; drains the legs and flows into the common iliac vein
Middle sacral vein
Drains the sacral region and flows into the left common iliac vein
Common iliac vein
Flows into the inferior vena cava at the level of L5; the left common iliac vein drains the sacral region, formed from the union of external and internal iliac veins near the inferior portion of the sacroiliac joint
Hepatic Portal System
The liver is a complex biochemical processing plant. It packages nutrients absorbed by the digestive system; produces plasma proteins, clotting factors, and bile; and disposes of worn-out cell components and waste products. Instead of entering the circulation directly, absorbed nutrients and certain wastes (for example, materials produced by the spleen) travel to the liver for processing. They do so via the hepatic portal system (Figure 6.10.22). Portal systems begin and end in capillaries. In this case, the initial capillaries from the stomach, small intestine, large intestine, and spleen lead to the hepatic portal vein and end in specialised capillaries within the liver, the hepatic sinusoids. You saw the only other portal system with the hypothalamic-hypophyseal portal vessel in the endocrine chapter.
The hepatic portal system consists of the hepatic portal vein and the veins that drain into it. The hepatic portal vein itself is relatively short, beginning at the level of L2 with the confluence of the superior mesenteric and splenic veins. It also receives branches from the inferior mesenteric vein, plus the splenic veins and all their tributaries. The superior mesenteric vein receives blood from the small intestine, two-thirds of the large intestine, and the stomach. The inferior mesenteric vein drains the distal third of the large intestine, including the descending colon, the sigmoid colon, and the rectum. The splenic vein is formed from branches from the spleen, pancreas, and portions of the stomach, and the inferior mesenteric vein. After its formation, the hepatic portal vein also receives branches from the gastric veins of the stomach and cystic veins from the gall bladder. The hepatic portal vein delivers materials from these digestive and circulatory organs directly to the liver for processing.
Because of the hepatic portal system, the liver receives its blood supply from two different sources: from normal systemic circulation via the hepatic artery and from the hepatic portal vein. The liver processes the blood from the portal system to remove certain wastes and excess nutrients, which are stored for later use. This processed blood, as well as the systemic blood that came from the hepatic artery, exits the liver via the right, left, and middle hepatic veins, and flows into the inferior vena cava. Overall systemic blood composition remains relatively stable, since the liver is able to metabolise the absorbed digestive components.
Figure 6.10.22. Hepatic portal system. The liver receives blood from the normal systemic circulation via the hepatic artery. It also receives and processes blood from other organs, delivered via the veins of the hepatic portal system. All blood exits the liver via the hepatic vein, which delivers the blood to the inferior vena cava. (Different colours are used to help distinguish among the different vessels in the system.)
Section Review
The right ventricle pumps oxygen-depleted blood into the pulmonary trunk and right and left pulmonary arteries, which carry it to the right and left lungs for gas exchange. Oxygen-rich blood is transported by pulmonary veins to the left atrium. The left ventricle pumps this blood into the aorta. The main regions of the aorta are the ascending aorta, aortic arch, and descending aorta, which is further divided into the thoracic and abdominal aorta. The coronary arteries branch from the ascending aorta. After oxygenating tissues in the capillaries, systemic blood is returned to the right atrium from the venous system via the superior vena cava, which drains most of the veins superior to the diaphragm, the inferior vena cava, which drains most of the veins inferior to the diaphragm, and the coronary veins via the coronary sinus. The hepatic portal system carries blood to the liver for processing before it enters circulation. Review the figures provided in this section for circulation of blood through the blood vessels.
6.11 Development of Blood Vessels and Foetal Circulation
Learning Objectives
By the end of this section, you will be able to:
Describe the development of blood vessels
Describe the foetal circulation
In a developing embryo, the heart has developed enough by day 21 post-fertilisation to begin beating. Circulation patterns are clearly established by the fourth week of embryonic life. It is critical to the survival of the developing human that the circulatory system forms early to supply the growing tissue with nutrients and gases, and to remove waste products. Blood cells and vessel production in structures outside the embryo proper called the yolk sac, chorion, and connecting stalk begin about 15 to 16 days following fertilisation. Development of these circulatory elements within the embryo itself begins approximately 2 days later. You will learn more about the formation and function of these early structures when you study the chapter on development. During those first few weeks, blood vessels begin to form from the embryonic mesoderm. The precursor cells are known as haemangioblasts. These in turn differentiate into angioblasts, which give rise to the blood vessels and pluripotent stem cells, which differentiate into the formed elements of blood. (Seek additional content for more detail on foetal development and circulation.) Together, these cells form masses known as blood islands scattered throughout the embryonic disc. Spaces appear on the blood islands that develop into vessel lumens. The endothelial lining of the vessels arise from the angioblasts within these islands. Surrounding mesenchymal cells give rise to the smooth muscle and connective tissue layers of the vessels. While the vessels are developing, the pluripotent stem cells begin to form the blood.
Vascular tubes also develop on the blood islands, and they eventually connect to one another as well as to the developing, tubular heart. Thus, the developmental pattern, rather than beginning from the formation of one central vessel and spreading outward, occurs in many regions simultaneously with vessels later joining together. This angiogenesis—the creation of new blood vessels from existing ones—continues as needed throughout life as we grow and develop.
Blood vessel development often follows the same pattern as nerve development and travels to the same target tissues and organs. This occurs because the many factors directing growth of nerves also stimulate blood vessels to follow a similar pattern. Whether a given vessel develops into an artery or a vein is dependent upon local concentrations of signalling proteins.
As the embryo grows within the mother’s uterus, its requirements for nutrients and gas exchange also grow. The placenta—a circulatory organ unique to pregnancy—develops jointly from the embryo and uterine wall structures to fill this need. Emerging from the placenta is the umbilical vein, which carries oxygen-rich blood from the mother to the foetal inferior vena cava via the ductus venosus to the heart that pumps it into foetal circulation. Two umbilical arteries carry oxygen-depleted foetal blood, including wastes and carbon dioxide, to the placenta. Remnants of the umbilical arteries remain in the adult.
There are three major shunts—alternate paths for blood flow—found in the circulatory system of the foetus. Two of these shunts divert blood from the pulmonary to the systemic circuit, whereas the third connects the umbilical vein to the inferior vena cava. The first two shunts are critical during foetal life, when the lungs are compressed, filled with amniotic fluid, and non-functional, and gas exchange is provided by the placenta. These shunts close shortly after birth, however, when the newborn begins to breathe. The third shunt persists a bit longer but becomes non-functional once the umbilical cord is severed. The three shunts are as follows (Figure 6.11.1).
The foramen ovale is an opening in the interatrial septum that allows blood to flow from the right atrium to the left atrium. A valve associated with this opening prevents backflow of blood during the foetal period. As the newborn begins to breathe and blood pressure in the atria increases, this shunt closes. The fossa ovalis remains in the interatrial septum after birth, marking the location of the former foramen ovale.
The ductus arteriosus is a short, muscular vessel that connects the pulmonary trunk to the aorta. Most of the blood pumped from the right ventricle into the pulmonary trunk is thereby diverted into the aorta. Only enough blood reaches the foetal lungs to maintain the developing lung tissue. When the newborn takes the first breath, pressure within the lungs drops dramatically, and both the lungs and the pulmonary vessels expand. As the amount of oxygen increases, the smooth muscles in the wall of the ductus arteriosus constrict, sealing off the passage. Eventually, the muscular and endothelial components of the ductus arteriosus degenerate, leaving only the connective tissue component of the ligamentum arteriosum.
The ductus venosus is a temporary blood vessel that branches from the umbilical vein, allowing much of the freshly oxygenated blood from the placenta—the organ of gas exchange between the mother and foetus—to bypass the foetal liver and go directly to the foetal heart. The ductus venosus closes slowly during the first weeks of infancy and degenerates to become the ligamentum venosum.
Figure 6.11.1. Foetal shunts. The foramen ovale in the interatrial septum allows blood to flow from the right atrium to the left atrium. The ductus arteriosus is a temporary vessel, connecting the aorta to the pulmonary trunk. The ductus venosus links the umbilical vein to the inferior vena cava largely through the liver.
Section Review
Blood vessels begin to form from the embryonic mesoderm. The precursor haemangioblasts differentiate into angioblasts, which give rise to the blood vessels and pluripotent stem cells that differentiate into the formed elements of the blood. Together, these cells form blood islands scattered throughout the embryo. Extensions known as vascular tubes eventually connect the vascular network. As the embryo grows within the mother’s womb, the placenta develops to supply blood rich in oxygen and nutrients via the umbilical vein and to remove wastes in oxygen-depleted blood via the umbilical arteries. Three major shunts found in the foetus are the foramen ovale and ductus arteriosus, which divert blood from the pulmonary to the systemic circuit, and the ductus venosus, which carries freshly oxygenated blood high in nutrients to the foetal heart.
Describe the structure and function of the lymphatic tissue (lymph fluid, vessels, ducts, and organs)
Describe the structure and function of the primary and secondary lymphatic organs
Discuss the cells of the immune system, how they function, and their relationship with the lymphatic system
The immune system is the complex collection of cells and organs that destroys or neutralises pathogens that would otherwise cause disease or death. The lymphatic system, for most people, is associated with the immune system to such a degree that the two systems are virtually indistinguishable. The lymphatic system is the system of vessels, cells, and organs that carries excess fluids to the bloodstream and filters pathogens from the blood. The swelling of lymph nodes during an infection and the transport of lymphocytes via the lymphatic vessels are just two examples of the many connections between these critical organ systems.
Functions of the Lymphatic System
A major function of the lymphatic system is to drain body fluids and return them to the bloodstream. Blood pressure causes leakage of fluid from the capillaries, resulting in the accumulation of fluid in the interstitial space—that is, spaces between individual cells in the tissues. In humans, 20 litres of plasma is released into the interstitial space of the tissues each day due to capillary filtration. Once this filtrate is out of the bloodstream and in the tissue spaces, it is referred to as interstitial fluid. Of this, 17 litres is reabsorbed directly by the blood vessels. But what happens to the remaining three litres? This is where the lymphatic system comes into play. It drains the excess fluid and empties it back into the bloodstream via a series of vessels, trunks, and ducts. Lymph is the term used to describe interstitial fluid once it has entered the lymphatic system. When the lymphatic system is damaged in some way, such as by being blocked by cancer cells or destroyed by injury, protein-rich interstitial fluid accumulates (sometimes “backs up” from the lymph vessels) in the tissue spaces. This inappropriate accumulation of fluid referred to as lymphedema may lead to serious medical consequences.
As the vertebrate immune system evolved, the network of lymphatic vessels became convenient avenues for transporting the cells of the immune system. Additionally, the transport of dietary lipids and fat-soluble vitamins absorbed in the gut uses this system.
Cells of the immune system not only use lymphatic vessels to make their way from interstitial spaces back into the circulation, but they also use lymph nodes as major staging areas for the development of critical immune responses. A lymph node is one of the small, bean-shaped organs located throughout the lymphatic system.
Structure of the Lymphatic System
The lymphatic vessels begin as open-ended capillaries, which feed into larger and larger lymphatic vessels, and eventually empty into the bloodstream by a series of ducts. Along the way, the lymph travels through the lymph nodes, which are commonly found near the groin, armpits, neck, chest, and abdomen. Humans have about 500–600 lymph nodes throughout the body (Figure 7.1.1).
Figure 7.1.1. Anatomy of the lymphatic system. Lymphatic vessels in the arms and legs convey lymph to the larger lymphatic vessels in the torso.
A major distinction between the lymphatic and cardiovascular systems in humans is that lymph is not actively pumped by the heart, but is forced through the vessels by the movements of the body, the contraction of skeletal muscles during body movements, and breathing. One-way valves (semi-lunar valves) in lymphatic vessels keep the lymph moving toward the heart. Lymph flows from the lymphatic capillaries, through lymphatic vessels, and then is dumped into the circulatory system via the lymphatic ducts located at the junction of the jugular and subclavian veins in the neck.
Lymphatic Capillaries
Lymphatic capillaries, also called the terminal lymphatics, are vessels where interstitial fluid enters the lymphatic system to become lymph fluid. Located in almost every tissue in the body, these vessels are interlaced among the arterioles and venules of the circulatory system in the soft connective tissues of the body (Figure 7.1.2). Exceptions are the central nervous system, bone marrow, bones, teeth and the cornea of the eye, which do not contain lymph vessels.
Figure 7.1.2. Lymphatic capillaries. Lymphatic capillaries are interlaced with the arterioles and venules of the cardiovascular system. Collagen fibres anchor a lymphatic capillary in the tissue (inset). Interstitial fluid slips through spaces between the overlapping endothelial cells that compose the lymphatic capillary.
Lymphatic capillaries are formed by a one cell-thick layer of endothelial cells and represent the open end of the system, allowing interstitial fluid to flow into them via overlapping cells (see Figure 7.1.2). When interstitial pressure is low, the endothelial flaps close to prevent “backflow.” As interstitial pressure increases, the spaces between the cells open up, allowing the fluid to enter. Entry of fluid into lymphatic capillaries is also enabled by the collagen filaments that anchor the capillaries to surrounding structures. As interstitial pressure increases, the filaments pull on the endothelial cell flaps, opening up them even further to allow easy entry of fluid.
In the small intestine, lymphatic capillaries called lacteals are critical for the transport of dietary lipids and lipid-soluble vitamins to the bloodstream. In the small intestine, dietary triglycerides combine with other lipids and proteins, and enter the lacteals to form a milky fluid called chyle. The chyle then travels through the lymphatic system, eventually entering the liver and then the bloodstream.
Larger Lymphatic Vessels, Trunks and Ducts
The lymphatic capillaries empty into larger lymphatic vessels, which are similar to veins in terms of their three-tunic structure and the presence of valves. These one-way valves are located fairly close to one another, and each one causes a bulge in the lymphatic vessel, giving the vessels a beaded appearance (see Figure 7.1.2).
The superficial and deep lymphatics eventually merge to form larger lymphatic vessels known as lymphatic trunks. On the right side of the body, the right sides of the head, thorax, and right upper limb drain lymph fluid into the right subclavian vein via the right lymphatic duct (Figure 7.1.3). On the left side of the body, the remaining portions of the body drain into the larger thoracic duct, which drains into the left subclavian vein. The thoracic duct itself begins just beneath the diaphragm in the cisterna chyli, a sac-like chamber that receives lymph from the lower abdomen, pelvis, and lower limbs by way of the left and right lumbar trunks and the intestinal trunk.
Figure 7.1.3. Major trunks and ducts of the lymphatic system. The thoracic duct drains a much larger portion of the body than does the right lymphatic duct.
The overall drainage system of the body is asymmetrical (see Figure 7.1.3). The right lymphatic duct receives lymph from only the upper right side of the body. The lymph from the rest of the body enters the bloodstream through the thoracic duct via all the remaining lymphatic trunks. In general, lymphatic vessels of the subcutaneous tissues of the skin, that is, the superficial lymphatics, follow the same routes as veins, whereas the deep lymphatic vessels of the viscera generally follow the paths of arteries.
The Organisation of Immune Function
The immune system is a collection of barriers, cells, and soluble proteins that interact and communicate with each other in extraordinarily complex ways. The modern model of immune function is organised into three phases based on the timing of their effects. The three temporal phases consist of the following:
Barrier defences such as the skin and mucous membranes, which act instantaneously to prevent pathogenic invasion into the body tissues
The rapid but nonspecific innate immune response, which consists of a variety of specialised cells and soluble factors
The slower but more specific and effective adaptive immune response, which involves many cell types and soluble factors, but is primarily controlled by white blood cells (leukocytes) known as lymphocytes, which help control immune responses
The cells of the blood, including all those involved in the immune response, arise in the bone marrow via various differentiation pathways from hematopoietic stem cells (Figure 7.1.4). In contrast with embryonic stem cells, hematopoietic stem cells are present throughout adulthood and allow for the continuous differentiation of blood cells to replace those lost to age or function. These cells can be divided into three classes based on function:
Phagocytic cells, which ingest pathogens to destroy them
Lymphocytes, which specifically coordinate the activities of adaptive immunity
Cells containing cytoplasmic granules, which help mediate immune responses against parasites and intracellular pathogens such as viruses
Figure 7.1.4. Haematopoietic system of the bone marrow. All the cells of the immune response as well as of the blood arise by differentiation from haematopoietic stem cells. Platelets are cell fragments involved in the clotting of blood.
Lymphocytes: B Cells, T cells, Plasma Cells and Natural Killer Cells
As stated above, lymphocytes are the primary cells of adaptive immune responses (Table 7.1.1). The two basic types of lymphocytes, B cells and T cells, are identical morphologically with a large central nucleus surrounded by a thin layer of cytoplasm. They are distinguished from each other by their surface protein markers as well as by the molecules they secrete. While B cells mature in red bone marrow and T cells mature in the thymus, they both initially develop from bone marrow. T cells migrate from bone marrow to the thymus gland where they further mature. B cells and T cells are found in many parts of the body, circulating in the bloodstream and lymph, and residing in secondary lymphoid organs, including the spleen and lymph nodes, which will be described later in this section. The human body contains approximately 2×1012 lymphocytes.
B Cells
B cells are immune cells that function primarily by producing antibodies. An antibody is any of the group of proteins that binds specifically to pathogen-associated molecules known as antigens. An antigen is a chemical structure on the surface of a pathogen that binds to T or B lymphocyte antigen receptors. Once activated by binding to antigen, B cells differentiate into cells that secrete a soluble form of their surface antibodies. These activated B cells are known as plasma cells.
T Cells
The T cell, on the other hand, does not secrete antibody but performs a variety of functions in the adaptive immune response. Different T cell types have the ability to either secrete soluble factors that communicate with other cells of the adaptive immune response or destroy cells infected with intracellular pathogens. The roles of T and B lymphocytes in the adaptive immune response will be discussed further in this chapter.
Plasma Cells
Another type of lymphocyte of importance is the plasma cell. A plasma cell is a B cell that has differentiated in response to antigen binding and has thereby gained the ability to secrete soluble antibodies. These cells differ in morphology from standard B and T cells in that they contain a large amount of cytoplasm packed with the protein-synthesising machinery known as rough endoplasmic reticulum.
Natural Killer (NK) Cells
A fourth important lymphocyte is the natural killer cell, a participant in the innate immune response. A natural killer cell (NK) is a circulating blood cell that contains cytotoxic (cell-killing) granules in its extensive cytoplasm. It shares this mechanism with the cytotoxic T cells of the adaptive immune response. NK cells are among the body’s first lines of defence against viruses and certain types of cancer.
Table 7.1.1. Lymphocytes
Type of Lymphocyte
Primary Function
B lymphocyte
Generates diverse antibodies
T lymphocyte
Secretes chemical messengers
Plasma cell
Secretes antibodies
NK cell
Destroys virally infected cells
Primary Lymphoid Organs and Lymphocyte Development
Understanding the differentiation and development of B and T cells is critical to the understanding of the adaptive immune response. It is through this process that the body (ideally) learns to destroy only pathogens and leaves the body’s own cells relatively intact. The primary lymphoid organs are the bone marrow and thymus gland. The lymphoid organs are where lymphocytes mature, proliferate, and are selected, which enables them to attack pathogens without harming the cells of the body.
Bone Marrow
Figure 7.1.5. Bone marrow. Red bone marrow fills the head of the femur, and a spot of yellow bone marrow is visible in the centre. The white reference bar is 1 cm.
In the embryo, blood cells are made in the yolk sac. As development proceeds, this function is taken over by the spleen, lymph nodes, and liver. Later, the bone marrow takes over most haematopoietic functions, although the final stages of the differentiation of some cells may take place in other organs. The red bone marrow is a loose collection of cells where haematopoiesis occurs, and the yellow bone marrow is a site of energy storage, which consists largely of fat cells (Figure 7.1.5). The B cell undergoes nearly all of its development in the red bone marrow, whereas the immature T cell, called a thymocyte, leaves the bone marrow and matures largely in the thymus gland.
Thymus
The thymus gland is a bilobed organ found in the space between the sternum and the aorta of the heart (Figure 7.1.6). Connective tissue holds the lobes closely together but also separates them and forms a capsule.
The connective tissue capsule further divides the thymus into lobules via extensions called trabeculae. The outer region of the organ is known as the cortex and contains large numbers of thymocytes with some epithelial cells, macrophages, and dendritic cells (two types of phagocytic cells that are derived from monocytes). The cortex is densely packed so it stains more intensely than the rest of the thymus (see Figure 7.1.6). The medulla, where thymocytes migrate before leaving the thymus, contains a less dense collection of thymocytes, epithelial cells, and dendritic cells.
Ageing and the Immune System
By the year 2050, the number of adults between 65 and 85 years is estimated to double, while number of adults aged above 85 is estimated to quadruple. Although the aging population shows the greatest burden of chronic conditions related to immune deficiencies, younger population is also affected. This loss of immune function with age is called immunosenescence. To treat this growing population, medical professionals must better understand the ageing process. One major cause of age-related immune deficiencies is thymic involution, the shrinking of the thymus gland that begins at birth, at a rate of about three percent tissue loss per year, and continues until 35–45 years of age, when the rate declines to about one percent loss per year for the rest of one’s life. At that pace, the total loss of thymic epithelial tissue and thymocytes would occur at about 120 years of age. Thus, this age is a theoretical limit to a healthy human lifespan.
Thymic involution has been observed in all vertebrate species that have a thymus gland. Animal studies have shown that transplanted thymic grafts between inbred strains of mice involuted according to the age of the donor and not of the recipient, implying the process is genetically programmed. There is evidence that the thymic microenvironment, so vital to the development of naïve T cells, loses thymic epithelial cells according to the decreasing expression of the FOXN1 gene with age.
It is also known that thymic involution can be altered by hormone levels. Sex hormones such as estrogen and testosterone enhance involution, and the hormonal changes in pregnant women cause a temporary thymic involution that reverses itself, when the size of the thymus and its hormone levels return to normal, usually after lactation ceases. What does all this tell us? Can we reverse immunosenescence, or at least slow it down? The potential is there for using thymic transplants from younger donors to keep thymic output of naïve T cells high. Gene therapies that target gene expression are also seen as future possibilities. The more we learn through immunosenescence research, the more opportunities there will be to develop therapies, even though these therapies will likely take decades to develop. The ultimate goal is for everyone to live and be healthy longer, but there may be limits to immortality imposed by our genes and hormones.
Secondary Lymphoid Organs and Their Roles in Active Immune Responses
Lymphocytes develop and mature in the primary lymphoid organs, but they mount immune responses from the secondary lymphoid organs. A naïve lymphocyte is one that has left the primary organ and entered a secondary lymphoid organ. Naïve lymphocytes are fully functional immunologically but have yet to encounter an antigen to respond to. In addition to circulating in the blood and lymph, lymphocytes concentrate in secondary lymphoid organs, which include the lymph nodes, spleen, and lymphoid nodules. All of these tissues have many features in common, including the following:
The presence of lymphoid follicles, the sites of the formation of lymphocytes, with specific B cell-rich and T cell-rich areas
An internal structure of reticular fibres with associated fixed macrophages
Germinal centres, which are the sites of rapidly dividing B lymphocytes and plasma cells, with the exception of the spleen
Specialised post-capillary vessels known as high endothelial venules; the cells lining these venules are thicker and more columnar than normal endothelial cells, which allow cells from the blood to directly enter these tissues
Lymph Nodes
Lymph nodes function to remove debris and pathogens from the lymph, and are thus sometimes referred to as the “filters of the lymph” (Figure 7.1.7). Any bacteria that infect the interstitial fluid are taken up by the lymphatic capillaries and transported to a regional lymph node. Dendritic cells and macrophages within this organ internalise and kill many of the pathogens that pass through, thereby removing them from the body. The lymph node is also the site of adaptive immune responses mediated by T cells, B cells, and accessory cells of the adaptive immune system. Like the thymus, the bean-shaped lymph nodes are surrounded by a tough capsule of connective tissue and are separated into compartments by trabeculae, the extensions of the capsule. In addition to the structure provided by the capsule and trabeculae, the structural support of the lymph node is provided by a series of reticular fibres laid down by fibroblasts.
The major routes into the lymph node are via afferent lymphatic vessels. Cells and lymph fluid that leave the lymph node may do so by another set of vessels known as the efferent lymphatic vessels. Lymph enters the lymph node via the subcapsular sinus, which is occupied by dendritic cells, macrophages, and reticular fibres. Within the cortex of the lymph node are lymphoid follicles, which consist of germinal centres of rapidly dividing B cells surrounded by a layer of T cells and other accessory cells. As the lymph continues to flow through the node, it enters the medulla, which consists of medullary cords of B cells and plasma cells, and the medullary sinuses where the lymph collects before leaving the node via the efferent lymphatic vessels.
Spleen
In addition to the lymph nodes, the spleen is a major secondary lymphoid organ (Figure 7.1.8). It is about 12 cm (5 in) long and is attached to the lateral border of the stomach via the gastrosplenic ligament. The spleen is a fragile organ without a strong capsule and is dark red due to its extensive vascularisation. The spleen is sometimes called the “filter of the blood” because of its extensive vascularisation and the presence of macrophages and dendritic cells that remove microbes and other materials from the blood, including dying red blood cells. The spleen also functions as the location of immune responses to blood-borne pathogens.
The spleen is also divided by trabeculae of connective tissue, and within each splenic nodule is an area of red pulp, consisting of mostly red blood cells, and white pulp, which resembles the lymphoid follicles of the lymph nodes. Upon entering the spleen, the splenic artery splits into several arterioles (surrounded by white pulp) and eventually into sinusoids. Blood from the capillaries subsequently collects in the venous sinuses and leaves via the splenic vein. The red pulp consists of reticular fibres with fixed macrophages attached, free macrophages, and all of the other cells typical of the blood, including some lymphocytes. The white pulp surrounds a central arteriole and consists of germinal centres of dividing B cells surrounded by T cells and accessory cells, including macrophages and dendritic cells. Thus, the red pulp primarily functions as a filtration system of the blood, using cells of the relatively nonspecific immune response, and white pulp is where adaptive T and B cell responses are mounted.
Lymphoid Nodules
The other lymphoid tissues, the lymphoid nodules, have a simpler architecture than the spleen and lymph nodes in that they consist of a dense cluster of lymphocytes without a surrounding fibrous capsule. These nodules are located in the respiratory and digestive tracts, areas routinely exposed to environmental pathogens.
Tonsils are lymphoid nodules located along the inner surface of the pharynx and are important in developing immunity to oral pathogens (Figure 7.1.9). The tonsil located at the back of the throat, the pharyngeal tonsil, is sometimes referred to as the adenoid when swollen. Such swelling is an indication of an active immune response to infection. Histologically, tonsils do not contain a complete capsule, and the epithelial layer invaginates deeply into the interior of the tonsil to form tonsillar crypts. These structures, which accumulate all sorts of materials taken into the body through eating and breathing, actually “encourage” pathogens to penetrate deep into the tonsillar tissues where they are acted upon by numerous lymphoid follicles and eliminated. This seems to be the major function of tonsils—to help children’s bodies recognise, destroy, and develop immunity to common environmental pathogens so that they will be protected in their later lives. Tonsils are often removed in those children who have recurring throat infections, especially those involving the palatine tonsils on either side of the throat, whose swelling may interfere with their breathing and/or swallowing.
Mucosa-associated lymphoid tissue (MALT) consists of an aggregate of lymphoid follicles directly associated with the mucous membrane epithelia. MALT makes up dome-shaped structures found underlying the mucosa of the gastrointestinal tract, breast tissue, lungs, and eyes. Peyer’s patches, a type of MALT in the small intestine, are especially important for immune responses against ingested substances (Figure 7.1.10). Peyer’s patches contain specialised endothelial cells called M (or microfold) cells that sample material from the intestinal lumen and transport it to nearby follicles so that adaptive immune responses to potential pathogens can be mounted.
Bronchus-associated lymphoid tissue (BALT) consists of lymphoid follicular structures with an overlying epithelial layer found along the bifurcations of the bronchi, and between bronchi and arteries. They also have the typically less-organised structure of other lymphoid nodules. These tissues, in addition to the tonsils, are effective against inhaled pathogens.
Section Review
The lymphatic system is a series of vessels, ducts, and trunks that remove interstitial fluid from the tissues and return it the blood. The lymphatics are also used to transport dietary lipids and cells of the immune system. Cells of the immune system all come from the hematopoietic system of the bone marrow. Primary lymphoid organs, the bone marrow and thymus gland, are the locations where lymphocytes of the adaptive immune system proliferate and mature. Secondary lymphoid organs are site in which mature lymphocytes congregate to mount immune responses. Many immune system cells use the lymphatic and circulatory systems for transport throughout the body to search for and then protect against pathogens.
7.2 Barrier Defences and the Innate Immune Response
Learning Objectives
By the end of this section, you will be able to:
Describe the barrier defences of the body
Show how the innate immune response is important and how it helps guide and prepare the body for adaptive immune responses
Describe various soluble factors that are part of the innate immune response
Explain the steps of inflammation and how they lead to destruction of a pathogen
Discuss early induced immune responses and their level of effectiveness
The immune system can be divided into two overlapping mechanisms to destroy pathogens: the innate immune response, which is relatively rapid but nonspecific and thus not always effective, and the adaptive immune response, which is slower in its development during an initial infection with a pathogen, but is highly specific and effective at attacking a wide variety of pathogens (Figure 7.2.1).
Figure 7.2.1. Cooperation between innate and adaptive immune responses. The innate immune system enhances adaptive immune responses so they can be more effective.
Any discussion of the innate immune response usually begins with the physical barriers that prevent pathogens from entering the body, destroy them after they enter, or flush them out before they can establish themselves in the hospitable environment of the body’s soft tissues. Barrier defences are part of the body’s most basic defence mechanisms. The barrier defences are not a response to infections, but they are continuously working to protect against a broad range of pathogens.
The different modes of barrier defences are associated with the external surfaces of the body, where pathogens may try to enter (Table 7.2.1). The primary barrier to the entrance of microorganisms into the body is the skin. Not only is the skin covered with a layer of dead, keratinised epithelium that is too dry for bacteria in which to grow, but as these cells are continuously sloughed off from the skin, they carry bacteria and other pathogens with them. Additionally, sweat and other skin secretions may lower pH, contain toxic lipids, and physically wash microbes away.
Table 7.2.1. Barrier defences
Site
Specific Defence
Protective Aspect
Skin
Epidermal surface
Keratinised cells of surface, Langerhans cells
Skin
(sweat/secretions)
Sweat glands, sebaceous glands
Low pH, washing action
Oral cavity
Salivary glands
Lysosome
Stomach
Gastrointestinal tract
Low pH
Mucosal surfaces
Mucosal epithelium
Nonkeratinised epithelial cells
Normal microbiome
(non-pathogenic bacteria)
Mucosal tissue
Prevent pathogens from growing on mucosal surfaces
Another barrier is the saliva in the mouth, which is rich in lysozyme—an enzyme that destroys bacteria by digesting their cell walls. The acidic environment of the stomach, which is fatal to many pathogens, is also a barrier. Additionally, the mucus layer of the gastrointestinal tract, respiratory tract, reproductive tract, eyes, ears, and nose traps both microbes and debris, and facilitates their removal. In the case of the upper respiratory tract, ciliated epithelial cells move potentially contaminated mucus upwards to the mouth, where it is then swallowed into the digestive tract, ending up in the harsh acidic environment of the stomach. Considering how often you breathe compared to how often you eat or perform other activities that expose you to pathogens, it is not surprising that multiple barrier mechanisms have evolved to work in concert to protect this vital area.
Cells of the Innate Immune Response
A phagocyte is a cell that is able to surround and engulf a particle or cell, a process called phagocytosis. The phagocytes of the immune system engulf other particles or cells, either to clean an area of debris, old cells, or to kill pathogenic organisms such as bacteria. The phagocytes are the body’s fast acting, first line of immunological defence against organisms that have breached barrier defences and have entered the vulnerable tissues of the body.
Phagocytes: Macrophages and Neutrophils
Many of the cells of the immune system have a phagocytic ability, at least at some point during their life cycles. Phagocytosis is an important and effective mechanism of destroying pathogens during innate immune responses. The phagocyte takes the organism inside itself as a phagosome, which subsequently fuses with a lysosome and its digestive enzymes, effectively killing many pathogens. On the other hand, some bacteria including Mycobacteria tuberculosis, the cause of tuberculosis, may be resistant to these enzymes and are therefore much more difficult to clear from the body. Macrophages, neutrophils, and dendritic cells are the major phagocytes of the immune system.
A macrophage is an irregularly shaped phagocyte that is amoeboid in nature and is the most versatile of the phagocytes in the body. Macrophages move through tissues and squeeze through capillary walls using pseudopodia. They not only participate in innate immune responses but have also evolved to cooperate with lymphocytes as part of the adaptive immune response. Macrophages exist in many tissues of the body, either freely roaming through connective tissues or fixed to reticular fibres within specific tissues such as lymph nodes. When pathogens breach the body’s barrier defences, macrophages are the first line of defence (Table 7.2.2). They are called different names, depending on the tissue: Kupffer cells in the liver, histiocytes in connective tissue, and alveolar macrophages in the lungs.
A neutrophil is a phagocytic cell that is attracted via chemotaxis from the bloodstream to infected tissues. These spherical cells are granulocytes. A granulocyte contains cytoplasmic granules, which in turn contain a variety of vasoactive mediators such as histamine. In contrast, macrophages are agranulocytes. An agranulocyte has few or no cytoplasmic granules. Whereas macrophages act like sentries, always on guard against infection, neutrophils can be thought of as military reinforcements that are called into a battle to hasten the destruction of the enemy. Although, usually thought of as the primary pathogen-killing cell of the inflammatory process of the innate immune response, new research has suggested that neutrophils play a role in the adaptive immune response as well, just as macrophages do.
A monocyte is a circulating precursor cell that differentiates into either a macrophage or dendritic cell, which can be rapidly attracted to areas of infection by signal molecules of inflammation.
Table 7.2.2. Phagocytic cells of the innate immune system
Cell
Cell type
Primary location
Function in the innate immune response
Macrophage
Agranulocyte
Body cavities/organs
Phagocytosis
Neutrophil
Granulocyte
Blood
Phagocytosis
Monocyte
Agranulocyte
Blood
Precursor of macrophage/dendritic cell
Natural Killer Cells
NK cells are a type of lymphocyte that have the ability to induce apoptosis, that is, programmed cell death, in cells infected with intracellular pathogens such as obligate intracellular bacteria and viruses. NK cells recognise these cells by mechanisms that are still not well understood, but that presumably involve their surface receptors. NK cells can induce apoptosis, in which a cascade of events inside the cell causes its own death by either of two mechanisms:
NK cells are able to respond to chemical signals and express the fas ligand. The fas ligand is a surface molecule that binds to the fas molecule on the surface of the infected cell, sending it apoptotic signals, thus killing the cell and the pathogen within it; or
The granules of the NK cells release perforins and granzymes. A perforin is a protein that forms pores in the membranes of infected cells. A granzyme is a protein-digesting enzyme that enters the cell via the perforin pores and triggers apoptosis intracellularly.
Both mechanisms are especially effective against virally infected cells. If apoptosis is induced before the virus has the ability to synthesise and assemble all its components, no infectious virus will be released from the cell, thus preventing further infection.
Recognition of Pathogens
Cells of the innate immune response, the phagocytic cells, and the cytotoxic NK cells recognise patterns of pathogen-specific molecules, such as bacterial cell wall components or bacterial flagellar proteins, using pattern recognition receptors. A pattern recognition receptor (PRR) is a membrane-bound receptor that recognises characteristic features of a pathogen and molecules released by stressed or damaged cells.
These receptors, which are thought to have evolved prior to the adaptive immune response, are present on the cell surface whether they are needed or not. Their variety, however, is limited by two factors. First, the fact that each receptor type must be encoded by a specific gene requires the cell to allocate most or all its DNA to make receptors able to recognise all pathogens. Secondly, the variety of receptors is limited by the finite surface area of the cell membrane. Thus, the innate immune system must “get by” using only a limited number of receptors that are active against as wide a variety of pathogens as possible. This strategy is in stark contrast to the approach used by the adaptive immune system, which uses large numbers of different receptors, each highly specific to a particular pathogen.
Should the cells of the innate immune system come into contact with a species of pathogen they recognise, the cell will bind to the pathogen and initiate phagocytosis (or cellular apoptosis in the case of an intracellular pathogen) in an effort to destroy the offending microbe. Receptors vary somewhat according to cell type, but they usually include receptors for bacterial components and for complement, discussed below.
Soluble Mediators of the Innate Immune Response
The previous discussions have alluded to chemical signals that can induce cells to change various physiological characteristics, such as the expression of a particular receptor. These soluble factors are secreted during innate or early induced responses, and later during adaptive immune responses.
Cytokines and Chemokines
A cytokine is signalling molecule that allows cells to communicate with each other over short distances. Cytokines are secreted into the intercellular space, and the action of the cytokine induces the receiving cell to change its physiology. A chemokine is a soluble chemical mediator similar to cytokines except that its function is to attract cells (chemotaxis) from longer distances.
Early Induced Proteins
Early induced proteins are those that are not constitutively present in the body, but are made as they are needed early during the innate immune response. Interferons are an example of early induced proteins. Cells infected with viruses secrete interferons that travel to adjacent cells and induce them to make antiviral proteins. Thus, even though the initial cell is sacrificed, the surrounding cells are protected. Other early induced proteins specific for bacterial cell wall components are mannose-binding protein and C-reactive protein, made in the liver, which bind specifically to polysaccharide components of the bacterial cell wall. Phagocytes such as macrophages have receptors for these proteins, and they are thus able to recognise them as they are bound to the bacteria. This brings the phagocyte and bacterium into close proximity and enhances the phagocytosis of the bacterium by the process known as opsonisation. Opsonisation is the tagging of a pathogen for phagocytosis by the binding of an antibody or an antimicrobial protein.
Complement System
The complement system is a series of proteins constitutively found in the blood plasma. As such, these proteins are not considered part of the early induced immune response, even though they share features with some of the antibacterial proteins of this class. Made in the liver, they have a variety of functions in the innate immune response, using what is known as the “alternate pathway” of complement activation. Additionally, complement functions in the adaptive immune response as well, in what is called the classical pathway. The complement system consists of several proteins that enzymatically alter and fragment later proteins in a series, which is why it is termed cascade. Once activated, the series of reactions is irreversible, and releases fragments that have the following actions:
Bind to the cell membrane of the pathogen that activates it, labelling it for phagocytosis (opsonisation)
Diffuse away from the pathogen and act as chemotactic agents to attract phagocytic cells to the site of inflammation
Form damaging pores in the plasma membrane of the pathogen
Figure 7.2.2 shows the classical pathway, which requires antibodies of the adaptive immune response. The alternate pathway does not require an antibody to become activated.
Figure 7.2.2. Complement cascade and function. The classical pathway, used during adaptive immune responses, occurs when C1 reacts with antibodies that have bound an antigen.
The splitting of the C3 protein is the common step to both pathways. In the alternate pathway, C3 is activated spontaneously and, after reacting with the molecules factor P, factor B, and factor D, splits apart. The larger fragment, C3b, binds to the surface of the pathogen and C3a, the smaller fragment, diffuses outward from the site of activation and attracts phagocytes to the site of infection. Surface-bound C3b then activates the rest of the cascade, with the last five proteins, C5–C9, forming the membrane-attack complex (MAC). The MAC can kill certain pathogens by disrupting their osmotic balance. The MAC is especially effective against a broad range of bacteria. The classical pathway is similar, except the early stages of activation require the presence of antibody bound to antigen, and thus is dependent on the adaptive immune response. The earlier fragments of the cascade also have important functions. Phagocytic cells such as macrophages and neutrophils are attracted to an infection site by chemotactic attraction to smaller complement fragments. Additionally, once they arrive, their receptors for surface-bound C3b opsonise the pathogen for phagocytosis and destruction.
Inflammatory Response
The hallmark of the innate immune response is inflammation. Inflammation is something everyone has experienced. Stub a toe, cut a finger, or do any activity that causes tissue damage and inflammation will result, with its four characteristics: heat, redness, pain, and swelling (“loss of function” is sometimes mentioned as a fifth characteristic). It is important to note that inflammation does not have to be initiated by an infection but can also be caused by tissue injuries. The release of damaged cellular contents into the site of injury is enough to stimulate the response, even in the absence of breaks in physical barriers that would allow pathogens to enter (by hitting your thumb with a hammer, for example). The inflammatory reaction brings in phagocytic cells to the damaged area to clear cellular debris and to set the stage for wound repair (Figure 7.2.3).
Figure 7.2.3. Inflammatory response.
This reaction also brings in the cells of the innate immune system, allowing them to get rid of the sources of a possible infection. Inflammation is part of a very basic form of immune response. The process not only brings fluid and cells into the site to destroy the pathogen and remove it and debris from the site, but also helps to isolate the site, limiting the spread of the pathogen. Acute inflammation is a short-term inflammatory response to an insult to the body. If the cause of the inflammation is not resolved, however, it can lead to chronic inflammation, which is associated with major tissue destruction and fibrosis. Chronic inflammation is ongoing inflammation. It can be caused by foreign bodies, persistent pathogens, and autoimmune diseases such as rheumatoid arthritis.
There are four important parts to the inflammatory response:
Tissue Injury. The released contents of injured cells stimulate the release of mast cell granules and their potent inflammatory mediators such as histamine, leukotrienes, and prostaglandins. Histamine increases the diameter of local blood vessels (vasodilation), causing an increase in blood flow. Histamine also increases the permeability of local capillaries, causing plasma to leak out and form interstitial fluid. This causes the swelling associated with inflammation.
Additionally, injured cells, phagocytes, and basophils are sources of inflammatory mediators, including prostaglandins and leukotrienes. Leukotrienes attract neutrophils from the blood by chemotaxis and increase vascular permeability. Prostaglandins cause vasodilation by relaxing vascular smooth muscle and are a major cause of the pain associated with inflammation. Nonsteroidal anti-inflammatory drugs such as aspirin and ibuprofen relieve pain by inhibiting prostaglandin production.
Vasodilation. Many inflammatory mediators such as histamine are vasodilators that increase the diameters of local capillaries. This causes increased blood flow and is responsible for the heat and redness of inflamed tissue. It allows greater access of the blood to the site of inflammation.
Increased Vascular Permeability. At the same time, inflammatory mediators increase the permeability of the local vasculature, causing leakage of fluid into the interstitial space, resulting in the swelling, or oedema, associated with inflammation.
Recruitment of Phagocytes. Leukotrienes are particularly good at attracting neutrophils from the blood to the site of infection by chemotaxis. Following an early neutrophil infiltrate stimulated by macrophage cytokines, more macrophages are recruited to clean up the debris left over at the site. When local infections are severe, neutrophils are attracted to the sites of infections in large numbers, and as they phagocytose the pathogens and subsequently die, their accumulated cellular remains are visible as pus at the infection site.
Overall, inflammation is valuable for many reasons. Not only are the pathogens killed and debris removed, but the increase in vascular permeability encourages the entry of clotting factors, the first step towards wound repair. Inflammation also facilitates the transport of antigen to lymph nodes by dendritic cells for the development of the adaptive immune response.
Section Review
Innate immune responses are critical to the early control of infections. Whereas barrier defences are the body’s first line of physical defence against pathogens, innate immune responses are the first line of physiological defence. Innate responses occur rapidly, but with less specificity and effectiveness than the adaptive immune response. Innate responses can be caused by a variety of cells, mediators, and antibacterial proteins such as complement. Within the first few days of an infection, another series of antibacterial proteins are induced, each with activities against certain bacteria, including opsonisation of certain species. Additionally, interferons are induced that protect cells from viruses in their vicinity. Finally, the innate immune response does not stop when the adaptive immune response is developed. In fact, both can cooperate and one can influence the other in their responses against pathogens.
7.3 The Adaptive Immune Response: T Lymphocytes and their Functional Types
Learning Objectives
By the end of this section, you will be able to:
Explain the advantages of the adaptive immune response over the innate immune response
List the various characteristics of an antigen
Describe the types of T cell antigen receptors
Outline the steps of T cell development
Describe the major T cell types and their functions
Innate immune responses (and early induced responses) are in many cases ineffective at completely controlling pathogen growth. However, they slow pathogen growth and allow time for the adaptive immune response to strengthen and either control or eliminate the pathogen. The innate immune system also sends signals to the cells of the adaptive immune system, guiding them in how to attack the pathogen. Thus, these are the two important arms of the immune response.
The Benefits of the Adaptive Immune Response
The specificity of the adaptive immune response—its ability to specifically recognise and make a response against a wide variety of pathogens—is its great strength. Antigens, the small chemical groups often associated with pathogens, are recognised by receptors on the surface of B and T lymphocytes. The adaptive immune response to these antigens is so versatile that it can respond to nearly any pathogen. This increase in specificity comes because the adaptive immune response has a unique way to develop as many as 1011, or 100 trillion, different receptors to recognise nearly every conceivable pathogen. How could so many different types of antibodies be encoded? And what about the many specificities of T cells? There is not nearly enough DNA in a cell to have a separate gene for each specificity. The mechanism was finally worked out in the 1970s and 1980s using the new tools of molecular genetics
Primary Disease and Immunological Memory
The immune system’s first exposure to a pathogen is called a primary adaptive response. Symptoms of a first infection, called primary disease, are always relatively severe because it takes time for an initial adaptive immune response to a pathogen to become effective.
Upon re-exposure to the same pathogen, a secondary adaptive immune response is generated, which is stronger and faster that the primary response. The secondary adaptive response often eliminates a pathogen before it can cause significant tissue damage or any symptoms. Without symptoms, there is no disease, and the individual is not even aware of the infection. This secondary response is the basis of immunological memory, which protects us from getting diseases repeatedly from the same pathogen. By this mechanism, an individual’s exposure to pathogens early in life spares the person from these diseases later in life.
Self Recognition
A third important feature of the adaptive immune response is its ability to distinguish between self-antigens, those that are normally present in the body, and foreign antigens, those that might be on a potential pathogen. As T and B cells mature, there are mechanisms in place that prevent them from recognising self-antigen, preventing a damaging immune response against the body. These mechanisms are not 100 percent effective, however, and their breakdown leads to autoimmune diseases, which will be discussed later in this section.
T Cell-Mediated Immune Responses
The primary cells that control the adaptive immune response are the lymphocytes, the T and B cells. T cells are particularly important, as they not only control a multitude of immune responses directly, but also control B cell immune responses in many cases as well. Thus, many of the decisions about how to attack a pathogen are made at the T cell level, and knowledge of their functional types is crucial to understanding the functioning and regulation of adaptive immune responses as a whole.
T lymphocytes recognise antigens based on a two-chain protein receptor. The most common and important of these are the alpha-beta T cell receptors (Figure 7.3.1).
Figure 7.3.1. Alpha-beta T cell receptor. Notice the constant and variable regions of each chain, anchored by the transmembrane region.
There are two chains in the T cell receptor, and each chain consists of two domains. The variable region domain is furthest away from the T cell membrane and is so named because its amino acid sequence varies between receptors. In contrast, the constant region domain has less variation. The differences in the amino acid sequences of the variable domains are the molecular basis of the diversity of antigens the receptor can recognise. Thus, the antigen-binding site of the receptor consists of the terminal ends of both receptor chains, and the amino acid sequences of those two areas combine to determine its antigenic specificity. Each T cell produces only one type of receptor and thus is specific for a single particular antigen.
Antigens
Antigens on pathogens are usually large and complex and consist of many antigenic determinants. An antigenic determinant (epitope) is one of the small regions within an antigen to which a receptor can bind, and antigenic determinants are limited by the size of the receptor itself. They usually consist of six or fewer amino acid residues in a protein, or one or two sugar moieties in a carbohydrate antigen. Antigenic determinants on a carbohydrate antigen are usually less diverse than on a protein antigen. Carbohydrate antigens are found on bacterial cell walls and on red blood cells (the ABO blood group antigens). Protein antigens are complex because of the variety of three-dimensional shapes that proteins can assume and are especially important for the immune responses to viruses and worm parasites. It is the interaction of the shape of the antigen and the complementary shape of the amino acids of the antigen-binding site that accounts for the chemical basis of specificity (Figure 7.3.2).
Figure 7.3.2. Antigenic determinants. A typical protein antigen has multiple antigenic determinants, shown by the ability of T cells with three different specificities to bind to different parts of the same antigen.
Antigen Processing and Presentation
Although Figure 7.3.12 shows T cell receptors interacting with antigenic determinants directly, the mechanism that T cells use to recognise antigens is, in reality, much more complex. T cells do not recognise free-floating or cell-bound antigens as they appear on the surface of the pathogen. They only recognise antigen on the surface of specialised cells called antigen-presenting cells. Antigens are internalised by these cells. Antigen processing is a mechanism that enzymatically cleaves the antigen into smaller pieces. The antigen fragments are then brought to the cell’s surface and associated with a specialised type of antigen-presenting protein known as a major histocompatibility complex (MHC) molecule. The MHC is the cluster of genes that encode these antigen-presenting molecules. The association of the antigen fragments with an MHC molecule on the surface of a cell is known as antigen presentation and results in the recognition of antigen by a T cell. This association of antigen and MHC occurs inside the cell, and it is the complex of the two that is brought to the surface. The peptide-binding cleft is a small indentation at the end of the MHC molecule that is furthest away from the cell membrane; it is here that the processed fragment of antigen sits. MHC molecules can present a variety of antigens, depending on the amino acid sequence, in their peptide-binding clefts. It is the combination of the MHC molecule and the fragment of the original peptide or carbohydrate that is actually physically recognised by the T cell receptor (Figure 7.3.3).
Figure 7.3.3. Antigen processing and presentation.
Two distinct types of MHC molecules, MHC class I and MHC class II, play roles in antigen presentation. Although produced from different genes, they both have similar functions. They bring processed antigen to the surface of the cell via a transport vesicle and present the antigen to the T cell and its receptor. Antigens from different classes of pathogens, however, use different MHC classes and take different routes through the cell to get to the surface for presentation. The basic mechanism, though, is the same. Antigens are processed by digestion, are brought into the endomembrane system of the cell, and then are expressed on the surface of the antigen-presenting cell for antigen recognition by a T cell. Intracellular antigens are typical of viruses, which replicate inside the cell, and certain other intracellular parasites and bacteria. These antigens are processed in the cytosol by an enzyme complex known as the proteasome and are then brought into the endoplasmic reticulum by the transporter associated with antigen processing (TAP) system, where they interact with class I MHC molecules and are eventually transported to the cell surface by a transport vesicle.
Extracellular antigens, characteristic of many bacteria, parasites, and fungi that do not replicate inside the cell’s cytoplasm, are brought into the endomembrane system of the cell by receptor-mediated endocytosis. The resulting vesicle fuses with vesicles from the Golgi complex, which contain pre-formed MHC class II molecules. After fusion of these two vesicles and the association of antigen and MHC, the new vesicle makes its way to the cell surface.
Antigen-Presenting Cells (APC)
Many cell types express class I molecules for the presentation of intracellular antigens. These MHC molecules may then stimulate a cytotoxic T cell immune response, eventually destroying the cell and the pathogen within. This is especially important when it comes to the most common class of intracellular pathogens, the virus. Viruses infect nearly every tissue of the body, so all these tissues must necessarily be able to express class I MHC or no T cell response can be made.
On the other hand, class II MHC molecules are expressed only on the cells of the immune system, specifically cells that affect other arms of the immune response. Thus, these cells are called “professional” antigen-presenting cells to distinguish them from those that bear class I MHC. The three types of professional antigen presenters are macrophages, dendritic cells, and B cells (Table 7.3.1).
Macrophages stimulate T cells to release cytokines that enhance phagocytosis. Dendritic cells also kill pathogens by phagocytosis, but their major function is to bring antigens to regional draining lymph nodes. The lymph nodes are the locations in which most T cell responses against pathogens of the interstitial tissues are mounted. Macrophages are found in the skin and in the lining of mucosal surfaces, such as the nasopharynx, stomach, lungs, and intestines. B cells may also present antigens to T cells, which are necessary for certain types of antibody responses, to be covered later in this chapter.
Table 7.3.1. Classes of antigen-presenting cells
MHC
Cell type
Phagocytic
Function
I
Many
No
Stimulates cytotoxic T cell immune response
II
Macrophage
Yes
Stimulates phagocytosis and presentation at primary infection site
II
Dendritic
Yes, in tissues
Brings antigens to regional lymph nodes
II
B cell
Yes, internalises surface Ig and antigen
Stimulates antibody secretion by B cells
T Cell Development and Differentiation
The process of eliminating T cells that might attack the cells of one’s own body is referred to as T cell tolerance. While thymocytes are in the cortex of the thymus, they are referred to as “double negatives,” meaning that they do not bear the CD4 or CD8 molecules that you can use to follow their pathways of differentiation (Figure 7.3.4). In the cortex of the thymus, they are exposed to cortical epithelial cells. In a process known as positive selection, double-negative thymocytes bind to the MHC molecules they observe on the thymic epithelia, and the MHC molecules of “self” are selected. This mechanism kills many thymocytes during T cell differentiation. In fact, only two percent of the thymocytes that enter the thymus leave it as mature, functional T cells.
Figure 7.3.4. Differentiation of T cells within the thymus. Thymocytes enter the thymus and go through a series of developmental stages that ensures both function and tolerance before they leave and become functional components of the adaptive immune response.
Later, the cells become double positives that express both CD4 and CD8 markers and move from the cortex to the junction between the cortex and medulla. It is here that negative selection takes place. In negative selection, self-antigens are brought into the thymus from other parts of the body by professional antigen-presenting cells. The T cells that bind to these self-antigens are selected for negatively and are killed by apoptosis. In summary, the only T cells left are those that can bind to MHC molecules of the body with foreign antigens presented on their binding clefts, preventing an attack on one’s own body tissues, at least under normal circumstances. Tolerance can be broken, however, by the development of an autoimmune response, to be discussed later in this chapter.
The cells that leave the thymus become single positives, expressing either CD4 or CD8, but not both (see Figure 7.3.14). The CD4+ T cells will bind to class II MHC and the CD8+ cells will bind to class I MHC. The discussion that follows explains the functions of these molecules and how they can be used to differentiate between the different T cell functional types.
Mechanisms of T Cell-Mediated Immune Responses
Mature T cells become activated by recognising processed foreign antigen in association with a self-MHC molecule and begin dividing rapidly by mitosis. This proliferation of T cells is called clonal expansion and is necessary to make the immune response strong enough to effectively control a pathogen. How does the body select only those T cells that are needed against a specific pathogen? Again, the specificity of a T cell is based on the amino acid sequence and the three-dimensional shape of the antigen-binding site formed by the variable regions of the two chains of the T cell receptor (Figure 7.3.5). Clonal selection is the process of antigen binding only to those T cells that have receptors specific to that antigen. Each T cell that is activated has a specific receptor “hard-wired” into its DNA, and all of its progeny will have identical DNA and T cell receptors, forming clones of the original T cell.
Figure 7.3.5. Clonal selection and expansion of T lymphocytes. Stem cells differentiate into T cells with specific receptors, called clones. The clones with receptors specific for antigens on the pathogen are selected for and expanded.
Clonal Selection and Expansion
The clonal selection theory was proposed by Frank Burnet in the 1950s. However, the term clonal selection is not a complete description of the theory, as clonal expansion goes hand in glove with the selection process. The main tenet of the theory is that a typical individual has a multitude (~4 x 1011) of different types of T cell clones based on their receptors. In this use, a clone is a group of lymphocytes that share the same antigen receptor. Each clone is necessarily present in the body in low numbers. Otherwise, the body would not have room for lymphocytes with so many specificities.
Only those clones of lymphocytes whose receptors are activated by the antigen are stimulated to proliferate. Keep in mind that most antigens have multiple antigenic determinants, so a T cell response to a typical antigen involves a polyclonal response. A polyclonal response is the stimulation of multiple T cell clones. Once activated, the selected clones increase in number and make many copies of each cell type, each clone with its unique receptor. By the time this process is complete, the body will have large numbers of specific lymphocytes available to fight the infection (see Figure 7.3.5).
The Cellular Basis of Immunological Memory
As already discussed, one of the major features of an adaptive immune response is the development of immunological memory.
During a primary adaptive immune response, both memory T cells and effector T cells are generated. Memory T cells are long-lived and can even persist for a lifetime. Memory cells are primed to act rapidly. Thus, any subsequent exposure to the pathogen will elicit a very rapid T cell response. This rapid, secondary adaptive response generates large numbers of effector T cells so fast that the pathogen is often overwhelmed before it can cause any symptoms of disease. This is what is meant by immunity to a disease. The same pattern of primary and secondary immune responses occurs in B cells and the antibody response, as will be discussed later in the chapter.
T Cell Types and Their Functions
In the discussion of T cell development, you saw that mature T cells express either the CD4 marker or the CD8 marker, but not both. These markers are cell adhesion molecules that keep the T cell in close contact with the antigen-presenting cell by directly binding to the MHC molecule (to a different part of the molecule than does the antigen). Thus, T cells and antigen-presenting cells are held together in two ways: by CD4 or CD8 attaching to MHC and by the T cell receptor binding to antigen (Figure 7.3.6).
Figure 7.3.6. Pathogen presentation. (a) CD4 is associated with helper and regulatory T cells. An extracellular pathogen is processed and presented in the binding cleft of a class II MHC molecule, and this interaction is strengthened by the CD4 molecule. (b) CD8 is associated with cytotoxic T cells. An intracellular pathogen is presented by a class I MHC molecule, and CD8 interacts with it.
Although the correlation is not 100 percent, CD4-bearing T cells are associated with helper functions and CD8-bearing T cells are associated with cytotoxicity. These functional distinctions based on CD4 and CD8 markers are useful in defining the function of each type.
Helper T Cells and Their Cytokines
Helper T cells (Th), bearing the CD4 molecule, function by secreting cytokines that act to enhance other immune responses. There are two classes of Th cells, and they act on different components of the immune response. These cells are not distinguished by their surface molecules but by the characteristic set of cytokines they secrete (Table 7.3.2).
Th1 cells are a type of helper T cell that secretes cytokines that regulate the immunological activity and development of a variety of cells, including macrophages and other types of T cells.
Th2 cells, on the other hand, are cytokine-secreting cells that act on B cells to drive their differentiation into plasma cells that make antibody. In fact, T cell help is required for antibody responses to most protein antigens, and these are called T cell-dependent antigens.
Cytotoxic T Cells
Cytotoxic T cells (Tc) are T cells that kill target cells by inducing apoptosis using the same mechanism as NK cells. They either express Fas ligand, which binds to the fas molecule on the target cell, or act by using perforins and granzymes contained in their cytoplasmic granules. As was discussed earlier with NK cells, killing a virally infected cell before the virus can complete its replication cycle results in the production of no infectious particles. As more Tc cells are developed during an immune response, they overwhelm the ability of the virus to cause disease. In addition, each Tc cell can kill more than one target cell, making them especially effective. Tc cells are so important in the antiviral immune response that some speculate that this was the main reason the adaptive immune response evolved in the first place.
Regulatory T Cells
Regulatory T cells (Treg), or suppressor T cells, are the most recently discovered of the types listed here, so less is understood about them. In addition to CD4, they bear the molecules CD25 and FOXP3. Exactly how they function is still under investigation, but it is known that they suppress other T cell immune responses. This is an important feature of the immune response, because if clonal expansion during immune responses could continue uncontrolled, these responses could lead to autoimmune diseases and other medical issues.
Not only do T cells directly destroy pathogens, but they regulate nearly all other types of the adaptive immune response as well, as evidenced by the functions of the T cell types, their surface markers, the cells they work on, and the types of pathogens they work against (see Table 5).
Table 7.3.2. Functions of T cell types and their cytokines
T cell
Main target
Function
Pathogen
Surface marker
MHC
Cytokines or mediators
Tc
Infected cells
Cytotoxicity
Intracellular
CD8
Class I
Perforins, granzymes, and fas ligand
Th1
Macrophage
Helper inducer
Extracellular
CD4
Class II
Interferon-gamma and TGF-β
Th2
B cell
Helper inducer
Extracellular
CD4
Class II
IL-4, IL-6, IL-10, and others
Treg
Th cell
Suppressor
None
CD4, CD25
?
TGF-β and IL-10
Section Review
T cells recognise antigens with their antigen receptor, a complex of two protein chains on their surface. They do not recognise self-antigens, however, but only processed antigen presented on their surfaces in a binding groove of a major histocompatibility complex molecule. T cells develop in the thymus, where they learn to use self-MHC molecules to recognise only foreign antigens, thus making them tolerant to self-antigens. There are several functional types of T lymphocytes, the major ones being helper, regulatory and cytotoxic T cells.
7.4 The Adaptive Immune Response: B-Lymhocytes and Antibodies
Learning Objectives
By the end of this section, you will be able to:
Explain how B cells mature and how B cell tolerance develops
Discuss how B cells are activated and differentiate into plasma cells
Describe the structure of the antibody classes and their functions
Antibodies were the first component of the adaptive immune response to be characterised by scientists working on the immune system. It was already known that individuals who survived a bacterial infection were immune to re-infection with the same pathogen. Early microbiologists took serum from an immune patient and mixed it with a fresh culture of the same type of bacteria, then observed the bacteria under a microscope. The bacteria became clumped in a process called agglutination. When a different bacterial species was used, the agglutination did not happen. Thus, there was something in the serum of immune individuals that could specifically bind to and agglutinate bacteria.
Scientists now know the cause of the agglutination is an antibody molecule, also called an immunoglobulin. What is an antibody? An antibody protein is essentially a secreted form of a B cell receptor. (In fact, surface immunoglobulin is another name for the B cell receptor.) Not surprisingly, the same genes encode both the secreted antibodies and the surface immunoglobulins. One minor difference in the way these proteins are synthesised distinguishes a naïve B cell with antibody on its surface from an antibody-secreting plasma cell with no antibodies on its surface. The antibodies of the plasma cell have the exact same antigen-binding site and specificity as their B cell precursors.
There are five different classes of antibody found in humans: IgM, IgD, IgG, IgA, and IgE. Each of these has specific functions in the immune response, so by learning about them, researchers can learn about the great variety of antibody functions critical to many adaptive immune responses.
B cells do not recognise antigen in the complex fashion of T cells. B cells can recognise native, unprocessed antigen and do not require the participation of MHC molecules and antigen-presenting cells.
B Cell Differentiation and Activation
B cells differentiate in the bone marrow. During the process of maturation, up to 100 trillion different clones of B cells are generated, which is similar to the diversity of antigen receptors seen in T cells.
B cell differentiation and the development of tolerance are not quite as well understood as it is in T cells. Central tolerance is the destruction or inactivation of B cells that recognise self-antigens in the bone marrow, and its role is critical and well established. In the process of clonal deletion, immature B cells that bind strongly to self-antigens expressed on tissues are signalled to commit suicide by apoptosis, removing them from the population. In the process of clonal anergy, however, B cells exposed to soluble antigen in the bone marrow are not physically deleted but become unable to function.
Another mechanism called peripheral tolerance is a direct result of T cell tolerance. In peripheral tolerance, functional, mature B cells leave the bone marrow but have yet to be exposed to self-antigen. Most protein antigens require signals from helper T cells (Th2) to proceed to make antibody. When a B cell binds to a self-antigen but receives no signals from a nearby Th2 cell to produce antibody, the cell is signalled to undergo apoptosis and is destroyed. This is yet another example of the control that T cells have over the adaptive immune response.
After B cells are activated by their binding to antigen, they differentiate into plasma cells. Plasma cells often leave the secondary lymphoid organs, where the response is generated, and migrate back to the bone marrow, where the whole differentiation process started. After secreting antibodies for a specific period, they die, as most of their energy is devoted to making antibodies and not to maintaining themselves. Thus, plasma cells are said to be terminally differentiated.
The final B cell of interest is the memory B cell, which results from the clonal expansion of an activated B cell. Memory B cells function in a way similar to memory T cells. They lead to a stronger and faster secondary response when compared to the primary response, as illustrated below.
Antibody Structure
Antibodies are glycoproteins consisting of two types of polypeptide chains with attached carbohydrates. The heavy chain and the light chain are the two polypeptides that form the antibody. The main differences between the classes of antibodies are in the differences between their heavy chains, but as you shall see, the light chains have an important role, forming part of the antigen-binding site on the antibody molecules.
Four-Chain Models of Antibody Structures
All antibody molecules have two identical heavy chains and two identical light chains. (Some antibodies contain multiple units of this four-chain structure.) The Fc region of the antibody is formed by the two heavy chains coming together, usually linked by disulphide bonds (Figure 7.4.1). The Fc portion of the antibody is important in that many effector cells of the immune system have Fc receptors. Cells having these receptors can then bind to antibody-coated pathogens, greatly increasing the specificity of the effector cells. At the other end of the molecule are two identical antigen-binding sites.
Figure 7.4.1. Antibody and IgG2 structures. The typical four chain structure of a generic antibody (a) and the corresponding three-dimensional structure of the antibody IgG2 (b). (credit b: modification of work by Tim Vickers).
Five Classes of Antibodies and their Functions
In general, antibodies have two basic functions. They can act as the B cell antigen receptor or they can be secreted, circulate, and bind to a pathogen, often labelling it for identification by other forms of the immune response. Of the five antibody classes, notice that only two can function as the antigen receptor for naïve B cells: IgM and IgD (Figure 7.4.2). Mature B cells that leave the bone marrow express both IgM and IgD, but both antibodies have the same antigen specificity. Only IgM is secreted, however, and no other nonreceptor function for IgD has been discovered.
Figure 7.4.2. Five Classes of antibodies.
IgM consists of five four-chain structures (20 total chains with 10 identical antigen-binding sites) and is thus the largest of the antibody molecules. IgM is usually the first antibody made during a primary response. Its 10 antigen-binding sites and large shape allow it to bind well to many bacterial surfaces. It is excellent at binding complement proteins and activating the complement cascade, consistent with its role in promoting chemotaxis, opsonisation, and cell lysis. Thus, it is a very effective antibody against bacteria at early stages of a primary antibody response. As the primary response proceeds, the antibody produced in a B cell can change to IgG, IgA, or IgE by the process known as class switching. Class switching is the change of one antibody class to another. While the class of antibody changes, the specificity and the antigen-binding sites do not. Thus, the antibodies made are still specific to the pathogen that stimulated the initial IgM response.
IgG is a major antibody of late primary responses and the main antibody of secondary responses in the blood. This is because class switching occurs during primary responses. IgG is a monomeric antibody that clears pathogens from the blood and can activate complement proteins (although not as well as IgM), taking advantage of its antibacterial activities. Furthermore, this class of antibody is the one that crosses the placenta to protect the developing foetus from disease exits the blood to the interstitial fluid to fight extracellular pathogens.
IgA exists in two forms, a four-chain monomer in the blood and an eight-chain structure, or dimer, in exocrine gland secretions of the mucous membranes, including mucus, saliva and tears. Thus, dimeric IgA is the only antibody to leave the interior of the body to protect body surfaces. IgA is also of importance to newborns, because this antibody is present in mother’s breast milk (colostrum), which serves to protect the infant from disease.
IgE is usually associated with allergies and anaphylaxis. It is present in the lowest concentration in the blood, because its Fc region binds strongly to an IgE-specific Fc receptor on the surfaces of mast cells. IgE makes mast cell degranulation very specific, such that if a person is allergic to peanuts, there will be peanut-specific IgE bound to his or her mast cells. In this person, eating peanuts will cause the mast cells to degranulate, sometimes causing severe allergic reactions, including anaphylaxis, a severe, systemic allergic response that can cause death.
Clonal Selection of B Cells
Clonal selection and expansion work much the same way in B cells as in T cells. Only B cells with appropriate antigen specificity are selected for and expanded (Figure 7.4.3). Eventually, the plasma cells secrete antibodies with antigenic specificity identical to those that were on the surfaces of the selected B cells. Notice in the figure that both plasma cells and memory B cells are generated simultaneously.
Figure 7.4.3. Clonal selection of B cells. During a primary B cell immune response, both antibody-secreting plasma cells and memory B cells are produced. These memory cells lead to the differentiation of more plasma cells and memory B cells during secondary responses.
Primary Versus Secondary B Cell Reponses
Figure 7.4.4. Primary and secondary antibody responses. Antigen A is given once to generate a primary response and later to generate a secondary response. When a different antigen is given for the first time, a new primary response is made.
Primary and secondary responses as they relate to T cells were discussed earlier. This section will look at these responses with B cells and antibody production. Because antibodies are easily obtained from blood samples, they are easy to follow and graph (Figure 7.4.4). As you will see from the figure, the primary response to an antigen (representing a pathogen) is delayed by several days. This is the time it takes for the B cell clones to expand and differentiate into plasma cells. The level of antibody produced is low, but it is sufficient for immune protection. The second time a person encounters the same antigen, there is no time delay, and the amount of antibody made is much higher. Thus, the secondary antibody response overwhelms the pathogens quickly and, in most situations, no symptoms are felt. When a different antigen is used, another primary response is made with its low antibody levels and time delay.
Active Versus Passive Immunity
Immunity to pathogens, and the ability to control pathogen growth so that damage to the tissues of the body is limited, can be acquired by (1) the active development of an immune response in the infected individual or (2) the passive transfer of immune components from an immune individual to a nonimmune one. Both active and passive immunity have examples in the natural world and as part of medicine.
Active immunity is the resistance to pathogens acquired during an adaptive immune response within an individual (Table 7.4.1). Naturally acquired active immunity, the response to a pathogen, is the focus of this chapter. Artificially acquired active immunity involves the use of vaccines. A vaccine is a killed or weakened pathogen or its components that, when administered to a healthy individual, leads to the development of immunological memory (a weakened primary immune response) without causing much in the way of symptoms. Thus, with the use of vaccines, one can avoid the damage from disease that results from the first exposure to the pathogen yet reap the benefits of protection from immunological memory. The advent of vaccines was one of the major medical advances of the twentieth century and led to the eradication of smallpox and the control of many infectious diseases, including polio, measles, and whooping cough.
Table 7.4.1. Active versus passive immunity
Natural
Artificial
Active
Adaptive immune response
Vaccine response
Passive
Trans-placental antibodies/breastfeeding
Immune globulin injections
Passive immunity arises from the transfer of antibodies to an individual without requiring them to mount their own active immune response. Naturally acquired passive immunity is seen during foetal development. IgG is transferred from the maternal circulation to the foetus via the placenta, protecting the foetus from infection and protecting the newborn for the first few months of its life. As already stated, a newborn benefits from the IgA antibodies it obtains from milk during breastfeeding. The foetus and newborn thus benefit from the immunological memory of the mother to the pathogens to which she has been exposed. In medicine, artificially acquired passive immunity usually involves injections of immunoglobulins, taken from animals previously exposed to a specific pathogen. This treatment is a fast-acting method of temporarily protecting an individual who was possibly exposed to a pathogen. The downside to both types of passive immunity is the lack of the development of immunological memory. Once the antibodies are transferred, they are effective for only a limited time before they degrade.
T Cell-Development versus T Cell-Independent Antigens
As discussed previously, Th2 cells secrete cytokines that drive the production of antibodies in a B cell, responding to complex antigens such as those made by proteins. On the other hand, some antigens are T cell independent. A T cell-independent antigen usually is in the form of repeated carbohydrate moieties found on the cell walls of bacteria. Each antibody on the B cell surface has two binding sites, and the repeated nature of T cell-independent antigen leads to crosslinking of the surface antibodies on the B cell. The crosslinking is enough to activate it in the absence of T cell cytokines.
A T cell-dependent antigen, on the other hand, usually is not repeated to the same degree on the pathogen and thus does not crosslink surface antibody with the same efficiency. To elicit a response to such antigens, the B and T cells must come close together (Figure 7.4.5). The B cell must receive two signals to become activated. Its surface immunoglobulin must recognise native antigen. Some of this antigen is internalised, processed, and presented to the Th2 cells on a class II MHC molecule. The T cell then binds using its antigen receptor and is activated to secrete cytokines that diffuse to the B cell, finally activating it completely. Thus, the B cell receives signals from both its surface antibody and the T cell via its cytokines and acts as a professional antigen-presenting cell in the process.
Figure 7.4.5. T and B cell binding. To elicit a response to a T cell-dependent antigen, the B and T cells must come close together. To become fully activated, the B cell must receive two signals from the native antigen and the T cell’s cytokines.
Section Review
B cells, which develop within the bone marrow, are responsible for making five different classes of antibodies, each with its own functions. B cells have their own mechanisms for tolerance, but in peripheral tolerance, the B cells that leave the bone marrow remain inactive due to T cell tolerance. Some B cells do not need T cell cytokines to make antibody, and they bypass this need by the crosslinking of their surface immunoglobulin by repeated carbohydrate residues found in the cell walls of many bacterial species. Others require T cells to become activated.
Explain the development of immunological competence
Describe the mucosal immune response
Discuss immune responses against bacterial, viral, fungal, and animal pathogens
Describe different ways pathogens evade immune responses
Now that you understand the development of mature, naïve B cells and T cells, and some of their major functions, how do all of these various cells, proteins, and cytokines come together to actually resolve an infection? Ideally, the immune response will rid the body of a pathogen entirely. The adaptive immune response, with its rapid clonal expansion, is well suited to this purpose. Think of a primary infection as a race between the pathogen and the immune system. The pathogen bypasses barrier defences and starts multiplying in the host’s body. During the first 4 to 5 days, the innate immune response will partially control, but not stop, pathogen growth. As the adaptive immune response gears up, however, it will begin to clear the pathogen from the body, while at the same time becoming stronger and stronger. When following antibody responses in patients with a particular disease such as a virus, this clearance is referred to as seroconversion (sero- = “serum”). Seroconversion is the reciprocal relationship between virus levels in the blood and antibody levels. As the antibody levels rise, the virus levels decline, and this is a sign that the immune response is being at least partially effective (partially, because in many diseases, seroconversion does not necessarily mean a patient is getting well).
An excellent example of this is seroconversion during HIV disease (Figure 7.5.1). Notice that antibodies are made early in this disease, and the increase in anti-HIV antibodies correlates with a decrease in detectable virus in the blood. Although these antibodies are an important marker for diagnosing the disease, they are not sufficient to completely clear the virus. Several years later, the vast majority of these individuals, if untreated, will lose their entire adaptive immune response, including the ability to make antibodies, during the final stages of AIDS.
Figure 7.5.1. HIV disease progression. Seroconversion, the rise of anti-HIV antibody levels and the concomitant decline in measurable virus levels, happens during the first several months of HIV disease. Unfortunately, this antibody response is ineffective at controlling the disease, as seen by the progression of the disease towards AIDS, in which all adaptive immune responses are compromised.
Everyday Connection Disinfectants: Fighting the Good Fight?
“Wash your hands!” Parents have been telling their children this for generations. Dirty hands can spread disease. But is it possible to get rid of enough pathogens that children will never get sick? Are children who avoid exposure to pathogens better off? The answers to both these questions appears to be no.
Antibacterial wipes, soaps, gels, and even toys with antibacterial substances embedded in their plastic are ubiquitous in our society. Still, these products do not rid the skin and gastrointestinal tract of bacteria, and it would be harmful to our health if they did. We need these non-pathogenic bacteria on and within our bodies to keep the pathogenic ones from growing. The urge to keep children perfectly clean is thus probably misguided. Children will get sick anyway, and the later benefits of immunological memory far outweigh the minor discomforts of most childhood diseases. In fact, getting diseases such as chickenpox or measles later in life is much harder on the adult and are associated with symptoms significantly worse than those seen in the childhood illnesses. Of course, vaccinations help children avoid some illnesses, but there are so many pathogens, we will never be immune to them all.
Could over-cleanliness be the reason that allergies are increasing in more developed countries? Some scientists think so. Allergies are based on an IgE antibody response. Many scientists think the system evolved to help the body rid itself of worm parasites. The hygiene theory is the idea that the immune system is geared to respond to antigens, and if pathogens are not present, it will respond instead to inappropriate antigens such as allergens and self-antigens. This is one explanation for the rising incidence of allergies in developed countries, where the response to nonpathogens like pollen, shrimp, and cat dander cause allergic responses while not serving any protective function.
The Mucosal Immune Response
Mucosal tissues are major barriers to the entry of pathogens into the body. The IgA (and sometimes IgM) antibodies in mucus and other secretions can bind to the pathogen, and in the cases of many viruses and bacteria, neutralise them. Neutralisation is the process of coating a pathogen with antibodies, making it physically impossible for the pathogen to bind to receptors. Neutralisation, which occurs in the blood, lymph, and other body fluids and secretions, protects the body constantly. Neutralising antibodies are the basis for the disease protection offered by vaccines. Vaccinations for diseases that commonly enter the body via mucous membranes, such as influenza, are usually formulated to enhance IgA production.
Immune responses in some mucosal tissues such as the Peyer’s patches in the small intestine take up particulate antigens by specialised cells known as microfold or M cells (Figure 7.5.2). These cells allow the body to sample potential pathogens from the intestinal lumen. Dendritic cells then take the antigen to the regional lymph nodes, where an immune response is mounted.
Figure 7.5.2. IgA immunity. The nasal-associated lymphoid tissue and Peyer’s patches of the small intestine generate IgA immunity. Both use M cells to transport antigen inside the body so that immune responses can be mounted.
Defences Against Bacteria and Fungi
The body fights bacterial pathogens with a wide variety of immunological mechanisms, essentially trying to find one that is effective. Bacteria such as Mycobacterium leprae, the cause of leprosy, are resistant to lysosomal enzymes and can persist in macrophage organelles or escape into the cytosol. In such situations, infected macrophages receiving cytokine signals from Th1 cells turn on special metabolic pathways. Macrophage oxidative metabolism is hostile to intracellular bacteria, often relying on the production of nitric oxide to kill the bacteria inside the macrophage.
Fungal infections, such as those from Aspergillus, Candida, and Pneumocystis, are largely opportunistic infections that take advantage of suppressed immune responses. Most of the same immune mechanisms effective against bacteria have similar effects on fungi, both of which have characteristic cell wall structures that protect their cells.
Defences Against Parasites
Worm parasites such as helminths are the primary reason why the mucosal immune response, IgE-mediated allergy and asthma, and eosinophils evolved. These parasites were at one time very common in human society. When infecting a human, often via contaminated food, some worms take up residence in the gastrointestinal tract. Eosinophils are attracted to the site by T cell cytokines, which release their granule contents upon their arrival. Mast cell degranulation also occurs, and the fluid leakage caused by the increase in local vascular permeability is thought to have a flushing action on the parasite, expelling its larvae from the body. Furthermore, if IgE labels the parasite, the eosinophils can bind to it by its Fc receptor.
Defences Against Viruses
The primary mechanisms against viruses are NK cells, interferons, and cytotoxic T cells. Antibodies are effective against viruses mostly during protection, where an immune individual can neutralise them based on a previous exposure. Antibodies have no effect on viruses or other intracellular pathogens once they enter the cell, since antibodies are not able to penetrate the plasma membrane of the cell. Many cells respond to viral infections by downregulating their expression of MHC class I molecules. This is to the advantage of the virus, because without class I expression, cytotoxic T cells have no activity. NK cells, however, can recognise virally infected class I-negative cells and destroy them. Thus, NK and cytotoxic T cells have complementary activities against virally infected cells.
Interferons have activity in slowing viral replication and are used in the treatment of certain viral diseases, such as hepatitis B and C, but their ability to eliminate the virus completely is limited. The cytotoxic T cell response, though, is key, as it eventually overwhelms the virus and kills infected cells before the virus can complete its replicative cycle. Clonal expansion and the ability of cytotoxic T cells to kill more than one target cell make these cells especially effective against viruses. In fact, without cytotoxic T cells, it is likely that humans would all die at some point from a viral infection (if no vaccine were available).
Evasion of the Immune System by Pathogens
It is important to keep in mind that although the immune system has evolved to be able to control many pathogens, pathogens themselves have evolved ways to evade the immune response. An example already mentioned is in Mycobacterium tuberculosis, which has evolved a complex cell wall that is resistant to the digestive enzymes of the macrophages that ingest them, and thus persists in the host, causing the chronic disease tuberculosis. This section briefly summarises other ways in which pathogens can “outwit” immune responses. But keep in mind, although it seems as if pathogens have a will of their own, they do not. All of these evasive “strategies” arose strictly by evolution, driven by selection.
Bacteria sometimes evade immune responses because they exist in multiple strains, such as different groups of Staphylococcus aureus. S. aureus is commonly found in minor skin infections, such as boils, and some healthy people harbour it in their nose. One small group of strains of this bacterium, however, called methicillin-resistant Staphylococcus aureus (MRSA), has become resistant to multiple antibiotics and is essentially untreatable. Different bacterial strains differ in the antigens on their surfaces. The immune response against one strain (antigen) does not affect the other; thus, the species survives.
Another method of immune evasion is mutation. Because viruses’ surface molecules mutate continuously, viruses like influenza change enough each year that the flu vaccine for one year may not protect against the flu common to the next. New vaccine formulations must be derived for each flu season.
Genetic recombination—the combining of gene segments from two different pathogens—is an efficient form of immune evasion. For example, the influenza virus contains gene segments that can recombine when two different viruses infect the same cell. Recombination between human and pig influenza viruses led to the 2010 H1N1 swine flu outbreak.
Pathogens can produce immunosuppressive molecules that impair immune function, and there are several different types. Viruses are especially good at evading the immune response in this way, and many types of viruses have been shown to suppress the host immune response in ways much more subtle than the wholesale destruction caused by HIV.
Section Review
Early childhood is a time when the body develops much of its immunological memory that protects it from diseases in adulthood. The components of the immune response that have the maximum effectiveness against a pathogen are often associated with the class of pathogen involved. Bacteria and fungi are especially susceptible to damage by complement proteins, whereas viruses are taken care of by interferons and cytotoxic T cells. Worms are attacked by eosinophils. Pathogens have shown the ability, however, to evade the body’s immune responses, some leading to chronic infections or even death. The immune system and pathogens are in a slow, evolutionary race to see who stays on top. Modern medicine, hopefully, will keep the results skewed in humans’ favour.
7.6 Diseases Associated with Depressed or Overactive Immune Responses
Learning Objectives
By the end of this section, you will be able to:
Discuss inherited and acquired immunodeficiencies
Explain the four types of hypersensitivity and how they differ
Give an example of how autoimmune disease breaks tolerance
This section is about how the immune system goes wrong. When it goes haywire, and becomes too weak or too strong, it leads to a state of disease. The factors that maintain immunological homeostasis are complex and incompletely understood.
Immunodeficiencies
As you have seen, the immune system is quite complex. It has many pathways using many cell types and signals. Because it is so complex, there are many ways for it to go wrong. Inherited immunodeficiencies arise from gene mutations that affect specific components of the immune response. There are also acquired immunodeficiencies with potentially devastating effects on the immune system, such as HIV.
Inherited Immunodeficiencies
A list of all inherited immunodeficiencies is well beyond the scope of this chapter. The list is almost as long as the list of cells, proteins, and signalling molecules of the immune system itself. Some deficiencies, such as those for complement, cause only a higher susceptibility to some Gram-negative bacteria. Others are more severe in their consequences. Certainly, the most serious of the inherited immunodeficiencies is severe combined immunodeficiency disease (SCID). This disease is complex because it is caused by many different genetic defects. What groups them together is the fact that both the B cell and T cell arms of the adaptive immune response are affected.
Children with this disease usually die of opportunistic infections within their first year of life unless they receive a bone marrow transplant. Such a procedure had not yet been perfected for David Vetter, the “boy in the bubble,” who was treated for SCID by having to live almost his entire life in a sterile plastic cocoon for the 12 years before his death from infection in 1984. One of the features that make bone marrow transplants work as well as they do is the proliferative capability of haematopoietic stem cells of the bone marrow. Only a small amount of bone marrow from a healthy donor is given intravenously to the recipient. It finds its own way to the bone where it populates it, eventually reconstituting the patient’s immune system, which is usually destroyed beforehand by treatment with radiation or chemotherapeutic drugs.
New treatments for SCID using gene therapy, inserting non-defective genes into cells taken from the patient and giving them back, have the advantage of not needing the tissue match required for standard transplants. Although not a standard treatment, this approach holds promise, especially for those in whom standard bone marrow transplantation has failed.
Human Immunodeficiency Viruses/AIDS
Although many viruses cause suppression of the immune system, only one wipes it out completely, and that is the previously mentioned HIV. It is worth discussing the biology of this virus, which can lead to the well-known AIDS, so that its full effects on the immune system can be understood. The virus is transmitted through semen, vaginal fluids, and blood, and can be caught by risky sexual behaviours and the sharing of needles by intravenous drug users. There are sometimes, but not always, flu-like symptoms in the first one to two weeks after infection. This is later followed by seroconversion. The anti-HIV antibodies formed during seroconversion are the basis for most initial HIV screening done in Australia, and many other countries such as United States and England. Because seroconversion takes different lengths of time in different individuals, multiple AIDS tests are given months apart to confirm or eliminate the possibility of infection.
After seroconversion, the amount of virus circulating in the blood drops and stays at a low level for several years. During this time, the levels of CD4+ cells, especially helper T cells, decline steadily, until at some point, the immune response is so weak that opportunistic disease and eventually death result. CD4 is the receptor that HIV uses to get inside T cells and reproduce. Given that CD4+ helper T cells play an important role in other in T cell immune responses and antibody responses, it should be no surprise that both types of immune responses are eventually seriously compromised.
Treatment for the disease consists of drugs that target virally encoded proteins that are necessary for viral replication but are absent from normal human cells. By targeting the virus itself and sparing the cells, this approach has been successful in significantly prolonging the lives of HIV-positive individuals. On the other hand, an HIV vaccine has been 30 years in development and is still years away. Because the virus mutates rapidly to evade the immune system, scientists have been looking for parts of the virus that do not change and thus would be good targets for a vaccine candidate.
Hypersensitivities
The word “hypersensitivity” simply means sensitive beyond normal levels of activation. Allergies and inflammatory responses to non-pathogenic environmental substances have been observed since the dawn of history. Hypersensitivity is a medical term describing symptoms that are now known to be caused by unrelated mechanisms of immunity. Still, it is useful for this discussion to use the four types of hypersensitivities as a guide to understand these mechanisms (Figure 7.6.1).
Figure 7.6.1. Immune hypersensitivity. Components of the immune system cause four types of hypersensitivity. Notice that types I–III are B cell mediated, whereas type IV hypersensitivity is exclusively a T cell phenomenon.
Immediate (Type I) Hypersensitivity
Antigens that cause allergic responses are often referred to as allergens. The specificity of the immediate hypersensitivity response is predicated on the binding of allergen-specific IgE to the mast cell surface. The process of producing allergen-specific IgE is called sensitisation, and is a necessary prerequisite for the symptoms of immediate hypersensitivity to occur. Allergies and allergic asthma are mediated by mast cell degranulation that is caused by the crosslinking of the antigen-specific IgE molecules on the mast cell surface. The mediators released have various vasoactive effects already discussed, but the major symptoms of inhaled allergens are the nasal oedema and runny nose caused by the increased vascular permeability and increased blood flow of nasal blood vessels. As these mediators are released with mast cell degranulation, type I hypersensitivity reactions are usually rapid and occur within just a few minutes, hence the term immediate hypersensitivity.
Most allergens are in themselves non-pathogenic and therefore innocuous. Some individuals develop mild allergies, which are usually treated with antihistamines. Others develop severe allergies that may cause anaphylactic shock, which can potentially be fatal within 20 to 30 minutes if untreated. This drop in blood pressure (shock) with accompanying contractions of bronchial smooth muscle is caused by systemic mast cell degranulation when an allergen is eaten (for example, shellfish and peanuts), injected (by a bee sting or being administered penicillin), or inhaled (asthma). Because adrenaline raises blood pressure and relaxes bronchial smooth muscle, it is routinely used to counteract the effects of anaphylaxis and can be lifesaving. Patients with known severe allergies are encouraged to keep automatic adrenaline injectors with them at all times, especially when away from easy access to hospitals.
Allergists use skin testing to identify allergens in type I hypersensitivity. In skin testing, allergen extracts are injected into the epidermis, and a positive result of a soft, pale swelling at the site surrounded by a red zone (called the wheal and flare response), caused by the release of histamine and the granule mediators, usually occurs within 30 minutes. The soft centre is due to fluid leaking from the blood vessels and the redness is caused by the increased blood flow to the area that results from the dilation of local blood vessels at the site.
Type II and Type III Hypersensitivities
Type II hypersensitivity, which involves IgG-mediated lysis of cells by complement proteins, occurs during mismatched blood transfusions and blood compatibility diseases such as erythroblastosis fetalis (see section on transplantation). Type III hypersensitivity occurs with diseases such as systemic lupus erythematosus, where soluble antigens, mostly DNA and other material from the nucleus, and antibodies accumulate in the blood to the point that the antigen and antibody precipitate along blood vessel linings. These immune complexes often lodge in the kidneys, joints, and other organs where they can activate complement proteins and cause inflammation.
Delayed (Type IV) Hypersensitivity
Delayed hypersensitivity, or type IV hypersensitivity, is basically a standard cellular immune response. In delayed hypersensitivity, the first exposure to an antigen is called sensitisation, such that on re-exposure, a secondary cellular response results, secreting cytokines that recruit macrophages and other phagocytes to the site. These sensitised T cells, of the Th1 class, will also activate cytotoxic T cells. The time it takes for this reaction to occur accounts for the 24- to 72-hour delay in development.
The classical test for delayed hypersensitivity is the tuberculin test for tuberculosis, where bacterial proteins from M. tuberculosis are injected into the skin. A couple of days later, a positive test is indicated by a raised red area that is hard to the touch, called an induration, which is a consequence of the cellular infiltrate, an accumulation of activated macrophages. A positive tuberculin test means that the patient has been exposed to the bacteria and exhibits a cellular immune response to it.
Another type of delayed hypersensitivity is contact sensitivity, where substances such as the metal nickel cause a red and swollen area upon contact with the skin. The individual must have been previously sensitised to the metal. A much more severe case of contact sensitivity is poison ivy (uncommon in Australia) but many of the harshest symptoms of the reaction are associated with the toxicity of its oils and are not T cell mediated.
Autoimmune Responses
The worst cases of the immune system over-reacting are autoimmune diseases. Somehow, tolerance breaks down and the immune systems in individuals with these diseases begin to attack their own bodies, causing significant damage. The trigger for these diseases is, more often than not, unknown, and the treatments are usually based on resolving the symptoms using immunosuppressive and anti-inflammatory drugs such as steroids. These diseases can be localised and crippling, as in rheumatoid arthritis, or diffuse in the body with multiple symptoms that differ in different individuals, as is the case with systemic lupus erythematosus (Figure 7.6.2).
Figure 7.6.2. Autoimmune disorders: rheumatoid arthritis and lupus. (a) Extensive damage to the right hand of a rheumatoid arthritis sufferer is shown in the x-ray. (b) The diagram shows a variety of possible symptoms of systemic lupus erythematosus.
Environmental triggers seem to play large roles in autoimmune responses. One explanation for the breakdown of tolerance is that, after certain bacterial infections, an immune response to a component of the bacterium cross-reacts with a self-antigen. This mechanism is seen in rheumatic fever, a result of infection with Streptococcus bacteria, which causes strep throat. The antibodies to this pathogen’s M protein cross-react with an antigenic component of heart myosin, a major contractile protein of the heart that is critical to its normal function. The antibody binds to these molecules and activates complement proteins, causing damage to the heart, especially to the heart valves. On the other hand, some theories propose that having multiple common infectious diseases prevents autoimmune responses. The fact that autoimmune diseases are rare in countries that have a high incidence of infectious diseases supports this idea, another example of the hygiene hypothesis discussed earlier in this chapter.
There are genetic factors in autoimmune diseases as well. Some diseases are associated with the MHC genes that an individual expresses. The reason for this association is likely because if one’s MHC molecules are not able to present a certain self-antigen, then that particular autoimmune disease cannot occur. Overall, there are more than 80 different autoimmune diseases, which are a significant health problem in the elderly. Table 7.6.1 lists several of the most common autoimmune diseases, the antigens that are targeted, and the segment of the adaptive immune response that causes the damage.
Table 7.6.1. Autoimmune diseases
Disease
Autoantigen
Symptoms
Coeliac disease
Tissue transglutaminase
Damage to small intestine
Diabetes mellitus type I
Beta cells of pancreas
Low insulin production; inability to regulate serum glucose
Thyroid-stimulating hormone receptor (antibody mimics hormone and stimulates receptor)
Hypothyroidism
Lupus erythematosus
Nuclear DNA and proteins
Damage of many body systems
Myasthenia gravis
Acetylcholine receptor in neuromuscular junctions
Debilitating muscle weakness
Rheumatoid arthritis
Joint capsule antigens
Chronic inflammation of joints
Section Review
The immune response can be under-reactive or over-reactive. Suppressed immunity can result from inherited genetic defects or by acquiring viruses. Over-reactive immune responses include the hypersensitivities: B cell- and T cell-mediated immune responses designed to control pathogens, but that lead to symptoms or medical complications. The worst cases of over-reactive immune responses are autoimmune diseases, where an individual’s immune system attacks his or her own body because of the breakdown of immunological tolerance. These diseases are more common in the aged, so treating them will be a challenge in the future as the aged population in the world increases.
Explain why blood typing is important and what happens when mismatched blood is used in a transfusion
Describe how tissue typing is done during organ transplantation and the role of transplant anti-rejection drugs
Show how the immune response is able to control some cancers and how this immune response might be enhanced by cancer vaccines
The immune responses to transplanted organs and to cancer cells are both important medical issues. With the use of tissue typing and anti-rejection drugs, transplantation of organs and the control of the anti-transplant immune response have made huge strides in the past 50 years. Today, these procedures are commonplace. Tissue typing is the determination of MHC molecules in the tissue to be transplanted to better match the donor to the recipient. The immune response to cancer, on the other hand, has been more difficult to understand and control. Although the immune system can recognise some cancers and control them, others seem to be resistant to immune mechanisms.
The Rh Factor
Red blood cells can be typed based on their surface antigens. ABO blood type, in which individuals are type A, B, AB, or O according to their genetics, is one example. A separate antigen system seen on red blood cells is the Rh antigen. When someone is “A positive” for example, the positive refers to the presence of the Rh antigen, whereas someone who is “A negative” would lack this molecule.
An interesting consequence of Rh factor expression is seen in erythroblastosis foetalis, a haemolytic disease of the newborn (Figure 7.7.1). This disease occurs when mothers negative for Rh antigen have multiple Rh-positive children. During the birth of a first Rh-positive child, the mother makes a primary anti-Rh antibody response to the foetal blood cells that enter the maternal bloodstream. If the mother has a second Rh-positive child, IgG antibodies against Rh-positive blood mounted during this secondary response cross the placenta and attack the foetal blood, causing anaemia. This is a consequence of the fact that the foetus is not genetically identical to the mother, and thus the mother can mount an immune response against it. This disease is treated with antibodies specific for Rh factor. These are given to the mother during the subsequent births, destroying any foetal blood that might enter her system and preventing the immune response.
Figure 7.7.1. Erythroblastosis foetalis. Erythroblastosis foetalis (haemolytic disease of the newborn) is the result of an immune response in an Rh-negative mother who has multiple children with an Rh-positive father. During the first birth, foetal blood enters the mother’s circulatory system, and anti-Rh antibodies are made. During the gestation of the second child, these antibodies cross the placenta and attack the blood of the foetus. The treatment for this disease is to give the mother anti-Rh antibodies (RhoGAM) during the first pregnancy to destroy Rh-positive foetal red blood cells from entering her system and causing the anti-Rh antibody response in the first place.
Tissue Transplantation
Tissue transplantation is more complicated than blood transfusions because of two characteristics of MHC molecules. These molecules are the major cause of transplant rejection (hence the name “histocompatibility”). MHC polygeny refers to the multiple MHC proteins on cells, and MHC polymorphism refers to the multiple alleles for each individual MHC locus. Thus, there are many alleles in the human population that can be expressed (Table 7.7.1 and Table 7.7.2). When a donor organ expresses MHC molecules that are different from the recipient, the latter will often mount a cytotoxic T cell response to the organ and reject it. Histologically, if a biopsy of a transplanted organ exhibits massive infiltration of T lymphocytes within the first weeks after transplant, it is a sign that the transplant is likely to fail. The response is a classical, and very specific, primary T cell immune response. As far as medicine is concerned, the immune response in this scenario does the patient no good at all and causes significant harm.
Table 7.7.1. Partial table of alleles of the human MHC (Class I)
Gene
# of alleles
# of possible MHC I protein components
A
2132
1527
B
2798
2110
C
1672
1200
E
11
3
F
22
4
G
50
16
Table 7.7.2. Partial table of alleles of the human MHC (Class II)
Gene
# of alleles
# of possible MHC II protein components
DRA
7
2
DRB
1297
958
DQA1
49
31
DQB1
179
128
DPA1
36
18
DPB1
158
136
DMA
7
4
DMB
13
7
DOA
12
3
DOB
13
5
Immunosuppressive drugs such as cyclosporine A have made transplants more successful but matching the MHC molecules is still key. In humans, there are six MHC molecules that show the most polymorphisms, three class I molecules (A, B, and C) and three class II molecules called DP, DQ, and DR. A successful transplant usually requires a match between at least 3–4 of these molecules, with more matches associated with greater success. Family members, since they share a similar genetic background, are much more likely to share MHC molecules than unrelated individuals do. In fact, due to the extensive polymorphisms in these MHC molecules, unrelated donors are found only through a worldwide database. The system is not foolproof however, as there are not enough individuals in the system to provide the organs necessary to treat all patients needing them.
One disease of transplantation occurs with bone marrow transplants, which are used to treat various diseases, including SCID and leukaemia. Because the bone marrow cells being transplanted contain lymphocytes capable of mounting an immune response, and because the recipient’s immune response has been destroyed before receiving the transplant, the donor cells may attack the recipient tissues, causing graft-versus-host disease. Symptoms of this disease, which usually include a rash and damage to the liver and mucosa, are variable, and attempts have been made to moderate the disease by first removing mature T cells from the donor bone marrow before transplanting it.
Immune Responses Against Cancer
Figure 7.7.2. Kaposi’s sarcoma lesions. (credit: National Cancer Institute).
It is clear that with some cancers, for example Kaposi’s sarcoma, a healthy immune system does a good job at controlling them (Figure 7.7.2). This disease, which is caused by the human herpesvirus, is almost never observed in individuals with strong immune systems, such as the young and immunocompetent. Other examples of cancers caused by viruses include liver cancer caused by the hepatitis B virus and cervical cancer caused by the human papilloma virus. As these last two viruses have vaccines available for them, getting vaccinated can help prevent these two types of cancer by stimulating the immune response.
On the other hand, as cancer cells are often able to divide and mutate rapidly, they may escape the immune response, just as certain pathogens such as HIV do. There are three stages in the immune response to many cancers: elimination, equilibrium, and escape. Elimination occurs when the immune response first develops toward tumour-specific antigens specific to the cancer and actively kills most cancer cells, followed by a period of controlled equilibrium during which the remaining cancer cells are held in check. Unfortunately, many cancers mutate, so they no longer express any specific antigens for the immune system to respond to, and a subpopulation of cancer cells escapes the immune response, continuing the disease process.
This fact has led to extensive research in trying to develop ways to enhance the early immune response to eliminate the early cancer and thus prevent a later escape. One method that has shown some success is the use of cancer vaccines, which differ from viral and bacterial vaccines in that they are directed against the cells of one’s own body. Treated cancer cells are injected into cancer patients to enhance their anti-cancer immune response and thereby prolong survival. The immune system has the capability to detect these cancer cells and proliferate faster than the cancer cells do, overwhelming the cancer in a similar way as they do for viruses. Cancer vaccines have been developed for malignant melanoma, a highly fatal skin cancer, and renal (kidney) cell carcinoma. These vaccines are still in the development stages, but some positive and encouraging results have been obtained clinically.
It is tempting to focus on the complexity of the immune system and the problems it causes as a negative. The upside to immunity, however, is so much greater: The benefit of staying alive far outweighs the negatives caused when the system does sometimes go awry. Working on “autopilot,” the immune system helps to maintain your health and kill pathogens. The only time you really miss the immune response is when it is not being effective and illness results, or, as in the extreme case of HIV disease, the immune system is gone completely.
Everyday Connection
How Stress Affects the Immune Response: The Connections between the Immune, Nervous, and Endocrine Systems of the Body
The immune system cannot exist in isolation. After all, it must protect the entire body from infection. Therefore, the immune system is required to interact with other organ systems, sometimes in complex ways. Thirty years of research focusing on the connections between the immune system, the central nervous system, and the endocrine system have led to a new science with the unwieldy name of called psychoneuroimmunology. The physical connections between these systems have been known for centuries: All primary and secondary organs are connected to sympathetic nerves. What is more complex, though, is the interaction of neurotransmitters, hormones, cytokines, and other soluble signalling molecules, and the mechanism of “crosstalk” between the systems, for example white blood cells, including lymphocytes and phagocytes, have receptors for various neurotransmitters released by associated neurons. Additionally, hormones such as cortisol (naturally produced by the adrenal cortex) and prednisone (synthetic) are well known for their abilities to suppress T cell immune mechanisms, hence, their prominent use in medicine as long-term, anti-inflammatory drugs.
One well-established interaction of the immune, nervous, and endocrine systems is the effect of stress on immune health. In the human vertebrate evolutionary past, stress was associated with the fight-or-flight response, largely mediated by the central nervous system and the adrenal medulla. This stress was necessary for survival. The physical action of fighting or running, whichever the animal decides, usually resolves the problem in one way or another. On the other hand, there are no physical actions to resolve most modern-day stresses, including short-term stressors like taking examinations and long-term stressors such as being unemployed or losing a spouse. The effect of stress can be felt by nearly every organ system, and the immune system is no exception (Table 7.7.3).
Table 7.7.3. Effects of stress on body systems
System
Stress-related illness
Integumentary system
Acne, skin rashes, irritation
Nervous system
Headaches, depression, anxiety, irritability, loss of appetite, lack of motivation, reduced mental performance
Muscular and skeletal systems
Muscle and joint pain, neck and shoulder pain
Circulatory system
Increased heart rate, hypertension, increased probability of heart attacks
Digestive system
Indigestion, heartburn, stomach pain, nausea, diarrhoea, constipation, weight gain or loss
Immune System
Depressed ability to fight infections
Male reproductive system
Lowered sperm production, impotence, reduced sexual desire
Female reproductive system
Irregular menstrual cycle, reduced sexual desire
At one time, it was assumed that all types of stress reduced all aspects of the immune response, but the last few decades of research have painted a different picture. First, most short-term stress does not impair the immune system in healthy individuals enough to lead to a greater incidence of diseases. However, older individuals and those with suppressed immune responses due to disease or immunosuppressive drugs may respond even to short-term stressors by getting sicker more often. It has been found that short-term stress diverts the body’s resources towards enhancing innate immune responses, which have the ability to act fast and would seem to help the body prepare better for possible infections associated with the trauma that may result from a fight-or-flight exchange. The diverting of resources away from the adaptive immune response, however, causes its own share of problems in fighting disease.
Chronic stress, unlike short-term stress, may inhibit immune responses even in otherwise healthy adults. The suppression of both innate and adaptive immune responses is clearly associated with increases in some diseases, as seen when individuals lose a spouse or have other long-term stresses, such as taking care of a spouse with a fatal disease or dementia. The new science of psychoneuroimmunology, while still in its relative infancy, has great potential to make exciting advances in our understanding of how the nervous, endocrine, and immune systems have evolved together and communicate with each other.
Section Review
Blood transfusion and organ transplantation both require an understanding of the immune response to prevent medical complications. Blood needs to be typed so that natural antibodies against mismatched blood will not destroy it, causing more harm than good to the recipient. Transplanted organs must be matched by their MHC molecules and, with the use of immunosuppressive drugs, can be successful even if an exact tissue match cannot be made. Another aspect to the immune response is its ability to control and eradicate cancer. Although this has been shown to occur with some rare cancers and those caused by known viruses, the normal immune response to most cancers is not sufficient to control cancer growth. Thus, cancer vaccines designed to enhance these immune responses show promise for certain types of cancer.
8.1 Organs and Structures of the Respiratory System
Learning Objectives
By the end of this section, you will be able to:
List the structures that make up the respiratory system
Describe how the respiratory system processes oxygen and CO2
Compare and contrast the functions of upper respiratory tract with the lower respiratory tract
The major organs of the respiratory system function primarily to provide oxygen to body tissues for cellular respiration, remove the waste product carbon dioxide, and help to maintain acid-base balance. Portions of the respiratory system are also used for non-vital functions, such as sensing odours, speech production, and for straining, such as during childbirth or coughing (Figure 8.1.1).
Figure 8.1.1. Major respiratory structures. The major respiratory structures span the nasal cavity to the diaphragm.
Functionally, the respiratory system can be divided into a conducting zone and a respiratory zone. The conducting zone of the respiratory system includes the organs and structures not directly involved in gas exchange. The gas exchange occurs in the respiratory zone.
Conducting Zone
The major functions of the conducting zone are to provide a route for incoming and outgoing air, remove debris and pathogens from the incoming air, and warm and humidify the incoming air. Several structures within the conducting zone perform other functions as well. The epithelium of the nasal passages, for example, is essential to sensing odours, and the bronchial epithelium that lines the lungs can metabolise some airborne carcinogens.
The Nose and its Adjacent Structures
The major entrance and exit for the respiratory system is through the nose. When discussing the nose, it is helpful to divide it into two major sections: the external nose, and the nasal cavity or internal nose.
The external nose consists of the surface and skeletal structures that result in the outward appearance of the nose and contribute to its numerous functions (Figure 8.1.2). The root is the region of the nose located between the eyebrows. The bridge is the part of the nose that connects the root to the rest of the nose. The dorsum nasi is the length of the nose. The apex is the tip of the nose. On either side of the apex, the nostrils are formed by the alae (singular = ala). An ala is a cartilaginous structure that forms the lateral side of each naris (plural = nares), or nostril opening. The philtrum is the concave surface that connects the apex of the nose to the upper lip.
Figure 8.1.2. Nose. This illustration shows features of the external nose (top) and skeletal features of the nose (bottom).
Underneath the thin skin of the nose are its skeletal features (see Figure 8.1.2, lower illustration). While the root and bridge of the nose consist of bone, the protruding portion of the nose is composed of cartilage. As a result, when looking at a skull, the nose is missing. The nasal bone is one of a pair of bones that lies under the root and bridge of the nose. The nasal bone articulates superiorly with the frontal bone and laterally with the maxillary bones. Septal cartilage is flexible hyaline cartilage connected to the nasal bone, forming the dorsum nasi. The alar cartilage consists of the apex of the nose; it surrounds the naris.
The nares open into the nasal cavity, which is separated into left and right sections by the nasal septum (Figure 8.1.3). The nasal septum is formed anteriorly by a portion of the septal cartilage (the flexible portion you can touch with your fingers) and posteriorly by the perpendicular plate of the ethmoid bone (a cranial bone located just posterior to the nasal bones) and the thin vomer bones (whose name refers to its plough shape). Each lateral wall of the nasal cavity has three bony projections, called the superior, middle, and inferior nasal conchae. The inferior conchae are separate bones, whereas the superior and middle conchae are portions of the ethmoid bone. Conchae serve to increase the surface area of the nasal cavity and to disrupt the flow of air as it enters the nose, causing air to bounce along the epithelium, where it is cleaned and warmed. The conchae and meatuses also conserve water and prevent dehydration of the nasal epithelium by trapping water during exhalation. The floor of the nasal cavity is composed of the palate. The hard palate at the anterior region of the nasal cavity is composed of bone. The soft palate at the posterior portion of the nasal cavity consists of muscle tissue. Air exits the nasal cavities via the internal nares and moves into the pharynx.
Figure 8.1.3. Upper Airway
Several bones that help form the walls of the nasal cavity have air-containing spaces called the paranasal sinuses, which serve to warm and humidify incoming air. Sinuses are lined with a mucosa. Each paranasal sinus is named for its associated bone: frontal sinus, maxillary sinus, sphenoidal sinus, and ethmoidal sinus. The sinuses produce mucus and lighten the weight of the skull.
The nares and anterior portion of the nasal cavities are lined with mucous membranes, containing sebaceous glands and hair follicles that serve to prevent the passage of large debris, such as dirt, through the nasal cavity. An olfactory epithelium used to detect odours is found deeper in the nasal cavity.
The conchae, meatuses, and paranasal sinuses are lined by respiratory epithelium composed of pseudostratified ciliated columnar epithelium (Figure 8.1.4). The epithelium contains goblet cells, one of the specialised, columnar epithelial cells that produce mucus to trap debris. The cilia of the respiratory epithelium help remove the mucus and debris from the nasal cavity with a constant beating motion, sweeping materials towards the throat to be swallowed. Interestingly, cold air slows the movement of the cilia, resulting in accumulation of mucus that may in turn lead to a runny nose during cold weather. This moist epithelium functions to warm and humidify incoming air. Capillaries located just beneath the nasal epithelium warm the air by convection. Serous and mucus-producing cells also secrete the lysozyme enzyme and proteins called defensins, which have antibacterial properties. Immune cells that patrol the connective tissue deep to the respiratory epithelium provide additional protection.
The pharynx is a tube formed by skeletal muscle and lined by mucous membrane that is continuous with that of the nasal cavities (see Figure 8.1.3). The pharynx is divided into three major regions: the nasopharynx, the oropharynx and the laryngopharynx (Figure 8.1.5).
Figure 8.1.5. Divisions of the pharynx. The pharynx is divided into three regions: the nasopharynx, the oropharynx and the laryngopharynx.
The nasopharynx is flanked by the conchae of the nasal cavity, and it serves only as an airway. At the top of the nasopharynx are the pharyngeal tonsils. A pharyngeal tonsil, also called an adenoid, is an aggregate of lymphoid reticular tissue similar to a lymph node that lies at the superior portion of the nasopharynx. The function of the pharyngeal tonsil is not well understood, but it contains a rich supply of lymphocytes and is covered with ciliated epithelium that traps and destroys invading pathogens that enter during inhalation. The pharyngeal tonsils are large in children, but interestingly, tend to regress with age and may even disappear. The uvula is a small bulbous, teardrop-shaped structure located at the apex of the soft palate. Both the uvula and soft palate move like a pendulum during swallowing, swinging upward to close off the nasopharynx to prevent ingested materials from entering the nasal cavity. In addition, auditory (Eustachian or pharyngotympanic) tubes that connect to each middle ear cavity open into the nasopharynx. This connection is why colds often lead to ear infections.
The oropharynx is a passageway for both air and food. The oropharynx is bordered superiorly by the nasopharynx and anteriorly by the oral cavity. The fauces is the opening at the connection between the oral cavity and the oropharynx. As the nasopharynx becomes the oropharynx, the epithelium changes from pseudostratified ciliated columnar epithelium to stratified squamous epithelium. The oropharynx contains two distinct sets of tonsils, the palatine and lingual tonsils. A palatine tonsil is one of a pair of structures located laterally in the oropharynx in the area of the fauces. The lingual tonsil is located at the base of the tongue. Similar to the pharyngeal tonsil, the palatine and lingual tonsils are composed of lymphoid tissue, and trap and destroy pathogens entering the body through the oral or nasal cavities.
The laryngopharynx is inferior to the oropharynx and posterior to the larynx. It continues the route for ingested material and air until its inferior end, where the digestive and respiratory systems diverge. The stratified squamous epithelium of the oropharynx is continuous with the laryngopharynx. Anteriorly, the laryngopharynx opens into the larynx, whereas posteriorly, it enters the oesophagus.
Larynx
The larynx is a cartilaginous structure inferior to the laryngopharynx that connects the pharynx to the trachea and helps regulate the volume of air that enters and leaves the lungs (Figure 8.1.6). The structure of the larynx is formed by several pieces of cartilage. Three large cartilage pieces—the thyroid cartilage (anterior), epiglottis (superior), and cricoid cartilage (inferior)—form the major structure of the larynx. The thyroid cartilage is the largest piece of cartilage that makes up the larynx. The thyroid cartilage consists of the laryngeal prominence, or “Adam’s apple,” which tends to be more prominent in males. The thick cricoid cartilage forms a ring, with a wide posterior region and a thinner anterior region. Three smaller, paired cartilages—the arytenoids, corniculates, and cuneiforms—attach to the epiglottis and the vocal cords and muscle that help move the vocal cords to produce speech.
Figure 8.1.6. Larynx. The larynx extends from the laryngopharynx and the hyoid bone to the trachea.
The epiglottis, attached to the thyroid cartilage, is a very flexible piece of elastic cartilage that covers the opening of the trachea (see Figure 8.1.3). When in the “closed” position, the unattached end of the epiglottis rests on the glottis. The glottis is composed of the vestibular folds, the true vocal cords, and the space between these folds (Figure 8.1.7). A vestibular fold, or false vocal cord, is one of a pair of folded sections of mucous membrane. A true vocal cord is one of the white, membranous folds attached by muscle to the thyroid and arytenoid cartilages of the larynx on their outer edges. The inner edges of the true vocal cords are free, allowing oscillation to produce sound. The size of the membranous folds of the true vocal cords differs between individuals, producing voices with different pitch ranges. Folds in males tend to be larger than those in females, which create a deeper voice. The act of swallowing causes the pharynx and larynx to lift upward, allowing the pharynx to expand and the epiglottis of the larynx to swing downward, closing the opening to the trachea. These movements produce a larger area for food to pass through, while preventing food and beverages from entering the trachea.
Figure 8.1.7. Vocal cords. The true vocal cords and vestibular folds of the larynx are viewed inferiorly from the laryngopharynx.
Continuous with the laryngopharynx, the superior portion of the larynx is lined with stratified squamous epithelium, transitioning into pseudostratified ciliated columnar epithelium that contains goblet cells. Similar to the nasal cavity and nasopharynx, this specialised epithelium produces mucus to trap debris and pathogens as they enter the trachea. The cilia beat the mucus upward towards the laryngopharynx, where it can be swallowed down the oesophagus.
Trachea
The trachea (windpipe) extends from the larynx toward the lungs (Figure 8.1.8a). The trachea is formed by 16 to 20 stacked, C-shaped pieces of hyaline cartilage that are connected by dense connective tissue. The trachealis muscle and elastic connective tissue together form the fibroelastic membrane, a flexible membrane that closes the posterior surface of the trachea, connecting the C-shaped cartilages. The fibroelastic membrane allows the trachea to stretch and expand slightly during inhalation and exhalation, whereas the rings of cartilage provide structural support and prevent the trachea from collapsing. In addition, the trachealis muscle can be contracted to force air through the trachea during exhalation. The trachea is lined with pseudostratified ciliated columnar epithelium, which is continuous with the larynx. The oesophagus borders the trachea posteriorly.
The trachea branches into the right and left primary bronchi at the carina. These bronchi are also lined by pseudostratified ciliated columnar epithelium containing mucus-producing goblet cells (Figure 8.1.8b). The carina is a raised structure that contains specialised nervous tissue that induces violent coughing if a foreign body, such as food, is present. Rings of cartilage, similar to those of the trachea, support the structure of the bronchi and prevent their collapse. The primary bronchi enter the lungs at the hilum, a concave region where blood vessels, lymphatic vessels, and nerves also enter the lungs. The bronchi continue to branch into bronchial a tree. A bronchial tree (or respiratory tree) is the collective term used for these multiple-branched bronchi. The main function of the bronchi, like other conducting zone structures, is to provide a passageway for air to move into and out of each lung. In addition, the mucous membrane traps debris and pathogens.
A bronchiole branches from the tertiary bronchi. Bronchioles, which are about 1 mm in diameter, further branch until they become the tiny terminal bronchioles, which lead to the structures of gas exchange. There are more than 1000 terminal bronchioles in each lung. The muscular walls of the bronchioles do not contain cartilage like those of the bronchi. This muscular wall can change the size of the tubing to increase or decrease airflow through the tube.
Respiratory Zone
In contrast to the conducting zone, the respiratory zone includes structures that are directly involved in gas exchange. The respiratory zone begins where the terminal bronchioles join a respiratory bronchiole, the smallest type of bronchiole (Figure 8.1.9), which then leads to an alveolar duct, opening into a cluster of alveoli.
Figure 8.1.9. Respiratory zone. Bronchioles lead to alveolar sacs in the respiratory zone, where gas exchange occurs.
Alveoli
An alveolar duct is a tube composed of smooth muscle and connective tissue, which opens into a cluster of alveoli. An alveolus is one of the many small, grape-like sacs that are attached to the alveolar ducts.
An alveolar sac is a cluster of many individual alveoli that are responsible for gas exchange. An alveolus is approximately 200 μm in diameter with elastic walls that allow the alveolus to stretch during air intake, which greatly increases the surface area available for gas exchange. Alveoli are connected to their neighbours by alveolar pores, which help maintain equal air pressure throughout the alveoli and lung (Figure 8.1.10).
The alveolar wall consists of three major cell types: type I alveolar cells, type II alveolar cells, and alveolar macrophages. A type I alveolar cell is a squamous epithelial cell of the alveoli, which constitute up to 97 percent of the alveolar surface area. These cells are about 25 nm thick and are highly permeable to gases. A type II alveolar cell is interspersed among the type I cells and secretes pulmonary surfactant, a substance composed of phospholipids and proteins that reduces the surface tension of the alveoli. Roaming around the alveolar wall is the alveolar macrophage, a phagocytic cell of the immune system that removes debris and pathogens that have reached the alveoli.
The simple squamous epithelium formed by type I alveolar cells is attached to a thin, elastic basement membrane. This epithelium is extremely thin and borders the endothelial membrane of capillaries. Taken together, the alveoli and capillary membranes form a respiratory membrane that is approximately 0.5 mm thick. The respiratory membrane allows gases to cross by simple diffusion, allowing oxygen to be picked up by the blood for transport and CO2 to be released into the air of the alveoli.
Diseases of the Respiratory System: Asthma
Asthma is common condition that affects the lungs in both adults and children. Almost 3 million Australians suffer from asthma (over 10% of the population). In United States 18.7 million adults (8.2%) and 7 million children (9.4%) suffer from asthma. In addition, asthma is the most frequent cause of hospitalisation in children.
Asthma is a chronic disease characterised by inflammation and oedema of the airway, and bronchospasms (that is, constriction of the bronchioles), which can inhibit air from entering the lungs. In addition, excessive mucus secretion can occur, which further contributes to airway occlusion (Figure 8.1.11). Cells of the immune system, such as eosinophils and mononuclear cells, may also be involved in infiltrating the walls of the bronchi and bronchioles. In 2016, a high pollen count mixed with meteorological conditions resulted in a thunderstorm-asthma event that resulted with over 3,500 people to be hospitalised in Melbourne, Australia.
Bronchospasms occur periodically and lead to an “asthma attack.” An attack may be triggered by environmental factors such as dust, pollen, pet hair, or dander, changes in the weather, mould, tobacco smoke, wood smoke, and respiratory infections, or by exercise and stress.
Figure 8.1.11. Normal and bronchial asthma tissues. (a) Normal lung tissue does not have the characteristics of lung tissue during (b) an asthma attack, which include thickened mucosa, increased mucus-producing goblet cells, and eosinophil infiltrates.
Symptoms of an asthma attack involve coughing, shortness of breath, wheezing, and tightness of the chest. Symptoms of a severe asthma attack that requires immediate medical attention would include difficulty breathing that results in blue (cyanotic) lips or face, confusion, drowsiness, a rapid pulse, sweating, and severe anxiety. The severity of the condition, frequency of attacks, and identified triggers influence the type of medication that an individual may require. Longer-term treatments are used for those with more severe asthma. Short-term, fast-acting drugs that are used to treat an asthma attack are typically administered via an inhaler. For young children or individuals who have difficulty using an inhaler, asthma medications can be administered via a nebuliser.
In many cases, the underlying cause of the condition is unknown. However, recent research has demonstrated that certain viruses, such as human rhinovirus C (HRVC), and the bacteria Mycoplasma pneumoniae and Chlamydia pneumoniae that are contracted in infancy or early childhood, may contribute to the development of many cases of asthma.
Section Review
The respiratory system is responsible for obtaining oxygen and getting rid of carbon dioxide and aiding in speech production and in sensing odours. From a functional perspective, the respiratory system can be divided into two major areas: the conducting zone and the respiratory zone. The conducting zone consists of all the structures that provide passageways for air to travel into and out of the lungs: the nasal cavity, pharynx, trachea, bronchi, and most bronchioles. The nasal passages contain the conchae and meatuses that expand the surface area of the cavity, which helps to warm and humidify incoming air, while removing debris and pathogens. The pharynx is composed of three major sections: the nasopharynx, which is continuous with the nasal cavity; the oropharynx, which borders the nasopharynx and the oral cavity; and the laryngopharynx, which borders the oropharynx, trachea, and oesophagus. The respiratory zone includes the structures of the lung that are directly involved in gas exchange: the terminal bronchioles and alveoli.
The lining of the conducting zone is composed mostly of pseudostratified ciliated columnar epithelium with goblet cells. The mucus traps pathogens and debris, whereas beating cilia move the mucus superiorly toward the throat, where it is swallowed. As the bronchioles become smaller and smaller, and nearer the alveoli, the epithelium thins and is simple squamous epithelium in the alveoli. The endothelium of the surrounding capillaries, together with the alveolar epithelium, forms the respiratory membrane. This is a blood-air barrier through which gas exchange occurs by simple diffusion.
Summarise the blood flow pattern associated with the lungs
Outline the anatomy of the blood supply to the lungs
Describe the pleura of the lungs and their function
A major organ of the respiratory system, each lung houses structures of both the conducting and respiratory zones. The main function of the lungs is to perform the exchange of oxygen and carbon dioxide with air from the atmosphere. To this end, the lungs exchange respiratory gases across a very large epithelial surface area—about 70 square metres—that is highly permeable to gases.
Gross Anatomy of the Lungs
The lungs are pyramid-shaped, paired organs that are connected to the trachea by the right and left bronchi; on the inferior surface, the lungs are bordered by the diaphragm. The diaphragm is the flat, dome-shaped muscle located at the base of the lungs and thoracic cavity. The lungs are enclosed by the pleurae, which are attached to the mediastinum. The right lung is shorter and wider than the left lung, and the left lung occupies a smaller volume than the right. The cardiac notch is an indentation on the surface of the left lung, and it allows space for the heart (Figure 8.2.1). The apex of the lung is the superior region, whereas the base is the opposite region near the diaphragm. The costal surface of the lung borders the ribs. The mediastinal surface faces the midline.
Figure 8.2.1. Gross anatomy of the lungs.
Each lung is composed of smaller units called lobes. Fissures separate these lobes from each other. The right lung consists of three lobes: the superior, middle, and inferior lobes. The left lung consists of two lobes: the superior and inferior lobes. A bronchopulmonary segment is a division of a lobe, and each lobe houses multiple bronchopulmonary segments. Each segment receives air from its own tertiary bronchus and is supplied with blood by its own artery. Some diseases of the lungs typically affect one or more bronchopulmonary segments, and in some cases, the diseased segments can be surgically removed with little influence on neighbouring segments. A pulmonary lobule is a subdivision formed as the bronchi branch into bronchioles. Each lobule receives its own large bronchiole that has multiple branches. An interlobular septum is a wall, composed of connective tissue, which separates lobules from one another.
Blood Supply and Nervous Innervation of the Lungs
The blood supply of the lungs plays an important role in gas exchange and serves as a transport system for gases throughout the body. In addition, innervation by the both the parasympathetic and sympathetic nervous systems provides an important level of control through dilation and constriction of the airway.
Blood Supply
The major function of the lungs is to perform gas exchange, which requires blood from the pulmonary circulation. This blood supply contains deoxygenated blood and travels to the lungs where erythrocytes, also known as red blood cells, pick up oxygen to be transported to tissues throughout the body. The pulmonary artery is an artery that arises from the pulmonary trunk and carries deoxygenated, arterial blood to the alveoli. The pulmonary artery branches multiple times as it follows the bronchi, and each branch becomes progressively smaller in diameter. One arteriole and an accompanying venule supply and drain one pulmonary lobule. As they near the alveoli, the pulmonary arteries become the pulmonary capillary network. The pulmonary capillary network consists of tiny vessels with very thin walls that lack smooth muscle fibres. The capillaries branch and follow the bronchioles and structure of the alveoli. It is at this point that the capillary wall meets the alveolar wall, creating the respiratory membrane. Once the blood is oxygenated, it drains from the alveoli by way of multiple pulmonary veins, which exit the lungs through the hilum.
Nervous Innervation
Dilation and constriction of the airway are achieved through nervous control by the parasympathetic and sympathetic nervous systems. The parasympathetic system causes bronchoconstriction, whereas the sympathetic nervous system stimulates bronchodilation. Reflexes such as coughing, and the ability of the lungs to regulate oxygen and carbon dioxide levels, also result from this autonomic nervous system control. Sensory nerve fibres arise from the vagus nerve, and from the second to fifth thoracic ganglia. The pulmonary plexus is a region on the lung root formed by the entrance of the nerves at the hilum. The nerves then follow the bronchi in the lungs and branch to innervate muscle fibres, glands, and blood vessels.
Pleurae of the Lungs
Each lung is enclosed within a cavity that is surrounded by the pleura. The pleura (plural = pleurae) is a serous membrane that surrounds the lung. The right and left pleurae, which enclose the right and left lungs, respectively, are separated by the mediastinum. The pleurae consist of two layers. The visceral pleura is the layer that is superficial to the lungs, and extends into and lines the lung fissures (Figure 8.2.2). In contrast, the parietal pleura is the outer layer that connects to the thoracic wall, the mediastinum, and the diaphragm. The visceral and parietal pleurae connect to each other at the hilum. The pleural cavity is the space between the visceral and parietal layers.
Figure 8.2.2. Parietal and visceral pleurae of the lungs.
The pleurae perform two major functions: They produce pleural fluid and create cavities that separate the major organs. Pleural fluid is secreted by mesothelial cells from both pleural layers and acts to lubricate their surfaces. This lubrication reduces friction between the two layers to prevent trauma during breathing and creates surface tension that helps maintain the position of the lungs against the thoracic wall. This adhesive characteristic of the pleural fluid causes the lungs to enlarge when the thoracic wall expands during ventilation, allowing the lungs to fill with air. The pleurae also create a division between major organs that prevents interference due to the movement of the organs, while preventing the spread of infection.
Everyday Connection
The Effects of Second-Hand Tobacco Smoke
The burning of a tobacco cigarette creates multiple chemical compounds that are released through mainstream smoke, which is inhaled by the smoker, and through side stream smoke, which is the smoke that is given off by the burning cigarette. Second-hand smoke, which is a combination of side stream smoke and the mainstream smoke that is exhaled by the smoker, has been demonstrated by numerous scientific studies to cause disease. At least 40 chemicals in side stream smoke have been identified that negatively impact human health, leading to the development of cancer or other conditions, such as immune system dysfunction, liver toxicity, cardiac arrhythmias, pulmonary oedema, and neurological dysfunction. Furthermore, second-hand smoke has been found to harbor at least 250 compounds that are known to be toxic, carcinogenic, or both. Some major classes of carcinogens in second-hand smoke are polyaromatic hydrocarbons (PAHs), N-nitrosamines, aromatic amines, formaldehyde, and acetaldehyde.
Tobacco and second-hand smoke are considered to be carcinogenic. Exposure to second-hand smoke can cause lung cancer in individuals who are not tobacco users themselves. It is estimated that the risk of developing lung cancer is increased by up to 30 percent in non-smokers who live with an individual who smokes in the house, as compared to non-smokers who are not regularly exposed to second-hand smoke. Children are especially affected by second-hand smoke. Children who live with an individual who smokes inside the home have a larger number of lower respiratory infections, which are associated with hospitalisations, and higher risk of sudden infant death syndrome (SIDS). Second-hand smoke in the home has also been linked to a greater number of ear infections in children, as well as worsening symptoms of asthma.
Section Review
The lungs are the major organs of the respiratory system and are responsible for performing gas exchange. The lungs are paired and separated into lobes; The left lung consists of two lobes, whereas the right lung consists of three lobes. Blood circulation is very important, as blood is required to transport oxygen from the lungs to other tissues throughout the body. The function of the pulmonary circulation is to aid in gas exchange. The pulmonary artery provides deoxygenated blood to the capillaries that form respiratory membranes with the alveoli, and the pulmonary veins return newly oxygenated blood to the heart for further transport throughout the body. The lungs are innervated by the parasympathetic and sympathetic nervous systems, which coordinate the bronchodilation and bronchoconstriction of the airways. The lungs are enclosed by the pleura, a membrane that is composed of visceral and parietal pleural layers. The space between these two layers is called the pleural cavity. The mesothelial cells of the pleural membrane create pleural fluid, which serves as both a lubricant (to reduce friction during breathing) and as an adhesive to adhere the lungs to the thoracic wall (to facilitate movement of the lungs during ventilation).
Discuss how pressure, volume, and resistance are related
List the steps involved in pulmonary ventilation
Discuss the physical factors related to breathing
Discuss the meaning of respiratory volume and capacities
Define respiratory rate
Outline the mechanisms behind the control of breathing
Describe the respiratory centres of the medulla oblongata
Describe the respiratory centres of the pons
Discuss factors that can influence the respiratory rate
Pulmonary ventilation is the act of breathing, which can be described as the movement of air into and out of the lungs. The major mechanisms that drive pulmonary ventilation are atmospheric pressure (Patm); the air pressure within the alveoli, called alveolar pressure (Palv); and the pressure within the pleural cavity, called intrapleural pressure (Pip).
Mechanisms of Breathing
The alveolar and intrapleural pressures are dependent on certain physical features of the lung. However, the ability to breathe—to have air enter the lungs during inspiration and air leave the lungs during expiration—is dependent on the air pressure of the atmosphere and the air pressure within the lungs.
Pressure Relationships
Inspiration (or inhalation) and expiration (or exhalation) are dependent on the differences in pressure between the atmosphere and the lungs. In a gas, pressure is a force created by the movement of gas molecules that are confined. For example, a certain number of gas molecules in a two-litre container has more room than the same number of gas molecules in a one-litre container (Figure 8.3.1). In this case, the force exerted by the movement of the gas molecules against the walls of the two-litre container is lower than the force exerted by the gas molecules in the one-litre container. Therefore, the pressure is lower in the two-litre container and higher in the one-litre container. At a constant temperature, changing the volume occupied by the gas changes the pressure, as does changing the number of gas molecules. Boyle’s law describes the relationship between volume and pressure in a gas at a constant temperature. Boyle discovered that the pressure of a gas is inversely proportional to its volume: If volume increases, pressure decreases. Likewise, if volume decreases, pressure increases. Pressure and volume are inversely related (P = k/V). Therefore, the pressure in the one-litre container (one-half the volume of the two-litre container) would be twice the pressure in the two-litre container. Boyle’s law is expressed by the following formula:
P1V1 = P2V2
In this formula, P1 represents the initial pressure and V1 represents the initial volume, whereas the final pressure and volume are represented by P2 and V2, respectively. If the two- and one-litre containers were connected by a tube and the volume of one of the containers were changed, then the gases would move from higher pressure (lower volume) to lower pressure (higher volume).
Figure 8.3.1. Boyle’s law. In a gas, pressure increases as volume decreases.
Pulmonary ventilation is dependent on three types of pressure: atmospheric, intra-alveolar, and interpleural. Atmospheric pressure is the amount of force that is exerted by gases in the air surrounding any given surface, such as the body. Atmospheric pressure can be expressed in terms of the unit atmosphere, abbreviated atm, or in millimetres of mercury (mm Hg). One atm is equal to 760 mm Hg, which is the atmospheric pressure at sea level. Typically, for respiration, other pressure values are discussed in relation to atmospheric pressure. Therefore, negative pressure is pressure lower than the atmospheric pressure, whereas positive pressure is pressure that it is greater than the atmospheric pressure. A pressure that is equal to the atmospheric pressure is expressed as zero.
Intra-alveolar pressure is the pressure of the air within the alveoli, which changes during the different phases of breathing (Figure 8.3.2). Because the alveoli are connected to the atmosphere via the tubing of the airways (similar to the two- and one-litre containers in the example above), the interpulmonary pressure of the alveoli always equalises with the atmospheric pressure.
Figure 8.3.2. Intrapulmonary and intrapleural pressure relationships. Alveolar pressure changes during the different phases of the cycle. It equalises at 760 mm Hg but does not remain at 760 mm Hg.
Intrapleural pressure is the pressure of the air within the pleural cavity, between the visceral and parietal pleurae. Similar to intra-alveolar pressure, intrapleural pressure also changes during the different phases of breathing. However, due to certain characteristics of the lungs, the intrapleural pressure is always lower than, or negative to, the intra-alveolar pressure (and therefore also to atmospheric pressure). Although it fluctuates during inspiration and expiration, intrapleural pressure remains approximately –4 mm Hg throughout the breathing cycle.
Competing forces within the thorax cause the formation of the negative intrapleural pressure. One of these forces relates to the elasticity of the lungs themselves—elastic tissue pulls the lungs inward, away from the thoracic wall. Surface tension of alveolar fluid, which is mostly water, also creates an inward pull of the lung tissue. This inward tension from the lungs is countered by opposing forces from the pleural fluid and thoracic wall. Surface tension within the pleural cavity pulls the lungs outward. Too much or too little pleural fluid would hinder the creation of the negative intrapleural pressure; therefore, the level must be closely monitored by the mesothelial cells and drained by the lymphatic system. Since the parietal pleura is attached to the thoracic wall, the natural elasticity of the chest wall opposes the inward pull of the lungs. Ultimately, the outward pull is slightly greater than the inward pull, creating the –4 mm Hg intrapleural pressure relative to the intra-alveolar pressure. Transpulmonary pressure is the difference between the intrapleural and intra-alveolar pressures, and it determines the size of the lungs. A higher transpulmonary pressure corresponds to a larger lung.
Physical Factors Affecting Ventilation
In addition to the differences in pressures, breathing is also dependent upon the contraction and relaxation of muscle fibres of both the diaphragm and thorax. The lungs themselves are passive during breathing, meaning they are not involved in creating the movement that helps inspiration and expiration. This is because of the adhesive nature of the pleural fluid, which allows the lungs to be pulled outward when the thoracic wall moves during inspiration. The recoil of the thoracic wall during expiration causes compression of the lungs. Contraction and relaxation of the diaphragm and intercostals muscles (found between the ribs) cause most of the pressure changes that result in inspiration and expiration. These muscle movements and subsequent pressure changes cause air to either rush in or be forced out of the lungs.
Other characteristics of the lungs influence the effort that must be expended to ventilate. Resistance is a force that slows motion, in this case, the flow of gases. The size of the airway is the primary factor affecting resistance. A small tubular diameter forces air through a smaller space, causing more collisions of air molecules with the walls of the airways. The following formula helps to describe the relationship between airway resistance and pressure changes:
F = ∆P / R
As noted earlier, there is surface tension within the alveoli caused by water present in the lining of the alveoli. This surface tension tends to inhibit expansion of the alveoli. However, pulmonary surfactant secreted by type II alveolar cells mixes with that water and helps reduce this surface tension. Without pulmonary surfactant, the alveoli would collapse during expiration.
Thoracic wall compliance is the ability of the thoracic wall to stretch while under pressure. This can also affect the effort expended in the process of breathing. In order for inspiration to occur, the thoracic cavity must expand. The expansion of the thoracic cavity directly influences the capacity of the lungs to expand. If the tissues of the thoracic wall are not very compliant, it will be difficult to expand the thorax to increase the size of the lungs.
Pulmonary Ventilation
The difference in pressures drives pulmonary ventilation because air flows down a pressure gradient, that is, air flows from an area of higher pressure to an area of lower pressure. Air flows into the lungs largely due to a difference in pressure; atmospheric pressure is greater than intra-alveolar pressure, and intra-alveolar pressure is greater than intrapleural pressure. Air flows out of the lungs during expiration based on the same principle; pressure within the lungs becomes greater than the atmospheric pressure.
Pulmonary ventilation comprises two major steps: inspiration and expiration. Inspiration is the process that causes air to enter the lungs, and expiration is the process that causes air to leave the lungs (Figure 8.3.3). A respiratory cycle is one sequence of inspiration and expiration. In general, two muscle groups are used during normal inspiration: the diaphragm and the external intercostal muscles. Additional muscles can be used if a bigger breath is required. When the diaphragm contracts, it moves inferiorly toward the abdominal cavity, creating a larger thoracic cavity and more space for the lungs. Contraction of the external intercostal muscles moves the ribs upward and outward, causing the rib cage to expand, which increases the volume of the thoracic cavity. Due to the adhesive force of the pleural fluid, the expansion of the thoracic cavity forces the lungs to stretch and expand as well. This increase in volume leads to a decrease in intra-alveolar pressure, creating a pressure lower than atmospheric pressure. As a result, a pressure gradient is created that drives air into the lungs.
Figure 8.3.3. Inspiration and expiration. Inspiration and expiration occur due to the expansion and contraction of the thoracic cavity, respectively.
The process of normal expiration is passive, meaning that energy is not required to push air out of the lungs. Instead, the elasticity of the lung tissue causes the lung to recoil, as the diaphragm and intercostal muscles relax following inspiration. In turn, the thoracic cavity and lungs decrease in volume, causing an increase in interpulmonary pressure. The interpulmonary pressure rises above atmospheric pressure, creating a pressure gradient that causes air to leave the lungs.
There are different types, or modes, of breathing that require a slightly different process to allow inspiration and expiration. Quiet breathing, also known as eupnoea, is a mode of breathing that occurs at rest and does not require the cognitive thought of the individual. During quiet breathing, the diaphragm and external intercostals must contract.
A deep breath, called diaphragmatic breathing, requires the diaphragm to contract. As the diaphragm relaxes, air passively leaves the lungs. A shallow breath, called costal breathing, requires contraction of the intercostal muscles. As the intercostal muscles relax, air passively leaves the lungs.
In contrast, forced breathing, also known as hyperpnea, is a mode of breathing that can occur during exercise or actions that require the active manipulation of breathing, such as singing. During forced breathing, inspiration and expiration both occur due to muscle contractions. In addition to the contraction of the diaphragm and intercostal muscles, other accessory muscles must also contract. During forced inspiration, muscles of the neck, including the scalenes, contract and lift the thoracic wall, increasing lung volume. During forced expiration, accessory muscles of the abdomen, including the obliques, contract, forcing abdominal organs upward against the diaphragm. This helps to push the diaphragm further into the thorax, pushing more air out. In addition, accessory muscles (primarily the internal intercostals) help to compress the rib cage, which also reduces the volume of the thoracic cavity.
Respiratory Volumes and Capacities
Respiratory volume is the term used for various volumes of air moved by or associated with the lungs at a given point in the respiratory cycle. There are four major types of respiratory volumes: tidal, residual, inspiratory reserve, and expiratory reserve (Figure 8.3.4). Tidal volume (TV) is the amount of air that normally enters the lungs during quiet breathing, which is about 500 millilitres. Expiratory reserve volume (ERV) is the amount of air you can forcefully exhale past a normal tidal expiration, up to 1,200 millilitres for men. Inspiratory reserve volume (IRV) is produced by a deep inhalation, past a tidal inspiration. This is the extra volume that can be brought into the lungs during a forced inspiration. Residual volume (RV) is the air left in the lungs if you exhale as much air as possible. The residual volume makes breathing easier by preventing the alveoli from collapsing. Respiratory volume is dependent on a variety of factors, and measuring the different types of respiratory volumes can provide important clues about a person’s respiratory health (Figure 8.3.5).
Figure 8.3.4. Respiratory volumes and capacities. These two graphs show (a) respiratory volumes and (b) the combination of volumes that results in respiratory capacity.Figure 8.3.5. Pulmonary function testing.
Respiratory capacity is the combination of two or more selected volumes, which further describes the amount of air in the lungs during a given time. For example, total lung capacity (TLC) is the sum of all of the lung volumes (TV, ERV, IRV, and RV), which represents the total amount of air a person can hold in the lungs after a forceful inhalation. TLC is about 6,000 mL air for men, and about 4,200 mL for women. Vital capacity (VC) is the amount of air a person can move into or out of his or her lungs and is the sum of all of the volumes except residual volume (TV, ERV, and IRV), which is between 4,000 and 5,000 millilitres. Inspiratory capacity (IC) is the maximum amount of air that can be inhaled past a normal tidal expiration, is the sum of the tidal volume and inspiratory reserve volume. On the other hand, the functional residual capacity (FRC) is the amount of air that remains in the lung after a normal tidal expiration; it is the sum of expiratory reserve volume and residual volume (see Figure 8.3.4).
In addition to the air that creates respiratory volumes, the respiratory system also contains anatomical dead space, which is air that is present in the airway that never reaches the alveoli and therefore never participates in gas exchange. Alveolar dead space involves air found within alveoli that are unable to function, such as those affected by disease or abnormal blood flow. Total dead space is the anatomical dead space and alveolar dead space together and represents all of the air in the respiratory system that is not being used in the gas exchange process.
Respiratory Rate and Control of Ventilation
Breathing usually occurs without thought, although at times you can consciously control it, such as when you swim under water, sing a song, or blow bubbles. The respiratory rate is the total number of breaths, or respiratory cycles, that occur each minute. Respiratory rate can be an important indicator of disease, as the rate may increase or decrease during an illness or in a disease condition. The respiratory rate is controlled by the respiratory centre located within the medulla oblongata in the brain, which responds primarily to changes in carbon dioxide, oxygen, and pH levels in the blood.
The normal respiratory rate of a child decreases from birth to adolescence. A child under 1 year of age has a normal respiratory rate between 30 and 60 breaths per minute, but by the time a child is about 10 years old, the normal rate is closer to 18 to 30. By adolescence, the normal respiratory rate is similar to that of adults, 12 to 18 breaths per minute.
Ventilation Control Centres
The control of ventilation is a complex interplay of multiple regions in the brain that signal the muscles used in pulmonary ventilation to contract (Table 8.3.1). The result is typically a rhythmic, consistent ventilation rate that provides the body with sufficient amounts of oxygen, while adequately removing carbon dioxide.
Table 8.3.1. Summary of ventilation regulation
System Component
Function
Medullary respiratory centre
Sets the basic rhythm of breathing
Ventral respiratory group (VRG)
Generates the breathing rhythm and integrates data coming into the medulla
Dorsal respiratory group (DRG)
Integrates input from the stretch receptors and the chemoreceptors in the periphery
Pontine respiratory group (PRG)
Influences and modifies the medulla oblongata’s functions
Aortic body
Monitors blood PCO2, PO2, and pH
Carotid body
Monitors blood PCO2, PO2, and pH
Hypothalamus
Monitors emotional state and body temperature
Cortical areas of the brain
Control voluntary breathing
Proprioceptors
Sends impulses regarding joint and muscle movements
Pulmonary irritant reflexes
Protects the respiratory zones of the system from foreign material
Inflation reflex
Protects the lungs from over-inflating
Neurons that innervate the muscles of the respiratory system are responsible for controlling and regulating pulmonary ventilation. The major brain centres involved in pulmonary ventilation are the medulla oblongata and the pontine respiratory group (Figure 8.3.6).
Figure 8.3.6. Respiratory centres of the brain.
The medulla oblongata contains the dorsal respiratory group (DRG) and the ventral respiratory group (VRG). The DRG is involved in maintaining a constant breathing rhythm by stimulating the diaphragm and intercostal muscles to contract, resulting in inspiration. When activity in the DRG ceases, it no longer stimulates the diaphragm and intercostals to contract, allowing them to relax, resulting in expiration. The VRG is involved in forced breathing, as the neurons in the VRG stimulate the accessory muscles involved in forced breathing to contract, resulting in forced inspiration. The VRG also stimulates the accessory muscles involved in forced expiration to contract.
The second respiratory centre of the brain is located within the pons, called the pontine respiratory group, and consists of the apneustic and pneumotaxic centre. The apneustic centre is a double cluster of neuronal cell bodies that stimulate neurons in the DRG, controlling the depth of inspiration, particularly for deep breathing. The pneumotaxic centre is a network of neurons that inhibits the activity of neurons in the DRG, allowing relaxation after inspiration, and thus controlling the overall rate.
Factors that Affect the Rate and Depth of Respiration
The respiratory rate and the depth of inspiration are regulated by the medulla oblongata and pons; however, these regions of the brain do so in response to systemic stimuli. It is a dose-response, positive-feedback relationship in which the greater the stimulus, the greater the response. Thus, increasing stimuli results in forced breathing. Multiple systemic factors are involved in stimulating the brain to produce pulmonary ventilation.
The major factor that stimulates the medulla oblongata and pons to produce respiration is surprisingly not oxygen concentration, but rather the concentration of carbon dioxide in the blood. As you recall, carbon dioxide is a waste product of cellular respiration and can be toxic. Concentrations of chemicals are sensed by chemoreceptors. A central chemoreceptor is one of the specialised receptors that are located in the brain and brainstem, whereas a peripheral chemoreceptor is one of the specialised receptors located in the carotid arteries and aortic arch. Concentration changes in certain substances, such as carbon dioxide or hydrogen ions, stimulate these receptors, which in turn signal the respiration centres of the brain. In the case of carbon dioxide, as the concentration of CO2 in the blood increases, it readily diffuses across the blood-brain barrier, where it collects in the extracellular fluid. As will be explained in more detail later, increased carbon dioxide levels lead to increased levels of hydrogen ions, decreasing pH. The increase in hydrogen ions in the brain triggers the central chemoreceptors to stimulate the respiratory centres to initiate contraction of the diaphragm and intercostal muscles. As a result, the rate and depth of respiration increase, allowing more carbon dioxide to be expelled, which brings more air into and out of the lungs promoting a reduction in the blood levels of carbon dioxide, and therefore hydrogen ions, in the blood. In contrast, low levels of carbon dioxide in the blood cause low levels of hydrogen ions in the brain, leading to a decrease in the rate and depth of pulmonary ventilation, producing shallow, slow breathing.
Another factor involved in influencing the respiratory activity of the brain is systemic arterial concentrations of hydrogen ions. Increasing carbon dioxide levels can lead to increased H+ levels, as mentioned above, as well as other metabolic activities, such as lactic acid accumulation after strenuous exercise. Peripheral chemoreceptors of the aortic arch and carotid arteries sense arterial levels of hydrogen ions. When peripheral chemoreceptors sense decreasing, or more acidic, pH levels, they stimulate an increase in ventilation to remove carbon dioxide from the blood at a quicker rate. Removal of carbon dioxide from the blood helps to reduce hydrogen ions, thus increasing systemic pH.
Blood levels of oxygen are also important in influencing respiratory rate. The peripheral chemoreceptors are responsible for sensing large changes in blood oxygen levels. If blood oxygen levels become quite low—about 60 mm Hg or less—then peripheral chemoreceptors stimulate an increase in respiratory activity. The chemoreceptors are only able to sense dissolved oxygen molecules, not the oxygen that is bound to haemoglobin. As you recall, the majority of oxygen is bound by haemoglobin; when dissolved levels of oxygen drop, haemoglobin releases oxygen. Therefore, a large drop in oxygen levels is required to stimulate the chemoreceptors of the aortic arch and carotid arteries.
The hypothalamus and other brain regions associated with the limbic system also play roles in influencing the regulation of breathing by interacting with the respiratory centres. The hypothalamus and other regions associated with the limbic system are involved in regulating respiration in response to emotions, pain, and temperature. For example, an increase in body temperature causes an increase in respiratory rate. Feeling excited or the fight-or-flight response will also result in an increase in respiratory rate.
Disorders of the Respiratory System: Sleep Apnoea
Sleep apnoea is a chronic disorder that can occur in children or adults and is characterised by the cessation of breathing during sleep. These episodes may last for several seconds or several minutes and may differ in the frequency with which they are experienced. Sleep apnoea leads to poor sleep, which is reflected in the symptoms of fatigue, evening napping, irritability, memory problems, and morning headaches. In addition, many individuals with sleep apnoea experience a dry throat in the morning after waking from sleep, which may be due to excessive snoring.
There are two types of sleep apnoea: obstructive sleep apnoea and central sleep apnoea. Obstructive sleep apnoea is caused by an obstruction of the airway during sleep, which can occur at different points in the airway, depending on the underlying cause of the obstruction. For example, the tongue and throat muscles of some individuals with obstructive sleep apnoea may relax excessively, causing the muscles to push into the airway. Another example is obesity, which is a known risk factor for sleep apnoea, as excess adipose tissue in the neck region can push the soft tissues towards the lumen of the airway, causing the trachea to narrow.
In central sleep apnoea, the respiratory centres of the brain do not respond properly to rising carbon dioxide levels and therefore do not stimulate the contraction of the diaphragm and intercostal muscles regularly. As a result, inspiration does not occur and breathing stops for a short period. In some cases, the cause of central sleep apnoea is unknown. However, some medical conditions, such as stroke and congestive heart failure, may cause damage to the pons or medulla oblongata. In addition, some pharmacologic agents, such as morphine, can affect the respiratory centres, causing a decrease in the respiratory rate. The symptoms of central sleep apnoea are similar to those of obstructive sleep apnoea.
A diagnosis of sleep apnoea is usually done during a sleep study, where the patient is monitored in a sleep laboratory for several nights. The patient’s blood oxygen levels, heart rate, respiratory rate, and blood pressure are monitored, as are brain activity and the volume of air that is inhaled and exhaled. Treatment of sleep apnoea commonly includes the use of a device called a continuous positive airway pressure (CPAP) machine during sleep. The CPAP machine has a mask that covers the nose, or the nose and mouth, and forces air into the airway at regular intervals. This pressurised air can help to gently force the airway to remain open, allowing more normal ventilation to occur. Other treatments include lifestyle changes to decrease weight, eliminate alcohol and other sleep apnoea–promoting drugs, and changes in sleep position. In addition to these treatments, patients with central sleep apnoea may need supplemental oxygen during sleep.
Section Review
Pulmonary ventilation is the process of breathing, which is driven by pressure differences between the lungs and the atmosphere. Atmospheric pressure is the force exerted by gases present in the atmosphere. The force exerted by gases within the alveoli is called intra-alveolar (intrapulmonary) pressure, whereas the force exerted by gases in the pleural cavity is called intrapleural pressure. Typically, intrapleural pressure is lower, or negative to, intra-alveolar pressure. The difference in pressure between intrapleural and intra-alveolar pressures is called transpulmonary pressure. In addition, intra-alveolar pressure will equalise with the atmospheric pressure. Pressure is determined by the volume of the space occupied by a gas and is influenced by resistance. Air flows when a pressure gradient is created, from a space of higher pressure to a space of lower pressure. Boyle’s law describes the relationship between volume and pressure. A gas is at lower pressure in a larger volume because the gas molecules have more space to in which to move. The same quantity of gas in a smaller volume results in gas molecules crowding together, producing increased pressure.
Resistance is created by inelastic surfaces, as well as the diameter of the airways. Resistance reduces the flow of gases. The surface tension of the alveoli also influences pressure, as it opposes the expansion of the alveoli. However, pulmonary surfactant helps to reduce the surface tension so that the alveoli do not collapse during expiration. The ability of the lungs to stretch, called lung compliance, also plays a role in gas flow. The more the lungs can stretch, the greater the potential volume of the lungs. The greater the volume of the lungs, the lower the air pressure within the lungs.
Pulmonary ventilation consists of the process of inspiration (or inhalation), where air enters the lungs, and expiration (or exhalation), where air leaves the lungs. During inspiration, the diaphragm and external intercostal muscles contract, causing the rib cage to expand and move outward, and expanding the thoracic cavity and lung volume. This creates a lower pressure within the lung than that of the atmosphere, causing air to be drawn into the lungs. During expiration, the diaphragm and intercostals relax, causing the thorax and lungs to recoil. The air pressure within the lungs increases to above the pressure of the atmosphere, causing air to be forced out of the lungs. However, during forced exhalation, the internal intercostals and abdominal muscles may be involved in forcing air out of the lungs.
Respiratory volume describes the amount of air in a given space within the lungs, or which can be moved by the lung, and is dependent on a variety of factors. Tidal volume refers to the amount of air that enters the lungs during quiet breathing, whereas inspiratory reserve volume is the amount of air that enters the lungs when a person inhales past the tidal volume. Expiratory reserve volume is the extra amount of air that can leave with forceful expiration, following tidal expiration. Residual volume is the amount of air that is left in the lungs after expelling the expiratory reserve volume. Respiratory capacity is the combination of two or more volumes. Anatomical dead space refers to the air within the respiratory structures that never participates in gas exchange, because it does not reach functional alveoli. Respiratory rate is the number of breaths taken per minute, which may change during certain diseases or conditions.
Both respiratory rate and depth are controlled by the respiratory centres of the brain, which are stimulated by factors such as chemical and pH changes in the blood. These changes are sensed by central chemoreceptors, which are located in the brain, and peripheral chemoreceptors, which are located in the aortic arch and carotid arteries. A rise in carbon dioxide or a decline in oxygen levels in the blood stimulates an increase in respiratory rate and depth.
Compare the composition of atmospheric air and alveolar air
Describe the mechanisms that drive gas exchange
Discuss the importance of sufficient ventilation and perfusion and how the body adapts when they are insufficient
Discuss the process of external respiration
Describe the process of internal respiration
The purpose of the respiratory system is to perform gas exchange. Pulmonary ventilation provides air to the alveoli for this gas exchange process. At the respiratory membrane, where the alveolar and capillary walls meet, gases move across the membranes, with oxygen entering the bloodstream and carbon dioxide exiting. It is through this mechanism that blood is oxygenated and carbon dioxide, the waste product of cellular respiration, is removed from the body.
Gas Exchange
In order to understand the mechanisms of gas exchange in the lung, it is important to understand the underlying principles of gases and their behaviour. In addition to Boyle’s law, several other gas laws help to describe the behaviour of gases.
Gas Laws and Air Composition
Gas molecules exert force on the surfaces with which they are in contact; this force is called pressure. In natural systems, gases are normally present as a mixture of different types of molecules. For example, the atmosphere consists of oxygen, nitrogen, carbon dioxide, and other gaseous molecules, and this gaseous mixture exerts a certain pressure referred to as atmospheric pressure (Table 8.4.1). Partial pressure (Px) is the pressure of a single type of gas in a mixture of gases. For example, in the atmosphere, oxygen exerts a partial pressure, and nitrogen exerts another partial pressure, independent of the partial pressure of oxygen (Figure 8.4.1). Total pressure is the sum of all the partial pressures of a gaseous mixture. Dalton’s law describes the behaviour of nonreactive gases in a gaseous mixture and states that a specific gas type in a mixture exerts its own pressure; thus, the total pressure exerted by a mixture of gases is the sum of the partial pressures of the gases in the mixture.
Table 8.4.1. Partial pressures of atmospheric gases
Gas
Percent of total composition
Partial pressure (mm Hg)
Nitrogen (N2)
78.6
597.4
Oxygen (O2)
20.9
158.8
Water (H2O)
0.04
3.0
Carbon dioxide (CO2)
0.004
0.3
Others
0.0006
0.5
Total composition/total atmospheric pressure
100%
760.0
Figure 8.4.1. Partial and total pressures of a gas. Partial pressure is the force exerted by a gas. The sum of the partial pressures of all the gases in a mixture equals the total pressure.
Partial pressure is extremely important in predicting the movement of gases. Recall that gases tend to equalise their pressure in two regions that are connected. A gas will move from an area where its partial pressure is higher to an area where its partial pressure is lower. In addition, the greater the partial pressure difference between the two areas, the more rapid is the movement of gases.
Solubility of Gases in Liquids
Henry’s law describes the behaviour of gases when they come into contact with a liquid, such as blood. Henry’s law states that the concentration of gas in a liquid is directly proportional to the solubility and partial pressure of that gas. The greater the partial pressure of the gas, the greater the number of gas molecules that will dissolve in the liquid. The concentration of the gas in a liquid is also dependent on the solubility of the gas in the liquid. For example, although nitrogen is present in the atmosphere, very little nitrogen dissolves into the blood, because the solubility of nitrogen in blood is very low. The exception to this occurs in scuba divers; the composition of the compressed air that divers breathe causes nitrogen to have a higher partial pressure than normal, causing it to dissolve in the blood in greater amounts than normal. Too much nitrogen in the bloodstream results in a serious condition that can be fatal if not corrected. Gas molecules establish an equilibrium between those molecules dissolved in liquid and those in air.
The composition of air in the atmosphere and in the alveoli differs. In both cases, the relative concentration of gases is nitrogen > oxygen > water vapor > carbon dioxide. The amount of water vapor present in alveolar air is greater than that in atmospheric air (Table 8.4.2). Recall that the respiratory system works to humidify incoming air, thereby causing the air present in the alveoli to have a greater amount of water vapor than atmospheric air. In addition, alveolar air contains a greater amount of carbon dioxide and less oxygen than atmospheric air. This is no surprise, as gas exchange removes oxygen from and adds carbon dioxide to alveolar air. Both deep and forced breathing cause the alveolar air composition to be changed more rapidly than during quiet breathing. As a result, the partial pressures of oxygen and carbon dioxide change, affecting the diffusion process that moves these materials across the membrane. This will cause oxygen to enter and carbon dioxide to leave the blood more quickly.
Table 8.4.2. Composition and partial Pressures of alveolar air
Gas
Percent of total composition
in Alveoli air
Alveolar air
Partial pressure (mm Hg)
Nitrogen (N2)
74.9
569
Oxygen (O2)
13.7
104
Water (H2O)
6.2
40
Carbon dioxide (CO2)
5.2
47
Total composition/total atmospheric pressure
100%
760.0
Ventilation and Perfusion
Two important aspects of gas exchange in the lung are ventilation and perfusion. Ventilation is the movement of air into and out of the lungs, and perfusion is the flow of blood in the pulmonary capillaries. For gas exchange to be efficient, the volumes involved in ventilation and perfusion should be compatible. However, factors such as regional gravity effects on blood, blocked alveolar ducts, or disease can cause ventilation and perfusion to be imbalanced.
The partial pressure of oxygen in alveolar air is about 104 mm Hg, whereas the partial pressure of the oxygenated pulmonary venous blood is about 100 mm Hg. When ventilation is sufficient, oxygen enters the alveoli at a high rate, and the partial pressure of oxygen in the alveoli remains high. In contrast, when ventilation is insufficient, the partial pressure of oxygen in the alveoli drops. Without the large difference in partial pressure between the alveoli and the blood, oxygen does not diffuse efficiently across the respiratory membrane. The body has mechanisms that counteract this problem. In cases when ventilation is not sufficient for an alveolus, the body redirects blood flow to alveoli that are receiving sufficient ventilation. This is achieved by constricting the pulmonary arterioles that serve the dysfunctional alveolus, which redirects blood to other alveoli that have sufficient ventilation. At the same time, the pulmonary arterioles that serve alveoli receiving sufficient ventilation vasodilate, which brings in greater blood flow. Factors such as carbon dioxide, oxygen and pH levels can all serve as stimuli for adjusting blood flow in the capillary networks associated with the alveoli.
Ventilation is regulated by the diameter of the airways, whereas perfusion is regulated by the diameter of the blood vessels. The diameter of the bronchioles is sensitive to the partial pressure of carbon dioxide in the alveoli. A greater partial pressure of carbon dioxide in the alveoli causes the bronchioles to increase their diameter as will a decreased level of oxygen in the blood supply, allowing carbon dioxide to be exhaled from the body at a greater rate. As mentioned above, a greater partial pressure of oxygen in the alveoli causes the pulmonary arterioles to dilate, increasing blood flow.
Gas Exchange
Gas exchange occurs at two sites in the body: in the lungs, where oxygen is picked up and carbon dioxide is released at the respiratory membrane, and at the tissues, where oxygen is released and carbon dioxide is picked up. External respiration is the exchange of gases with the external environment and occurs in the alveoli of the lungs. Internal respiration is the exchange of gases with the internal environment and occurs in the tissues. The actual exchange of gases occurs due to simple diffusion. Energy is not required to move oxygen or carbon dioxide across membranes. Instead, these gases follow pressure gradients that allow them to diffuse. The anatomy of the lung maximises the diffusion of gases: The respiratory membrane is highly permeable to gases; the respiratory and blood capillary membranes are very thin; and there is a large surface area throughout the lungs.
External Respiration
The pulmonary artery carries deoxygenated blood into the lungs from the heart, where it branches and eventually becomes the capillary network composed of pulmonary capillaries. These pulmonary capillaries create the respiratory membrane with the alveoli (Figure 8.4.2). As the blood is pumped through this capillary network, gas exchange occurs. Although a small amount of the oxygen is able to dissolve directly into plasma from the alveoli, most of the oxygen is picked up by erythrocytes (red blood cells) and binds to a protein called haemoglobin, a process described later in this chapter. Oxygenated haemoglobin is red, causing the overall appearance of bright red oxygenated blood, which returns to the heart through the pulmonary veins. Carbon dioxide is released in the opposite direction of oxygen, from the blood to the alveoli. Some of the carbon dioxide is returned on haemoglobin but can also be dissolved in plasma or is present as a converted form, also explained in greater detail later in this section.
External respiration occurs as a function of partial pressure differences in oxygen and carbon dioxide between the alveoli and the blood in the pulmonary capillaries.
Figure 8.4.2. External respiration. In external respiration, oxygen diffuses across the respiratory membrane from the alveolus to the capillary, whereas carbon dioxide diffuses out of the capillary into the alveolus.
Although the solubility of oxygen in blood is not high, there is a drastic difference in the partial pressure of oxygen in the alveoli versus in the blood of the pulmonary capillaries. This difference is about 64 mm Hg: The partial pressure of oxygen in the alveoli is about 104 mm Hg, whereas its partial pressure in the blood of the capillary is about 40 mm Hg. This large difference in partial pressure creates a very strong pressure gradient that causes oxygen to rapidly cross the respiratory membrane from the alveoli into the blood.
The partial pressure of carbon dioxide is also different between the alveolar air and the blood of the capillary. However, the partial pressure difference is less than that of oxygen, about 5 mm Hg. The partial pressure of carbon dioxide in the blood of the capillary is about 45 mm Hg, whereas its partial pressure in the alveoli is about 40 mm Hg. However, the solubility of carbon dioxide is much greater than that of oxygen—by a factor of about 20—in both blood and alveolar fluids. As a result, the relative concentrations of oxygen and carbon dioxide that diffuse across the respiratory membrane are similar.
Internal Respiration
Internal respiration is gas exchange that occurs at the level of body tissues (Figure 8.4.3). Similar to external respiration, internal respiration also occurs as simple diffusion due to a partial pressure gradient. However, the partial pressure gradients are opposite of those present at the respiratory membrane. The partial pressure of oxygen in tissues is low, about 40 mm Hg, because oxygen is continuously used for cellular respiration. In contrast, the partial pressure of oxygen in the blood is about 100 mm Hg. This creates a pressure gradient that causes oxygen to dissociate from haemoglobin, diffuse out of the blood, cross the interstitial space, and enter the tissue. Haemoglobin that has little oxygen bound to it loses much of its brightness, so that blood returning to the heart is more burgundy in colour.
Considering that cellular respiration continuously produces carbon dioxide, the partial pressure of carbon dioxide is lower in the blood than it is in the tissue, causing carbon dioxide to diffuse out of the tissue, cross the interstitial fluid, and enter the blood. It is then carried back to the lungs either bound to haemoglobin, dissolved in plasma, or in a converted form. By the time blood returns to the heart, the partial pressure of oxygen has returned to about 40 mm Hg, and the partial pressure of carbon dioxide has returned to about 45 mm Hg. The blood is then pumped back to the lungs to be oxygenated once again during external respiration.
Figure 8.4.3. Internal respiration. Oxygen diffuses out of the capillary and into cells, whereas carbon dioxide diffuses out of cells and into the capillary.
Everyday Connection
Hyperbaric Chamber Treatment
A type of device used in some areas of medicine that exploits the behaviour of gases is hyperbaric chamber treatment. A hyperbaric chamber is a unit that can be sealed and expose a patient to either 100 percent oxygen with increased pressure or a mixture of gases that includes a higher concentration of oxygen than normal atmospheric air, also at a higher partial pressure than the atmosphere. There are two major types of chambers: monoplace and multiplace. Monoplace chambers are typically for one patient, and the staff tending to the patient observes the patient from outside of the chamber (Figure 8.4.4). Some facilities have special monoplace hyperbaric chambers that allow multiple patients to be treated at once, usually in a sitting or reclining position, to help ease feelings of isolation or claustrophobia. Multiplace chambers are large enough for multiple patients to be treated at one time, and the staff attending these patients is present inside the chamber. In a multiplace chamber, patients are often treated with air via a mask or hood, and the chamber is pressurised.
Hyperbaric chamber treatment is based on the behaviour of gases. As you recall, gases move from a region of higher partial pressure to a region of lower partial pressure. In a hyperbaric chamber, the atmospheric pressure is increased, causing a greater amount of oxygen than normal to diffuse into the bloodstream of the patient. Hyperbaric chamber therapy is used to treat a variety of medical problems, such as wound and graft healing, anaerobic bacterial infections, and carbon monoxide poisoning. Exposure to and poisoning by carbon monoxide is difficult to reverse, because haemoglobin’s affinity for carbon monoxide is much stronger than its affinity for oxygen, causing carbon monoxide to replace oxygen in the blood. Hyperbaric chamber therapy can treat carbon monoxide poisoning, because the increased atmospheric pressure causes more oxygen to diffuse into the bloodstream. At this increased pressure and increased concentration of oxygen, carbon monoxide is displaced from haemoglobin. Another example is the treatment of anaerobic bacterial infections, which are created by bacteria that cannot or prefer not to live in the presence of oxygen. An increase in blood and tissue levels of oxygen helps to kill the anaerobic bacteria that are responsible for the infection, as oxygen is toxic to anaerobic bacteria. For wounds and grafts, the chamber stimulates the healing process by increasing energy production needed for repair. Increasing oxygen transport allows cells to ramp up cellular respiration and thus ATP production, the energy needed to build new structures.
Section Review
The behaviour of gases can be explained by the principles of Dalton’s law and Henry’s law, both of which describe aspects of gas exchange. Dalton’s law states that each specific gas in a mixture of gases exerts force (its partial pressure) independently of the other gases in the mixture. Henry’s law states that the amount of a specific gas that dissolves in a liquid is a function of its partial pressure. The greater the partial pressure of a gas, the more of that gas will dissolve in a liquid, as the gas moves toward equilibrium. Gas molecules move down a pressure gradient; in other words, gas moves from a region of high pressure to a region of low pressure. The partial pressure of oxygen is high in the alveoli and low in the blood of the pulmonary capillaries. As a result, oxygen diffuses across the respiratory membrane from the alveoli into the blood. In contrast, the partial pressure of carbon dioxide is high in the pulmonary capillaries and low in the alveoli. Therefore, carbon dioxide diffuses across the respiratory membrane from the blood into the alveoli. The amount of oxygen and carbon dioxide that diffuses across the respiratory membrane is similar.
Ventilation is the process that moves air into and out of the alveoli, and perfusion affects the flow of blood in the capillaries. Both are important in gas exchange, as ventilation must be sufficient to create a high partial pressure of oxygen in the alveoli. If ventilation is insufficient and the partial pressure of oxygen drops in the alveolar air, the capillary is constricted and blood flow is redirected to alveoli with sufficient ventilation. External respiration refers to gas exchange that occurs in the alveoli, whereas internal respiration refers to gas exchange that occurs in the tissue. Both are driven by partial pressure differences.
Describe the principles of carbon dioxide transport
The other major activity in the lungs is the process of respiration, the process of gas exchange. The function of respiration is to provide oxygen for use by body cells during cellular respiration and to eliminate carbon dioxide, a waste product of cellular respiration, from the body. In order for the exchange of oxygen and carbon dioxide to occur, both gases must be transported between the external and internal respiration sites. Although carbon dioxide is more soluble than oxygen in blood, both gases require a specialised transport system for the majority of the gas molecules to be moved between the lungs and other tissues.
Oxygen Transport in the Blood
Even though oxygen is transported via the blood, you may recall that oxygen is not very soluble in liquids. A small amount of oxygen does dissolve in the blood and is transported in the bloodstream, but it is only about 1.5% of the total amount. The majority of oxygen molecules are carried from the lungs to the body’s tissues by a specialised transport system, which relies on the erythrocyte—the red blood cell. Erythrocytes contain a metalloprotein, haemoglobin, which serves to bind oxygen molecules to the erythrocyte (Figure 8.5.1). Haem is the portion of haemoglobin that contains iron, and it is haem that binds oxygen. One haemoglobin molecule contains iron-containing Haem molecules, and because of this, each haemoglobin molecule is capable of carrying up to four molecules of oxygen. As oxygen diffuses across the respiratory membrane from the alveolus to the capillary, it also diffuses into the red blood cell and is bound by haemoglobin. The following reversible chemical reaction describes the production of the final product, oxyhaemoglobin (Hb–O2), which is formed when oxygen binds to haemoglobin. Oxyhaemoglobin is a bright red-coloured molecule that contributes to the bright red colour of oxygenated blood.
Hb + O2 ↔ Hb − O2
In this formula, Hb represents reduced haemoglobin, that is, haemoglobin that does not have oxygen bound to it. There are multiple factors involved in how readily haem binds to and dissociates from oxygen, which will be discussed in the subsequent sections.
Figure 8.5.1. Erythrocyte and haemoglobin. Haemoglobin consists of four subunits, each of which contains one molecule of iron.
Function of Haemoglobin
Haemoglobin is composed of subunits, a protein structure that is referred to as a quaternary structure. Each of the four subunits that make up haemoglobin is arranged in a ring-like fashion, with an iron atom covalently bound to the haem in the centre of each subunit. Binding of the first oxygen molecule causes a conformational change in haemoglobin that allows the second molecule of oxygen to bind more readily. As each molecule of oxygen is bound, it further facilitates the binding of the next molecule, until all four haem sites are occupied by oxygen. The opposite occurs as well: After the first oxygen molecule dissociates and is “dropped off” at the tissues, the next oxygen molecule dissociates more readily. When all four haem sites are occupied, the haemoglobin is said to be saturated. When one to three haem sites are occupied, the haemoglobin is said to be partially saturated. Therefore, when considering the blood as a whole, the percent of the available haem units that are bound to oxygen at a given time is called haemoglobin saturation. Haemoglobin saturation of 100 percent means that every haem unit in all of the erythrocytes of the body is bound to oxygen. In a healthy individual with normal haemoglobin levels, haemoglobin saturation generally ranges from 95 percent to 99 percent.
Oxygen Dissociation from Haemoglobin
Partial pressure is an important aspect of the binding of oxygen to and disassociation from haem. An oxygen–haemoglobin dissociation curve is a graph that describes the relationship of partial pressure to the binding of oxygen to haem and its subsequent dissociation from haem (Figure 8.5.2). Remember that gases travel from an area of higher partial pressure to an area of lower partial pressure. In addition, the affinity of an oxygen molecule for haem increases as more oxygen molecules are bound. Therefore, in the oxygen–haemoglobin saturation curve, as the partial pressure of oxygen increases, a proportionately greater number of oxygen molecules are bound by haem. Not surprisingly, the oxygen–haemoglobin saturation/dissociation curve also shows that the lower the partial pressure of oxygen, the fewer oxygen molecules are bound to haem. As a result, the partial pressure of oxygen plays a major role in determining the degree of binding of oxygen to haem at the site of the respiratory membrane, as well as the degree of dissociation of oxygen from haem at the site of body tissues.
Figure 8.5.2. Oxygen-haemoglobin dissociation curves and effects of pH and temperature. These three graphs show (a) the relationship between the partial pressure of oxygen and haemoglobin saturation, (b) the effect of pH on the oxygen–haemoglobin dissociation curve, and (c) the effect of temperature on the oxygen–haemoglobin dissociation curve.
The mechanisms behind the oxygen–haemoglobin saturation/dissociation curve also serve as automatic control mechanisms that regulate how much oxygen is delivered to different tissues throughout the body. This is important because some tissues have a higher metabolic rate than others. Highly active tissues, such as muscle, rapidly use oxygen to produce ATP, lowering the partial pressure of oxygen in the tissue to about 20 mm Hg. The partial pressure of oxygen inside capillaries is about 100 mm Hg, so the difference between the two becomes quite high, about 80 mm Hg. As a result, a greater number of oxygen molecules dissociate from haemoglobin and enter the tissues. The reverse is true of tissues, such as adipose (body fat), which have lower metabolic rates. Because less oxygen is used by these cells, the partial pressure of oxygen within such tissues remains relatively high, resulting in fewer oxygen molecules dissociating from haemoglobin and entering the tissue interstitial fluid. Although venous blood is said to be deoxygenated, some oxygen is still bound to haemoglobin in its red blood cells. This provides an oxygen reserve that can be used when tissues suddenly demand more oxygen.
Factors other than partial pressure also affect the oxygen–haemoglobin saturation/dissociation curve. For example, a higher temperature promotes haemoglobin and oxygen to dissociate faster, whereas a lower temperature inhibits dissociation (see Figure 8.5.2, middle). However, the human body tightly regulates temperature, so this factor may not affect gas exchange throughout the body. The exception to this is in highly active tissues, which may release a larger amount of energy than is given off as heat. As a result, oxygen readily dissociates from haemoglobin, which is a mechanism that helps to provide active tissues with more oxygen.
Certain hormones, such as androgens, adrenaline, thyroid hormones, and growth hormone, can affect the oxygen–haemoglobin saturation/dissociation curve by stimulating the production of a compound called 2,3-bisphosphoglycerate (BPG) by erythrocytes. BPG is a by-product of glycolysis. Because erythrocytes do not contain mitochondria, glycolysis is the sole method by which these cells produce ATP. BPG promotes the dissociation of oxygen from haemoglobin. Therefore, the greater the concentration of BPG, the more readily oxygen dissociates from haemoglobin, despite its partial pressure.
The pH of the blood is another factor that influences the oxygen–haemoglobin saturation/dissociation curve (see Figure 8.5.2). The Bohr effect is a phenomenon that arises from the relationship between pH and oxygen’s affinity for haemoglobin: A lower, more acidic pH promotes oxygen dissociation from haemoglobin. In contrast, a higher, or more basic, pH inhibits oxygen dissociation from haemoglobin. The greater the amount of carbon dioxide in the blood, the more molecules that must be converted, which in turn generates hydrogen ions and thus lowers blood pH. Furthermore, blood pH may become more acidic when certain by-products of cell metabolism, such as lactic acid, carbonic acid, and carbon dioxide, are released into the bloodstream.
Haemoglobin of the Foetus
The foetus has its own circulation with its own erythrocytes; however, it is dependent on the mother for oxygen. Blood is supplied to the foetus by way of the umbilical cord, which is connected to the placenta and separated from maternal blood by the chorion. The mechanism of gas exchange at the chorion is similar to gas exchange at the respiratory membrane. However, the partial pressure of oxygen is lower in the maternal blood in the placenta, at about 35 to 50 mm Hg, than it is in maternal arterial blood. The difference in partial pressures between maternal and foetal blood is not large, as the partial pressure of oxygen in foetal blood at the placenta is about 20 mm Hg. Therefore, there is not as much diffusion of oxygen into the foetal blood supply. The foetus’ haemoglobin overcomes this problem by having a greater affinity for oxygen than maternal haemoglobin (Figure 8.5.3). Both foetal and adult haemoglobin have four subunits, but two of the subunits of foetal haemoglobin have a different structure that causes foetal haemoglobin to have a greater affinity for oxygen than does adult haemoglobin.
Carbon Dioxide Transport in the Blood
Carbon dioxide is transported by three major mechanisms. The first mechanism of carbon dioxide transport is by blood plasma, as some carbon dioxide molecules dissolve in the blood. The second mechanism is transport in the form of bicarbonate (HCO3–), which also dissolves in plasma. The third mechanism of carbon dioxide transport is similar to the transport of oxygen by erythrocytes (Figure 8.5.4).
Figure 8.5.4. Carbon dioxide transport. Carbon dioxide is transported by three different methods: (a) in erythrocytes; (b) after forming carbonic acid (H2CO3), which is dissolved in plasma; (c) and in plasma.
Dissolved Carbon Dioxide
Although carbon dioxide is not considered to be highly soluble in blood, a small fraction—about 7 to 10 percent—of the carbon dioxide that diffuses into the blood from the tissues dissolves in plasma. The dissolved carbon dioxide then travels in the bloodstream and when the blood reaches the pulmonary capillaries, the dissolved carbon dioxide diffuses across the respiratory membrane into the alveoli, where it is then exhaled during pulmonary ventilation.
Bicarbonate Buffer
A large fraction—about 70 percent—of the carbon dioxide molecules that diffuse into the blood is transported to the lungs as bicarbonate. Most bicarbonate is produced in erythrocytes after carbon dioxide diffuses into the capillaries, and subsequently into red blood cells. Carbonic anhydrase (CA) causes carbon dioxide and water to form carbonic acid (H2CO3), which dissociates into two ions: bicarbonate (HCO3–) and hydrogen (H+). The following formula depicts this reaction:
Bicarbonate tends to build up in the erythrocytes, so that there is a greater concentration of bicarbonate in the erythrocytes than in the surrounding blood plasma. As a result, some of the bicarbonate will leave the erythrocytes and move down its concentration gradient into the plasma in exchange for chloride (Cl–) ions. This phenomenon is referred to as the chloride shift and occurs because by exchanging one negative ion for another negative ion, neither the electrical charge of the erythrocytes nor that of the blood is altered.
At the pulmonary capillaries, the chemical reaction that produced bicarbonate (shown above) is reversed, and carbon dioxide and water are the products. Much of the bicarbonate in the plasma re-enters the erythrocytes in exchange for chloride ions. Hydrogen ions and bicarbonate ions join to form carbonic acid, which is converted into carbon dioxide and water by carbonic anhydrase. Carbon dioxide diffuses out of the erythrocytes and into the plasma, where it can further diffuse across the respiratory membrane into the alveoli to be exhaled during pulmonary ventilation.
Carbaminohaemoglobin
About 20 percent of carbon dioxide is bound by haemoglobin and is transported to the lungs. Carbon dioxide does not bind to iron as oxygen does; instead, carbon dioxide binds amino acid moieties on the globin portions of haemoglobin to form carbaminohaemoglobin, which forms when haemoglobin and carbon dioxide bind. When haemoglobin is not transporting oxygen, it tends to have a bluish-purple tone to it, creating the darker maroon colour typical of deoxygenated blood. The following formula depicts this reversible reaction:
CO2 + Hb ↔ HbCO2
Similar to the transport of oxygen by haem, the binding and dissociation of carbon dioxide to and from haemoglobin is dependent on the partial pressure of carbon dioxide. Because carbon dioxide is released from the lungs, blood that leaves the lungs and reaches body tissues has a lower partial pressure of carbon dioxide than is found in the tissues. As a result, carbon dioxide leaves the tissues because of its higher partial pressure, enters the blood, and then moves into red blood cells, binding to haemoglobin. In contrast, in the pulmonary capillaries, the partial pressure of carbon dioxide is high compared to within the alveoli. As a result, carbon dioxide dissociates readily from haemoglobin and diffuses across the respiratory membrane into the air.
In addition to the partial pressure of carbon dioxide, the oxygen saturation of haemoglobin and the partial pressure of oxygen in the blood also influence the affinity of haemoglobin for carbon dioxide. The Haldane effect is a phenomenon that arises from the relationship between the partial pressure of oxygen and the affinity of haemoglobin for carbon dioxide. haemoglobin that is saturated with oxygen does not readily bind carbon dioxide. However, when oxygen is not bound to haem and the partial pressure of oxygen is low, haemoglobin readily binds to carbon dioxide.
Section Review
Oxygen is primarily transported through the blood by erythrocytes. These cells contain a metalloprotein called haemoglobin, which is composed of four subunits with a ring-like structure. Each subunit contains one atom of iron bound to a molecule of haem. Haem binds oxygen so that each haemoglobin molecule can bind up to four oxygen molecules. When all the haem units in the blood are bound to oxygen, haemoglobin is considered to be saturated. Haemoglobin is partially saturated when only some haem units are bound to oxygen. An oxygen–haemoglobin saturation/dissociation curve is a common way to depict the relationship of how easily oxygen binds to or dissociates from haemoglobin as a function of the partial pressure of oxygen. As the partial pressure of oxygen increases, the more readily haemoglobin binds to oxygen. At the same time, once one molecule of oxygen is bound by haemoglobin, additional oxygen molecules more readily bind to haemoglobin. Other factors such as temperature, pH, the partial pressure of carbon dioxide, and the concentration of 2,3-bisphosphoglycerate can enhance or inhibit the binding of haemoglobin and oxygen as well. Foetal haemoglobin has a different structure than adult haemoglobin, which results in foetal haemoglobin having a greater affinity for oxygen than adult haemoglobin.
Carbon dioxide is transported in blood by three different mechanisms: as dissolved carbon dioxide, as bicarbonate, or as carbaminohaemoglobin. A small portion of carbon dioxide remains. The largest amount of transported carbon dioxide is as bicarbonate, formed in erythrocytes. For this conversion, carbon dioxide is combined with water with the aid of an enzyme called carbonic anhydrase. This combination forms carbonic acid, which spontaneously dissociates into bicarbonate and hydrogen ions. As bicarbonate builds up in erythrocytes, it is moved across the membrane into the plasma in exchange for chloride ions by a mechanism called the chloride shift. At the pulmonary capillaries, bicarbonate re-enters erythrocytes in exchange for chloride ions, and the reaction with carbonic anhydrase is reversed, recreating carbon dioxide and water. Carbon dioxide then diffuses out of the erythrocyte and across the respiratory membrane into the air. An intermediate amount of carbon dioxide binds directly to haemoglobin to form carbaminohaemoglobin. The partial pressures of carbon dioxide and oxygen, as well as the oxygen saturation of haemoglobin, influence how readily haemoglobin binds carbon dioxide. The less saturated haemoglobin is and the lower the partial pressure of oxygen in the blood is, the more readily haemoglobin binds to carbon dioxide. This is an example of the Haldane effect.
Describe the effect of exercise on the respiratory system
Describe the effect of high altitude on the respiratory system
Discuss the process of acclimatisation
At rest, the respiratory system performs its functions at a constant, rhythmic pace, as regulated by the respiratory centres of the brain. At this pace, ventilation provides sufficient oxygen to all the tissues of the body. However, there are times that the respiratory system must alter the pace of its functions in order to accommodate the oxygen demands of the body.
Hyperpnoea
Hyperpnoea is an increased depth and rate of ventilation to meet an increase in oxygen demand as might be seen in exercise or disease, particularly diseases that target the respiratory or digestive tracts. This does not significantly alter blood oxygen or carbon dioxide levels, but merely increases the depth and rate of ventilation to meet the demand of the cells. In contrast, hyperventilation is an increased ventilation rate that is independent of the cellular oxygen needs and leads to abnormally low blood carbon dioxide levels and high (alkaline) blood pH.
Interestingly, exercise does not cause hyperpnoea as one might think. Muscles that perform work during exercise do increase their demand for oxygen, stimulating an increase in ventilation. However, hyperpnoea during exercise appears to occur before a drop in oxygen levels within the muscles can occur. Therefore, hyperpnoea must be driven by other mechanisms, either instead of or in addition to a drop in oxygen levels. The exact mechanisms behind exercise hyperpnoea are not well understood, and some hypotheses are somewhat controversial. However, in addition to low oxygen, high carbon dioxide, and low pH levels, there appears to be a complex interplay of factors related to the nervous system and the respiratory centres of the brain.
First, a conscious decision to partake in exercise, or another form of physical exertion, results in a psychological stimulus that may trigger the respiratory centres of the brain to increase ventilation. In addition, the respiratory centres of the brain may be stimulated through the activation of motor neurons that innervate muscle groups that are involved in the physical activity. Finally, physical exertion stimulates proprioceptors, which are receptors located within the muscles, joints, and tendons, which sense movement and stretching; proprioceptors thus create a stimulus that may also trigger the respiratory centres of the brain. These neural factors are consistent with the sudden increase in ventilation that is observed immediately as exercise begins. Because the respiratory centres are stimulated by psychological, motor neuron, and proprioceptor inputs throughout exercise, the fact that there is also a sudden decrease in ventilation immediately after the exercise ends when these neural stimuli cease, further supports the idea that they are involved in triggering the changes of ventilation.
High Altitude Effects
An increase in altitude results in a decrease in atmospheric pressure. Although the proportion of oxygen relative to gases in the atmosphere remains at 21 percent, its partial pressure decreases (Table 8.6.1). As a result, it is more difficult for a body to achieve the same level of oxygen saturation at high altitude than at low altitude, due to lower atmospheric pressure. In fact, haemoglobin saturation is lower at high altitudes compared to haemoglobin saturation at sea level. For example, haemoglobin saturation is about 67 percent at 5791 meters above sea level, whereas it reaches about 98 percent at sea level.
Table 8.6.1. Partial pressure of oxygen at different altitudes
Example Location
Altitude
(metres above sea level)
Atmospheric pressure
(mm Hg)
Partial pressure of oxygen
(mm Hg)
New York City, New York
0
760
159
Boulder, Colorado
1,524
632
133
Aspen, Colorado
2,438
565
118
Pike’s Peak, Colorado
4,267
447
94
Denali (Mt. McKinley), Alaska
6,096
350
73
Mt. Everest, Tibet
8,839
260
54
As you recall, partial pressure is extremely important in determining how much gas can cross the respiratory membrane and enter the blood of the pulmonary capillaries. A lower partial pressure of oxygen means that there is a smaller difference in partial pressures between the alveoli and the blood, so less oxygen crosses the respiratory membrane. As a result, fewer oxygen molecules are bound by haemoglobin. Despite this, the tissues of the body still receive a sufficient amount of oxygen during rest at high altitudes. This is due to two major mechanisms. First, the number of oxygen molecules that enter the tissue from the blood is nearly equal between sea level and high altitudes. At sea level, haemoglobin saturation is higher, but only a quarter of the oxygen molecules are actually released into the tissue. At high altitudes, a greater proportion of molecules of oxygen are released into the tissues. Secondly, at high altitudes, a greater amount of BPG is produced by erythrocytes, which enhances the dissociation of oxygen from haemoglobin. Physical exertion, such as skiing or hiking, can lead to altitude sickness due to the low amount of oxygen reserves in the blood at high altitudes. At sea level, there is a large amount of oxygen reserve in venous blood (even though venous blood is thought of as “deoxygenated”) from which the muscles can draw during physical exertion. Because the oxygen saturation is much lower at higher altitudes, this venous reserve is small, resulting in pathological symptoms of low blood oxygen levels. You may have heard that it is important to drink more water when traveling at higher altitudes than you are accustomed to. This is because your body will increase micturition (urination) at high altitudes to counteract the effects of lower oxygen levels. By removing fluids, blood plasma levels drop but not the total number of erythrocytes. In this way, the overall concentration of erythrocytes in the blood increases, which helps tissues obtain the oxygen they need.
Acute mountain sickness (AMS), or altitude sickness, is a condition that results from acute exposure to high altitudes due to a low partial pressure of oxygen at high altitudes. AMS typically can occur at 2400 metres (8000 feet) above sea level. AMS is a result of low blood oxygen levels, as the body has acute difficulty adjusting to the low partial pressure of oxygen. In serious cases, AMS can cause pulmonary or cerebral oedema. Symptoms of AMS include nausea, vomiting, fatigue, light-headedness, drowsiness, feeling disoriented, increased pulse, and nosebleeds. The only treatment for AMS is descending to a lower altitude; however, pharmacologic treatments and supplemental oxygen can improve symptoms. AMS can be prevented by slowly ascending to the desired altitude, allowing the body to acclimate, as well as maintaining proper hydration.
Acclimatisation
Especially in situations where the ascent occurs too quickly, traveling to areas of high altitude can cause AMS. Acclimatisation is the process of adjustment that the respiratory system makes due to chronic exposure to a high altitude. Over a period of time, the body adjusts to accommodate the lower partial pressure of oxygen. The low partial pressure of oxygen at high altitudes results in a lower oxygen saturation level of haemoglobin in the blood. In turn, the tissue levels of oxygen are also lower. As a result, the kidneys are stimulated to produce the hormone erythropoietin (EPO), which stimulates the production of erythrocytes, resulting in a greater number of circulating erythrocytes in an individual at a high altitude over a long period. This process, however, is slow-acting as it will take approximately 3-4 days for reticulocytosis to become apparent following the rise in plasma EPO. With more red blood cells, there is more haemoglobin to help transport the available oxygen. Even though there is low saturation of each haemoglobin molecule, there will be more haemoglobin present, and therefore more oxygen in the blood. Over time, this allows the person to partake in physical exertion without developing AMS.
Section Review
Normally, the respiratory centres of the brain maintain a consistent, rhythmic breathing cycle. However, in certain cases, the respiratory system must adjust to situational changes in order to supply the body with sufficient oxygen. For example, exercise results in increased ventilation, and chronic exposure to a high altitude results in a greater number of circulating erythrocytes. Hyperpnea, an increase in the rate and depth of ventilation, appears to be a function of three neural mechanisms that include a psychological stimulus, motor neuron activation of skeletal muscles, and the activation of proprioceptors in the muscles, joints, and tendons. As a result, hyperpnea related to exercise is initiated when exercise begins, as opposed to when tissue oxygen demand increases.
In contrast, acute exposure to a high altitude, particularly during times of physical exertion, does result in low blood and tissue levels of oxygen. This change is caused by a low partial pressure of oxygen in the air, because the atmospheric pressure at high altitudes is lower than the atmospheric pressure at sea level. This can lead to a condition called acute mountain sickness (AMS) with symptoms that include headaches, disorientation, fatigue, nausea, and light-headedness. Over a long period of time, a person’s body will adjust to the high altitude, a process called acclimatisation. During acclimatisation, the low tissue levels of oxygen will cause the kidneys to produce greater amounts of the hormone erythropoietin, which stimulates the production of erythrocytes. Increased levels of circulating erythrocytes provide an increased amount of haemoglobin that helps supply an individual with more oxygen, preventing the symptoms of AMS.
8.7 Embryonic Development of the Respiratory System
Learning Objectives
By this section, you will be able to:
Create a timeline of the phases of respiratory development in the foetus
Propose reasons for foetal breathing movements
Explain how the lungs become inflated after birth
Development of the respiratory system begins early in the foetus. It is a complex process that includes many structures, most of which arise from the endoderm. From as early as week 10 of gestation the foetus can be observed making breathing movements, however it is more evident in later weeks with ~15% of cases observing foetal breathing between weeks 24 to 28, and ~35% of foetal breathing can be observed in weeks 30 onwards. Until birth, however, the mother provides all of the oxygen to the foetus as well as removes all of the foetal carbon dioxide via the placenta.
Timeline
The development of the respiratory system begins at about week 4 of gestation. By week 28, enough alveoli have matured that a baby born prematurely at this time can usually breathe on its own. The respiratory system, however, is not fully developed until early childhood, when a full complement of mature alveoli is present.
Weeks 4–7
Respiratory development in the embryo begins around week 4. Ectodermal tissue from the anterior head region invaginates posteriorly to form olfactory pits, which fuse with endodermal tissue of the developing pharynx. An olfactory pit is one of a pair of structures that will enlarge to become the nasal cavity. At about this same time, the lung bud forms. The lung bud is a dome-shaped structure composed of tissue that bulges from the foregut. The foregut is endoderm just inferior to the pharyngeal pouches. The laryngotracheal bud is a structure that forms from the longitudinal extension of the lung bud as development progresses. The portion of this structure nearest the pharynx becomes the trachea, whereas the distal end becomes more bulbous, forming bronchial buds. A bronchial bud is one of a pair of structures that will eventually become the bronchi and all other lower respiratory structures (Figure 8.7.1).
Figure 8.7.1. Development of the lower respiratory system.
Weeks 7–16
Bronchial buds continue to branch as development progresses until all of the segmental bronchi have been formed. Beginning around week 13, the lumens of the bronchi begin to expand in diameter. By week 16, respiratory bronchioles form. The foetus now has all major lung structures involved in the airway.
Weeks 16–24
Once the respiratory bronchioles form, further development includes extensive vascularisation, or the development of the blood vessels, as well as the formation of alveolar ducts and alveolar precursors. At about week 19, the respiratory bronchioles have formed. In addition, cells lining the respiratory structures begin to differentiate to form type I and type II pneumocytes. Once type II cells have differentiated, they begin to secrete small amounts of pulmonary surfactant. Around week 20, foetal breathing movements may begin.
Weeks 24–TERM
Major growth and maturation of the respiratory system occurs from week 24 until term. More alveolar precursors develop, and larger amounts of pulmonary surfactant are produced. Surfactant levels are not generally adequate to create effective lung compliance until about the eighth month of pregnancy. The respiratory system continues to expand, and the surfaces that will form the respiratory membrane develop further. At this point, pulmonary capillaries have formed and continue to expand, creating a large surface area for gas exchange. The major milestone of respiratory development occurs at around week 28, when sufficient alveolar precursors have matured so that a baby born prematurely at this time can usually breathe on its own. However, alveoli continue to develop and mature into childhood. A full complement of functional alveoli does not appear until around 8 years of age.
Foetal “Breathing”
Although the function of foetal breathing movements is not entirely clear, they can be observed starting at 20–21 weeks of development. Foetal breathing movements involve muscle contractions that cause the inhalation of amniotic fluid and exhalation of the same fluid, with pulmonary surfactant and mucus. Foetal breathing movements are not continuous and may include periods of frequent movements and periods of no movements. Maternal factors can influence the frequency of breathing movements. For example, high blood glucose levels, called hyperglycaemia, can boost the number of breathing movements. Conversely, low blood glucose levels, called hypoglycaemia, can reduce the number of foetal breathing movements. Tobacco use is also known to lower foetal breathing rates. foetal breathing may help tone the muscles in preparation for breathing movements once the foetus is born. It may also help the alveoli to form and mature. Foetal breathing movements are considered a sign of robust health.
Birth
Prior to birth, the lungs are filled with amniotic fluid, mucus, and surfactant. As the foetus is squeezed through the birth canal, the foetal thoracic cavity is compressed, expelling much of this fluid. Some fluid remains, however, but is rapidly absorbed by the body shortly after birth. The first inhalation occurs within 10 seconds after birth and not only serves as the first inspiration, but also acts to inflate the lungs. However, at birth the alveoli are not fully develop, this process will continue throughout the first two years of a child’s life. Pulmonary surfactant is critical for inflation to occur, as it reduces the surface tension of the alveoli. Preterm birth around 26 weeks frequently results in severe respiratory distress, although with current medical advancements, some babies may survive. Prior to 26 weeks, sufficient pulmonary surfactant is not produced, and the surfaces for gas exchange have not formed adequately; therefore, survival is low.
Disorders of the Respiratory System: Respiratory Distress Syndrome
Respiratory distress syndrome (RDS) (or infant respiratory distress syndrome IRDS) primarily occurs in infants born prematurely. Up to 50 percent of infants born between 26 and 28 weeks and fewer than 30 percent of infants born between 30 and 31 weeks develop RDS. RDS results from insufficient production of pulmonary surfactant, thereby preventing the lungs from properly inflating at birth. A small amount of pulmonary surfactant is produced beginning at around 20 weeks; however, this is not sufficient for inflation of the lungs. As a result, dyspnoea occurs and gas exchange cannot be performed properly. Blood oxygen levels are low, whereas blood carbon dioxide levels and pH are high.
The primary cause of RDS is premature birth, which may be due to a variety of known or unknown causes. Other risk factors include gestational diabetes, caesarean delivery, second-born twins, and family history of RDS. The presence of RDS can lead to other serious disorders, such as septicaemia (infection of the blood) or pulmonary haemorrhage. Therefore, it is important that RDS is immediately recognised and treated to prevent death and reduce the risk of developing other disorders.
Medical advances have resulted in an improved ability to treat RDS and support the infant until proper lung development can occur. At the time of delivery, treatment may include resuscitation and intubation if the infant does not breathe on his or her own. These infants would need to be placed on a ventilator to mechanically assist with the breathing process. If spontaneous breathing occurs, application of nasal continuous positive airway pressure (CPAP) may be required. In addition, pulmonary surfactant is typically administered. Death due to RDS has been reduced by 50 percent due to the introduction of pulmonary surfactant therapy. Other therapies may include corticosteroids, supplemental oxygen, and assisted ventilation. Supportive therapies, such as temperature regulation, nutritional support, and antibiotics, may be administered to the premature infant as well.
Section Review
The development of the respiratory system in the foetus begins at about 4 weeks and continues into childhood. Ectodermal tissue in the anterior portion of the head region invaginates posteriorly, forming olfactory pits, which ultimately fuse with endodermal tissue of the early pharynx. At about this same time, a protrusion of endodermal tissue extends anteriorly from the foregut, producing a lung bud, which continues to elongate until it forms the laryngotracheal bud. The proximal portion of this structure will mature into the trachea, whereas the bulbous end will branch to form two bronchial buds. These buds then branch repeatedly, so that at about week 16, all major airway structures are present. Development progresses after week 16 as respiratory bronchioles and alveolar ducts form, and extensive vascularisation occurs. Alveolar type I cells also begin to take shape. Type II pulmonary cells develop and begin to produce small amounts of surfactant. As the foetus grows, the respiratory system continues to expand as more alveoli develop and more surfactant is produced. Beginning at about week 36 and lasting into childhood, alveolar precursors mature to become fully functional alveoli. At birth, compression of the thoracic cavity forces much of the fluid in the lungs to be expelled. The first inhalation inflates the lungs, foetal breathing movements begin around week 20 or 21 and occur when contractions of the respiratory muscles cause the foetus to inhale and exhale amniotic fluid. These movements continue until birth and may help to tone the muscles in preparation for breathing after birth and are a sign of good health.
Muscle is one of the four primary tissue types of the body. The body contains three types of muscle tissue: skeletal muscle, cardiac muscle, and smooth muscle (Figure 9.1.1). All three muscle tissues have some properties in common; they all exhibit a quality called excitability as their plasma membranes can change their electrical states (from polarised to depolarised) and send an electrical wave called an action potential along the entire length of the membrane. While the nervous system can influence the excitability of cardiac and smooth muscle to some degree, skeletal muscle completely depends on signalling from the nervous system to work properly. On the other hand, both cardiac muscle and smooth muscle can respond to other stimuli, such as hormones and local stimuli.
The muscles all begin the actual process of contracting (shortening) when a protein called actin is pulled by a protein called myosin. This occurs in striated muscle (skeletal and cardiac) after specific binding sites on the actin have been exposed in response to the interaction between calcium ions (Ca2+) and proteins (troponin and tropomyosin) that “shield” the actin-binding sites. Ca2+ also is required for the contraction of smooth muscle, although its role is different: here Ca2+ activates enzymes, which in turn activate myosin heads. All muscles require adenosine triphosphate (ATP) to continue the process of contracting, and they all relax when the Ca2+ is removed and the actin-binding sites are re-shielded.
A muscle can return to its original length when relaxed due to a quality of muscle tissue called elasticity. It can recoil back to its original length due to elastic fibres. Muscle tissue also has the quality of extensibility; it can stretch or extend. Contractility allows muscle tissue to pull on its attachment points and shorten with force.
Differences among the three muscle types include the microscopic organisation of their contractile proteins—actin and myosin. The actin and myosin proteins are arranged very regularly in the cytoplasm of individual muscle cells (referred to as fibres) in both skeletal muscle and cardiac muscle, which creates a pattern, or stripes, called striations. The striations are visible with a light microscope under high magnification (see Figure 9.1.1). Skeletal muscle fibres are multinucleated structures that compose the skeletal muscle. Cardiac muscle fibres each have one to two nuclei and are physically and electrically connected to each other so that the entire heart contracts as one unit (called a syncytium).
Because the actin and myosin are not arranged in such regular fashion in smooth muscle, the cytoplasm of a smooth muscle fibre (which has only a single nucleus) has a uniform, nonstriated appearance (resulting in the name smooth muscle). However, the less organised appearance of smooth muscle should not be interpreted as less efficient. Smooth muscle in the walls of arteries is a critical component that regulates blood pressure necessary to push blood through the circulatory system; and smooth muscle in the skin, visceral organs, and internal passageways is essential for moving all materials through the body.
Section Review
Muscle is the tissue in animals that allows for active movement of the body or materials within the body. There are three types of muscle tissue: skeletal muscle, cardiac muscle, and smooth muscle. Most of the body’s skeletal muscle produces movement by acting on the skeleton. Cardiac muscle is found in the wall of the heart and pumps blood through the circulatory system.
Smooth muscle is found in the skin, where it is associated with hair follicles; it also is found in the walls of internal organs, blood vessels and internal passageways, where it assists in moving materials.
Describe the layers of connective tissues packaging skeletal muscle
Explain how muscles work with tendons to move the body
Identify areas of the skeletal muscle fibres
Describe excitation-contraction coupling
The best-known feature of skeletal muscle is its ability to contract and cause movement. Skeletal muscles act not only to produce movement but also to stop movement, such as resisting gravity to maintain posture. Small, constant adjustments of the skeletal muscles are needed to hold a body upright or balanced in any position. Muscles also prevent excess movement of the bones and joints, maintaining skeletal stability and preventing skeletal structure damage or deformation. Joints can become misaligned or dislocated entirely by pulling on the associated bones; muscles work to keep joints stable. Skeletal muscles are located throughout the body at the openings of internal tracts to control the movement of various substances. These muscles allow functions, such as swallowing, urination, and defecation, to be under voluntary control. Skeletal muscles also protect internal organs (particularly abdominal and pelvic organs) by acting as an external barrier or shield to external trauma and by supporting the weight of the organs.
Skeletal muscles contribute to the maintenance of homeostasis in the body by generating heat. Muscle contraction requires energy, and when ATP is broken down, heat is produced. This heat is very noticeable during exercise, when sustained muscle movement causes body temperature to rise, and in cases of extreme cold, when shivering produces random skeletal muscle contractions to generate heat.
Each skeletal muscle is an organ that consists of various integrated tissues. These tissues include the skeletal muscle fibres, blood vessels, nerve fibres, and connective tissue. Each skeletal muscle has three layers of connective tissue (called “mysia”) that enclose it and provide structure to the muscle as a whole, and compartmentalise the muscle fibres within the muscle (Figure 9.2.1). Each muscle is wrapped in a sheath of dense, irregular connective tissue called the epimysium, which allows a muscle to contract and move powerfully while maintaining its structural integrity. The epimysium also separates muscle from other tissues and organs in the area, allowing the muscle to move independently.
Figure 9.2.1. The Three connective tissue layers. Bundles of muscle fibres, called fascicles, are covered by the perimysium. Muscle fibres are covered by the endomysium.
Inside each skeletal muscle, muscle fibres are organised into individual bundles, each called a fascicle, by a middle layer of connective tissue called the perimysium. This fascicular organisation is common in muscles of the limbs; it allows the nervous system to trigger a specific movement of a muscle by activating a subset of muscle fibres within a bundle, or fascicle of the muscle. Inside each fascicle, each muscle fibre is encased in a thin connective tissue layer of collagen and reticular fibres called the endomysium. The endomysium contains the extracellular fluid and nutrients to support the muscle fibre. These nutrients are supplied via blood to the muscle tissue.
In skeletal muscles that work with tendons to pull on bones, the collagen in the three tissue layers (the mysia) intertwines with the collagen of a tendon. At the other end of the tendon, it fuses with the periosteum coating the bone. The tension created by contraction of the muscle fibres is then transferred though the mysia, to the tendon, and then to the periosteum to pull on the bone for movement of the skeleton. In other places, the mysia may fuse with a broad, tendon-like sheet called an aponeurosis, or fascia, the connective tissue between skin and bones. The broad sheet of connective tissue in the lower back that the latissimus dorsi muscles (the “lats”) fuse into is an example of an aponeurosis.
Every skeletal muscle is also richly supplied by blood vessels for nourishment, oxygen delivery, and waste removal. In addition, every muscle fibre in a skeletal muscle is supplied by the axon branch of a somatic motor neuron, which signals the fibre to contract. Unlike cardiac and smooth muscle, the only way to functionally contract a skeletal muscle is through signalling from the nervous system.
Skeletal Muscle Fibres
Because skeletal muscle cells are long and cylindrical, they are commonly referred to as muscle fibres. Skeletal muscle fibres can be quite large for human cells, with diameters up to 100 μm and lengths up to 30 cm (11.8 in) in the Sartorius of the upper leg. During early development, embryonic myoblasts, each with its own nucleus, fuse with up to hundreds of other myoblasts to form the multinucleated skeletal muscle fibres. Multiple nuclei mean multiple copies of genes, permitting the production of the large amounts of proteins and enzymes needed for muscle contraction.
Some other terminology associated with muscle fibres is rooted in the Greek sarco, which means “flesh.” The plasma membrane of muscle fibres is called the sarcolemma, the cytoplasm is referred to as sarcoplasm, and the specialised smooth endoplasmic reticulum, which stores, releases, and retrieves calcium ions (Ca2+) is called the sarcoplasmic reticulum (SR) (Figure 9.2.2). As will soon be described, the functional unit of a skeletal muscle fibre is the sarcomere, a highly organised arrangement of the contractile myofilaments actin (thin filament) and myosin (thick filament), along with other support proteins.
Figure 9.2.2. Muscle fibre. A skeletal muscle fibre is surrounded by a plasma membrane called the sarcolemma, which contains sarcoplasm, the cytoplasm of muscle cells. A muscle fibre is composed of many fibrils, which give the cell its striated appearance.
The Sarcomere
The sarcomere is the functional unit of the muscle fibre. The sarcomere itself is bundled within the myofibril that runs the entire length of the muscle fibre and attaches to the sarcolemma at its end. As myofibrils contract, the entire muscle cell contracts. Because myofibrils are only approximately 1.2 μm in diameter, hundreds to thousands (each with thousands of sarcomeres) can be found inside one muscle fibre. Each sarcomere is approximately 2 μm in length with a three-dimensional cylinder-like arrangement and is bordered by structures called Z-discs (also called Z-lines, because pictures are two-dimensional), to which the actin myofilaments are anchored (Figure 9.2.3). Because the actin and its troponin-tropomyosin complex (projecting from the Z-discs toward the centre of the sarcomere) form strands that are thinner than the myosin, it is called the thin filament of the sarcomere. Likewise, because the myosin strands and their multiple heads (projecting from the centre of the sarcomere, toward but not all to way to, the Z-discs) have more mass and are thicker, they are called the thick filament of the sarcomere.
Figure 9.2.3. The sarcomere. The sarcomere, the region from one Z-line to the next Z-line, is the functional unit of a skeletal muscle fibre.
The Neuromuscular Junction
Another specialisation of the skeletal muscle is the site where a motor neuron’s terminal meets the muscle fibre — called the neuromuscular junction (NMJ). This is where the muscle fibre first responds to signalling by the motor neuron. Every skeletal muscle fibre in every skeletal muscle is innervated by a motor neuron at the NMJ. Excitation signals from the neuron are the only way to functionally activate the fibre to contract.
Excitation–Contraction Coupling
All living cells have membrane potentials, or electrical gradients across their membranes. The inside of the membrane is usually around -60 to -90 mV, relative to the outside. This is referred to as a cell’s membrane potential. Neurons and muscle cells can use their membrane potentials to generate electrical signals. They do this by controlling the movement of charged particles, called ions, across their membranes to create electrical currents. This is achieved by opening and closing specialised proteins in the membrane called ion channels. Although the currents generated by ions moving through these channel proteins are very small, they form the basis of both neural signalling and muscle contraction.
Both neurons and skeletal muscle cells are electrically excitable, meaning that they can generate action potentials. An action potential is a special type of electrical signal that can travel along a cell membrane as a wave. This allows a signal to be transmitted quickly and faithfully over long distances.
Although the term excitation-contraction coupling confuses or scares some students, it comes down to this: for a skeletal muscle fibre to contract, its membrane must first be “excited”—in other words, it must be stimulated to fire an action potential. The muscle fibre action potential, which sweeps along the sarcolemma as a wave, is “coupled” to the actual contraction through the release of calcium ions (Ca2+) from the SR. Once released, the Ca2+ interacts with the shielding proteins, forcing them to move aside so that the actin-binding sites are available for attachment by myosin heads. The myosin then pulls the actin filaments toward the centre, shortening the muscle fibre.
In skeletal muscle, this sequence begins with signals from the somatic motor division of the nervous system. In other words, the “excitation” step in skeletal muscles is always triggered by signalling from the nervous system (Figure 9.2.4).
Figure 9.2.4. Motor end-plate and innervation. At the NMJ, the axon terminal releases ACh. The motor end-plate is the location of the ACh-receptors in the muscle fibre sarcolemma. When ACh molecules are released, they diffuse across a minute space called the synaptic cleft and bind to the receptors.
The motor neurons that tell the skeletal muscle fibres to contract originate in the spinal cord, with a smaller number located in the brainstem for activation of skeletal muscles of the face, head, and neck. These neurons have long processes, called axons, which are specialised to transmit action potentials long distances— in this case, all the way from the spinal cord to the muscle itself (which may be up to three feet away). The axons of multiple neurons bundle together to form nerves, like wires bundled together in a cable.
Signalling begins when a neuronal action potential travels along the axon of a motor neuron, and then along the individual branches to terminate at the NMJ. At the NMJ, the axon terminal releases a chemical messenger, or neurotransmitter, called acetylcholine (ACh). The ACh molecules diffuse across a minute space called the synaptic cleft and bind to ACh receptors located within the motor end-plate of the sarcolemma on the other side of the synapse. Once ACh binds, a channel in the ACh receptor opens and positively charged ions can pass through into the muscle fibre, causing it to depolarise, meaning that the membrane potential of the muscle fibre becomes less negative (closer to zero.)
As the membrane depolarises, another set of ion channels called voltage-gated sodium channels are triggered to open. Sodium ions enter the muscle fibre, and an action potential rapidly spreads (or “fires”) along the entire membrane to initiate excitation-contraction coupling.
Things happen very quickly in the world of excitable membranes (just think about how quickly you can snap your fingers as soon as you decide to do it). Immediately following depolarisation of the membrane, it repolarises, re-establishing the negative membrane potential. Meanwhile, the ACh in the synaptic cleft is degraded by the enzyme acetylcholinesterase (AChE) so that the ACh cannot rebind to a receptor and reopen its channel, which would cause unwanted extended muscle excitation and contraction.
Propagation of an action potential along the sarcolemma is the excitation portion of excitation-contraction coupling. Recall that this excitation triggers the release of calcium ions (Ca2+) from its storage in the cell’s SR. For the action potential to reach the membrane of the SR, there are periodic invaginations in the sarcolemma, called T-tubules (“T” stands for “transverse”). You will recall that the diameter of a muscle fibre can be up to 100 μm, so these T-tubules ensure that the membrane can get close to the SR in the sarcoplasm. The arrangement of a T-tubule with the membranes of SR on either side is called a triad (Figure 9.2.5). The triad surrounds the cylindrical structure called a myofibril, which contains actin and myosin.
Figure 9.2.5. The T-tubule. Narrow T-tubules permit the conduction of electrical impulses. The SR functions to regulate intracellular levels of calcium. Two terminal cisternae (where enlarged SR connects to the T-tubule) and one T-tubule comprise a triad—a “threesome” of membranes, with those of SR on two sides and the T-tubule sandwiched between them.
The T-tubules carry the action potential into the interior of the cell, which triggers the opening of calcium channels in the membrane of the adjacent SR, causing Ca2+ to diffuse out of the SR and into the sarcoplasm. It is the arrival of Ca2+ in the sarcoplasm that initiates contraction of the muscle fibre by its contractile units, or sarcomeres.
Section Review
Skeletal muscles contain connective tissue, blood vessels and nerves. There are three layers of connective tissue: epimysium, perimysium, and endomysium. Skeletal muscle fibres are organised into groups called fascicles. Blood vessels and nerves enter the connective tissue and branch in the cell. Muscles attach to bones directly or through tendons or aponeuroses. Skeletal muscles maintain posture, stabilise bones and joints, control internal movement, and generate heat.
Skeletal muscle fibres are long, multinucleated cells. The membrane of the cell is the sarcolemma; the cytoplasm of the cell is the sarcoplasm. The sarcoplasmic reticulum (SR) is a form of endoplasmic reticulum. Muscle fibres are composed of myofibrils. The striations are created by the organisation of actin and myosin resulting in the banding pattern of myofibrils.
Describe the components involved in a muscle contraction
Explain how muscles contract and relax
Describe the sliding filament model of muscle contraction
The sequence of events that result in the contraction of an individual muscle fibre begins with a signal—the neurotransmitter, ACh—from the motor neuron innervating that fibre. The local membrane of the fibre will depolarise as positively charged sodium ions (Na+) enter, triggering an action potential that spreads to the rest of the membrane will depolarise, including the T-tubules. This triggers the release of calcium ions (Ca2+) from storage in the sarcoplasmic reticulum (SR). The Ca2+ then initiates contraction, which is sustained by ATP (Figure 9.3.1). As long as Ca2+ ions remain in the sarcoplasm to bind to troponin, which keeps the actin-binding sites “unshielded,” and as long as ATP is available to drive the cross-bridge cycling and the pulling of actin strands by myosin, the muscle fibre will continue to shorten to an anatomical limit. Muscle contraction usually stops when signalling from the motor neuron ends, which repolarises the sarcolemma and T-tubules, and closes the voltage-gated calcium channels in the SR. Ca2+ ions are then pumped back into the SR, which causes the tropomyosin to re-shield (or re-cover) the binding sites on the actin strands. A muscle also can stop contracting when it runs out of ATP and becomes fatigued (Figure 9.3.6 ).
Figure 9.3.1. Contraction of a muscle fibre. A cross-bridge forms between actin and the myosin heads triggering contraction. As long as Ca2+ ions remain in the sarcoplasm to bind to troponin, and as long as ATP is available, the muscle fibre will continue to shorten.
The molecular events of muscle fibre shortening occur within the fibre’s sarcomeres (see Figure 9.3.2). The contraction of a striated muscle fibre occurs as the sarcomeres, linearly arranged within myofibrils, shorten as myosin heads pull on the actin filaments.
The region where thick and thin filaments overlap has a dense appearance, as there is little space between the filaments. This zone where thin and thick filaments overlap is very important to muscle contraction, as it is the site where filament movement starts. Thin filaments, anchored at their ends by the Z-discs, do not extend completely into the central region that only contains thick filaments, anchored at their bases at a spot called the M-line. A myofibril is composed of many sarcomeres running along its length; thus, myofibrils and muscle cells contract as the sarcomeres contract.
The Sliding Filament Model of Contraction
When signalled by a motor neuron, a skeletal muscle fibre contracts as the thin filaments are pulled and then slide past the thick filaments within the fibre’s sarcomeres. This process is known as the sliding filament model of muscle contraction (Figure 9.3.2). The sliding can only occur when myosin-binding sites on the actin filaments are exposed by a series of steps that begins with Ca2+ entry into the sarcoplasm.
Figure 9.3.2. The Sliding filament model of muscle contraction. When a sarcomere contracts, the Z lines move closer together, and the I band becomes smaller. The A band stays the same width. At full contraction, the thin and thick filaments overlap.
Tropomyosin is a protein that winds around the chains of the actin filament and covers the myosin-binding sites to prevent actin from binding to myosin. Tropomyosin binds to troponin to form a troponin-tropomyosin complex. The troponin-tropomyosin complex prevents the myosin “heads” from binding to the active sites on the actin microfilaments. Troponin also has a binding site for Ca2+ ions.
To initiate muscle contraction, tropomyosin must expose the myosin-binding site on an actin filament to allow cross-bridge formation between the actin and myosin microfilaments. The first step in the process of contraction is for Ca2+ to bind to troponin so that tropomyosin can slide away from the binding sites on the actin strands. This allows the myosin heads to bind to these exposed binding sites and form cross-bridges. The thin filaments are then pulled by the myosin heads to slide past the thick filaments toward the centre of the sarcomere. But each head can only pull a very short distance before it has reached its limit and must be “re-cocked” before it can pull again, a step that requires ATP.
ATP and Muscle Contraction
For thin filaments to continue to slide past thick filaments during muscle contraction, myosin heads must pull the actin at the binding sites, detach, re-cock, attach to more binding sites, pull, detach, re-cock, etc. This repeated movement is known as the cross-bridge cycle. This motion of the myosin heads is similar to the oars when an individual rows a boat: The paddle of the oars (the myosin heads) pull, are lifted from the water (detach), repositioned (re-cocked) and then immersed again to pull (Figure 9.3.3). Each cycle requires energy, and the action of the myosin heads in the sarcomeres repetitively pulling on the thin filaments also requires energy, which is provided by ATP.
Figure 9.3.3. Skeletal muscle contraction. (a) The active site on actin is exposed as calcium binds to troponin. (b) The myosin head is attracted to actin, and myosin binds actin at its actin-binding site, forming the cross-bridge. (c) During the power stroke, the phosphate generated in the previous contraction cycle is released. This results in the myosin head pivoting toward the centre of the sarcomere, after which the attached ADP and phosphate group are released. (d) A new molecule of ATP attaches to the myosin head, causing the cross-bridge to detach. (e) The myosin head hydrolyses ATP to ADP and phosphate, which returns the myosin to the cocked position.
Cross-bridge formation occurs when the myosin head attaches to the actin while adenosine diphosphate (ADP) and inorganic phosphate (Pi) are still bound to myosin (Figure 9.3.3a,b). Pi is then released, causing myosin to form a stronger attachment to the actin, after which the myosin head moves toward the M-line, pulling the actin along with it. As actin is pulled, the filaments move approximately 10 nm toward the M-line. This movement is called the power stroke, as movement of the thin filament occurs at this step (Figure 9.3.3c). In the absence of ATP, the myosin head will not detach from actin.
One part of the myosin head attaches to the binding site on the actin, but the head has another binding site for ATP. ATP binding causes the myosin head to detach from the actin (Figure 9.3.3d). After this occurs, ATP is converted to ADP and Pi by the intrinsic ATPase activity of myosin. The energy released during ATP hydrolysis changes the angle of the myosin head into a cocked position (Figure 9.3.3e). The myosin head is now in position for further movement.
When the myosin head is cocked, myosin is in a high-energy configuration. This energy is expended as the myosin head moves through the power stroke, and at the end of the power stroke, the myosin head is in a low-energy position. After the power stroke, ADP is released; however, the formed cross-bridge is still in place, and actin and myosin are bound together. If ATP is available, it readily attaches to myosin, the cross-bridge cycle can recur, and muscle contraction can continue.
Note that each thick filament of roughly 300 myosin molecules has multiple myosin heads, and many cross-bridges form and break continuously during muscle contraction. Multiply this by all of the sarcomeres in one myofibril, all the myofibrils in one muscle fibre, and all of the muscle fibres in one skeletal muscle, and you can understand why so much energy (ATP) is needed to keep skeletal muscles working. In fact, it is the loss of ATP that results in the rigor mortis observed soon after someone dies. With no further ATP production possible, there is no ATP available for myosin heads to detach from the actin-binding sites, so the cross-bridges stay in place, causing the rigidity in the skeletal muscles.
Sources of ATP
ATP supplies the energy for muscle contraction to take place. In addition to its direct role in the cross-bridge cycle, ATP also provides the energy for the active-transport Ca2+ pumps in the SR. Muscle contraction does not occur without enough ATP. The amount of ATP stored in muscle is very low, only sufficient to power a few seconds worth of contractions. As it is broken down, ATP must therefore be regenerated and replaced quickly to allow for sustained contraction. There are three mechanisms by which ATP can be regenerated: creatine phosphate metabolism, anaerobic glycolysis, fermentation and aerobic respiration.
Creatine phosphate is a molecule that can store energy in its phosphate bonds. In a resting muscle, excess ATP transfers its energy to creatine, producing ADP and creatine phosphate. This acts as an energy reserve that can be used to quickly create more ATP. When the muscle starts to contract and needs energy, creatine phosphate transfers its phosphate back to ADP to form ATP and creatine. This reaction is catalysed by the enzyme creatine kinase and occurs very quickly; thus, creatine phosphate-derived ATP powers the first few seconds of muscle contraction. However, creatine phosphate can only provide approximately 15 seconds worth of energy, at which point another energy source has to be used (Figure 9.3.4).
Figure 9.3.4. Muscle metabolism. (a) Some ATP is stored in a resting muscle. As contraction starts, it is used up in seconds. More ATP is generated from creatine phosphate for about 15 seconds. (b) Each glucose molecule produces two ATP and two molecules of pyruvic acid, which can be used in aerobic respiration or converted to lactic acid. If oxygen is not available, pyruvic acid is converted to lactic acid, which may contribute to muscle fatigue. This occurs during strenuous exercise when high amounts of energy are needed but oxygen cannot be sufficiently delivered to muscle. (c) Aerobic respiration is the breakdown of glucose in the presence of oxygen (O2) to produce carbon dioxide, water, and ATP. Approximately 95 percent of the ATP required for resting or moderately active muscles is provided by aerobic respiration, which takes place in mitochondria.
As the ATP produced by creatine phosphate is depleted, muscles turn to glycolysis as an ATP source. Glycolysis is an anaerobic (non-oxygen-dependent) process that breaks down glucose (sugar) to produce ATP; however, glycolysis cannot generate ATP as quickly as creatine phosphate. Thus, the switch to glycolysis results in a slower rate of ATP availability to the muscle. The sugar used in glycolysis can be provided by blood glucose or by metabolising glycogen that is stored in the muscle. The breakdown of one glucose molecule produces two ATP and two molecules of pyruvic acid, which can be used in aerobic respiration or when oxygen levels are low, converted to lactic acid (Figure 9.3.4b).
If oxygen is available, pyruvic acid is used in aerobic respiration. However, if oxygen is not available, pyruvic acid is converted to lactic acid, which may contribute to muscle fatigue. This conversion allows the recycling of the enzyme NAD+ from NADH, which is needed for glycolysis to continue. This occurs during strenuous exercise when high amounts of energy are needed but oxygen cannot be sufficiently delivered to muscle. Glycolysis itself cannot be sustained for very long (approximately 1 minute of muscle activity), but it is useful in facilitating short bursts of high-intensity output. This is because glycolysis does not utilise glucose very efficiently, producing a net gain of two ATPs per molecule of glucose, and the end-product of lactic acid, which may contribute to muscle fatigue as it accumulates.
Aerobic respiration is the breakdown of glucose or other nutrients in the presence of oxygen (O2) to produce carbon dioxide, water, and ATP. Approximately 95 percent of the ATP required for resting or moderately active muscles is provided by aerobic respiration, which takes place in mitochondria. The inputs for aerobic respiration include glucose circulating in the bloodstream, pyruvic acid, and fatty acids. Aerobic respiration is much more efficient than anaerobic glycolysis, producing approximately 36 ATPs per molecule of glucose versus four from glycolysis. However, aerobic respiration cannot be sustained without a steady supply of O2 to the skeletal muscle and is much slower (Figure 9.3.4c). To compensate, muscles store small amount of excess oxygen in proteins call myoglobin, allowing for more efficient muscle contractions and less fatigue. Aerobic training also increases the efficiency of the circulatory system so that O2 can be supplied to the muscles for longer periods of time.
Muscle fatigue occurs when a muscle can no longer contract in response to signals from the nervous system. The exact causes of muscle fatigue are not fully known, although certain factors have been correlated with the decreased muscle contraction that occurs during fatigue. ATP is needed for normal muscle contraction, and as ATP reserves are reduced, muscle function may decline. This may be more of a factor in brief, intense muscle output rather than sustained, lower intensity efforts. Lactic acid build-up may lower intracellular pH, affecting enzyme and protein activity. Imbalances in Na+ and K+ levels as a result of membrane depolarisation may disrupt Ca2+ flow out of the SR. Long periods of sustained exercise may damage the SR and the sarcolemma, resulting in impaired Ca2+ regulation.
Intense muscle activity results in an oxygen debt, which is the amount of oxygen needed to compensate for ATP produced without oxygen during muscle contraction. Oxygen is required to restore ATP and creatine phosphate levels, convert lactic acid to pyruvic acid, and, in the liver, to convert lactic acid into glucose or glycogen. Other systems used during exercise also require oxygen, and all these combined processes result in the increased breathing rate that occurs after exercise. Until the oxygen debt has been met, oxygen intake is elevated, even after exercise has stopped.
Relaxation of a Skeletal Muscle
Relaxing skeletal muscle fibres, and ultimately, the skeletal muscle, begins with the motor neuron, which stops releasing its chemical signal, ACh, into the synapse at the NMJ. The muscle fibre will repolarise, which closes the gates in the SR where Ca2+ was being released. ATP-driven pumps will move Ca2+ out of the sarcoplasm back into the SR. This results in the “re-shielding” of the actin-binding sites on the thin filaments. Without the ability to form cross-bridges between the thin and thick filaments, the muscle fibre loses its tension and relaxes.
Figure 9.3.5. Relaxation of a muscle fibre. Ca2+ ions are pumped back into the SR, which causes the tropomyosin to re-shield the binding sites on the actin strands. A muscle may also stop contracting when it runs out of ATP and becomes fatigued.
Muscle Strength
The number of skeletal muscle fibres in each muscle is genetically determined and does not change. Muscle strength is directly related to the number of myofibrils and sarcomeres within each fibre. Factors, such as hormones and stress (and artificial anabolic steroids), acting on the muscle can increase the production of sarcomeres and myofibrils within the muscle fibres, a change called hypertrophy, which results in the increased mass and bulk in a skeletal muscle. Likewise, decreased use of a skeletal muscle results in atrophy, where the number of sarcomeres and myofibrils disappear (but not the number of muscle fibres). It is common for a limb in a cast to show atrophied muscles when the cast is removed and certain diseases, such as polio, show atrophied muscles.
Disorders of the Muscular System
Duchenne muscular dystrophy (DMD) is a progressive weakening of the skeletal muscles which affects 1:3500 male newborns worldwide. It is one of several diseases collectively referred to as “muscular dystrophy.” DMD is caused by a lack of the protein dystrophin, which helps the thin filaments of myofibrils bind to the sarcolemma. Without sufficient dystrophin, muscle contractions cause the sarcolemma to tear, causing an influx of Ca2+, leading to cellular damage and muscle fibre degradation. Over time, as muscle damage accumulates, muscle mass is lost, and greater functional impairments develop.
DMD is an inherited disorder caused by an abnormal X chromosome. It primarily affects males, and it is usually diagnosed in early childhood. DMD usually first appears as difficulty with balance and motion, and then progresses to an inability to walk. It continues progressing upward in the body from the lower extremities to the upper body, where it affects the muscles responsible for breathing and circulation. It ultimately causes death due to respiratory failure, and those afflicted do not usually live past their 20s.
Because DMD is caused by a mutation in the gene that codes for dystrophin, it was thought that introducing healthy myoblasts into patients might be an effective treatment. Myoblasts are the embryonic cells responsible for muscle development, and ideally, they would carry healthy genes that could produce the dystrophin needed for normal muscle contraction. This approach has been largely unsuccessful in humans. A recent approach has involved attempting to boost the muscle’s production of utrophin, a protein like dystrophin that may be able to assume the role of dystrophin and prevent cellular damage from occurring.
Section Review
A sarcomere is the smallest contractile portion of a muscle. Myofibrils are composed of thick and thin filaments. Thick filaments are composed of the protein myosin; thin filaments are composed of the protein actin. Troponin and tropomyosin are regulatory proteins.
Muscle contraction is described by the sliding filament model of contraction. ACh is the neurotransmitter that binds at the neuromuscular junction (NMJ) to trigger depolarisation, and an action potential travels along the sarcolemma to trigger calcium release from SR. The actin sites are exposed after Ca2+ enters the sarcoplasm from its SR storage to activate the troponin-tropomyosin complex so that the tropomyosin shifts away from the sites. The cross-bridging of myosin heads docking into actin-binding sites is followed by the “power stroke”—the sliding of the thin filaments by thick filaments. The power strokes are powered by ATP. Ultimately, the sarcomeres, myofibrils, and muscle fibres shorten to produce movement.
Explain concentric, isotonic and eccentric contractions
Describe the length-tension relationship
Describe the three phases of a muscle twitch
Define wave summation, tetanus and treppe
To move an object, referred to as load, the sarcomeres in the muscle fibres of the skeletal muscle must shorten. The force generated by the contraction of the muscle (or shortening of the sarcomeres) is called muscle tension. However, muscle tension also is generated when the muscle is contracting against a load that does not move, resulting in two main types of skeletal muscle contractions: isotonic contractions and isometric contractions.
In isotonic contractions, where the tension in the muscle stays constant, a load is moved as the length of the muscle changes (shortens). There are two types of isotonic contractions: concentric and eccentric. A concentric contraction involves the muscle shortening to move a load. An example of this is the biceps brachii muscle contracting when a hand weight is brought upward with increasing muscle tension. As the biceps brachii contract, the angle of the elbow joint decreases as the forearm is brought toward the body. Here, the biceps brachii contracts as sarcomeres in its muscle fibres are shortening and cross-bridges form; the myosin heads pull the actin. An eccentric contraction occurs as the muscle tension diminishes and the muscle lengthens. In this case, the hand weight is lowered in a slow and controlled manner as the amount of cross-bridges being activated by nervous system stimulation decreases. In this case, as tension is released from the biceps brachii, the angle of the elbow joint increases. Eccentric contractions are also used for movement and balance of the body.
An isometric contraction occurs as the muscle produces tension without changing the angle of a skeletal joint. Isometric contractions involve sarcomere shortening and increasing muscle tension, but do not move a load, as the force produced cannot overcome the resistance provided by the load. For example, if one attempts to lift a hand weight that is too heavy, there will be sarcomere activation and shortening to a point, and ever-increasing muscle tension, but no change in the angle of the elbow joint. In everyday living, isometric contractions are active in maintaining posture and maintaining bone and joint stability. However, holding your head in an upright position occurs not because the muscles cannot move the head, but because the goal is to remain stationary and not produce movement. Most actions of the body are the result of a combination of isotonic and isometric contractions working together to produce a wide range of outcomes (Figure 9.4.1).
Figure 9.4.1. Types of muscle contractions. During isotonic contractions, muscle length changes to move a load. During isometric contractions, muscle length does not change because the load exceeds the tension the muscle can generate.
All these muscle activities are under the exquisite control of the nervous system. Neural control regulates concentric, eccentric and isometric contractions, muscle fibre recruitment, and muscle tone. A crucial aspect of nervous system control of skeletal muscles is the role of motor units.
Motor Units
As you have learned, every skeletal muscle fibre must be innervated by the axon terminal of a motor neuron in order to contract. Each muscle fibre is innervated by only one motor neuron. The actual group of muscle fibres in a muscle innervated by a single motor neuron is called a motor unit. The size of a motor unit is variable depending on the nature of the muscle.
A small motor unit is an arrangement where a single motor neuron supplies a small number of muscle fibres in a muscle. Small motor units permit very fine motor control of the muscle. The best example in humans is the small motor units of the extraocular eye muscles that move the eyeballs. There are thousands of muscle fibres in each muscle, but every six or so fibres are supplied by a single motor neuron, as the axons branch to form synaptic connections at their individual NMJs. This allows for exquisite control of eye movements so that both eyes can quickly focus on the same object. Small motor units are also involved in the many fine movements of the fingers and thumb of the hand for grasping, texting, etc.
A large motor unit is an arrangement where a single motor neuron supplies many muscle fibres in a muscle. Large motor units are concerned with simple, or “gross,” movements, such as powerfully extending the knee joint. The best example is the large motor units of the thigh muscles or back muscles, where a single motor neuron will supply thousands of muscle fibres in a muscle, as its axon splits into thousands of branches.
There is a wide range of motor units within many skeletal muscles, which gives the nervous system a wide range of control over the muscle. The small motor units in the muscle will have smaller, lower-threshold motor neurons that are more excitable, firing first to their skeletal muscle fibres, which also tend to be the smallest. Activation of these smaller motor units, results in a relatively small degree of contractile strength (tension) generated in the muscle. As more strength is needed, larger motor units, with bigger, higher-threshold motor neurons are enlisted to activate larger muscle fibres. This increasing activation of motor units produces an increase in muscle contraction known as recruitment. As more motor units are recruited, the muscle contraction grows progressively stronger. In some muscles, the largest motor units may generate a contractile force of 50 times more than the smallest motor units in the muscle. This allows a feather to be picked up using the biceps brachii arm muscle with minimal force, and a heavy weight to be lifted by the same muscle by recruiting the largest motor units.
When necessary, the maximal number of motor units in a muscle can be recruited simultaneously, producing the maximum force of contraction for that muscle, but this cannot last for very long because of the energy requirements to sustain the contraction. To prevent complete muscle fatigue, motor units are generally not all simultaneously active, but instead some motor units rest while others are active, which allows for longer muscle contractions. The nervous system uses recruitment as a mechanism to efficiently utilise a skeletal muscle.
The Length-Tension Range of a Sarcomere
When a skeletal muscle fibre contracts, myosin heads attach to actin to form cross-bridges followed by the thin filaments sliding over the thick filaments as the heads pull the actin, and this results in sarcomere shortening, creating the tension of the muscle contraction. The cross-bridges can only form where thin and thick filaments already overlap, so that the length of the sarcomere has a direct influence on the force generated when the sarcomere shortens. This is called the length-tension relationship.
The ideal length of a sarcomere to produce maximal tension occurs at 80 percent to 120 percent of its resting length, with 100 percent being the state where the medial edges of the thin filaments are just at the most-medial myosin heads of the thick filaments (Figure 9.4.2). This length maximises the overlap of actin-binding sites and myosin heads. If a sarcomere is stretched past this ideal length (beyond 120 percent), thick and thin filaments do not overlap sufficiently, which results in less tension produced. If a sarcomere is shortened beyond 80 percent, the zone of overlap is reduced with the thin filaments jutting beyond the last of the myosin heads and shrinks the H zone, which is normally composed of myosin tails. Eventually, there is nowhere else for the thin filaments to go and the amount of tension is diminished. If the muscle is stretched to the point where thick and thin filaments do not overlap at all, no cross-bridges can be formed, and no tension is produced in that sarcomere. This amount of stretching does not usually occur, as accessory proteins and connective tissue oppose extreme stretching.
Figure 9.4.2. The ideal length of a sarcomere. Sarcomeres produce maximal tension when thick and thin filaments overlap between about 80 percent to 120 percent.
The Frequency of Motor Neuron Stimulation
A single action potential from a motor neuron will produce a single contraction in the muscle fibres of its motor unit. This isolated contraction is called a twitch. A twitch can last for a few milliseconds or 100 milliseconds, depending on the muscle type. The tension produced by a single twitch can be measured by a myogram, an instrument that measures the amount of tension produced over time (Figure 9.4.3). Each twitch undergoes three phases. The first phase is the latent period, during which the action potential is being propagated along the sarcolemma and Ca2+ ions are released from the SR. This is the phase during which excitation and contraction are being coupled but contraction has yet to occur. The contraction phase occurs next. The Ca2+ ions in the sarcoplasm have bound to troponin, tropomyosin has shifted away from actin-binding sites, cross-bridges formed, and sarcomeres are actively shortening to the point of peak tension. The last phase is the relaxation phase, when tension decreases as contraction stops. Ca2+ ions are pumped out of the sarcoplasm into the SR, and cross-bridge cycling stops, returning the muscle fibres to their resting state.
Figure 9.4.3. A myogram of a muscle twitch. A single muscle twitch has a latent period, a contraction phase when tension increases, and a relaxation phase when tension decreases. During the latent period, the action potential is being propagated along the sarcolemma. During the contraction phase, Ca2+ ions in the sarcoplasm bind to troponin, tropomyosin moves from actin-binding sites, cross-bridges form, and sarcomeres shorten. During the relaxation phase, tension decreases as Ca2+ ions are pumped out of the sarcoplasm and cross-bridge cycling stops.
Although a person can experience a muscle “twitch,” a single twitch does not produce any significant muscle activity in a living body. A series of action potentials to the muscle fibres is necessary to produce a muscle contraction that can produce work. Normal muscle contraction is more sustained, and it can be modified by input from the nervous system to produce varying amounts of force; this is called a graded muscle response. The frequency of action potentials (nerve impulses) from a motor neuron and the number of motor neurons transmitting action potentials both affect the tension produced in skeletal muscle.
The rate at which a motor neuron fires action potentials affects the tension produced in the skeletal muscle. If the fibres are stimulated while a previous twitch is still occurring, the second twitch will be stronger. This response is called wave summation, because the excitation-contraction coupling effects of successive motor neuron signalling is summed, or added together (Figure 9.4.4a). At the molecular level, summation occurs because the second stimulus triggers the release of more Ca2+ ions, which become available to activate additional sarcomeres while the muscle is still contracting from the first stimulus. Summation results in greater contraction of the motor unit.
Figure 9.4.4. Wave summation and tetanus. (a) The excitation-contraction coupling effects of successive motor neuron signalling is added together which is referred to as wave summation. The bottom of each wave, the end of the relaxation phase, represents the point of stimulus. (b) When the stimulus frequency is so high that the relaxation phase disappears completely, the contractions become continuous; this is called tetanus.
If the frequency of motor neuron signalling increases, summation and subsequent muscle tension in the motor unit continues to rise until it reaches a peak point. The tension at this point is about three to four times greater than the tension of a single twitch, a state referred to as incomplete tetanus. During incomplete tetanus, the muscle goes through quick cycles of contraction with a short relaxation phase for each. If the stimulus frequency is so high that the relaxation phase disappears completely, contractions become continuous in a process called complete tetanus (Figure 9.4.4b).
During tetanus, the concentration of Ca2+ ions in the sarcoplasm allows virtually all of the sarcomeres to form cross-bridges and shorten, so that a contraction can continue uninterrupted (until the muscle fatigues and can no longer produce tension).
Treppe
When a skeletal muscle has been dormant for an extended period and then activated to contract, with all other things being equal, the initial contractions generate about one-half the force of later contractions. The muscle tension increases in a graded manner that to some looks like a set of stairs. This tension increase is called treppe, a condition where muscle contractions become more efficient. It’s also known as the “staircase effect” (Figure 9.4.5).
Figure 9.4.5. Treppe. When muscle tension increases in a graded manner that looks like a set of stairs, it is called treppe. The bottom of each wave represents the point of stimulus.
It is believed that treppe results from a higher concentration of Ca2+ in the sarcoplasm resulting from the steady stream of signals from the motor neuron. It can only be maintained with adequate ATP.
Muscle Tone
Skeletal muscles are rarely completely relaxed, or flaccid. Even if a muscle is not producing movement, it is contracted a small amount to maintain its contractile proteins and produce muscle tone. The tension produced by muscle tone allows muscles to continually stabilise joints and maintain posture.
Muscle tone is accomplished by a complex interaction between the nervous system and skeletal muscles that results in the activation of a few motor units at a time, most likely in a cyclical manner. In this manner, muscles never fatigue completely, as some motor units can recover while others are active.
The absence of the low-level contractions that lead to muscle tone is referred to as hypotonia or atrophy and can result from damage to parts of the central nervous system (CNS), such as the cerebellum, or from loss of innervations to a skeletal muscle, as in poliomyelitis. Hypotonic muscles have a flaccid appearance and display functional impairments, such as weak reflexes. Conversely, excessive muscle tone is referred to as hypertonia, accompanied by hyperreflexia (excessive reflex responses), often the result of damage to upper motor neurons in the CNS. Hypertonia can present with muscle rigidity (as seen in Parkinson’s disease) or spasticity, a phasic change in muscle tone, where a limb will “snap” back from passive stretching (as seen in some strokes).
Section Review
The number of cross-bridges formed between actin and myosin determines the amount of tension produced by a muscle. The length of a sarcomere is optimal when the zone of overlap between thin and thick filaments is greatest. Muscles that are stretched or compressed too greatly do not produce maximal amounts of power. A motor unit is formed by a motor neuron and all the muscle fibres that are innervated by that same motor neuron. A single contraction is called a twitch. A muscle twitch has a latent period, a contraction phase, and a relaxation phase. A graded muscle response allows variation in muscle tension. Summation occurs as successive stimuli are added together to produce a stronger muscle contraction. Tetanus is the fusion of contractions to produce a continuous contraction. Increasing the number of motor neurons involved increases the amount of motor units activated in a muscle, which is called recruitment. Muscle tone is the constant low-level contractions that allow for posture and stability.
Two criteria to consider when classifying the types of muscle fibres are how fast some fibres contract relative to others, and how fibres produce ATP. Using these criteria, there are three main types of skeletal muscle fibres. Slow oxidative (SO) fibres contract relatively slowly and use aerobic respiration (oxygen and glucose) to produce ATP. Fast oxidative (FO) fibres have fast contractions and primarily use aerobic respiration, but because they may switch to anaerobic respiration (glycolysis), can fatigue more quickly than SO fibres. Lastly, fast glycolytic (FG) fibres have fast contractions and primarily use anaerobic glycolysis. The FG fibres fatigue more quickly than the others. Most skeletal muscles in a human contain(s) all three types, although in varying proportions.
The speed of contraction is dependent on how quickly myosin’s ATPase hydrolyses ATP to produce cross-bridge action. Fast fibres hydrolyse ATP approximately twice as quickly as slow fibres, resulting in much quicker cross-bridge cycling (which pulls the thin filaments toward the centre of the sarcomeres at a faster rate). The primary metabolic pathway used by a muscle fibre determines whether the fibre is classified as oxidative or glycolytic. If a fibre primarily produces ATP through aerobic pathways it is oxidative. More ATP can be produced during each metabolic cycle, making the fibre more resistant to fatigue. Glycolytic fibres primarily create ATP through anaerobic glycolysis, which produces less ATP per cycle. As a result, glycolytic fibres fatigue at a quicker rate.
The oxidative fibres contain many more mitochondria than the glycolytic fibres, because aerobic metabolism, which uses oxygen (O2) in the metabolic pathway, occurs in the mitochondria. The SO fibres possess many mitochondria and can contract for longer periods because of the large amount of ATP they can produce, but they have a relatively small diameter and do not produce a large amount of tension. SO fibres are extensively supplied with blood capillaries to supply O2 from the red blood cells in the bloodstream. The SO fibres also possess myoglobin, an O2-carrying molecule like O2-carrying haemoglobin in the red blood cells. The myoglobin stores some of the needed O2 within the fibres themselves (and gives SO fibres their red colour). All these features allow SO fibres to produce large quantities of ATP, which can sustain muscle activity without fatiguing for long periods of time.
The fact that SO fibres can function for long periods without fatiguing makes them useful in maintaining posture, producing isometric contractions, stabilising bones and joints, and making small movements that happen often but do not require large amounts of energy. They do not produce high tension, and thus they are not used for powerful, fast movements that require high amounts of energy and rapid cross-bridge cycling.
FO fibres are sometimes called intermediate fibres because they possess characteristics that are intermediate between fast fibres and slow fibres. They produce ATP relatively quickly, more quickly than SO fibres, and thus can produce relatively high amounts of tension. They are oxidative because they produce ATP aerobically, possess high amounts of mitochondria, and do not fatigue quickly. However, FO fibres do not possess significant myoglobin, giving them a lighter colour than the red SO fibres. FO fibres are used primarily for movements, such as walking, that require more energy than postural control but less energy than an explosive movement, such as sprinting. FO fibres are useful for this type of movement because they produce more tension than SO fibres but they are more fatigue-resistant than FG fibres.
FG fibres primarily use anaerobic glycolysis as their ATP source. They have a large diameter and possess high amounts of glycogen, which is used in glycolysis to generate ATP quickly to produce high levels of tension. Because they do not primarily use aerobic metabolism, they do not possess substantial numbers of mitochondria or significant amounts of myoglobin and therefore have a white colour. FG fibres are used to produce rapid, forceful contractions to make quick, powerful movements. These fibres fatigue quickly, permitting them to only be used for short periods. Most muscles possess a mixture of each fibre type. The predominant fibre type in a muscle is determined by the primary function of the muscle.
Section Review
ATP provides the energy for muscle contraction. The three mechanisms for ATP regeneration are creatine phosphate, anaerobic glycolysis, and aerobic metabolism. Creatine phosphate provides about the first 15 seconds of ATP at the beginning of muscle contraction. Anaerobic glycolysis produces small amounts of ATP in the absence of oxygen for a short period. Aerobic metabolism utilises oxygen to produce much more ATP, allowing a muscle to work for longer periods. Muscle fatigue, which has many contributing factors, occurs when muscle can no longer contract. An oxygen debt is created as a result of muscle use. The three types of muscle fibre are slow oxidative (SO), fast oxidative (FO) and fast glycolytic (FG). SO fibres use aerobic metabolism to produce low power contractions over long periods and are slow to fatigue. FO fibres use aerobic metabolism to produce ATP but produce higher tension contractions than SO fibres. FG fibres use anaerobic metabolism to produce powerful, high-tension contractions but fatigue quickly.
Explain how resistance exercise builds skeletal muscle
Explain how performance-enhancing substances affect muscle
Physical training alters the appearance of skeletal muscles and can produce changes in muscle performance. Conversely, a lack of use can result in decreased performance and muscle appearance. Although muscle cells can change in size, new cells are not formed when muscles grow. Instead, structural proteins are added to muscle fibres in a process called hypertrophy, so cell diameter increases. The reverse, when structural proteins are lost and muscle mass decreases, is called atrophy. Age-related muscle atrophy is called sarcopenia. Cellular components of muscles can also undergo changes in response to changes in muscle use.
Endurance Exercise
Slow fibres are predominantly used in endurance exercises that require little force but involve numerous repetitions. The aerobic metabolism used by slow-twitch fibres allows them to maintain contractions over long periods. Endurance training modifies these slow fibres to make them even more efficient by producing more mitochondria to enable more aerobic metabolism and more ATP production. Endurance exercise can also increase the amount of myoglobin in a cell, as increased aerobic respiration increases the need for oxygen. Myoglobin is found in the sarcoplasm and acts as an oxygen storage supply for the mitochondria.
The training can trigger the formation of more extensive capillary networks around the fibre, a process called angiogenesis, to supply oxygen and remove metabolic waste. To allow these capillary networks to supply the deep portions of the muscle, muscle mass does not greatly increase in order to maintain a smaller area for the diffusion of nutrients and gases. All these cellular changes result in the ability to sustain low levels of muscle contractions for greater periods without fatiguing.
The proportion of SO muscle fibres in muscle determines the suitability of that muscle for endurance and may benefit those participating in endurance activities. Postural muscles have a large number of SO fibres and relatively few FO and FG fibres, to keep the back straight (Figure 9.6.1). Endurance athletes, like marathon-runners also would benefit from a larger proportion of SO fibres, but it is unclear if the most-successful marathoners are those with naturally high numbers of SO fibres, or whether the most successful marathon runners develop high numbers of SO fibres with repetitive training. Endurance training can result in overuse injuries such as stress fractures and joint and tendon inflammation.
Figure 9.6.1. Marathoners. Long-distance runners have many SO fibres and relatively few FO and FG fibres. (credit: “Tseo2”/Wikimedia Commons).
Resistance Exercise
Resistance exercises, as opposed to endurance exercise, require large amounts of FG fibres to produce short, powerful movements that are not repeated over long periods. The high rates of ATP hydrolysis and cross-bridge formation in FG fibres result in powerful muscle contractions. Muscles used for power have a higher ratio of FG to SO/FO fibres, and trained athletes possess even higher levels of FG fibres in their muscles. Resistance exercise affects muscles by increasing the formation of myofibrils, thereby increasing the thickness of muscle fibres. This added structure causes hypertrophy, or the enlargement of muscles, exemplified by the fibres skeletal muscles seen in body builders and other athletes (Figure 9.6.2). Because this muscular enlargement is achieved by the addition of structural proteins, athletes trying to build muscle mass often ingest large amounts of protein.
Figure 9.6.2. Hypertrophy. Body builders have many FG fibres and relatively few FO and SO fibres. (credit: Lin Mei/flickr).
Except for the hypertrophy that follows an increase in the number of sarcomeres and myofibrils in a skeletal muscle, the cellular changes observed during endurance training do not usually occur with resistance training. There is usually no significant increase in mitochondria or capillary density. However, resistance training does increase the development of connective tissue, which adds to the overall mass of the muscle and helps to contain muscles as they produce increasingly powerful contractions. Tendons also become stronger to prevent tendon damage, as the force produced by muscles is transferred to tendons that attach the muscle to bone.
For effective strength training, the intensity of the exercise must continually be increased. For instance, continued weightlifting without increasing the weight of the load does not increase muscle size. To produce ever-greater results, the weights lifted must become increasingly heavier, making it more difficult for muscles to move the load. The muscle then adapts to this heavier load, and an even heavier load must be used if even greater muscle mass is desired.
If done improperly, resistance training can lead to overuse injuries of the muscle, tendon, or bone. These injuries can occur if the load is too heavy or if the muscles are not given sufficient time between workouts to recover or if joints are not aligned properly during the exercises. Cellular damage to muscle fibres that occurs after intense exercise includes damage to the sarcolemma and myofibrils. This muscle damage contributes to the feeling of soreness after strenuous exercise, but muscles gain mass as this damage is repaired, and additional structural proteins are added to replace the damaged ones. Overworking skeletal muscles can also lead to tendon damage and even skeletal damage if the load is too great for the muscles to bear.
Performance-Enhancing Substances
Some athletes attempt to boost their performance by using various agents that may enhance muscle performance. Anabolic steroids are one of the more widely known agents used to boost muscle mass and increase power output. Anabolic steroids are a form of testosterone, a male sex hormone that stimulates muscle formation, leading to increased muscle mass.
Endurance athletes may also try to boost the availability of oxygen to muscles to increase aerobic respiration by using substances such as erythropoietin (EPO), a hormone normally produced in the kidneys, which triggers the production of red blood cells. The extra oxygen carried by these blood cells can then be used by muscles for aerobic respiration. Human growth hormone (hGH) is another supplement, and although it can facilitate building muscle mass, its main role is to promote the healing of muscle and other tissues after strenuous exercise. Increased hGH may allow for faster recovery after muscle damage, reducing the rest required after exercise, and allowing for more sustained high-level performance.
Although performance-enhancing substances often do improve performance, most are banned by governing bodies in sports and are illegal for nonmedical purposes. Their use to enhance performance raises ethical issues of cheating because they give users an unfair advantage over nonusers. A greater concern, however, is that their use carries serious health risks. The side effects of these substances are often significant, non-reversible, and in some cases fatal. The physiological strain caused by these substances is often greater than what the body can handle, leading to effects that are unpredictable and dangerous. Anabolic steroid use has been linked to infertility, aggressive behaviour, cardiovascular disease, and brain cancer.
Similarly, some athletes have used creatine to increase power output. Creatine phosphate provides quick bursts of ATP to muscles in the initial stages of contraction. Increasing the amount of creatine available to cells is thought to produce more ATP and therefore increase explosive power output, although its effectiveness as a supplement has been questioned.
Everyday Connection
Ageing and Muscle Tissue
Although atrophy due to disuse can often be reversed with exercise, muscle atrophy with age, referred to as sarcopenia, is irreversible. This is a primary reason why even highly trained athletes succumb to declining performance with age. This decline is noticeable in athletes whose sports require strength and powerful movements, such as sprinting, whereas the effects of age are less noticeable in endurance athletes such as marathon runners or long-distance cyclists. As muscles age, muscle fibres die, and they are replaced by connective tissue and adipose tissue (Figure 9.6.3). Because those tissues cannot contract and generate force as muscle can, muscles lose the ability to produce powerful contractions. The decline in muscle mass causes a loss of strength, including the strength required for posture and mobility. This may be caused by a reduction in FG fibres that hydrolyse ATP quickly to produce short, powerful contractions. Muscles in older people sometimes possess greater numbers of SO fibres, which are responsible for longer contractions and do not produce powerful movements. There may also be a reduction in the size of motor units, resulting in fewer fibres being stimulated and less muscle tension being produced.
Figure 9.6.3. Atrophy. Muscle mass is reduced as muscles atrophy with disuse.
Sarcopenia can be delayed to some extent by exercise, as training adds structural proteins and causes cellular changes that can offset the effects of atrophy. Increased exercise can produce greater numbers of cellular mitochondria, increase capillary density, and increase the mass and strength of connective tissue. The effects of age-related atrophy are especially pronounced in people who are sedentary, as the loss of muscle cells is displayed as functional impairments such as trouble with locomotion, balance, and posture. This can lead to a decrease in quality of life and medical problems, such as joint problems because the muscles that stabilise bones and joints are weakened. Problems with locomotion and balance can also cause various injuries due to falls.
Section Review
Hypertrophy is an increase in muscle mass due to the addition of structural proteins. The opposite of hypertrophy is atrophy, the loss of muscle mass due to the breakdown of structural proteins. Endurance exercise causes an increase in cellular mitochondria, myoglobin, and capillary networks in SO fibres. Endurance athletes have a high level of SO fibres relative to the other fibre types. Resistance exercise causes hypertrophy. Power-producing muscles have a higher number of FG fibres than of slow fibres. Strenuous exercise causes muscle cell damage that requires time to heal. Some athletes use performance-enhancing substances to enhance muscle performance. Muscle atrophy due to age is called sarcopenia and occurs as muscle fibres die and are replaced by connective and adipose tissue.
Cardiac muscle tissue is only found in the heart. Highly coordinated contractions of cardiac muscle pump blood into the vessels of the circulatory system. Similar to skeletal muscle, cardiac muscle is striated and organised into sarcomeres, possessing the same banding organisation as skeletal muscle (Figure 9.7.1). However, cardiac muscle fibres are shorter than skeletal muscle fibres and usually contain only one nucleus, which is in the central region of the cell. Cardiac muscle fibres also possess many mitochondria and myoglobin, as ATP is produced primarily through aerobic metabolism. Cardiac muscle fibres cells also are extensively branched and are connected to one another at their ends by intercalated discs. An intercalated disc allows the cardiac muscle cells to contract in a wave-like pattern so that the heart can work as a pump.
Intercalated discs are part of the sarcolemma and contain two structures important in cardiac muscle contraction: gap junctions and desmosomes. A gap junction forms channels between adjacent cardiac muscle fibres that allow the depolarising current produced by cations to flow from one cardiac muscle cell to the next. This joining is called electric coupling, and in cardiac muscle it allows the quick transmission of action potentials and the coordinated contraction of the entire heart. This network of electrically connected cardiac muscle cells creates a functional unit of contraction called a syncytium. The remainder of the intercalated disc is composed of desmosomes. A desmosome is a cell structure that anchors the ends of cardiac muscle fibres together so the cells do not pull apart during the stress of individual fibres contracting (Figure 9.7.2).
Figure 9.7.2. Cardiac muscle. Intercalated discs are part of the cardiac muscle sarcolemma and they contain gap junctions and desmosomes.
Contractions of the heart (heartbeats) are controlled by specialised cardiac muscle cells called pacemaker cells that directly control heart rate. Although cardiac muscle cannot be consciously controlled, the pacemaker cells respond to signals from the autonomic nervous system (ANS) to speed up or slow down the heart rate. The pacemaker cells can also respond to various hormones that modulate heart rate to control blood pressure.
The wave of contraction that allows the heart to work as a unit, called a functional syncytium, begins with the pacemaker cells. This group of cells is self-excitable and able to depolarise to threshold and fire action potentials on their own, a feature called autorhythmicity; they do this at set intervals which determine heart rate. Because they are connected with gap junctions to surrounding muscle fibres and the specialised fibres of the heart’s conduction system, the pacemaker cells can transfer the depolarisation to the other cardiac muscle fibres in a manner that allows the heart to contract in a coordinated manner.
Another feature of cardiac muscle is its relatively long action potentials in its fibres, having a sustained depolarisation “plateau.” The plateau is produced by Ca2+ entry though voltage-gated calcium channels in the sarcolemma of cardiac muscle fibres. This sustained depolarisation (and Ca2+ entry) provides for a longer contraction than is produced by an action potential in skeletal muscle. Unlike skeletal muscle, a large percentage of the Ca2+ that initiates contraction in cardiac muscles comes from outside the cell rather than from the SR.
Section Review
Cardiac muscle is striated muscle that is present only in the heart. Cardiac muscle fibres have a single nucleus, are branched, and joined to one another by intercalated discs that contain gap junctions for depolarisation between cells and desmosomes to hold the fibres together when the heart contracts. Contraction in each cardiac muscle fibre is triggered by Ca2+ ions in a similar manner as skeletal muscle, but here the Ca2+ ions come from SR and through voltage-gated calcium channels in the sarcolemma. Pacemaker cells stimulate the spontaneous contraction of cardiac muscle as a functional unit, called a syncytium.
Explain how smooth muscle works with internal organs and passageways through the body
Explain how smooth muscles differ from skeletal and cardiac muscles
Explain the difference between single-unit and multi-unit smooth muscle
Smooth muscle (so-named because the cells do not have striations) is present in the walls of hollow organs like the urinary bladder, uterus, stomach, intestines, and in the walls of passageways, such as the arteries and veins of the circulatory system, and the tracts of the respiratory, urinary, and reproductive systems (Figure 9.8.1ab). Smooth muscle is also present in the eyes, where it functions to change the size of the iris and alter the shape of the lens; and in the skin where it causes hair to stand erect in response to cold temperature or fear.
Smooth muscle fibres are spindle-shaped (wide in the middle and tapered at both ends, somewhat like a football) and have a single nucleus; they range from about 30 to 200 μm (thousands of times shorter than skeletal muscle fibres), and they produce their own connective tissue, endomysium. Although they do not have striations and sarcomeres, smooth muscle fibres do have actin and myosin contractile proteins, and thick and thin filaments. These thin filaments are anchored by dense bodies. A dense body is analogous to the Z-discs of skeletal and cardiac muscle fibres and is fastened to the sarcolemma. Calcium ions are supplied by the SR in the fibres and by sequestration from the extracellular fluid through membrane indentations called calveoli.
Because smooth muscle cells do not contain troponin, cross-bridge formation is not regulated by the troponin-tropomyosin complex but instead by the regulatory protein calmodulin. In a smooth muscle fibre, external Ca2+ ions passing through opened calcium channels in the sarcolemma, and additional Ca2+ released from SR, bind to calmodulin. The Ca2+-calmodulin complex then activates an enzyme called myosin (light chain) kinase, which, in turn, activates the myosin heads by phosphorylating them (converting ATP to ADP and Pi, with the Pi attaching to the head). The heads can then attach to actin-binding sites and pull on the thin filaments. The thin filaments also are anchored to the dense bodies; the structures invested in the inner membrane of the sarcolemma (at adherens junctions) that also have cord-like intermediate filaments attached to them. When the thin filaments slide past the thick filaments, they pull on the dense bodies, structures tethered to the sarcolemma, which then pull on the intermediate filaments networks throughout the sarcoplasm. This arrangement causes the entire muscle fibre to contract in a manner whereby the ends are pulled toward the centre, causing the midsection to bulge in a corkscrew motion (Figure 9.8.2).
Figure 9.8.2. Muscle contraction. The dense bodies and intermediate filaments are networked through the sarcoplasm, which cause the muscle fibre to contract.
Although smooth muscle contraction relies on the presence of Ca2+ ions, smooth muscle fibres have a much smaller diameter than skeletal muscle cells. T-tubules are not required to reach the interior of the cell and therefore not necessary to transmit an action potential deep into the fibre. Smooth muscle fibres have a limited calcium-storing SR but have calcium channels in the sarcolemma (similar to cardiac muscle fibres) that open during the action potential along the sarcolemma. The influx of extracellular Ca2+ ions, which diffuse into the sarcoplasm to reach the calmodulin, accounts for most of the Ca2+ that triggers contraction of a smooth muscle cell.
Muscle contraction continues until ATP-dependent calcium pumps actively transport Ca2+ ions back into the SR and out of the cell. However, a low concentration of calcium remains in the sarcoplasm to maintain muscle tone. This remaining calcium keeps the muscle slightly contracted, which is important in certain tracts and around blood vessels.
Because most smooth muscles must function for long periods without rest, their power output is relatively low, but contractions can continue without using large amounts of energy. Some smooth muscle can also maintain contractions even as Ca2+ is removed, and myosin kinase is inactivated/dephosphorylated. This can happen as a subset of cross-bridges between myosin heads and actin, called latch-bridges, keep the thick and thin filaments linked together for a prolonged period, and without the need for ATP. This allows for the maintaining of muscle “tone” in smooth muscle that lines arterioles and other visceral organs with very little energy expenditure.
Smooth muscle is not under voluntary control; thus, it is called involuntary muscle. The triggers for smooth muscle contraction include hormones, neural stimulation by the ANS, and local factors. In certain locations, such as the walls of visceral organs, stretching the muscle can trigger its contraction (the stretch-relaxation response).
Axons of neurons in the ANS do not form the highly organised NMJs with smooth muscle, as seen between motor neurons and skeletal muscle fibres. Instead, there is a series of neurotransmitter-filled bulges called varicosities as an axon courses through smooth muscle, loosely forming motor units (Figure 9.8.3). A varicosity releases neurotransmitters into the synaptic cleft. Also, visceral muscle in the walls of the hollow organs (except the heart) contains pacesetter cells. A pacesetter cell can spontaneously trigger action potentials and contractions in the muscle.
Figure 9.8.3. Motor units. A series of axon-like swelling, called varicosities or “boutons,” from autonomic neurons form motor units through the smooth muscle.
Smooth muscle is organised in two ways: as single-unit smooth muscle, which is much more common; and as multiunit smooth muscle. The two types have different locations in the body and have different characteristics. Single-unit muscle has its muscle fibres joined by gap junctions so that the muscle contracts as a single unit. This type of smooth muscle is found in the walls of all visceral organs except the heart (which has cardiac muscle in its walls), and so it is commonly called visceral muscle. Because the muscle fibres are not constrained by the organisation and stretchability limits of sarcomeres, visceral smooth muscle has a stress-relaxation response. This means that as the muscle of a hollow organ is stretched when it fills, the mechanical stress of the stretching will trigger contraction, but this is immediately followed by relaxation so that the organ does not empty its contents prematurely. This is important for hollow organs, such as the stomach or urinary bladder, which continuously expand as they fill. The smooth muscle around these organs also can maintain a muscle tone when the organ empties and shrinks, a feature that prevents “flabbiness” in the empty organ. In general, visceral smooth muscle produces slow, steady contractions that allow substances, such as food in the digestive tract, to move through the body.
Multiunit smooth muscle cells rarely possess gap junctions, and thus are not electrically coupled. As a result, contraction does not spread from one cell to the next but is instead confined to the cell that was originally stimulated. Stimuli for multiunit smooth muscles come from autonomic nerves or hormones but not from stretching. This type of tissue is found around large blood vessels, in the respiratory airways, and in the eyes.
Hyperplasia in Smooth Muscle
Similar to skeletal and cardiac muscle cells, smooth muscle can undergo hypertrophy to increase in size. Unlike other muscle, smooth muscle can also divide to produce more cells, a process called hyperplasia. This can most evidently be observed in the uterus at puberty, which responds to increased estrogen levels by producing more uterine smooth muscle fibres, and greatly increases the size of the myometrium.
Section Review
Smooth muscle is found throughout the body around various organs and tracts. Smooth muscle cells have a single nucleus and are spindle-shaped. Smooth muscle cells can undergo hyperplasia, mitotically dividing to produce new cells. The smooth cells are nonstriated, but their sarcoplasm is filled with actin and myosin, along with dense bodies in the sarcolemma to anchor the thin filaments and a network of intermediate filaments involved in pulling the sarcolemma toward the fibre’s middle, shortening it in the process. Ca2+ ions trigger contraction when they are released from SR and enter through opened voltage-gated calcium channels. Smooth muscle contraction is initiated when the Ca2+ binds to intracellular calmodulin, which then activates an enzyme called myosin kinase that phosphorylates myosin heads so they can form the cross-bridges with actin and then pull on the thin filaments. Smooth muscle can be stimulated by pacesetter cells, by the autonomic nervous system, by hormones, spontaneously, or by stretching. The fibres in some smooth muscle have latch-bridges, cross-bridges that cycle slowly without the need for ATP; these muscles can maintain low-level contractions for long periods. Single-unit smooth muscle tissue contains gap junctions to synchronise membrane depolarisation and contractions so that the muscle contracts as a single unit. Single-unit smooth muscle in the walls of the viscera, called visceral muscle, has a stress-relaxation response that permits muscle to stretch, contract, and relax as the organ expands. Multiunit smooth muscle cells do not possess gap junctions, and contraction does not spread from one cell to the next.
Explain which muscle has the greatest regeneration ability
Most muscle tissue of the body arises from embryonic mesoderm. Paraxial mesodermal cells adjacent to the neural tube form blocks of cells called somites. Skeletal muscles, excluding those of the head and limbs, develop from mesodermal somites, whereas skeletal muscle in the head and limbs develop from general mesoderm. Somites give rise to myoblasts. A myoblast is a muscle-forming stem cell that migrates to different regions in the body and then fuse(s) to form a syncytium, or myotube. As a myotube is formed from many different myoblast cells, it contains many nuclei, but has a continuous cytoplasm. This is why skeletal muscle cells are multinucleate, as the nucleus of each contributing myoblast remains intact in the mature skeletal muscle cell. However, cardiac and smooth muscle cells are not multinucleate because the myoblasts that form their cells do not fuse.
Gap junctions develop in the cardiac and single-unit smooth muscle in the early stages of development. In skeletal muscles, ACh receptors are initially present along most of the surface of the myoblasts, but spinal nerve innervation causes the release of growth factors that stimulate the formation of motor end-plates and NMJs. As neurons become active, electrical signals that are sent through the muscle influence the distribution of slow and fast fibres in the muscle.
Although the number of muscle cells is set during development, satellite cells help to repair skeletal muscle cells. A satellite cell is similar to a myoblast because it is a type of stem cell; however, satellite cells are incorporated into muscle cells and facilitate the protein synthesis required for repair and growth. These cells are located outside the sarcolemma and are stimulated to grow and fuse with muscle cells by growth factors that are released by muscle fibres under certain forms of stress. Satellite cells can regenerate muscle fibres to a very limited extent, but they primarily help to repair damage in living cells. If a cell is damaged to a greater extent than can be repaired by satellite cells, the muscle fibres are replaced by scar tissue in a process called fibrosis. Because scar tissue cannot contract, muscle that has sustained significant damage loses strength and cannot produce the same amount of power or endurance as it could before being damaged.
Smooth muscle tissue can regenerate from a type of stem cell called a pericyte, which is found in some small blood vessels. Pericytes allow smooth muscle cells to regenerate and repair much more readily than skeletal and cardiac muscle tissue. Like skeletal muscle tissue, cardiac muscle does not regenerate to a great extent. Dead cardiac muscle tissue is replaced by scar tissue, which cannot contract. As scar tissue accumulates, the heart loses its ability to pump because of the loss of contractile power. However, some minor regeneration may occur due to stem cells found in the blood that occasionally enter cardiac tissue.
Career Connections
Physical Therapist
As muscle cells die, they are not regenerated but instead are replaced by connective tissue and adipose tissue, which do not possess the contractile abilities of muscle tissue. Muscles atrophy when they are not used, and over time if atrophy is prolonged, muscle cells die. It is therefore important that those who are susceptible to muscle atrophy exercise to maintain muscle function and prevent the complete loss of muscle tissue. In extreme cases, when movement is not possible, electrical stimulation can be introduced to a muscle from an external source. This acts as a substitute for endogenous neural stimulation, stimulating the muscle to contract and preventing the loss of proteins that occurs with a lack of use.
Physiotherapists work with patients to maintain muscles. They are trained to target muscles susceptible to atrophy, and to prescribe and monitor exercises designed to stimulate those muscles. There are various causes of atrophy, including mechanical injury, disease, and age. After breaking a limb or undergoing surgery, muscle use is impaired and can lead to disuse atrophy. If the muscles are not exercised, this atrophy can lead to long-term muscle weakness. A stroke can also cause muscle impairment by interrupting neural stimulation to certain muscles. Without neural inputs, these muscles do not contract and thus begin to lose structural proteins. Exercising these muscles can help to restore muscle function and minimise functional impairments. Age-related muscle loss is also a target of physical therapy, as exercise can reduce the effects of age-related atrophy and improve muscle function.
The goal of a physiotherapist is to improve physical functioning and reduce functional impairments; this is achieved by understanding the cause of muscle impairment and assessing the capabilities of a patient, after which a program to enhance these capabilities is designed. Some factors that are assessed include strength, balance, and endurance, which are continually monitored as exercises are introduced to track improvements in muscle function. Physiotherapists can also instruct patients on the proper use of equipment, such as crutches, and assess whether someone has sufficient strength to use the equipment and when they can function without it.
Section Review
Muscle tissue arises from embryonic mesoderm. Somites give rise to myoblasts and fuse to form a myotube. The nucleus of each contributing myoblast remains intact in the mature skeletal muscle cell, resulting in a mature, multinucleate cell. Satellite cells help to repair skeletal muscle cells. Smooth muscle tissue can regenerate from stem cells called pericytes, whereas dead cardiac muscle tissue is replaced by scar tissue. Ageing causes muscle mass to decrease and be replaced by noncontractile connective tissue and adipose tissue.
List and describe the functions of the skeletal system
Bone, or osseous tissue, is a hard, dense connective tissue that forms most of the adult skeleton, the support structure of the body. In the areas of the skeleton where bones move (for example, the ribcage and joints), cartilage, a semi-rigid form of connective tissue, provides flexibility and smooth surfaces for movement. The skeletal system is the body system composed of bones and cartilage and performs the following critical functions for the human body:
supports the body
facilitates movement
protects internal organs
produces blood cells
stores and releases minerals and fat
Support, Movement and Protection
Figure 10.1.1. Bones support movement. Bones act as levers when muscles span a joint and contract. (credit: Benjamin J. DeLong).
The most apparent functions of the skeletal system are the gross functions—those visible by observation. Simply by looking at a person, you can see how the bones support, facilitate movement, and protect the human body.
Just as the steel beams of a building provide a scaffold to support its weight, the bones and cartilage of your skeletal system compose the scaffold that supports the rest of your body. Without the skeletal system, you would be a limp mass of organs, muscle, and skin.
Bones also facilitate movement by serving as points of attachment for your muscles. While some bones only serve as a support for the muscles, others also transmit the forces produced when your muscles contract. From a mechanical point of view, bones act as levers and joints serve as fulcrums (Figure 10.1.1). Unless a muscle spans a joint and contracts, a bone is not going to move.
Bones also protect internal organs from injury by covering or surrounding them. Your ribs protect your lungs and heart, the bones of your vertebral column (spine) protect your spinal cord, and the bones of your cranium (skull) protect your brain (Figure 10.1.2).
Figure 10.1.2. Bones protect brain. The cranium surrounds and protects the brain from non-traumatic injury.
Career Connection
Orthopaedist
An orthopaedist is a doctor who specialises in diagnosing and treating disorders and injuries related to the musculoskeletal system. Some orthopaedic problems can be treated with medications, exercises, braces, and other devices, but others may be best treated with surgery (Figure 10.1.3).
While the origin of the word “orthopaedics” (ortho- = “straight”; paed- = “child”), literally means “straightening of the child,” orthopaedists can have patients who range from paediatric to geriatric. In recent years, orthopaedists have even performed prenatal surgery to correct spina bifida, a congenital defect in which the neural canal in the spine of the foetus fails to close completely during embryologic development.
Figure 10.1.3. Arm brace. An orthopaedist will sometimes prescribe the use of a brace that reinforces the underlying bone structure it is being used to support. (credit: Juhan Sonin).
Orthopaedists commonly treat bone and joint injuries but they also treat other bone conditions including curvature of the spine. Lateral curvatures (scoliosis) can be severe enough to slip under the shoulder blade (scapula) forcing it up as a hump. Spinal curvatures can also be excessive dorsoventrally (kyphosis) causing a hunch back and thoracic compression. These curvatures often appear in preteens as the result of poor posture, abnormal growth, or indeterminate causes. Mostly, they are readily treated by orthopaedists. As people age, accumulated spinal column injuries and diseases like osteoporosis can also lead to curvatures of the spine, hence the stooping you sometimes see in the elderly.
Some orthopaedists sub-specialise in sports medicine, which addresses both simple injuries, such as a sprained ankle, and complex injuries, such as a torn rotator cuff in the shoulder. Treatment can range from exercise to surgery. The Australian Orthopaedic Association (AOA) is a not-for-profit organisation that provides specialist education and training, ensuring high standard of orthopaedic care, and is a leading authority body who actively supports scientific research and orthopaedic humanitarian initiatives in Australia and worldwide.
Mineral Storage, Energy Storage and Haematopoiesis
On a metabolic level, bone tissue performs several critical functions. For one, the bone matrix acts as a reservoir for several minerals important to the functioning of the body, especially calcium and phosphorus. These minerals, incorporated into bone tissue, can be released back into the bloodstream to maintain levels needed to support physiological processes. Calcium ions, for example, are essential for muscle contractions and controlling the flow of other ions involved in the transmission of nerve impulses.
Bone also serves as a site for fat storage and blood cell production. The softer connective tissue that fills the interior of most bone is referred to as bone marrow (Figure 10.1.4). There are two types of bone marrow: yellow marrow and red marrow. Yellow marrow contains adipose tissue; the triglycerides stored in the adipocytes of the tissue can serve as a source of energy. Red marrow is where haematopoiesis—the production of blood cells—takes place. Red blood cells, white blood cells, and platelets are all produced in the red marrow.
Figure 10.1.4. Head of femur showing red and yellow marrow. The head of the femur contains both yellow and red marrow. Yellow marrow stores fat. Red marrow is responsible for haematopoiesis. (credit: modification of work by “stevenfruitsmaak”/Wikimedia Commons).
Section Review
The major functions of the bones are body support, facilitation of movement, protection of internal organs, storage of minerals and fat and haematopoiesis. Together, the muscular system and skeletal system are known as the musculoskeletal system.
The 206 bones that compose the adult skeleton are divided into five categories based on their shapes (Figure 10.2.1). Their shapes and their functions are related such that each categorical shape of bone has a distinct function.
Figure 10.2.1. Classifications of bones. Bones are classified according to their shape.
Long Bones
A long bone is one that is cylindrical in shape, being longer than it is wide. Keep in mind, however, that the term describes the shape of a bone, not its size. Long bones are found in the arms (humerus, ulna, radius) and legs (femur, tibia, fibula), as well as in the fingers (metacarpals, phalanges) and toes (metatarsals, phalanges). Long bones function as levers; they move when muscles contract.
Short Bones
A short bone is one that is cube-like in shape, being approximately equal in length, width, and thickness. The only short bones in the human skeleton are in the carpals of the wrists and the tarsals of the ankles. Short bones provide stability and support as well as some limited motion.
Flat Bones
The term “flat bone” is somewhat of a misnomer because, although a flat bone is typically thin, it is also often curved. Examples include the cranial (skull) bones, the scapulae (shoulder blades), the sternum (breastbone), and the ribs. Flat bones serve as points of attachment for muscles and often protect internal organs.
Irregular Bones
An irregular bone is one that does not have any easily characterised shape and therefore does not fit any other classification. These bones tend to have more complex shapes, like the vertebrae that support the spinal cord and protect it from compressive forces. Many facial bones, particularly the ones containing sinuses, are classified as irregular bones.
Sesamoid Bones
A sesamoid bone is a small, round bone that, as the name suggests, is shaped like a sesame seed. These bones form in tendons (the sheaths of tissue that connect bones to muscles) where a great deal of pressure is generated in a joint. The sesamoid bones protect tendons by helping them overcome compressive forces. Sesamoid bones vary in number and placement from person to person but are typically found in tendons associated with the feet, hands, and knees. The patellae (singular = patella) are the only sesamoid bones found in common with every person. Table 10.2.1 reviews bone classifications with their associated features, functions, and examples.
Cube-like shape, approximately equal in length, width, and thickness
Provides stability, support, while allowing for some motion
Carpals, tarsals
Flat
Thin and curved
Points of attachment for muscles; protectors of internal organs
Sternum, ribs, scapulae, cranial bones
Irregular
Complex shape
Protect internal organs
Vertebrae, facial bones
Sesamoid
Small and round; embedded in tendons
Protect tendons from compressive forces
Patellae
Section Review
Bones can be classified according to their shapes. Long bones, such as the femur, are longer than they are wide. Short bones, such as the carpals, are approximately equal in length, width, and thickness. Flat bones are thin, but are often curved, such as the ribs. Irregular bones such as those of the face have no characteristic shape. Sesamoid bones, such as the patellae, are small and round and are in tendons.
Identify the structures that compose compact and spongy bone
Describe how bones are nourished and innervated
Bone tissue (osseous tissue) differs from other tissues in the body. Bone is hard and many of its functions depend on that characteristic hardness. Later discussions in this chapter will show that bone is also dynamic in that its shape adjusts to accommodate stresses. This section will examine the gross anatomy of bone first and then move on to its histology.
Gross Anatomy of Bone
The structure of a long bone allows for the best visualisation of all the parts of a bone (Figure 10.3.1). A long bone has two parts: the diaphysis and the epiphysis. The diaphysis is the tubular shaft that runs between the proximal and distal ends of the bone. The hollow region in the diaphysis is called the medullary cavity, which is filled with yellow marrow. The walls of the diaphysis are composed of dense and hard compact bone.
Figure 10.3.1. Anatomy of a long bone. A typical long bone shows the gross anatomical characteristics of bone.
The wider section at each end of the bone is called the epiphysis (plural = epiphyses), which is filled with spongy bone. Red marrow fills the spaces in the spongy bone. Each epiphysis meets the diaphysis at the metaphysis, the narrow area that contains the epiphyseal plate (growth plate), a layer of hyaline (transparent) cartilage in a growing bone. When the bone stops growing in early adulthood (approximately 18–21 years), the cartilage is replaced by osseous tissue and the epiphyseal plate becomes an epiphyseal line.
The medullary cavity has a delicate membranous lining called the endosteum (end- = “inside”; oste- = “bone”), where bone growth, repair, and remodelling occur. The outer surface of the bone is covered with a fibrous membrane called the periosteum (peri– = “around” or “surrounding”). The periosteum contains blood vessels, nerves, and lymphatic vessels that nourish compact bone. Tendons and ligaments also attach to bones at the periosteum. The periosteum covers the entire outer surface except where the epiphyses meet other bones to form joints (Figure 10.3.2). In this region, the epiphyses are covered with articular cartilage, a thin layer of cartilage that reduces friction and acts as a shock absorber.
Figure 10.3.2. Periosteum and endosteum. The periosteum forms the outer surface of bone, and the endosteum lines the medullary cavity.
Flat bones, like those of the cranium, consist of a layer of diploë (spongy bone), lined on either side by a layer of compact bone (Figure 10.3.3). The two layers of compact bone and the interior spongy bone work together to protect the internal organs. If the outer layer of a cranial bone fractures, the brain is still protected by the intact inner layer.
Figure 10.3.3. Anatomy of a flat bone. This cross-section of a flat bone shows the spongy bone (diploë) lined on either side by a layer of compact bone.
Bone Markings
The surface features of bones vary depending on the function and location in the body. Table 10.3.1 describes the bone markings, which are illustrated in (Figure 10.3.4). There are three general classes of bone markings: (1) articulations, (2) projections and (3) holes. As the name implies, an articulation is where two bone surfaces come together (articulus = “joint”). These surfaces tend to conform to one another, such as one being rounded and the other cupped, to facilitate the function of the articulation. A projection is an area of a bone that projects above the surface of the bone. These are the attachment points for tendons and ligaments. In general, their size and shape are an indication of the forces exerted through the attachment to the bone. A hole is an opening or groove in the bone that allows blood vessels and nerves to enter the bone. As with the other markings, their size and shape reflect the size of the vessels and nerves that penetrate the bone at these points.
Table 10.3.1 Bone Markings
Marking
Description
Example
Articulations
Where two bones meet
Knee joint
Canal
Passage in bone
Auditory canal
Condyle
Rounded surface
Occipital condyles
Crest
Ridge
Iliac crest
Facet
Flat surface
Vertebrae
Fissure
Slit through bone
Auricular fissure
Foramen
Hole through bone
Foramen magnum in the occipital bone
Fossa
Elongated basin
Mandibular fossa
Fovea
Small pit
Fovea capitis on the head of the femur
Head
Prominent rounded surface
Head of femur
Holes
Holes and depressions
Foramen (holes through which blood vessels can pass through)
Line
Slight, elongated ridge
Temporal lines of the parietal bones
Meatus
Opening into canal
External auditory meatus
Process
Prominence feature
Transverse process of vertebra
Projections
Raised Markings
Spinous process of the vertebrae
Protuberance
Protruding
Chin
Sinus
Air-filled space in bone
Nasal sinus
Spine
Sharp process
Ischial spine
Sulcus
Groove
Sigmoid sulcus of the temporal bones
Tubercle
Small, rounded process
Deltoid tuberosity
Figure 10.3.4. Bone features. The surface features of bones depend on their function, location, attachment of ligaments and tendons, or the penetration of blood vessels and nerves.
Bone Cells and Tissue
Bone contains a relatively small number of cells entrenched in a matrix of collagen fibres that provide a surface for inorganic salt crystals to adhere. These salt crystals form when calcium phosphate and calcium carbonate combine to create hydroxyapatite, which incorporates other inorganic salts like magnesium hydroxide, fluoride and sulphate as it crystallises, or calcifies, on the collagen fibres. The hydroxyapatite crystals give bones their hardness and strength, while the collagen fibres give them flexibility so that they are not brittle.
Although bone cells compose a small amount of the bone volume, they are crucial to the function of bones. Four types of cells are found within bone tissue: osteoblasts, osteocytes, osteogenic cells and osteoclasts (Figure 10.3.5).
Figure 10.3.5. Bone cells. Four types of cells are found within bone tissue. Osteogenic cells are undifferentiated and develop into osteoblasts. When osteoblasts get trapped within the calcified matrix, their structure and function changes, and they become osteocytes. Osteoclasts develop from monocytes and macrophages and differ in appearance from other bone cells.
The osteoblast is the bone cell responsible for forming new bone and is found in the growing portions of bone, including the periosteum and endosteum. Osteoblasts, which do not divide, synthesise and secrete the collagen matrix and calcium salts. As the secreted matrix surrounding the osteoblast calcifies, the osteoblast become trapped within it; as a result, it changes in structure and becomes an osteocyte, the primary cell of mature bone and the most common type of bone cell. Each osteocyte is located in a space called a lacuna and is surrounded by bone tissue. Osteocytes maintain the mineral concentration of the matrix via the secretion of enzymes. Like osteoblasts, osteocytes lack mitotic activity. They can communicate with each other and receive nutrients via long cytoplasmic processes that extend through canaliculi (singular = canaliculus), channels within the bone matrix.
If osteoblasts and osteocytes are incapable of mitosis, then how are they replenished when old ones die? The answer lies in the properties of a third category of bone cells—the osteogenic cell. These osteogenic cells are undifferentiated with high mitotic activity and they are the only bone cells that divide. Immature osteogenic cells are found in the deep layers of the periosteum and the marrow. They differentiate and develop into osteoblasts.
The dynamic nature of bone means that new tissue is constantly formed, and old, injured, or unnecessary bone is dissolved for repair or for calcium release. The cell responsible for bone resorption, or breakdown, is the osteoclast. They are found on bone surfaces, are multinucleated, and originate from monocytes and macrophages, two types of white blood cells, not from osteogenic cells. Osteoclasts are continually breaking down old bone while osteoblasts are continually forming new bone. The ongoing balance between osteoblasts and osteoclasts is responsible for the constant but subtle reshaping of bone. Table 10.3.2 reviews the bone cells, their functions and locations.
Table 10.3.2. Bone Cells
Cell type
Function
Location
Osteogenic cells
Develop into osteoblasts
Deep layers of the periosteum and the marrow
Osteoblasts
Bone formation
Growing portions of bone, including periosteum and endosteum
Osteocytes
Maintain mineral concentration of matrix
Entrapped in matrix
Osteoclasts
Bone resorption
Bone surfaces and at sites of old, injured or unneeded bone
Compact and Spongy Bone
The differences between compact and spongy bone are best explored via their histology. Most bones contain compact and spongy osseous tissue, but their distribution and concentration vary based on the bone’s overall function. Compact bone is dense so that it can withstand compressive forces, while spongy (cancellous) bone has open spaces and supports shifts in weight distribution.
Compact Bone
Compact bone is the denser, stronger of the two types of bone tissue (Figure 10.3.6). It can be found under the periosteum and in the diaphyses of long bones, where it provides support and protection.
The microscopic structural unit of compact bone is called an osteon, or Haversian system. Each osteon is composed of concentric rings of calcified matrix called lamellae (singular = lamella). Running down the centre of each osteon is the central canal, or Haversian canal, which contains blood vessels, nerves, and lymphatic vessels. These vessels and nerves branch off at right angles through a perforating canal, also known as Volkmann’s canals, to extend to the periosteum and endosteum.
The osteocytes are located inside spaces called lacunae (singular = lacuna), found at the borders of adjacent lamellae. As described earlier, canaliculi connect with the canaliculi of other lacunae and eventually with the central canal. This system allows nutrients to be transported to the osteocytes and wastes to be removed from them.
Spongy (Cancellous) Bone
Like compact bone, spongy bone, also known as cancellous bone, contains osteocytes housed in lacunae, but they are not arranged in concentric circles. Instead, the lacunae and osteocytes are found in a lattice-like network of matrix spikes called trabeculae (singular = trabecula) (Figure 10.3.7). The trabeculae may appear to be a random network, but each trabecula forms along lines of stress to provide strength to the bone. The spaces of the trabeculated network provide balance to the dense and heavy compact bone by making bones lighter so that muscles can move them more easily. In addition, the spaces in some spongy bones contain red marrow, protected by the trabeculae, where haematopoiesis occurs.
Figure 10.3.7. Diagram of spongy bone. Spongy bone is composed of trabeculae that contain the osteocytes. Red marrow fills the spaces in some bones.
Ageing and the Skeletal System: Paget’s Disease
Paget’s disease usually occurs in adults over age 40. It is a disorder of the bone remodelling process that begins with overactive osteoclasts. This means more bone is resorbed than is laid down. The osteoblasts try to compensate but the new bone they lay down is weak and brittle and therefore prone to fracture.
While some people with Paget’s disease have no symptoms, others experience pain, bone fractures, and bone deformities (Figure 10.3.8). Bones of the pelvis, skull, spine, and legs are the most affected. When occurring in the skull, Paget’s disease can cause headaches and hearing loss.
Figure 10.3.8. Paget’s disease. Normal leg bones are relatively straight, but those affected by Paget’s disease are porous and curved.
What causes the osteoclasts to become overactive? The answer is still unknown, but hereditary factors play a role. Some scientists believe Paget’s disease is due to an as-yet-unidentified virus.
Paget’s disease is diagnosed via imaging studies and lab tests. X-rays may show bone deformities or areas of bone resorption. Bone scans are also useful. In these studies, a dye containing a radioactive ion is injected into the body. Areas of bone resorption have an affinity for the ion, so they will light up on the scan if the ions are absorbed. In addition, blood levels of an enzyme called alkaline phosphatase are typically elevated in people with Paget’s disease.
Bisphosphonates, drugs that decrease the activity of osteoclasts, are often used in the treatment of Paget’s disease. However, in a small percentage of cases, bisphosphonates themselves have been linked to an increased risk of fractures because the old bone that is left after bisphosphonates are administered becomes worn out and brittle. Still, most doctors feel that the benefits of bisphosphonates more than outweigh the risk; the medical professional must weigh the benefits and risks on a case-by-case basis. Bisphosphonate treatment can reduce the overall risk of deformities or fractures, which in turn reduces the risk of surgical repair and its associated risks and complications.
Blood and Nerve Supply
The spongy bone and medullary cavity receive nourishment from arteries that pass through the compact bone. The arteries enter through the nutrient foramen (plural = foramina), small openings in the diaphysis. The osteocytes in spongy bone are nourished by blood vessels of the periosteum that penetrate spongy bone and blood that circulates in the marrow cavities. As the blood passes through the marrow cavities, it is collected by veins, which then pass out of the bone through the foramina.
In addition to the blood vessels, nerves follow the same paths into the bone where they tend to concentrate in the more metabolically active regions of the bone. The nerves sense pain, and the nerves also play roles in regulating blood supplies and in bone growth, hence their concentrations in metabolically active sites of the bone.
Figure 10.3.9. Diagram of blood and nerve supply to bone. Blood vessels and nerves enter the bone through the nutrient foramen.
Section Review
A hollow medullary cavity filled with yellow marrow runs the length of the diaphysis of a long bone. The walls of the diaphysis are compact bone. The epiphyses, which are wider sections at each end of a long bone, are filled with spongy bone and red marrow. The epiphyseal plate, a layer of hyaline cartilage, is replaced by osseous tissue as the organ grows in length. The medullary cavity has a delicate membranous lining called the endosteum. The outer surface of bone, except in regions covered with articular cartilage, is covered with a fibrous membrane called the periosteum. Flat bones consist of two layers of compact bone surrounding a layer of spongy bone. Bone markings depend on the function and location of bones. Articulations are places where two bones meet. Projections stick out from the surface of the bone and provide attachment points for tendons and ligaments. Holes are openings or depressions in the bones.
Bone matrix consists of collagen fibres and organic ground substance, primarily hydroxyapatite formed from calcium salts. Osteogenic cells develop into osteoblasts. Osteoblasts are cells that make new bone. They become osteocytes, the cells of mature bone, when they get trapped in the matrix. Osteoclasts engage in bone resorption. Compact bone is dense and composed of osteons, while spongy bone is less dense and made up of trabeculae. Blood vessels and nerves enter the bone through the nutrient foramina to nourish and innervate bones.
Explain the growth activity at the epiphyseal plate
Compare and contrast the processes of modelling and remodelling
In the early stages of embryonic development, the embryo’s skeleton consists of fibrous membranes and hyaline cartilage. By the sixth or seventh week of embryonic life, the actual process of bone development, ossification (osteogenesis), begins. There are two osteogenic pathways—intramembranous ossification and endochondral ossification—but bone is the same regardless of the pathway that produces it.
Cartilage Templates
Bone is a replacement tissue; that is, it uses a model tissue on which to lay down its mineral matrix. For skeletal development, the most common template is cartilage. During foetal development, a framework is laid down that determines where bones will form. This framework is a flexible, semi-solid matrix produced by chondroblasts and consists of hyaluronic acid, chondroitin sulphate, collagen fibres, and water. As the matrix surrounds and isolates chondroblasts, they are called chondrocytes. Unlike most connective tissues, cartilage is avascular, meaning that it has no blood vessels supplying nutrients and removing metabolic wastes. All these functions are carried on by diffusion through the matrix. This is why damaged cartilage does not repair itself as readily as most tissues do.
Throughout foetal development and into childhood growth and development, bone forms on the cartilaginous matrix. By the time a foetus is born, most of the cartilage has been replaced with bone. Some additional cartilage will be replaced throughout childhood, and some cartilage remains in the adult skeleton.
Intermembranous Ossification
During intramembranous ossification, compact and spongy bone develops directly from sheets of mesenchymal (undifferentiated) connective tissue. The flat bones of the face, most of the cranial bones, and the clavicles (collarbones) are formed via intramembranous ossification.
The process begins when mesenchymal cells in the embryonic skeleton gather together and begin to differentiate into specialised cells (Figure 10.4.1a). Some of these cells will differentiate into capillaries, while others will become osteogenic cells and then osteoblasts. Although they will be spread out by the formation of bone tissue, early osteoblasts appear in a cluster called an ossification centre.
The osteoblasts secrete osteoid, uncalcified matrix, which calcifies (hardens) within a few days as mineral salts are deposited on it, thereby entrapping the osteoblasts within. Once entrapped, the osteoblasts become osteocytes (Figure 10.4.1b). As osteoblasts transform into osteocytes, osteogenic cells in the surrounding connective tissue differentiate into new osteoblasts.
Osteoid (unmineralised bone matrix) secreted around the capillaries results in a trabecular matrix, while osteoblasts on the surface of the spongy bone become the periosteum (Figure 10.4.1c). The periosteum then creates a protective layer of compact bone superficial to the trabecular bone. The trabecular bone crowds nearby blood vessels, which eventually condense into red marrow (Figure 10.4.1d).
Figure 10.4.1. Intramembranous ossification. Intramembranous ossification follows four steps. (a) Mesenchymal cells group into clusters, and ossification centres form. (b) Secreted osteoid traps osteoblasts, which then become osteocytes. (c) Trabecular matrix and periosteum form. (d) Compact bone develops superficial to the trabecular bone, and crowded blood vessels condense into red marrow.
Intramembranous ossification begins in utero during foetal development and continues into adolescence. At birth, the skull and clavicles are not fully ossified nor are the sutures of the skull closed. This allows the skull and shoulders to deform during passage through the birth canal. The last bones to ossify via intramembranous ossification are the flat bones of the face, which reach their adult size at the end of the adolescent growth spurt.
Endochondral Ossification
In endochondral ossification, bone develops by replacing hyaline cartilage. Cartilage does not become bone. Instead, cartilage serves as a template to be completely replaced by new bone. Endochondral ossification takes much longer than intramembranous ossification. Bones at the base of the skull and long bones form via endochondral ossification.
In a long bone, for example, at about 6 to 8 weeks after conception, some of the mesenchymal cells differentiate into chondrocytes (cartilage cells) that form the cartilaginous skeletal precursor of the bones (Figure 10.4.2a). Soon after, the perichondrium, a membrane that covers the cartilage, appears (Figure 10.4.2b).
Figure 10.4.2. Endochondral ossification. Endochondral ossification follows five steps. (a) Mesenchymal cells differentiate into chondrocytes. (b) The cartilage model of the future bony skeleton and the perichondrium form. (c) Capillaries penetrate cartilage. Perichondrium transforms into periosteum. Periosteal collar develops. Primary ossification centre develops. (d) Cartilage and chondrocytes continue to grow at ends of the bone. (e) Secondary ossification centres develop. (f) Cartilage remains at epiphyseal (growth) plate and at joint surface as articular cartilage.
As more matrix is produced, the chondrocytes in the centre of the cartilaginous model grow in size. As the matrix calcifies, nutrients can no longer reach the chondrocytes. This results in their death and the disintegration of the surrounding cartilage. Blood vessels invade the resulting spaces, not only enlarging the cavities but also carrying osteogenic cells with them, many of which will become osteoblasts. These enlarging spaces eventually combine to become the medullary cavity.
As the cartilage grows, capillaries penetrate it. This penetration initiates the transformation of the perichondrium into the bone-producing periosteum. Here, the osteoblasts form a periosteal collar of compact bone around the cartilage of the diaphysis. By the second or third month of foetal life, bone cell development and ossification ramps up and creates the primary ossification centre, a region deep in the periosteal collar where ossification begins (Figure 10.4.2c).
While these deep changes are occurring, chondrocytes and cartilage continue to grow at the ends of the bone (the future epiphyses), which increases the bone’s length at the same time bone is replacing cartilage in the diaphyses. By the time the foetal skeleton is fully formed, cartilage only remains at the joint surface as articular cartilage and between the diaphysis and epiphysis as the epiphyseal plate, the latter of which is responsible for the longitudinal growth of bones. After birth, this same sequence of events (matrix mineralisation, death of chondrocytes, invasion of blood vessels from the periosteum, and seeding with osteogenic cells that become osteoblasts) occurs in the epiphyseal regions, and each of these centres of activity is referred to as a secondary ossification centre (Figure 10.4.2e).
How Bones Grow in Length
The epiphyseal plate is the area of growth in a long bone. It is a layer of hyaline cartilage where ossification occurs in immature bones. On the epiphyseal side of the epiphyseal plate, cartilage is formed. On the diaphyseal side, cartilage is ossified, and the diaphysis grows in length. The epiphyseal plate is composed of four zones of cells and activity (Figure 10.4.3). The reserve zone is the region closest to the epiphyseal end of the plate and contains small chondrocytes within the matrix. These chondrocytes do not participate in bone growth but secure the epiphyseal plate to the osseous tissue of the epiphysis.
Figure 10.4.3. Longitudinal bone growth. The epiphyseal plate is responsible for longitudinal bone growth.
The proliferative zone is the next layer toward the diaphysis and contains stacks of slightly larger chondrocytes. It makes new chondrocytes (via mitosis) to replace those that die at the diaphyseal end of the plate. Chondrocytes in the next layer, the zone of maturation and hypertrophy, are older and larger than those in the proliferative zone. The more mature cells are situated closer to the diaphyseal end of the plate. The longitudinal growth of bone is a result of cellular division in the proliferative zone and the maturation of cells in the zone of maturation and hypertrophy.
Most of the chondrocytes in the zone of calcified matrix, the zone closest to the diaphysis, are dead because the matrix around them has calcified. Capillaries and osteoblasts from the diaphysis penetrate this zone, and the osteoblasts secrete bone tissue on the remaining calcified cartilage. Thus, the zone of calcified matrix connects the epiphyseal plate to the diaphysis. A bone grows in length when osseous tissue is added to the diaphysis.
Bones continue to grow in length until early adulthood. The rate of growth is controlled by hormones, which will be discussed later. When the chondrocytes in the epiphyseal plate cease their proliferation and bone replaces the cartilage, longitudinal growth stops. All that remains of the epiphyseal plate is the epiphyseal line (Figure 10.4.4).
Figure 10.4.4. Progression from epiphyseal plate to epiphyseal line. As a bone matures, the epiphyseal plate progresses to an epiphyseal line. (a) Epiphyseal plates are visible in a growing bone. (b) Epiphyseal lines are the remnants of epiphyseal plates in a mature bone.
How Bones Growth in Diameter
While bones are increasing in length, they are also increasing in diameter; growth in diameter can continue even after longitudinal growth ceases. This is called appositional growth. Osteoclasts resorb old bone that lines the medullary cavity, while osteoblasts, via intramembranous ossification, produce new bone tissue beneath the periosteum. The erosion of old bone along the medullary cavity and the deposition of new bone beneath the periosteum not only increase the diameter of the diaphysis but also increase the diameter of the medullary cavity. This process is called modelling.
Bone Remodelling
The process in which matrix is resorbed on one surface of a bone and deposited on another is known as bone modelling. Modelling primarily takes place during a bone’s growth. However, in adult life, bone undergoes remodelling, in which resorption of old or damaged bone takes place on the same surface where osteoblasts lay new bone to replace that which is resorbed. Injury, exercise, and other activities lead to remodelling. Those influences are discussed later in the chapter, but even without injury or exercise, about 5 to 10 percent of the skeleton is remodelled annually just by destroying old bone and renewing it with fresh bone.
Diseases of the Skeletal System
Osteogenesis imperfecta (OI) is a genetic disease in which bones do not form properly and therefore are fragile and break easily. It is also called brittle bone disease. The disease is present from birth and affects a person throughout life.
The genetic mutation that causes OI affects the body’s production of collagen, one of the critical components of bone matrix. The severity of the disease can range from mild to severe. Those with the most severe forms of the disease sustain many more fractures than those with a mild form. Frequent and multiple fractures typically lead to bone deformities and short stature. Bowing of the long bones and curvature of the spine are also common in people afflicted with OI. Curvature of the spine makes breathing difficult because the lungs are compressed.
Because collagen is such an important structural protein in many parts of the body, people with OI may also experience fragile skin, weak muscles, loose joints, easy bruising, frequent nosebleeds, brittle teeth, blue sclera, and hearing loss. There is no known cure for OI. Treatment focuses on helping the person retain as much independence as possible while minimising fractures and maximising mobility. Toward that end, safe exercises, like swimming, in which the body is less likely to experience collisions or compressive forces, are recommended. Braces to support legs, ankles, knees, and wrists are used as needed. Canes, walkers, or wheelchairs can also help compensate for weaknesses.
When bones do break, casts, splints, or wraps are used. In some cases, metal rods may be surgically implanted into the long bones of the arms and legs. Research is currently being conducted on using bisphosphonates to treat OI. Smoking and being overweight are especially risky in people with OI, since smoking is known to weaken bones, and extra body weight puts additional stress on the bones.
Section Review
All bone formation is a replacement process. Embryos develop a cartilaginous skeleton and various membranes. During development, these are replaced by bone during the ossification process. In intramembranous ossification, bone develops directly from sheets of mesenchymal connective tissue. In endochondral ossification, bone develops by replacing hyaline cartilage. Activity in the epiphyseal plate enables bones to grow in length. Modelling allows bones to grow in diameter. Remodelling occurs as bone is resorbed and replaced by new bone. Osteogenesis imperfecta is a genetic disease in which collagen production is altered, resulting in fragile, brittle bones.
Differentiate among the different types of fractures
Describe the steps involved in bone repair
A fracture is a broken bone. It will heal whether or not a physician resets it in its anatomical position. If the bone is not reset correctly, the healing process will keep the bone in its deformed position.
When a broken bone is manipulated and set into its natural position without surgery, the procedure is called a closed reduction. Open reduction requires surgery to expose the fracture and reset the bone. While some fractures can be minor, others are quite severe and result in grave complications. For example, a fractured diaphysis of the femur has the potential to release fat globules into the bloodstream. These can become lodged in the capillary beds of the lungs, leading to respiratory distress and if not treated quickly, death.
Types of Fractures
Fractures are classified by their complexity, location, and other features (Figure 10.5.1). Table 10.5.1 outlines common types of fractures. Some fractures may be described using more than one term because it may have the features of more than one type (e.g., an open transverse fracture).
Figure 10.5.1. Types of fractures. Compare healthy bone with different types of fractures: (a) closed fracture, (b) open fracture, (c) transverse fracture, (d) spiral fracture, (e) comminuted fracture, (f) impacted fracture, (g) greenstick fracture, and (h) oblique fracture.
Table 10.5.1 Types of Fractures
Type of fracture
Description
Closed (or simple)
A fracture in which the skin remains intact
Comminated
Several breaks result in many small pieces between two large segments
Greenstick
A partial fracture in which only one side of the bone is broken
Impact
One fragment is driven into the other, usually as a result of compression
Oblique
Occurs at an angle that is not 90 degrees
Open (or compound)
A fracture in which at least one end of the bone tears through the skin; carries high risk of infection
Spiral
Bone segments are pulled apart as a result of a twisting motion
Transverse
Occurs straight across the long axis of the bone
Bone Repair
When a bone breaks, blood flows from any vessel torn by the fracture. These vessels could be in the periosteum, osteons, and/or medullary cavity. The blood begins to clot, and about six to eight hours after the fracture, the clotting blood has formed a fracture haematoma (Figure 10.5.2a). The disruption of blood flow to the bone results in the death of bone cells around the fracture.
Figure 10.5.2. Stages in Fracture Repair. The healing of a bone fracture follows a series of progressive steps: (a) A fracture haematoma forms. (b) Internal and external calli form. (c) Cartilage of the calli is replaced by trabecular bone. (d) Remodelling occurs.
Within about 48 hours after the fracture, chondrocytes from the endosteum have created an internal callus (plural = calli) by secreting a fibrocartilaginous matrix between the two ends of the broken bone, while the periosteal chondrocytes and osteoblasts create an external callus of hyaline cartilage and bone, respectively, around the outside of the break (Figure 10.5.2b). This stabilises the fracture.
Over the next several weeks, osteoclasts resorb the dead bone; osteogenic cells become active, divide, and differentiate into osteoblasts. The cartilage in the calli is replaced by trabecular bone via endochondral ossification (Figure 10.5.2c).
Eventually, the internal and external calli unite, compact bone replaces spongy bone at the outer margins of the fracture, and healing is complete. A slight swelling may remain on the outer surface of the bone, but quite often, that region undergoes remodelling (Figure 10.5.2d), and no external evidence of the fracture remains.
Section Review
Fractured bones may be repaired by closed reduction or open reduction. Fractures are classified by their complexity, location, and other features. Common types of fractures are transverse, oblique, spiral, comminuted, impacted, greenstick, open (or compound), and closed (or simple). Healing of fractures begins with the formation of a haematoma, followed by internal and external calli. Osteoclasts resorb dead bone, while osteoblasts create new bone that replaces the cartilage in the calli. The calli eventually unite, remodelling occurs, and healing is complete.
10.6 Exercise, Nutrition, Hormones and Bone Tissue
Learning Objectives
By the end of this section, you will be able to:
Describe the effect exercise has on bone tissue
List the nutrients that affect bone health
Discuss the role those nutrients play in bone health
Describe the effects of hormones on bone tissue
All the organ systems of your body are interdependent, and the skeletal system is no exception. The food you take in via your digestive system and the hormones secreted by your endocrine system affect your bones. Even using your muscles to engage in exercise has an impact on your bones.
Exercise and Bone Tissue
During long space missions, astronauts can lose approximately 1 to 2 percent of their bone mass per month. This loss of bone mass is thought to be caused by the lack of mechanical stress on astronauts’ bones due to the low gravitational forces in space. Lack of mechanical stress causes bones to lose mineral salts and collagen fibres, and thus strength. Similarly, mechanical stress stimulates the deposition of mineral salts and collagen fibres. The internal and external structure of a bone will change as stress increases or decreases so that the bone is an ideal size and weight for the amount of activity it endures. That is why people who exercise regularly have thicker bones than people who are more sedentary. It is also why a broken bone in a cast atrophies while its contralateral mate maintains its concentration of mineral salts and collagen fibres. The bones undergo remodelling because of forces (or lack of forces) placed on them.
Numerous, controlled studies have demonstrated that people who exercise regularly have greater bone density than those who are more sedentary. Any type of exercise will stimulate the deposition of more bone tissue, but resistance training has a greater effect than cardiovascular activities. Resistance training is especially important to slow down the eventual bone loss due to ageing and for preventing osteoporosis.
Nutrition and Bone Tissue
The vitamins and minerals contained in all the food we consume are important for all our organ systems. However, there are certain nutrients that affect bone health.
Calcium and Vitamin D
You already know that calcium is a critical component of bone, especially in the form of calcium phosphate and calcium carbonate. Since the body cannot make calcium, it must be obtained from the diet. However, calcium cannot be absorbed from the small intestine without vitamin D. Therefore, intake of vitamin D is also critical to bone health. In addition to vitamin D’s role in calcium absorption, it also plays a role, though not as clearly understood, in bone remodelling.
Milk and other dairy foods are not the only sources of calcium. This important nutrient is also found in green leafy vegetables, broccoli, and intact salmon and canned sardines with their soft bones. Nuts, beans, seeds, and shellfish provide calcium in smaller quantities.
Except for fatty fish like salmon and tuna, or fortified milk or cereal, vitamin D is not found naturally in many foods. The action of sunlight on the skin triggers the body to produce its own vitamin D (Figure 10.6.1), but many people, especially those of darker complexion and those living in northern latitudes where the sun’s rays are not as strong, are deficient in vitamin D. In cases of deficiency, a doctor can prescribe a vitamin D supplement.
Figure 10.6.1. Synthesis of vitamin D. Sunlight is one source of vitamin D.
Other Nutrients
Vitamin K also supports bone mineralisation and may have a synergistic role with vitamin D in the regulation of bone growth. Green leafy vegetables are a reliable source of vitamin K.
The minerals magnesium and fluoride may also play a role in supporting bone health. While magnesium is only found in trace amounts in the human body, more than 60 percent of it is in the skeleton, suggesting it plays a role in the structure of bone. Fluoride can displace the hydroxyl group in bone’s hydroxyapatite crystals and form fluorapatite. Like its effect on dental enamel, fluorapatite helps stabilise and strengthen bone mineral. Fluoride can also enter spaces within hydroxyapatite crystals, thus increasing their density.
Omega-3 fatty acids have long been known to reduce inflammation in various parts of the body. Inflammation can interfere with the function of osteoblasts, so consuming omega-3 fatty acids, in the diet or in supplements, may also help enhance production of new osseous tissue. Table 10.6.1summarises the role of nutrients in bone health.
Table 10.6.1. Nutrients and bone health
Nutrient
Role in bone health
Calcium
Needed to make calcium phosphate and calcium carbonate, which form the hydroxyapatite crystals that give bone its hardness
Vitamin D
Needed for calcium absorption
Vitamin K
Supports bone mineralisation; may have synergistic effects with vitamin D
Magnesium
Structural component of bone
Fluoride
Structural component of bone
Omega-3 fatty acids
Reduces inflammation that may interfere with osteoblast function
Hormones and Bone Tissue
The endocrine system produces and secretes hormones, many of which interact with the skeletal system. These hormones are involved in controlling bone growth, maintaining bone once it is formed, and remodelling it.
Hormones that influence Osteoblasts and/or Maintain the Matrix
Several hormones are necessary for controlling bone growth and maintaining the bone matrix. The pituitary gland secretes growth hormone (GH), which, as its name implies, controls bone growth in several ways. It triggers chondrocyte proliferation in epiphyseal plates, resulting in the increasing length of long bones. GH also increases calcium retention, which enhances mineralisation, and stimulates osteoblastic activity, which improves bone density.
GH is not alone in stimulating bone growth and maintaining osseous tissue. Thyroxine, a hormone secreted by the thyroid gland promotes osteoblastic activity and the synthesis of bone matrix. During puberty, the sex hormones (oestrogen in girls, testosterone in boys) also come into play. They too promote osteoblastic activity and production of bone matrix, and in addition, are responsible for the growth spurt that often occurs during adolescence. They also promote the conversion of the epiphyseal plate to the epiphyseal line (i.e., cartilage to its bony remnant), thus bringing an end to the longitudinal growth of bones. Additionally, calcitriol, the active form of vitamin D, is produced by the kidneys and stimulates the absorption of calcium and phosphate from the digestive tract.
Ageing and the Skeletal System
Osteoporosis is a disease characterised by a decrease in bone mass that occurs when the rate of bone resorption exceeds the rate of bone formation, a common occurrence as the body ages. Notice how this is different from Paget’s disease. In Paget’s disease, new bone is formed to keep up with the resorption by the overactive osteoclasts, but that new bone is produced haphazardly. In fact, when a physician is evaluating a patient with thinning bone, they will typically test for osteoporosis and Paget’s disease (as well as other diseases). Osteoporosis does not have the elevated blood levels of alkaline phosphatase found in Paget’s disease.
Figure 10.6.2. Graph showing relationship between age and bone mass. Bone density peaks at about 30 years of age. Women lose bone mass more rapidly than men.
While osteoporosis can involve any bone, it most commonly affects the proximal ends of the femur, vertebrae, and wrist. As a result of the loss of bone density, the osseous tissue may not provide adequate support for everyday functions, and something as simple as a sneeze can cause a vertebral fracture. When an elderly person falls and breaks a hip (really, the femur), it is very likely the femur that broke first, which resulted in the fall. Histologically, osteoporosis is characterised by a reduction in the thickness of compact bone and the number and size of trabeculae in cancellous bone.
Figure 10.6.2 shows that women lose bone mass more quickly than men starting at about 50 years of age. This occurs because 50 is the approximate age at which women go through menopause. Not only do their menstrual periods lessen and eventually cease, but their ovaries reduce in size and then cease the production of oestrogen, a hormone that promotes osteoblastic activity and production of bone matrix. Thus, osteoporosis is more common in women than in men, but men can develop it, too. Anyone with a family history of osteoporosis has a greater risk of developing the disease, so the best treatment is prevention, which should start with a childhood diet that includes adequate intake of calcium and vitamin D and a lifestyle that includes weight-bearing exercise. These actions, as discussed above, are important in building bone mass. Promoting proper nutrition and weight-bearing exercise early in life can maximise bone mass before the age of 30, thus reducing the risk of osteoporosis.
For many elderly people, a hip fracture can be life threatening. The fracture itself may not be serious, but the immobility that comes during the healing process can lead to the formation of blood clots that can lodge in the capillaries of the lungs, resulting in respiratory failure; pneumonia due to the lack of poor air exchange that accompanies immobility; pressure sores (bed sores) that allow pathogens to enter the body and cause infections; and urinary tract infections from catheterisation.
Current treatments for managing osteoporosis include bisphosphonates (the same medications often used in Paget’s disease), calcitonin, and oestrogen (for women only). Minimising the risk of falls, for example, by removing tripping hazards, is also an important step in managing the potential outcomes from the disease.
Hormones That Influence Osteoclasts
Bone modelling and remodelling require osteoclasts to resorb unneeded, damaged, or old bone, and osteoblasts to lay down new bone. Two hormones that affect the osteoclasts are parathyroid hormone (PTH) and calcitonin.
PTH stimulates osteoclast proliferation and activity. As a result, calcium is released from the bones into the circulation, thus increasing the calcium ion concentration in the blood. PTH also promotes the reabsorption of calcium by the kidney tubules, which can affect calcium homeostasis (see below).
The small intestine is also affected by PTH, albeit indirectly. Because another function of PTH is to stimulate the synthesis of vitamin D, and because vitamin D promotes intestinal absorption of calcium, PTH indirectly increases calcium uptake by the small intestine. Calcitonin, a hormone secreted by the thyroid gland, has some effects that counteract those of PTH. Calcitonin inhibits osteoclast activity and stimulates calcium uptake by the bones, thus reducing the concentration of calcium ions in the blood. As evidenced by their opposing functions in maintaining calcium homeostasis, PTH and calcitonin are generally not secreted at the same time. Table 10.6.2 summarises the hormones that influence the skeletal system.
Table 10.6.2 Hormones That Affect the Skeletal System
Hormone
Role
Growth hormone
Increases length of long bones, enhances mineralisation, and improves bone density
Thyroxine
Stimulates bone growth and promotes synthesis of bone matrix
Sex hormones
Promote osteoblastic activity and production of bone matrix; responsible for adolescent growth spurt; promote conversion of epiphyseal plate to epiphyseal line
Calcitriol
Stimulates absorption of calcium and phosphate from digestive tract
Parathyroid hormone
Stimulates osteoclast proliferation and resorption of bone by osteoclasts; promotes reabsorption of calcium by kidney tubules; indirectly increases calcium absorption by small intestine
Calcitonin
Inhibits osteoclast activity and stimulates calcium uptake by bones
Section Review
Mechanical stress stimulates the deposition of mineral salts and collagen fibres within bones. Calcium, the predominant mineral in bone, cannot be absorbed from the small intestine if vitamin D is lacking. Vitamin K supports bone mineralisation and may have a synergistic role with vitamin D. Magnesium and fluoride, as structural elements, play a supporting role in bone health. Omega-3 fatty acids reduce inflammation and may promote production of new osseous tissue. Growth hormone increases the length of long bones, enhances mineralisation, and improves bone density. Thyroxine stimulates bone growth and promotes the synthesis of bone matrix. The sex hormones (oestrogen in women; testosterone in men) promote osteoblastic activity and the production of bone matrix, are responsible for the adolescent growth spurt, and promote closure of the epiphyseal plates. Osteoporosis is a disease characterised by decreased bone mass that is common in ageing adults. Calcitriol stimulates the digestive tract to absorb calcium and phosphate. Parathyroid hormone (PTH) stimulates osteoclast proliferation and resorption of bone by osteoclasts. Vitamin D plays a synergistic role with PTH in stimulating the osteoclasts. Additional functions of PTH include promoting reabsorption of calcium by kidney tubules and indirectly increasing calcium absorption from the small intestine. Calcitonin inhibits osteoclast activity and stimulates calcium uptake by bones.
10.7 Calcium Homeostasis: Interactions of the Skeletal System and Other Organ Systems
Learning Objectives
By the end of this section, you will be able to:
Describe the effect of too much or too little calcium on the body
Explain the process of calcium homeostasis
Calcium is not only the most abundant mineral in bone, it is also the most abundant mineral in the human body. Calcium ions are needed not only for bone mineralisation but for tooth health, regulation of the heart rate and strength of contraction, blood coagulation, contraction of smooth and skeletal muscle cells, and regulation of nerve impulse conduction. The normal concentration of calcium in the blood is about 10 mg/dL. When the body cannot maintain this level, a person will experience hypo- or hypercalcaemia.
Hypocalcaemia, a condition characterised by abnormally low levels of calcium, can have an adverse effect on several different body systems including circulation, muscles, nerves, and bone. Without adequate calcium, blood has difficulty coagulating, the heart may skip beats or stop beating altogether, muscles may have difficulty contracting, nerves may have difficulty functioning, and bones may become brittle. The causes of hypocalcaemia can range from hormonal imbalances to an improper diet. Treatments vary according to the cause, but prognoses are generally good.
Conversely, in hypercalcaemia, a condition characterised by abnormally elevated levels of calcium, the nervous system is underactive, which results in lethargy, sluggish reflexes, constipation and loss of appetite, confusion, and in severe cases, coma.
Obviously, calcium homeostasis is critical. The skeletal, endocrine, and digestive systems play a role in this, but the kidneys do, too. These body systems work together to maintain a normal calcium concentration in the blood (Figure 10.7.1).
Figure 10.7.1. Pathways in calcium homeostasis. The body regulates calcium homeostasis with two pathways; one is signalled to turn on when blood calcium levels drop below normal and one is the pathway that is signalled to turn on when blood calcium levels are elevated.
Calcium is a chemical element that cannot be produced by any biological processes. The only way it can enter the body is through the diet. The bones act as a storage site for calcium: The body deposits calcium in the bones when blood levels get too high, and it releases calcium when blood levels drop too low. This process is regulated by PTH, vitamin D, and calcitonin.
Cells of the parathyroid gland have plasma membrane receptors for calcium. When calcium is not binding to these receptors, the cells release PTH, which stimulates osteoclast proliferation and resorption of bone by osteoclasts. This demineralisation process releases calcium into the blood. PTH promotes reabsorption of calcium from the urine by the kidneys, so that the calcium returns to the blood. Finally, PTH stimulates the synthesis of vitamin D, which in turn, stimulates calcium absorption from any digested food in the small intestine.
When all these processes return blood calcium levels to normal, there is enough calcium to bind with the receptors on the surface of the cells of the parathyroid glands, and this cycle of events is turned off (Figure 10.7.1).
When blood concentrations of calcium get too high, the thyroid gland is stimulated to release calcitonin (Figure 10.7.1), which inhibits osteoclast activity and stimulates calcium uptake by the bones, but also decreases reabsorption of calcium by the kidneys. All these actions lower blood levels of calcium. When blood calcium levels return to normal, the thyroid gland stops secreting calcitonin.
Section Review
Calcium homeostasis, i.e., maintaining a blood calcium concentration of about 10 mg/dL, is critical for normal body functions. Hypocalcaemia can result in problems with blood coagulation, muscle contraction, nerve functioning, and bone strength. Hypercalcaemia can result in lethargy, sluggish reflexes, constipation and loss of appetite, confusion, and coma. Calcium homeostasis is controlled by PTH, vitamin D, and calcitonin and the interactions of the skeletal, endocrine, digestive and urinary systems.
Distinguish between the axial skeleton and appendicular skeleton
Define the axial skeleton and its components
Define the appendicular skeleton and its components
The skeletal system includes all the bones, cartilages, and ligaments of the body that support and give shape to the body and body structures. The skeleton consists of the bones of the body. For adults, there are 206 bones in the skeleton. Younger individuals have higher numbers of bones because some bones fuse together during childhood and adolescence to form an adult bone. The primary functions of the skeleton are to provide a rigid, internal structure that can support the weight of the body against the force of gravity, and to provide a structure upon which muscles can act to produce movements of the body. The lower portion of the skeleton is specialised for stability during walking or running. In contrast, the upper skeleton has greater mobility and ranges of motion, features that allow you to lift and carry objects or turn your head and trunk.
In addition to providing for support and movements of the body, the skeleton has protective and storage functions. It protects the internal organs, including the brain, spinal cord, heart, lungs, and pelvic organs. The bones of the skeleton serve as the primary storage site for important minerals such as calcium and phosphate. The bone marrow found within bones stores fat and houses the blood-cell producing tissue of the body.
The skeleton is subdivided into two major divisions—the axial and appendicular.
The Axial Skeleton
The skeleton is subdivided into two major divisions—the axial and appendicular. The axial skeleton forms the vertical, central axis of the body and includes all bones of the head, neck, chest, and back (Figure 10.8.1). It serves to protect the brain, spinal cord, heart, and lungs. It also serves as the attachment site for muscles that move the head, neck, and back, and for muscles that act across the shoulder and hip joints to move their corresponding limbs.
The axial skeleton of the adult consists of 80 bones, including the skull, the vertebral column, and the thoracic cage. The skull is formed by 22 bones. Also associated with the head are an additional seven bones, including the hyoid bone and the ear ossicles (three small bones found in each middle ear). The vertebral column consists of 24 bones, each called a vertebra, plus the sacrum and coccyx. The thoracic cage includes the 12 pairs of ribs, and the sternum, the flattened bone of the anterior chest.
Figure 10.8.1 Axial and appendicular skeleton. The axial skeleton supports the head, neck, back, and chest and thus forms the vertical axis of the body. It consists of the skull, vertebral column (including the sacrum and coccyx), and the thoracic cage, formed by the ribs and sternum. The appendicular skeleton is made up of all bones of the upper and lower limbs.
The Appendicular Skeleton
The appendicular skeleton includes all bones of the upper and lower limbs, plus the bones that attach each limb to the axial skeleton. There are 126 bones in the appendicular skeleton of an adult. The bones of the appendicular skeleton are covered in a separate chapter.
Section Review
The skeletal system includes all the bones, cartilages, and ligaments of the body. It serves to support the body, protect the brain and other internal organs, and provides a rigid structure upon which muscles can pull to generate body movements. It also stores fat and the tissue responsible for the production of blood cells. The skeleton is subdivided into two parts. The axial skeleton forms a vertical axis that includes the head, neck, back, and chest. It has 80 bones and consists of the skull, vertebral column, and thoracic cage. The adult vertebral column consists of 24 vertebrae plus the sacrum and coccyx. The thoracic cage is formed by 12 pairs of ribs and the sternum. The appendicular skeleton consists of 126 bones in the adult and includes all the bones of the upper and lower limbs plus the bones that anchor each limb to the axial skeleton.
List and identify the bones of the brain case and face
Locate the major suture lines of the skull and name the bones associated with each
Locate and define the boundaries of the anterior, middle and posterior cranial fossae, the temporal fossa and infratemporal fossa
Define the paranasal sinuses and identify the location of each
Name the bones that make up the walls of the orbit and identify the openings associated with the orbit
Identify the bones and structures that form the nasal septum and nasal conchae, and locate the hyoid bone
Identify the bony openings of the skull
The cranium (skull) is the skeletal structure of the head that supports the face and protects the brain. It is subdivided into the facial bones and the brain case, or cranial vault (Figure 10.9.1). The facial bones underlie the facial structures, form the nasal cavity, enclose the eyeballs, and support the teeth of the upper and lower jaws. The rounded brain case surrounds and protects the brain and houses the middle and inner ear structures.
In the adult, the skull consists of 22 individual bones, 21 of which are immobile and united into a single unit. The 22nd bone is the mandible (lower jaw), which is the only moveable bone of the skull.
Figure 10.9.1 Parts of the skull. The skull consists of the rounded brain case that houses the brain and the facial bones that form the upper and lower jaws, nose, orbits, and other facial structures.
Anterior View of the Skull
The anterior skull consists of the facial bones and provides the bony support for the eyes and structures of the face. This view of the skull is dominated by the openings of the orbits and the nasal cavity. Also seen are the upper and lower jaws, with their respective teeth (Figure 10.9.2).
The orbit is the bony socket that houses the eyeball and muscles that move the eyeball or open the upper eyelid. The upper margin of the anterior orbit is the supraorbital margin. Located near the midpoint of the supraorbital margin is a small opening called the supraorbital foramen. This provides for passage of a sensory nerve to the skin of the forehead. Below the orbit is the infraorbital foramen, which is the point of emergence for a sensory nerve that supplies the anterior face below the orbit.
Figure 10.9.2. Anterior view of the skull. An anterior view of the skull shows the bones that form the forehead, orbits (eye sockets), nasal cavity, nasal septum, and upper and lower jaws.
Inside the nasal area of the skull, the nasal cavity is divided into halves by the nasal septum. The upper portion of the nasal septum is formed by the perpendicular plate of the ethmoid bone and the lower portion is the vomer bone. Each side of the nasal cavity is triangular in shape, with a broad inferior space that narrows superiorly. When looking into the nasal cavity from the front of the skull, two bony plates are seen projecting from each lateral wall. The larger of these is the inferior nasal concha, an independent bone of the skull. Located just above the inferior concha is the middle nasal concha, which is part of the ethmoid bone. A third bony plate, also part of the ethmoid bone, is the superior nasal concha. It is much smaller and out of sight, above the middle concha. The superior nasal concha is located just lateral to the perpendicular plate, in the upper nasal cavity.
Lateral View of the Skull
A view of the lateral skull is dominated by the large, rounded brain case above and the upper and lower jaws with their teeth below (Figure 10.9.3). Separating these areas is the bridge of bone called the zygomatic arch. The zygomatic arch is the bony arch on the side of skull that spans from the area of the cheek to just above the ear canal. It is formed by the junction of two bony processes: a short anterior component, the temporal process of the zygomatic bone (the cheekbone) and a longer posterior portion, the zygomatic process of the temporal bone, extending forward from the temporal bone. Thus, the temporal process (anteriorly) and the zygomatic process (posteriorly) join, like the two ends of a drawbridge, to form the zygomatic arch. One of the major muscles that pulls the mandible upward during biting and chewing arises from the zygomatic arch.
On the lateral side of the brain case, above the level of the zygomatic arch, is a shallow space called the temporal fossa. Below the level of the zygomatic arch and deep to the vertical portion of the mandible is another space called the infratemporal fossa. Both the temporal fossa and infratemporal fossa contain muscles that act on the mandible during chewing.
Figure 10.9.3 Lateral view of skull. The lateral skull shows the large, rounded brain case, zygomatic arch, and the upper and lower jaws. The zygomatic arch is formed jointly by the zygomatic process of the temporal bone and the temporal process of the zygomatic bone. The shallow space above the zygomatic arch is the temporal fossa. The space inferior to the zygomatic arch and deep to the posterior mandible is the infratemporal fossa.
Bones of the Brain Case
The brain case contains and protects the brain. The interior space that is almost completely occupied by the brain is called the cranial cavity. This cavity is bounded superiorly by the rounded top of the skull, which is called the calvaria (skullcap), and the lateral and posterior sides of the skull. The bones that form the top and sides of the brain case are usually referred to as the “flat” bones of the skull.
The floor of the brain case is referred to as the base of the skull. This is a complex area that varies in depth and has numerous openings for the passage of cranial nerves, blood vessels, and the spinal cord. Inside the skull, the base is subdivided into three large spaces, called the anterior cranial fossa, middle cranial fossa, and posterior cranial fossa (fossa = “trench or ditch”) (Figure 10.9.4). From anterior to posterior, the fossae increase in depth. The shape and depth of each fossa corresponds to the shape and size of the brain region that each houses. The boundaries and openings of the cranial fossae (singular = fossa) will be described in a later section.
Figure 10.9.4 Cranial fossae. The bones of the brain case surround and protect the brain, which occupies the cranial cavity. The base of the brain case, which forms the floor of cranial cavity, is subdivided into the shallow anterior cranial fossa, the middle cranial fossa, and the deep posterior cranial fossa.
The brain case consists of eight bones. These include the paired parietal and temporal bones, plus the unpaired frontal, occipital, sphenoid, and ethmoid bones.
Parietal Bone
The parietal bone forms most of the upper lateral side of the skull (see Figure 10.9.3). These are paired bones, with the right and left parietal bones joining together at the top of the skull. Each parietal bone is also bounded anteriorly by the frontal bone, inferiorly by the temporal bone, and posteriorly by the occipital bone.
Temporal Bone
The temporal bone forms the lower lateral side of the skull (see Figure 10.9.3). Common wisdom has it that the temporal bone (temporal = “time”) is so named because this area of the head (the temple) is where hair typically first turns gray, indicating the passage of time.
The temporal bone is subdivided into several regions (Figure 10.9.5). The flattened, upper portion is the squamous portion of the temporal bone. Below this area and projecting anteriorly is the zygomatic process of the temporal bone, which forms the posterior portion of the zygomatic arch. Posteriorly is the mastoid portion of the temporal bone. Projecting inferiorly from this region is a large prominence, the mastoid process, which serves as a muscle attachment site. The mastoid process can easily be felt on the side of the head just behind your earlobe. On the interior of the skull, the petrous portion of each temporal bone forms the prominent, diagonally oriented petrous ridge in the floor of the cranial cavity. Located inside each petrous ridge are small cavities that house the structures of the middle and inner ears.
Figure 10.9.5 Temporal bone. A lateral view of the isolated temporal bone shows the squamous, mastoid, and zygomatic portions of the temporal bone.
Important landmarks of the temporal bone, as shown in Figure 10.9.6, include the following:
External acoustic meatus (ear canal)—This is the large opening on the lateral side of the skull that is associated with the ear.
Internal acoustic meatus—This opening is located inside the cranial cavity, on the medial side of the petrous ridge. It connects to the middle and inner ear cavities of the temporal bone.
Mandibular fossa—This is the deep, oval-shaped depression located on the external base of the skull, just in front of the external acoustic meatus. The mandible (lower jaw) joins with the skull at this site as part of the temporomandibular joint, which allows for movements of the mandible during opening and closing of the mouth.
Articular tubercle—The smooth ridge located immediately anterior to the mandibular fossa. Both the articular tubercle and mandibular fossa contribute to the temporomandibular joint, the joint that provides for movements between the temporal bone of the skull and the mandible.
Styloid process—Posterior to the mandibular fossa on the external base of the skull is an elongated, downward bony projection called the styloid process, so named because of its resemblance to a stylus (a pen or writing tool). This structure serves as an attachment site for several small muscles and for a ligament that supports the hyoid bone of the neck. (See also Figure 10.9.5)
Stylomastoid foramen—This small opening is located between the styloid process and mastoid process. This is the point of exit for the cranial nerve that supplies the facial muscles.
Carotid canal—The carotid canal is a zig-zag shaped tunnel that provides passage through the base of the skull for one of the major arteries that supplies the brain. Its entrance is located on the outside base of the skull, anteromedial to the styloid process. The canal then runs anteromedially within the bony base of the skull, and then turns upward to its exit in the floor of the middle cranial cavity, above the foramen lacerum.
Figure 10.9.6. External and internal views of base of skull. (a) The hard palate is formed anteriorly by the palatine processes of the maxilla bones and posteriorly by the horizontal plate of the palatine bones. (b) The complex floor of the cranial cavity is formed by the frontal, ethmoid, sphenoid, temporal, and occipital bones. The lesser wing of the sphenoid bone separates the anterior and middle cranial fossae. The petrous ridge (petrous portion of temporal bone) separates the middle and posterior cranial fossae.
Frontal Bone
The frontal bone is the single bone that forms the forehead. At its anterior midline, between the eyebrows, there is a slight depression called the glabella (see Figure 10.9.3). The frontal bone also forms the supraorbital margin of the orbit. Near the middle of this margin, is the supraorbital foramen, the opening that provides passage for a sensory nerve to the forehead. The frontal bone is thickened just above each supraorbital margin, forming rounded brow ridges. These are located just behind your eyebrows and vary in size among individuals, although they are larger in males. Inside the cranial cavity, the frontal bone extends posteriorly. This flattened region forms both the roof of the orbit below and the floor of the anterior cranial cavity above (see Figure 10.9.6b).
Occipital Bone
The occipital bone is the single bone that forms the posterior skull and posterior base of the cranial cavity (Figure 10.9.7; see also Figure 10.9.6). On its outside surface, at the posterior midline, is a small protrusion called the external occipital protuberance, which serves as an attachment site for a ligament of the posterior neck. Lateral to either side of this bump is a superior nuchal line (nuchal = “nape” or “posterior neck”). The nuchal lines represent the most superior point at which muscles of the neck attach to the skull, with only the scalp covering the skull above these lines. On the base of the skull, the occipital bone contains the large opening of the foramen magnum, which allows for passage of the spinal cord as it exits the skull. On either side of the foramen magnum is an oval-shaped occipital condyle. These condyles form joints with the first cervical vertebra and thus support the skull on top of the vertebral column.
Figure 10.9.7 Posterior view of skull. This view of the posterior skull shows attachment sites for muscles and joints that support the skull.
Sphenoid Bone
The sphenoid bone is a single, complex bone of the central skull (Figure 10.9.8). It serves as a “keystone” bone because it joins with almost every other bone of the skull. The sphenoid forms much of the base of the central skull (see Figure 10.9.6 and also extends laterally to contribute to the sides of the skull (see Figure 10.9.3). Inside the cranial cavity, the right and left lesser wings of the sphenoid bone, which resemble the wings of a flying bird, form the lip of a prominent ridge that marks the boundary between the anterior and middle cranial fossae. The sella turcica (“Turkish saddle”) is located at the midline of the middle cranial fossa. This bony region of the sphenoid bone is named for its resemblance to the horse saddles used by the Ottoman Turks, with a high back and a tall front. The rounded depression in the floor of the sella turcica is the hypophyseal (pituitary) fossa, which houses the pea-sized pituitary (hypophyseal) gland. The greater wings of the sphenoid bone extend laterally to either side away from the sella turcica, where they form the anterior floor of the middle cranial fossa. The greater wing is best seen on the outside of the lateral skull, where it forms a rectangular area immediately anterior to the squamous portion of the temporal bone.
On the inferior aspect of the skull, each half of the sphenoid bone forms two thin, vertically oriented bony plates. These are the medial pterygoid plate and lateral pterygoid plate (pterygoid = “wing-shaped”). The right and left medial pterygoid plates form the posterior, lateral walls of the nasal cavity. The larger lateral pterygoid plates serve as attachment sites for chewing muscles that fill the infratemporal space and act on the mandible.
Figure 10.9.8 Sphenoid bone. Shown in isolation in (a) superior and (b) posterior views, the sphenoid bone is a single midline bone that forms the anterior walls and floor of the middle cranial fossa. It has a pair of lesser wings and a pair of greater wings. The sella turcica surrounds the hypophyseal fossa. Projecting downward are the medial and lateral pterygoid plates. The sphenoid has multiple openings for the passage of nerves and blood vessels, including the optic canal, superior orbital fissure, foramen rotundum, foramen ovale, and foramen spinosum.
Ethmoid Bone
The ethmoid bone is a single, midline bone that forms the roof and lateral walls of the upper nasal cavity, the upper portion of the nasal septum, and contributes to the medial wall of the orbit (Figure 10.9.9 and Figure 10.9.10). On the interior of the skull, the ethmoid also forms a portion of the floor of the anterior cranial cavity (see Figure 10.9.6b).
Within the nasal cavity, the perpendicular plate of the ethmoid bone forms the upper portion of the nasal septum. The ethmoid bone also forms the lateral walls of the upper nasal cavity. Extending from each lateral wall are the superior nasal concha and middle nasal concha, which are thin, curved projections that extend into the nasal cavity (Figure 10.9.11).
In the cranial cavity, the ethmoid bone forms a small area at the midline in the floor of the anterior cranial fossa. This region also forms the narrow roof of the underlying nasal cavity. This portion of the ethmoid bone consists of two parts, the crista galli and cribriform plates. The crista galli (“rooster’s comb or crest”) is a small upward bony projection located at the midline. It functions as an anterior attachment point for one of the covering layers of the brain. To either side of the crista galli is the cribriform plate (cribrum = “sieve”), a small, flattened area with numerous small openings termed olfactory foramina. Small nerve branches from the olfactory areas of the nasal cavity pass through these openings to enter the brain.
The lateral portions of the ethmoid bone are located between the orbit and upper nasal cavity, and thus form the lateral nasal cavity wall and a portion of the medial orbit wall. Located inside this portion of the ethmoid bone are several small, air-filled spaces that are part of the paranasal sinus system of the skull.
Figure 10.9.9 Sagittal section of skull. This midline view of the sagittally sectioned skull shows the nasal septum.Figure 10.9.10. Ethmoid bone. The unpaired ethmoid bone is located at the midline within the central skull. It has an upward projection, the crista galli, and a downward projection, the perpendicular plate, which forms the upper nasal septum. The cribriform plates form both the roof of the nasal cavity and a portion of the anterior cranial fossa floor. The lateral sides of the ethmoid bone form the lateral walls of the upper nasal cavity, part of the medial orbit wall, and give rise to the superior and middle nasal conchae. The ethmoid bone also contains the ethmoid air cells.Figure 10.9.11. Lateral wall of nasal cavity. The three nasal conchae are curved bones that project from the lateral walls of the nasal cavity. The superior nasal concha and middle nasal concha are parts of the ethmoid bone. The inferior nasal concha is an independent bone of the skull.
Sutures of the Skull
A suture is an immobile joint between adjacent bones of the skull. The narrow gap between the bones is filled with dense, fibrous connective tissue that unites the bones. The long sutures located between the bones of the brain case are not straight, but instead follow irregular, tightly twisting paths. These twisting lines serve to tightly interlock the adjacent bones, thus adding strength to the skull for brain protection.
The two suture lines seen on the top of the skull are the coronal and sagittal sutures. The coronal suture runs from side to side across the skull, within the coronal plane of section (see Figure 10.9.3). It joins the frontal bone to the right and left parietal bones. The sagittal suture extends posteriorly from the coronal suture, running along the midline at the top of the skull in the sagittal plane of section (see Figure 10.9.7). It unites the right and left parietal bones. On the posterior skull, the sagittal suture terminates by joining the lambdoid suture. The lambdoid suture extends downward and laterally to either side away from its junction with the sagittal suture. The lambdoid suture joins the occipital bone to the right and left parietal and temporal bones. This suture is named for its upside-down “V” shape, which resembles the capital letter version of the Greek letter lambda (Λ). The squamous suture is located on the lateral skull. It unites the squamous portion of the temporal bone with the parietal bone (see Figure 10.9.3). At the intersection of four bones is the pterion, a small, capital-H-shaped suture line region that unites the frontal bone, parietal bone, squamous portion of the temporal bone, and greater wing of the sphenoid bone. It is the weakest part of the skull. The pterion is located approximately two finger widths above the zygomatic arch and a thumb’s width posterior to the upward portion of the zygomatic bone.
Disorders of the Skeletal System
Head and traumatic brain injuries are major causes of immediate death and disability, with bleeding and infections as possible additional complications. According to the Centres for Disease Control and Prevention (2010), approximately 30 percent of all injury-related deaths in the United States are caused by head injuries. Most head injuries involve falls. They are most common among young children (ages 0–4 years), adolescents (15–19 years), and the elderly (over 65 years). Additional causes vary, but prominent among these are automobile and motorcycle accidents.
Strong blows to the brain-case portion of the skull can produce fractures. These may result in bleeding inside the skull with subsequent injury to the brain. The most common is a linear skull fracture, in which fracture lines radiate from the point of impact. Other fracture types include a comminuted fracture, in which the bone is broken into several pieces at the point of impact, or a depressed fracture, in which the fractured bone is pushed inward. In a contrecoup (counterblow) fracture, the bone at the point of impact is not broken, but instead a fracture occurs on the opposite side of the skull. Fractures of the occipital bone at the base of the skull can occur in this manner, producing a basilar fracture that can damage the artery that passes through the carotid canal.
A blow to the lateral side of the head may fracture the bones of the pterion. The pterion is an important clinical landmark because located immediately deep to it on the inside of the skull is a major branch of an artery that supplies the skull and covering layers of the brain. A strong blow to this region can fracture the bones around the pterion. If the underlying artery is damaged, bleeding can cause the formation of a haematoma (collection of blood) between the brain and interior of the skull. As blood accumulates, it will put pressure on the brain. Symptoms associated with a haematoma may not be apparent immediately following the injury, but if untreated, blood accumulation will exert increasing pressure on the brain and can result in death within a few hours.
Facial Bones of the Skull
The facial bones of the skull form the upper and lower jaws, the nose, nasal cavity and nasal septum, and the orbit. The facial bones include 14 bones, with six paired bones and two unpaired bones. The paired bones are the maxilla, palatine, zygomatic, nasal, lacrimal, and inferior nasal conchae bones. The unpaired bones are the vomer and mandible bones. Although classified with the brain-case bones, the ethmoid bone also contributes to the nasal septum and the walls of the nasal cavity and orbit.
Maxillary Bone
The maxillary bone, often referred to simply as the maxilla (plural = maxillae), is one of a pair that together form the upper jaw, much of the hard palate, the medial floor of the orbit, and the lateral base of the nose (see Figure 10.9.2). The curved, inferior margin of the maxillary bone that forms the upper jaw and contains the upper teeth is the alveolar process of the maxilla (Figure 10.9.12). Each tooth is anchored into a deep socket called an alveolus. On the anterior maxilla, just below the orbit, is the infraorbital foramen. This is the point of exit for a sensory nerve that supplies the nose, upper lip, and anterior cheek. On the inferior skull, the palatine process from each maxillary bone can be seen joining together at the midline to form the anterior three-quarters of the hard palate (see Figure 10.9.6a). The hard palate is the bony plate that forms the roof of the mouth and floor of the nasal cavity, separating the oral and nasal cavities.
Figure 10.9.12. Maxillary bone. The maxillary bone forms the upper jaw and supports the upper teeth. Each maxilla also forms the lateral floor of each orbit and most of the hard palate.
Palatine Bone
The palatine bone is one of a pair of irregularly shaped bones that contribute small areas to the lateral walls of the nasal cavity and the medial wall of each orbit. The largest region of each of the palatine bone is the horizontal plate. The plates from the right and left palatine bones join together at the midline to form the posterior quarter of the hard palate (see Figure 10.9.6a). Thus, the palatine bones are best seen in an inferior view of the skull and hard palate.
Homeostatic Imbalances
Cleft Lip and Cleft Palate
During embryonic development, the right and left maxilla bones come together at the midline to form the upper jaw. At the same time, the muscle and skin overlying these bones join together to form the upper lip. Inside the mouth, the palatine processes of the maxilla bones, along with the horizontal plates of the right and left palatine bones, join together to form the hard palate. If an error occurs in these developmental processes, a birth defect of cleft lip or cleft palate may result.
Cleft lip is a common development defect that affects approximately 1 in every 600-800 newborns in Australia. This defect involves a partial or complete failure of the right and left portions of the upper lip to fuse together, leaving a cleft (gap).
A more severe developmental defect is cleft palate, which affects the hard palate. The hard palate is the bony structure that separates the nasal cavity from the oral cavity. It is formed during embryonic development by the midline fusion of the horizontal plates from the right and left palatine bones and the palatine processes of the maxilla bones. In Australia, cleft palate affects approximately 1 in every 800 newborns. It results from a failure of the two halves of the hard palate to completely come together and fuse at the midline, thus leaving a gap between them. This gap allows for communication between the nasal and oral cavities. In severe cases, the bony gap continues into the anterior upper jaw where the alveolar processes of the maxilla bones also do not properly join together above the front teeth. If this occurs, a cleft lip will also be seen. Because of the communication between the oral and nasal cavities, a cleft palate makes it very difficult for an infant to generate the suckling needed for nursing, thus leaving the infant at risk for malnutrition. Surgical repair is required to correct cleft palate defects.
Zygomatic Bone
The zygomatic bone is also known as the cheekbone. Each of the paired zygomatic bones forms much of the lateral wall of the orbit and the lateral-inferior margins of the anterior orbital opening (see Figure 10.9.2). The short temporal process of the zygomatic bone projects posteriorly, where it forms the anterior portion of the zygomatic arch (see Figure 10.9.3).
Nasal Bone
The nasal bone is one of two small bones that articulate (join) with each other to form the bony base (bridge) of the nose. They also support the cartilages that form the lateral walls of the nose (see Figure 10.9.9). These are the bones that are damaged when the nose is broken.
Lacrimal Bone
Each lacrimal bone is a small, rectangular bone that forms the anterior, medial wall of the orbit (see Figure 10.9.2 and Figure 10.9.3). The anterior portion of the lacrimal bone forms a shallow depression called the lacrimal fossa and extending inferiorly from this is the nasolacrimal canal. The lacrimal fluid (tears of the eye), which serves to maintain the moist surface of the eye, drains at the medial corner of the eye into the nasolacrimal canal. This duct then extends downward to open into the nasal cavity, behind the inferior nasal concha. In the nasal cavity, the lacrimal fluid normally drains posteriorly, but with an increased flow of tears due to crying or eye irritation, some fluid will also drain anteriorly, thus causing a runny nose.
Inferior Nasal Conchae
The right and left inferior nasal conchae form a curved bony plate that projects into the nasal cavity space from the lower lateral wall (see Figure 10.9.11). The inferior concha is the largest of the nasal conchae and can easily be seen when looking into the anterior opening of the nasal cavity.
Vomer Bone
The unpaired vomer bone, often referred to simply as the vomer, is triangular-shaped and forms the posterior-inferior part of the nasal septum (see Figure 10.9.9). The vomer is best seen when looking from behind into the posterior openings of the nasal cavity (see Figure 10.9.6a). In this view, the vomer is seen to form the entire height of the nasal septum. A much smaller portion of the vomer can also be seen when looking into the anterior opening of the nasal cavity.
Mandible
The mandible forms the lower jaw and is the only moveable bone of the skull. At the time of birth, the mandible consists of paired right and left bones, but these fuse together during the first year to form the single U-shaped mandible of the adult skull. Each side of the mandible consists of a horizontal body and posteriorly, a vertically oriented ramus of the mandible (ramus = “branch”). The outside margin of the mandible, where the body and ramus come together is called the angle of the mandible (Figure 10.9.13).
The ramus on each side of the mandible has two upward-going bony projections. The more anterior projection is the flattened coronoid process of the mandible, which provides attachment for one of the biting muscles. The posterior projection is the condylar process of the mandible, which is topped by the oval-shaped condyle. The condyle of the mandible articulates (joins) with the mandibular fossa and articular tubercle of the temporal bone. Together these articulations form the temporomandibular joint, which allows for opening and closing of the mouth (see Figure 10.9.3). The broad U-shaped curve located between the coronoid and condylar processes is the mandibular notch.
Important landmarks for the mandible include the following:
Alveolar process of the mandible—This is the upper border of the mandibular body and serves to anchor the lower teeth.
Mental protuberance—The forward projection from the inferior margin of the anterior mandible that forms the chin (mental = “chin”).
Mental foramen—The opening located on each side of the anterior-lateral mandible, which is the exit site for a sensory nerve that supplies the chin.
Mylohyoid line—This bony ridge extends along the inner aspect of the mandibular body (see Figure 10.9.9). The muscle that forms the floor of the oral cavity attaches to the mylohyoid lines on both sides of the mandible.
Mandibular foramen—This opening is located on the medial side of the ramus of the mandible. The opening leads into a tunnel that runs down the length of the mandibular body. The sensory nerve and blood vessels that supply the lower teeth enter the mandibular foramen and then follow this tunnel. Thus, to numb the lower teeth prior to dental work, the dentist must inject anaesthesia into the lateral wall of the oral cavity at a point prior to where this sensory nerve enters the mandibular foramen.
Lingula—This small flap of bone is named for its shape (lingula = “little tongue”). It is located immediately next to the mandibular foramen, on the medial side of the ramus. A ligament that anchors the mandible during opening and closing of the mouth extends down from the base of the skull and attaches to the lingula.
Figure 10.9.13. Isolated mandible. The mandible is the only moveable bone of the skull.
The Orbit
The orbit is the bony socket that houses the eyeball and contains the muscles that move the eyeball or open the upper eyelid. Each orbit is cone-shaped, with a narrow posterior region that widens toward the large anterior opening. To help protect the eye, the bony margins of the anterior opening are thickened and somewhat constricted. The medial walls of the two orbits are parallel to each other but each lateral wall diverges away from the midline at a 45° angle. This divergence provides greater lateral peripheral vision.
The walls of each orbit include contributions from seven skull bones (Figure 10.9.14). The frontal bone forms the roof and the zygomatic bone forms the lateral wall and lateral floor. The medial floor is primarily formed by the maxilla, with a small contribution from the palatine bone. The ethmoid bone and lacrimal bone make up much of the medial wall and the sphenoid bone forms the posterior orbit.
At the posterior apex of the orbit is the opening of the optic canal, which allows for passage of the optic nerve from the retina to the brain. Lateral to this is the elongated and irregularly shaped superior orbital fissure, which provides passage for the artery that supplies the eyeball, sensory nerves, and the nerves that supply the muscles involved in eye movements.
Figure 10.9.14. Bones of the orbit. Seven skull bones contribute to the walls of the orbit. Opening into the posterior orbit from the cranial cavity are the optic canal and superior orbital fissure.
The Nasal Septum and Nasal Chonchae
The nasal septum consists of both bone and cartilage components (Figure 10.9.15; see also Figure 10.9.9). The upper portion of the septum is formed by the perpendicular plate of the ethmoid bone. The lower and posterior parts of the septum are formed by the triangular-shaped vomer bone. In an anterior view of the skull, the perpendicular plate of the ethmoid bone is easily seen inside the nasal opening as the upper nasal septum, but only a small portion of the vomer is seen as the inferior septum. A better view of the vomer bone is seen when looking into the posterior nasal cavity with an inferior view of the skull, where the vomer forms the full height of the nasal septum. The anterior nasal septum is formed by the septal cartilage, a flexible plate that fills in the gap between the perpendicular plate of the ethmoid and vomer bones. This cartilage also extends outward into the nose where it separates the right and left nostrils.
Attached to the lateral wall on each side of the nasal cavity are the superior, middle, and inferior nasal conchae (singular = concha), which are named for their positions (see Figure 10.9.11). These are bony plates that curve downward as they project into the space of the nasal cavity. They serve to swirl the incoming air, which helps to warm and humidify it before the air moves into the delicate air sacs of the lungs. This also allows mucus, secreted by the tissue lining the nasal cavity, to trap incoming dust, pollen, bacteria, and viruses. The largest of the conchae is the inferior nasal concha, which is an independent bone of the skull. The middle concha and the superior conchae, which is the smallest, are both formed by the ethmoid bone. When looking into the anterior nasal opening of the skull, only the inferior and middle conchae can be seen. The small superior nasal concha is well hidden above and behind the middle concha.
Figure 10.9.15. Nasal septum. The nasal septum is formed by the perpendicular plate of the ethmoid bone and the vomer bone. The septal cartilage fills the gap between these bones and extends into the nose.
Cranial Fossae
Inside the skull, the floor of the cranial cavity is subdivided into three cranial fossae (spaces), which increase in depth from anterior to posterior (see Figure 10.9.4, Figure 10.9.6b, and Figure 10.9.9). Since the brain occupies these areas, the shape of each conforms to the shape of the brain regions that it contains. Each cranial fossa has anterior and posterior boundaries and is divided at the midline into right and left areas by a significant bony structure or opening.
Anterior Cranial Fossa
The anterior cranial fossa is the most anterior and the shallowest of the three cranial fossae. It overlies the orbits and contains the frontal lobes of the brain. Anteriorly, the anterior fossa is bounded by the frontal bone, which also forms most of the floor for this space. The lesser wings of the sphenoid bone form the prominent ledge that marks the boundary between the anterior and middle cranial fossae. Located in the floor of the anterior cranial fossa at the midline is a portion of the ethmoid bone, consisting of the upward projecting crista galli and to either side of this, the cribriform plates.
Middle Cranial Fossa
The middle cranial fossa is deeper and situated posterior to the anterior fossa. It extends from the lesser wings of the sphenoid bone anteriorly, to the petrous ridges (petrous portion of the temporal bones) posteriorly. The large, diagonally positioned petrous ridges give the middle cranial fossa a butterfly shape, making it narrow at the midline and broad laterally. The temporal lobes of the brain occupy this fossa. The middle cranial fossa is divided at the midline by the upward bony prominence of the sella turcica, a part of the sphenoid bone. The middle cranial fossa has several openings for the passage of blood vessels and cranial nerves (see Figure 10.9.6).
Openings in the middle cranial fossa are as follows:
Optic canal—This opening is located at the anterior lateral corner of the sella turcica. It provides for passage of the optic nerve into the orbit.
Superior orbital fissure—This large, irregular opening into the posterior orbit is located on the anterior wall of the middle cranial fossa, lateral to the optic canal and under the projecting margin of the lesser wing of the sphenoid bone. Nerves to the eyeball and associated muscles, and sensory nerves to the forehead pass through this opening.
Foramen rotundum—This rounded opening (rotundum = “round”) is located in the floor of the middle cranial fossa, just inferior to the superior orbital fissure. It is the exit point for a major sensory nerve that supplies the cheek, nose, and upper teeth.
Foramen ovale of the middle cranial fossa—This large, oval-shaped opening in the floor of the middle cranial fossa provides passage for a major sensory nerve to the lateral head, cheek, chin, and lower teeth.
Foramen spinosum—This small opening, located posterior-lateral to the foramen ovale, is the entry point for an important artery that supplies the covering layers surrounding the brain. The branching pattern of this artery forms readily visible grooves on the internal surface of the skull and these grooves can be traced back to their origin at the foramen spinosum.
Carotid canal—This is the zig-zag passageway through which a major artery to the brain enters the skull. The entrance to the carotid canal is located on the inferior aspect of the skull, anteromedial to the styloid process (see Figure 10.9.6a). From here, the canal runs anteromedially within the bony base of the skull. Just above the foramen lacerum, the carotid canal opens into the middle cranial cavity, near the posterior-lateral base of the sella turcica.
Foramen lacerum—This irregular opening is located in the base of the skull, immediately inferior to the exit of the carotid canal. This opening is an artifact of the dry skull, because in life it is filled with cartilage. All the openings of the skull that provide for passage of nerves or blood vessels have smooth margins; the word lacerum (“ragged” or “torn”) tells us that this opening has ragged edges and thus nothing passes through it.
Posterior Cranial Fossa
The posterior cranial fossa is the most posterior and deepest portion of the cranial cavity. It contains the cerebellum of the brain. The posterior fossa is bounded anteriorly by the petrous ridges, while the occipital bone forms the floor and posterior wall. It is divided at the midline by the large foramen magnum (“great aperture”), the opening that provides for passage of the spinal cord.
Located on the medial wall of the petrous ridge in the posterior cranial fossa is the internal acoustic meatus (see Figure 10.9.9). This opening provides for passage of the nerve from the hearing and equilibrium organs of the inner ear, and the nerve that supplies the muscles of the face. Located at the anterior-lateral margin of the foramen magnum is the hypoglossal canal. These emerge on the inferior aspect of the skull at the base of the occipital condyle and provide passage for an important nerve to the tongue.
Immediately inferior to the internal acoustic meatus is the large, irregularly shaped jugular foramen (see Figure 10.9.6a). Several cranial nerves from the brain exit the skull via this opening. It is also the exit point through the base of the skull for all the venous return blood leaving the brain. The venous structures that carry blood inside the skull form large, curved grooves on the inner walls of the posterior cranial fossa, which terminate at each jugular foramen.
Paranasal Sinuses
The paranasal sinuses are hollow, air-filled spaces located within certain bones of the skull (Figure 10.9.16). All the sinuses communicate with the nasal cavity (paranasal = “next to nasal cavity”) and are lined with nasal mucosa. They serve to reduce bone mass and thus lighten the skull, and they also add resonance to the voice. This second feature is most obvious when you have a cold or sinus congestion when swelling of the mucosa and excess mucus production can obstruct the narrow passageways between the sinuses and the nasal cavity, causing your voice to sound different to yourself and others. This blockage can also allow the sinuses to fill with fluid, with the resulting pressure producing pain and discomfort.
The paranasal sinuses are named for the skull bone that each occupies. The frontal sinus is located just above the eyebrows, within the frontal bone (see Figure 10.9.15). This irregular space may be divided at the midline into bilateral spaces, or these may be fused into a single sinus space. The frontal sinus is the most anterior of the paranasal sinuses. The largest sinus is the maxillary sinus. These are paired and located within the right and left maxillary bones, where they occupy the area just below the orbits. The maxillary sinuses are most commonly involved during sinus infections. Because their connection to the nasal cavity is located high on their medial wall, they are difficult to drain. The sphenoid sinus is a single, midline sinus. It is located within the body of the sphenoid bone, just anterior and inferior to the sella turcica, thus making it the most posterior of the paranasal sinuses. The lateral aspects of the ethmoid bone contain multiple small spaces separated by very thin bony walls. Each of these spaces is called an ethmoid air cell. These are located on both sides of the ethmoid bone, between the upper nasal cavity and medial orbit, just behind the superior nasal conchae.
Figure 10.9.16. Paranasal sinuses. The paranasal sinuses are hollow, air-filled spaces named for the skull bone that each occupies. The most anterior is the frontal sinus, located in the frontal bone above the eyebrows. The largest are the maxillary sinuses, located in the right and left maxillary bones below the orbits. The most posterior is the sphenoid sinus, located in the body of the sphenoid bone, under the sella turcica. The ethmoid air cells are multiple small spaces located in the right and left sides of the ethmoid bone, between the medial wall of the orbit and lateral wall of the upper nasal cavity.
Hyoid Bone
The hyoid bone is an independent bone that does not contact any other bone and thus is not part of the skull (Figure 10.9.17). It is a small U-shaped bone located in the upper neck near the level of the inferior mandible, with the tips of the “U” pointing posteriorly. The hyoid serves as the base for the tongue above and is attached to the larynx below and the pharynx posteriorly. The hyoid is held in position by a series of small muscles that attach to it either from above or below. These muscles act to move the hyoid up/down or forward/back. Movements of the hyoid are coordinated with movements of the tongue, larynx, and pharynx during swallowing and speaking.
Figure 10.9.17. Hyoid bone. The hyoid bone is located in the upper neck and does not join with any other bone. It provides attachments for muscles that act on the tongue, larynx, and pharynx.
Section Review
The skull consists of the brain case and the facial bones. The brain case surrounds and protects the brain, which occupies the cranial cavity inside the skull. It consists of the rounded calvaria and a complex base. The brain case is formed by eight bones, the paired parietal and temporal bones plus the unpaired frontal, occipital, sphenoid, and ethmoid bones. The narrow gap between the bones is filled with dense, fibrous connective tissue that unites the bones. The sagittal suture joins the right and left parietal bones. The coronal suture joins the parietal bones to the frontal bone, the lamboid suture joins them to the occipital bone, and the squamous suture joins them to the temporal bone.
The facial bones support the facial structures and form the upper and lower jaws. These consist of 14 bones, with the paired maxillary, palatine, zygomatic, nasal, lacrimal, and inferior conchae bones and the unpaired vomer and mandible bones. The ethmoid bone also contributes to the formation of facial structures. The maxilla forms the upper jaw and the mandible forms the lower jaw. The maxilla also forms the larger anterior portion of the hard palate, which is completed by the smaller palatine bones that form the posterior portion of the hard palate.
The floor of the cranial cavity increases in depth from front to back and is divided into three cranial fossae. The anterior cranial fossa is located between the frontal bone and lesser wing of the sphenoid bone. A small area of the ethmoid bone, consisting of the crista galli and cribriform plates, is located at the midline of this fossa. The middle cranial fossa extends from the lesser wing of the sphenoid bone to the petrous ridge (petrous portion of temporal bone). The right and left sides are separated at the midline by the sella turcica, which surrounds the shallow hypophyseal fossa. Openings through the skull in the floor of the middle fossa include the optic canal and superior orbital fissure, which open into the posterior orbit, the foramen rotundum, foramen ovale, and foramen spinosum, and the exit of the carotid canal with its underlying foramen lacerum. The deep posterior cranial fossa extends from the petrous ridge to the occipital bone. Openings here include the large foramen magnum, plus the internal acoustic meatus, jugular foramina, and hypoglossal canals. Additional openings located on the external base of the skull include the stylomastoid foramen and the entrance to the carotid canal.
The anterior skull has the orbits that house the eyeballs and associated muscles. The walls of the orbit are formed by contributions from seven bones: the frontal, zygomatic, maxillary, palatine, ethmoid, lacrimal, and sphenoid. Located at the superior margin of the orbit is the supraorbital foramen, and below the orbit is the infraorbital foramen. The mandible has two openings, the mandibular foramen on its inner surface and the mental foramen on its external surface near the chin. The nasal conchae are bony projections from the lateral walls of the nasal cavity. The large inferior nasal concha is an independent bone, while the middle and superior conchae are parts of the ethmoid bone. The nasal septum is formed by the perpendicular plate of the ethmoid bone, the vomer bone, and the septal cartilage. The paranasal sinuses are air-filled spaces located within the frontal, maxillary, sphenoid, and ethmoid bones.
On the lateral skull, the zygomatic arch consists of two parts, the temporal process of the zygomatic bone anteriorly and the zygomatic process of the temporal bone posteriorly. The temporal fossa is the shallow space located on the lateral skull above the level of the zygomatic arch. The infratemporal fossa is located below the zygomatic arch and deep to the ramus of the mandible.
The hyoid bone is located in the upper neck and does not join with any other bone. It is held in position by muscles and serves to support the tongue above, the larynx below, and the pharynx posteriorly.
Describe each region of the vertebral column and the number of bones in each region
Discuss the curves of the vertebral column and how these change after birth
Describe a typical vertebra and determine the distinguishing characteristics for vertebrae in each vertebral region and features of the sacrum and the coccyx
Define the structure of an intervertebral disc
Determine the location of the ligaments that provide support for the vertebral column
The vertebral column is also known as the spinal column or spine (Figure 10.10.1). It consists of a sequence of vertebrae (singular = vertebra), each of which is separated and united by an intervertebral disc. Together, the vertebrae and intervertebral discs form the vertebral column. It is a flexible column that supports the head, neck, and body and allows for their movements. It also protects the spinal cord, which passes down the back through openings in the vertebrae.
Figure 10.10.1. Vertebral column. The adult vertebral column consists of 24 vertebrae, plus the sacrum and coccyx. The vertebrae are divided into three regions: cervical C1–C7 vertebrae, thoracic T1–T12 vertebrae, and lumbar L1–L5 vertebrae. The vertebral column is curved, with two primary curvatures (thoracic and sacrococcygeal curves) and two secondary curvatures (cervical and lumbar curves).
Regions of the Vertebral Column
The vertebral column originally develops as a series of 33 vertebrae, but this number is eventually reduced to 24 vertebrae, plus the sacrum and coccyx. The vertebral column is subdivided into five regions, with the vertebrae in each area named for that region and numbered in descending order. In the neck, there are seven cervical vertebrae, each designated with the letter “C” followed by its number. Superiorly, the C1 vertebra articulates (forms a joint) with the occipital condyles of the skull. Inferiorly, C1 articulates with the C2 vertebra, and so on. Below these are the 12 thoracic vertebrae, designated T1–T12. The lower back contains the L1–L5 lumbar vertebrae. The single sacrum, which is also part of the pelvis, is formed by the fusion of five sacral vertebrae. Similarly, the coccyx, or tailbone, results from the fusion of four small coccygeal vertebrae. However, the sacral and coccygeal fusions do not start until age 20 and are not completed until middle age.
An interesting anatomical fact is that almost all mammals have seven cervical vertebrae, regardless of body size. This means that there are large variations in the size of cervical vertebrae, ranging from the very small cervical vertebrae of a shrew to the elongated vertebrae in the neck of a giraffe. In a full-grown giraffe, each cervical vertebra is 11 inches tall.
Curvatures of the Vertebral Column
The adult vertebral column does not form a straight line, but instead has four curvatures along its length (see Figure 10.10.1). These curves increase the vertebral column’s strength, flexibility, and ability to absorb shock. When the load on the spine is increased, by carrying a heavy backpack for example, the curvatures increase in depth (become more curved) to accommodate the extra weight. They then spring back when the weight is removed. The four adult curvatures are classified as either primary or secondary curvatures. Primary curves are retained from the original foetal curvature, while secondary curvatures develop after birth.
During foetal development, the body is flexed anteriorly into the foetal position, giving the entire vertebral column a single curvature that is concave anteriorly. In the adult, this foetal curvature is retained in two regions of the vertebral column as the thoracic curve, which involves the thoracic vertebrae, and the sacrococcygeal curve, formed by the sacrum and coccyx. Each of these is thus called a primary curve because they are retained from the original foetal curvature of the vertebral column.
A secondary curve develops gradually after birth as the child learns to sit upright, stand, and walk. Secondary curves are concave posteriorly, opposite in direction to the original foetal curvature. The cervical curve of the neck region develops as the infant begins to hold their head upright when sitting. Later, as the child begins to stand and then to walk, the lumbar curve of the lower back develops. In adults, the lumbar curve is deeper in females.
Disorders associated with the curvature of the spine include kyphosis (an excessive posterior curvature of the thoracic region), lordosis (an excessive anterior curvature of the lumbar region), and scoliosis (an abnormal, lateral curvature, accompanied by twisting of the vertebral column).
Disorders of the Vertebral Column
Developmental anomalies, pathological changes, or obesity can enhance the normal vertebral column curves, resulting in the development of abnormal or excessive curvatures (Figure 10.10.2). Kyphosis, also referred to as humpback or hunchback, is an excessive posterior curvature of the thoracic region. This can develop when osteoporosis causes weakening and erosion of the anterior portions of the upper thoracic vertebrae, resulting in their gradual collapse (Figure 10.10.3). Lordosis, or swayback, is an excessive anterior curvature of the lumbar region and is most associated with obesity or late pregnancy. The accumulation of body weight in the abdominal region results an anterior shift in the line of gravity that carries the weight of the body. This causes in an anterior tilt of the pelvis and a pronounced enhancement of the lumbar curve.
Scoliosis is an abnormal, lateral curvature, accompanied by twisting of the vertebral column. Compensatory curves may also develop in other areas of the vertebral column to help maintain the head positioned over the feet. Scoliosis is the most common vertebral abnormality among girls. The cause is usually unknown, but it may result from weakness of the back muscles, defects such as differential growth rates in the right and left sides of the vertebral column, or differences in the length of the lower limbs. When present, scoliosis tends to get worse during adolescent growth spurts. Although most individuals do not require treatment, a back brace may be recommended for growing children. In extreme cases, surgery may be required.
Excessive vertebral curves can be identified while an individual stands in the anatomical position. Observe the vertebral profile from the side and then from behind to check for kyphosis or lordosis. Then have the person bend forward. If scoliosis is present, an individual will have difficulty in bending directly forward, and the right and left sides of the back will not be level with each other in the bent position.
Figure 10.10.2. Abnormal curvatures of the vertebral column. (a) Scoliosis is an abnormal lateral bending of the vertebral column. (b) An excessive curvature of the upper thoracic vertebral column is called kyphosis. (c) Lordosis is an excessive curvature in the lumbar region of the vertebral column.Figure 10.10.3. Osteoporosis. Osteoporosis is an age-related disorder that causes the gradual loss of bone density and strength. When the thoracic vertebrae are affected, there can be a gradual collapse of the vertebrae. This results in kyphosis, an excessive curvature of the thoracic region.
General Structure of a Vertebra
Within the different regions of the vertebral column, vertebrae vary in size and shape, but they all follow a similar structural pattern. A typical vertebra will consist of a body, a vertebral arch and seven processes (Figure 10.10.4).
The body is the anterior portion of each vertebra and is the part that supports the body weight. Because of this, the vertebral bodies progressively increase in size and thickness going down the vertebral column. The bodies of adjacent vertebrae are separated and strongly united by an intervertebral disc.
The vertebral arch forms the posterior portion of each vertebra. It consists of four parts, the right and left pedicles and the right and left laminae. Each pedicle forms one of the lateral sides of the vertebral arch. The pedicles are anchored to the posterior side of the vertebral body. Each lamina forms part of the posterior roof of the vertebral arch. The large opening between the vertebral arch and body is the vertebral foramen, which contains the spinal cord. In the intact vertebral column, the vertebral foramina of all the vertebrae align to form the vertebral (spinal) canal, which serves as the bony protection and passageway for the spinal cord down the back. When the vertebrae are aligned together in the vertebral column, notches in the margins of the pedicles of adjacent vertebrae together form an intervertebral foramen, the opening through which a spinal nerve exits from the vertebral column (Figure 10.10.5).
Seven processes arise from the vertebral arch. Each paired transverse process projects laterally and arises from the junction point between the pedicle and lamina. The single spinous process (vertebral spine) projects posteriorly at the midline of the back. The vertebral spines can easily be felt as a series of bumps just under the skin down the middle of the back. The transverse and spinous processes serve as important muscle attachment sites. A superior articular process extends or faces upward, and an inferior articular process faces or projects downward on each side of a vertebrae. The paired superior articular processes of one vertebra join with the corresponding paired inferior articular processes from the next higher vertebra. These junctions form slightly moveable joints between the adjacent vertebrae. The shape and orientation of the articular processes vary in different regions of the vertebral column and play a key role in determining the type and range of motion available in each region.
Figure 10.10.4. Parts of a typical vertebra. A typical vertebra consists of a body and a vertebral arch. The arch is formed by the paired pedicles and paired laminae. Arising from the vertebral arch are the transverse, spinous, superior articular, and inferior articular processes. The vertebral foramen provides for passage of the spinal cord. Each spinal nerve exits through an intervertebral foramen, located between adjacent vertebrae. Intervertebral discs unite the bodies of adjacent vertebrae.Figure 10.10.5. Intervertebral disc. The bodies of adjacent vertebrae are separated and united by an intervertebral disc, which provides padding and allows for movements between adjacent vertebrae. The disc consists of a fibrous outer layer called the anulus fibrosus and a gel-like centre called the nucleus pulposus. The intervertebral foramen is the opening formed between adjacent vertebrae for the exit of a spinal nerve.
Regional Modifications of Vertebrae
In addition to the general characteristics of a typical vertebra described above, vertebrae also display characteristic size and structural features that vary between the different vertebral column regions. Thus, cervical vertebrae are smaller than lumbar vertebrae due to differences in the proportion of body weight that each supports. Thoracic vertebrae have sites for rib attachment, and the vertebrae that give rise to the sacrum and coccyx have fused together into single bones.
Cervical Vertebrae
Typical cervical vertebrae, such as C4 or C5, have several characteristic features that differentiate them from thoracic or lumbar vertebrae (Figure 10.10.6). Cervical vertebrae have a small body, reflecting the fact that they carry the least amount of body weight. Cervical vertebrae usually have a bifid (Y-shaped) spinous process. The spinous processes of the C3–C6 vertebrae are short, but the spine of C7 is much longer. You can find these vertebrae by running your finger down the midline of the posterior neck until you encounter the prominent C7 spine located at the base of the neck. The transverse processes of the cervical vertebrae are sharply curved (U-shaped) to allow for passage of the cervical spinal nerves. Each transverse process also has an opening called the transverse foramen. An important artery that supplies the brain ascends up the neck by passing through these openings. The superior and inferior articular processes of the cervical vertebrae are flattened and face upward or downward, respectively.
The first and second cervical vertebrae are further modified, giving each a distinctive appearance. The first cervical (C1) vertebra is also called the atlas, because this is the vertebra that supports the skull on top of the vertebral column (in Greek mythology, Atlas was the god who supported the heavens on his shoulders). The C1 vertebra does not have a body or spinous process. Instead, it is ring-shaped, consisting of an anterior arch and a posterior arch. The transverse processes of the atlas are longer and extend more laterally than do the transverse processes of any other cervical vertebrae. The superior articular processes face upward and are deeply curved for articulation with the occipital condyles on the base of the skull. The inferior articular processes are flat and face downward to join with the superior articular processes of the C2 vertebra.
The second cervical (C2) vertebra is called the axis, because it serves as the axis for rotation when turning the head toward the right or left. The axis resembles typical cervical vertebrae in most respects but is easily distinguished by the dens (odontoid process), a bony projection that extends upward from the vertebral body. The dens joins with the inner aspect of the anterior arch of the atlas, where it is held in place by transverse ligament.
Figure 10.10.6. Cervical vertebrae. A typical cervical vertebra has a small body, a bifid spinous process, transverse processes that have a transverse foramen and are curved for spinal nerve passage. The atlas (C1 vertebra) does not have a body or spinous process. It consists of an anterior and a posterior arch and elongated transverse processes. The axis (C2 vertebra) has the upward projecting dens, which articulates with the anterior arch of the atlas.
Thoracic Vertebrae
The bodies of the thoracic vertebrae are larger than those of cervical vertebrae (Figure 10.10.7). The characteristic feature for a typical midthoracic vertebra is the spinous process, which is long and has a pronounced downward angle that causes it to overlap the next inferior vertebra. The superior articular processes of thoracic vertebrae face anteriorly and the inferior processes face posteriorly. These orientations are important determinants for the type and range of movements available to the thoracic region of the vertebral column.
Thoracic vertebrae have several additional articulation sites, each of which is called a facet, where a rib is attached. Most thoracic vertebrae have two facets located on the lateral sides of the body, each of which is called a costal facet (costal = “rib”). These are for articulation with the head (end) of a rib. An additional facet is located on the transverse process for articulation with the tubercle of a rib.
Figure 10.10.7. Thoracic vertebrae. A typical thoracic vertebra is distinguished by the spinous process, which is long and projects downward to overlap the next inferior vertebra. It also has articulation sites (facets) on the vertebral body and a transverse process for rib attachment.Figure 10.10.8. Rib articulation in thoracic vertebrae. Thoracic vertebrae have superior and inferior articular facets on the vertebral body for articulation with the head of a rib and a transverse process facet for articulation with the rib tubercle.
Lumbar Vertebrae
Lumbar vertebrae carry the greatest amount of body weight and are thus characterised by the large size and thickness of the vertebral body (Figure 10.10.9). They have short transverse processes and a short, blunt spinous process that projects posteriorly. The articular processes are large, with the superior process facing backward and the inferior facing forward.
Figure 10.10.9. Lumbar vertebrae. Lumbar vertebrae are characterised by having a large, thick body and a short, rounded spinous process.
Sacrum and Coccyx
The sacrum is a triangular-shaped bone that is thick and wide across its superior base where it is weight bearing and then tapers down to an inferior, non-weight bearing apex (Figure 10.10.10). It is formed by the fusion of five sacral vertebrae, a process that does not begin until after the age of 20. On the anterior surface of the older adult sacrum, the lines of vertebral fusion can be seen as four transverse ridges. On the posterior surface, running down the midline, is the median sacral crest, a bumpy ridge that is the remnant of the fused spinous processes (median = “midline”; while medial = “toward, but not necessarily at, the midline”). Similarly, the fused transverse processes of the sacral vertebrae form the lateral sacral crest.
The sacral promontory is the anterior lip of the superior base of the sacrum. Lateral to this is the roughened auricular surface, which joins with the ilium portion of the hipbone to form the immobile sacroiliac joints of the pelvis. Passing inferiorly through the sacrum is a bony tunnel called the sacral canal, which terminates at the sacral hiatus near the inferior tip of the sacrum. The anterior and posterior surfaces of the sacrum have a series of paired openings called sacral foramina (singular = foramen) that connect to the sacral canal. Each of these openings is called a posterior (dorsal) sacral foramen or anterior (ventral) sacral foramen. These openings allow for the anterior and posterior branches of the sacral spinal nerves to exit the sacrum. The superior articular process of the sacrum, one of which is found on either side of the superior opening of the sacral canal, articulates with the inferior articular processes from the L5 vertebra.
The coccyx, or tailbone, is derived from the fusion of four very small coccygeal vertebrae (see Figure 10.10.10). It articulates with the inferior tip of the sacrum. It is not weight bearing in the standing position but may receive some body weight when sitting.
Figure 10.10.10. Sacrum and coccyx. The sacrum is formed from the fusion of five sacral vertebrae, whose lines of fusion are indicated by the transverse ridges. The fused spinous processes form the median sacral crest, while the lateral sacral crest arises from the fused transverse processes. The coccyx is formed by the fusion of four small coccygeal vertebrae.
Intervertebral Discs and Ligaments of the Vertebral Column
The bodies of adjacent vertebrae are strongly anchored to each other by an intervertebral disc. This structure provides padding between the bones during weight bearing, and because it can change shape, also allows for movement between the vertebrae. Although the total amount of movement available between any two adjacent vertebrae is small, when these movements are summed together along the entire length of the vertebral column, large body movements can be produced. Ligaments that extend along the length of the vertebral column also contribute to its overall support and stability.
Intervertebral Discs
An intervertebral disc is a fibrocartilaginous pad that fills the gap between adjacent vertebral bodies (see Figure 10.10.5). Each disc is anchored to the bodies of its adjacent vertebrae, thus strongly uniting these. The discs also provide padding between vertebrae during weight bearing. Because of this, intervertebral discs are thin in the cervical region and thickest in the lumbar region, which carries the most body weight. In total, the intervertebral discs account for approximately 25 percent of your body height between the top of the pelvis and the base of the skull. Intervertebral discs are also flexible and can change shape to allow for movements of the vertebral column.
Each intervertebral disc consists of two parts. The anulus fibrosus is the tough, fibrous outer layer of the disc. It forms a circle (anulus = “ring” or “circle”) and is firmly anchored to the outer margins of the adjacent vertebral bodies. Inside is the nucleus pulposus, consisting of a softer, more gel-like material. It has a high water content that serves to resist compression and thus is important for weight bearing. With increasing age, the water content of the nucleus pulposus gradually declines. This causes the disc to become thinner, decreasing total body height somewhat, and reduces the flexibility and range of motion of the disc, making bending more difficult.
The gel-like nature of the nucleus pulposus also allows the intervertebral disc to change shape as one vertebra rocks side to side or forward and back in relation to its neighbours during movements of the vertebral column. Thus, bending forward causes compression of the anterior portion of the disc but expansion of the posterior disc. If the posterior anulus fibrosus is weakened due to injury or increasing age, the pressure exerted on the disc when bending forward and lifting a heavy object can cause the nucleus pulposus to protrude posteriorly through the anulus fibrosus, resulting in a herniated disc (“ruptured” or “slipped” disc) (Figure 10.10.11). The posterior bulging of the nucleus pulposus can cause compression of a spinal nerve at the point where it exits through the intervertebral foramen, with resulting pain and/or muscle weakness in those body regions supplied by that nerve. The most common sites for disc herniation are the L4/L5 or L5/S1 intervertebral discs, which can cause sciatica, a widespread pain that radiates from the lower back down the thigh and into the leg. Similar injuries of the C5/C6 or C6/C7 intervertebral discs, following forcible hyperflexion of the neck from a collision accident or football injury, can produce pain in the neck, shoulder, and upper limb.
Figure 10.10.11. Herniated intervertebral disc. Weakening of the anulus fibrosus can result in herniation (protrusion) of the nucleus pulposus and compression of a spinal nerve, resulting in pain and/or muscle weakness in the body regions supplied by that nerve.
Ligaments of the Vertebral Column
Adjacent vertebrae are united by ligaments that run the length of the vertebral column along both its posterior and anterior aspects (Figure 10.10.12). These serve to resist excess forward or backward bending movements of the vertebral column, respectively.
The anterior longitudinal ligament runs down the anterior side of the entire vertebral column, uniting the vertebral bodies. It serves to resist excess backward bending of the vertebral column. Protection against this movement is particularly important in the neck, where extreme posterior bending of the head and neck can stretch or tear this ligament, resulting in a painful whiplash injury. Prior to the mandatory installation of seat headrests, whiplash injuries were common for passengers involved in a rear-end automobile collision.
The supraspinous ligament is located on the posterior side of the vertebral column, where it interconnects the spinous processes of the thoracic and lumbar vertebrae. This strong ligament supports the vertebral column during forward bending motions. In the posterior neck, where the cervical spinous processes are short, the supraspinous ligament expands to become the nuchal ligament (nuchae = “nape” or “back of the neck”). The nuchal ligament is attached to the cervical spinous processes and extends upward and posteriorly to attach to the midline base of the skull, out to the external occipital protuberance. It supports the skull and prevents it from falling forward. This ligament is much larger and stronger in four-legged animals such as cows, where the large skull hangs off the front end of the vertebral column. You can easily feel this ligament by first extending your head backward and pressing down on the posterior midline of your neck. Then tilt your head forward and you will fill the nuchal ligament popping out as it tightens to limit anterior bending of the head and neck.
Additional ligaments are located inside the vertebral canal, next to the spinal cord, along the length of the vertebral column. The posterior longitudinal ligament is found anterior to the spinal cord, where it is attached to the posterior sides of the vertebral bodies. Posterior to the spinal cord is the ligamentum flavum (“yellow ligament”). This consists of a series of short, paired ligaments, each of which interconnects the lamina regions of adjacent vertebrae. The ligamentum flavum has large numbers of elastic fibres, which have a yellowish colour, allowing it to stretch and then pull back. Both ligaments provide important support for the vertebral column when bending forward.
Figure 10.10.12. Ligaments of vertebral column. The anterior longitudinal ligament runs the length of the vertebral column, uniting the anterior sides of the vertebral bodies. The supraspinous ligament connects the spinous processes of the thoracic and lumbar vertebrae. In the posterior neck, the supraspinous ligament enlarges to form the nuchal ligament, which attaches to the cervical spinous processes and to the base of the skull.
Section Review
The vertebral column forms the neck and back. The vertebral column originally develops as 33 vertebrae, but is eventually reduced to 24 vertebrae, plus the sacrum and coccyx. The vertebrae are divided into the cervical region (C1–C7 vertebrae), the thoracic region (T1–T12 vertebrae), and the lumbar region (L1–L5 vertebrae). The sacrum arises from the fusion of five sacral vertebrae and the coccyx from the fusion of four small coccygeal vertebrae. The vertebral column has four curvatures, the cervical, thoracic, lumbar, and sacrococcygeal curves. The thoracic and sacrococcygeal curves are primary curves retained from the original foetal curvature. The cervical and lumbar curves develop after birth and thus are secondary curves. The cervical curve develops as the infant begins to hold up the head, and the lumbar curve appears with standing and walking.
A typical vertebra consists of an enlarged anterior portion called the body, which provides weight-bearing support. Attached posteriorly to the body is a vertebral arch, which surrounds and defines the vertebral foramen for passage of the spinal cord. The vertebral arch consists of the pedicles, which attach to the vertebral body, and the laminae, which come together to form the roof of the arch. Arising from the vertebral arch are the laterally projecting transverse processes and the posteriorly oriented spinous process. The superior articular processes project upward, where they articulate with the downward projecting inferior articular processes of the next higher vertebrae.
A typical cervical vertebra has a small body, a bifid (Y-shaped) spinous process, and U-shaped transverse processes with a transverse foramen. In addition to these characteristics, the axis (C2 vertebra) also has the dens projecting upward from the vertebral body. The atlas (C1 vertebra) differs from the other cervical vertebrae in that it does not have a body, but instead consists of bony ring formed by the anterior and posterior arches. The atlas articulates with the dens from the axis. A typical thoracic vertebra is distinguished by its long, downward projecting spinous process. Thoracic vertebrae also have articulation facets on the body and transverse processes for attachment of the ribs. Lumbar vertebrae support the greatest amount of body weight and thus have a large, thick body. They also have a short, blunt spinous process. The sacrum is triangular in shape. The median sacral crest is formed by the fused vertebral spinous processes and the lateral sacral crest is derived from the fused transverse processes. Anterior (ventral) and posterior (dorsal) sacral foramina allow branches of the sacral spinal nerves to exit the sacrum. The auricular surfaces are articulation sites on the lateral sacrum that anchor the sacrum to the hipbones to form the pelvis. The coccyx is small and derived from the fusion of four small vertebrae.
The intervertebral discs fill in the gaps between the bodies of adjacent vertebrae. They provide strong attachments and padding between the vertebrae. The outer, fibrous layer of a disc is called the anulus fibrosus. The gel-like interior is called the nucleus pulposus. The disc can change shape to allow for movement between vertebrae. If the anulus fibrosus is weakened or damaged, the nucleus pulposus can protrude outward, resulting in a herniated disc.
The anterior longitudinal ligament runs along the full length of the anterior vertebral column, uniting the vertebral bodies. The supraspinous ligament is located posteriorly and interconnects the spinous processes of the thoracic and lumbar vertebrae. In the neck, this ligament expands to become the nuchal ligament. The nuchal ligament is attached to the cervical spinous processes and superiorly to the base of the skull, out to the external occipital protuberance. The posterior longitudinal ligament runs within the vertebral canal and unites the posterior sides of the vertebral bodies. The ligamentum flavum unites the lamina of adjacent vertebrae.
Discuss the components that make up the thoracic cage
Identify the parts of the sternum and define the sternal angle
Discuss the parts of a rib and rib classifications
The thoracic cage (rib cage) forms the thorax (chest) portion of the body. It consists of the 12 pairs of ribs with their costal cartilages and the sternum (Figure 10.11.1). The ribs are anchored posteriorly to the 12 thoracic vertebrae (T1–T12). The thoracic cage protects the heart and lungs.
Figure 10.11.1. Thoracic cage. The thoracic cage is formed by the (a) sternum and (b) 12 pairs of ribs with their costal cartilages. The ribs are anchored posteriorly to the 12 thoracic vertebrae. The sternum consists of the manubrium, body, and xiphoid process. The ribs are classified as true ribs (1–7) and false ribs (8–12). The last two pairs of false ribs are also known as floating ribs (11–12).
Sternum
The sternum is the elongated bony structure that anchors the anterior thoracic cage. It consists of three parts: the manubrium, body, and xiphoid process. The manubrium is the wider, superior portion of the sternum. The top of the manubrium has a shallow, U-shaped border called the jugular (suprasternal) notch. This can be easily felt at the anterior base of the neck, between the medial ends of the clavicles. The clavicular notch is the shallow depression located on either side at the superior-lateral margins of the manubrium. This is the site of the sternoclavicular joint, between the sternum and clavicle. The first ribs also attach to the manubrium.
The elongated, central portion of the sternum is the body. The manubrium and body join together at the sternal angle, so called because the junction between these two components is not flat, but forms a slight bend. The second rib attaches to the sternum at the sternal angle. Since the first rib is hidden behind the clavicle, the second rib is the highest rib that can be identified by palpation. Thus, the sternal angle and second rib are important landmarks for the identification and counting of the lower ribs. Ribs 3–7 attach to the sternal body.
The inferior tip of the sternum is the xiphoid process. This small structure is cartilaginous early in life, but gradually becomes ossified starting during middle age.
Ribs
Each rib is a curved, flattened bone that contributes to the wall of the thorax. The ribs articulate posteriorly with the T1–T12 thoracic vertebrae, and most attach anteriorly via their costal cartilages to the sternum. There are 12 pairs of ribs. The ribs are numbered 1–12 in accordance with the thoracic vertebrae.
Parts of a Typical Rib
The posterior end of a typical rib is called the head of the rib. This region articulates primarily with the costal facet located on the body of the same numbered thoracic vertebra and to a lesser degree, with the costal facet located on the body of the next higher vertebra. Lateral to the head is the narrowed neck of the rib. A small bump on the posterior rib surface is the tubercle of the rib, which articulates with the facet located on the transverse process of the same numbered vertebra. The remainder of the rib is the body of the rib (shaft). Just lateral to the tubercle is the angle of the rib, the point at which the rib has its greatest degree of curvature. The angles of the ribs form the most posterior extent of the thoracic cage. In the anatomical position, the angles align with the medial border of the scapula. A shallow costal groove for the passage of blood vessels and a nerve is found along the inferior margin of each rib.
Rib Classifications
The bony ribs do not extend anteriorly completely around to the sternum. Instead, each rib ends in a costal cartilage. These cartilages are made of hyaline cartilage and can extend for several inches. Most ribs are then attached, either directly or indirectly, to the sternum via their costal cartilage (see Figure 10.11.1). The ribs are classified into three groups based on their relationship to the sternum.
Ribs 1–7 are classified as true ribs (vertebrosternal ribs). The costal cartilage from each of these ribs attaches directly to the sternum. Ribs 8–12 are called false ribs (vertebrochondral ribs). The costal cartilages from these ribs do not attach directly to the sternum. For ribs 8–10, the costal cartilages are attached to the cartilage of the next higher rib. Thus, the cartilage of rib 10 attaches to the cartilage of rib 9, rib 9 then attaches to rib 8, and rib 8 is attached to rib 7. The last two false ribs (11–12) are also called floating ribs (vertebral ribs). These are short ribs that do not attach to the sternum at all. Instead, their small costal cartilages terminate within the musculature of the lateral abdominal wall.
Section Review
The thoracic cage protects the heart and lungs. It is composed of 12 pairs of ribs with their costal cartilages and the sternum. The ribs are anchored posteriorly to the 12 thoracic vertebrae. The sternum consists of the manubrium, body, and xiphoid process. The manubrium and body are joined at the sternal angle, which is also the site for attachment of the second ribs.
Ribs are flattened, curved bones and are numbered 1–12. Posteriorly, the head of the rib articulates with the costal facets located on the bodies of thoracic vertebrae and the rib tubercle articulates with the facet located on the vertebral transverse process. The angle of the ribs forms the most posterior portion of the thoracic cage. The costal groove in the inferior margin of each rib carries blood vessels and a nerve. Anteriorly, each rib ends in a costal cartilage. True ribs (1–7) attach directly to the sternum via their costal cartilage. The false ribs (8–12) either attach to the sternum indirectly or not at all. Ribs 8–10 have their costal cartilages attached to the cartilage of the next higher rib. The floating ribs (11–12) are short and do not attach to the sternum or to another rib.
Discuss the two types of embryonic bone development within the skull
Describe the development of the vertebral column and thoracic cage
The axial skeleton begins to form during early embryonic development. However, growth, remodelling, and ossification (bone formation) continue for several decades after birth before the adult skeleton is fully formed. Knowledge of the developmental processes that give rise to the skeleton is important for understanding the abnormalities that may arise in skeletal structures.
Development of The Skull
During the third week of embryonic development, a rod-like structure called the notochord develops dorsally along the length of the embryo. The tissue overlying the notochord enlarges and forms the neural tube, which will give rise to the brain and spinal cord. By the fourth week, mesoderm tissue located on either side of the notochord thickens and separates into a repeating series of block-like tissue structures, each of which is called a somite. As the somites enlarge, each one will split into several parts. The most medial of these parts is called a sclerotome. The sclerotomes consist of an embryonic tissue called mesenchyme, which will give rise to the fibrous connective tissues, cartilages, and bones of the body.
The bones of the skull arise from mesenchyme during embryonic development in two different ways. The first mechanism produces the bones that form the top and sides of the brain case. This involves the local accumulation of mesenchymal cells at the site of the future bone. These cells then differentiate directly into bone producing cells, which form the skull bones through the process of intramembranous ossification. As the brain case bones grow in the foetal skull, they remain separated from each other by large areas of dense connective tissue, each of which is called a fontanelle (Figure 10.12.1). The fontanelles are the soft spots on an infant’s head. They are important during birth because these areas allow the skull to change shape as it squeezes through the birth canal. After birth, the fontanelles allow for continued growth and expansion of the skull as the brain enlarges. The largest fontanelle is located on the anterior head, at the junction of the frontal and parietal bones. The fontanelles decrease in size and disappear by age two. However, the skull bones remained separated from each other at the sutures, which contain dense fibrous connective tissue that unites the adjacent bones. The connective tissue of the sutures allows for continued growth of the skull bones as the brain enlarges during childhood growth.
The second mechanism for bone development in the skull produces the facial bones and floor of the brain case. This also begins with the localised accumulation of mesenchymal cells. However, these cells differentiate into cartilage cells, which produce a hyaline cartilage model of the future bone. As this cartilage model grows, it is gradually converted into bone through the process of endochondral ossification. This is a slow process and the cartilage is not completely converted to bone until the skull achieves its full adult size.
At birth, the brain case and orbits of the skull are disproportionally large compared to the bones of the jaws and lower face. This reflects the relative underdevelopment of the maxilla and mandible, which lack teeth, and the small sizes of the paranasal sinuses and nasal cavity. During early childhood, the mastoid process enlarges, the two halves of the mandible and frontal bone fuse together to form single bones, and the paranasal sinuses enlarge. The jaws also expand as the teeth begin to appear. These changes all contribute to the rapid growth and enlargement of the face during childhood.
Figure 10.12.1. Newborn skull. The bones of the newborn skull are not fully ossified and are separated by large areas called fontanelles, which are filled with fibrous connective tissue. The fontanelles allow for continued growth of the skull after birth. At the time of birth, the facial bones are small and underdeveloped, and the mastoid process has not yet formed.
Development of the Vertebral Column and Thoracic Cage
Development of the vertebrae begins with the accumulation of mesenchyme cells from each sclerotome around the notochord. These cells differentiate into a hyaline cartilage model for each vertebra, which then grow and eventually ossify into bone through the process of endochondral ossification. As the developing vertebrae grow, the notochord disappears. However, small areas of notochord tissue persist between the adjacent vertebrae and this contributes to the formation of each intervertebral disc.
The ribs and sternum also develop from mesenchyme. The ribs initially develop as part of the cartilage model for each vertebra, but in the thorax region, the rib portion separates from the vertebra by the eighth week. The cartilage model of the rib then ossifies, except for the anterior portion, which remains as the costal cartilage. The sternum initially forms as paired hyaline cartilage models on either side of the anterior midline, beginning during the fifth week of development. The cartilage models of the ribs become attached to the lateral sides of the developing sternum. Eventually, the two halves of the cartilaginous sternum fuse together along the midline and then ossify into bone. The manubrium and body of the sternum are converted into bone first, with the xiphoid process remaining as cartilage until late in life.
Homeostatic Imbalances
Craniosynostosis
The premature closure (fusion) of a suture line is a condition called craniosynostosis. This error in the normal developmental process results in abnormal growth of the skull and deformity of the head. It is produced either by defects in the ossification process of the skull bones or failure of the brain to properly enlarge. Genetic factors are involved, but the underlying cause is unknown. It is a common condition, occurring in approximately 1:3000 births in Australia, with males being more commonly affected. Primary craniosynostosis involves the early fusion of one cranial suture, whereas complex craniosynostosis results from the premature fusion of several sutures.
The early fusion of a suture in primary craniosynostosis prevents any additional enlargement of the cranial bones and skull along this line. Continued growth of the brain and skull is therefore diverted to other areas of the head, causing an abnormal enlargement of these regions. For example, the early disappearance of the anterior fontanelle and premature closure of the sagittal suture prevents growth across the top of the head. This is compensated by upward growth by the bones of the lateral skull, resulting in a long, narrow, wedge-shaped head. This condition, known as scaphocephaly, accounts for approximately 50 percent of craniosynostosis abnormalities. Although the skull is misshapen, the brain still has adequate room to grow and thus there is no accompanying abnormal neurological development.
In cases of complex craniosynostosis, several sutures close prematurely. The amount and degree of skull deformity is determined by the location and extent of the sutures involved. This results in more severe constraints on skull growth, which can alter or impede proper brain growth and development.
Cases of craniosynostosis are usually treated with surgery. A team of physicians will open the skull along the fused suture, which will then allow the skull bones to resume their growth in this area. In some cases, parts of the skull will be removed and replaced with an artificial plate. The earlier after birth that surgery is performed, the better the outcome. After treatment, most children continue to grow and develop normally and do not exhibit any neurological problems.
Section Review
Formation of the axial skeleton begins during early embryonic development with the appearance of the rod-like notochord along the dorsal length of the early embryo. Repeating, paired blocks of tissue called somites then appear along either side of notochord. As the somites grow, they split into parts, one of which is called a sclerotome. This consists of mesenchyme, the embryonic tissue that will become the bones, cartilages, and connective tissues of the body.
Mesenchyme in the head region will produce the bones of the skull via two different mechanisms. The bones of the brain case arise via intramembranous ossification in which embryonic mesenchyme tissue converts directly into bone. At the time of birth, these bones are separated by fontanelles, wide areas of fibrous connective tissue. As the bones grow, the fontanelles are reduced to sutures, which allow for continued growth of the skull throughout childhood. In contrast, the cranial base and facial bones are produced by the process of endochondral ossification, in which mesenchyme tissue initially produces a hyaline cartilage model of the future bone. The cartilage model allows for growth of the bone and is gradually converted into bone over a period of many years.
The vertebrae, ribs, and sternum also develop via endochondral ossification. Mesenchyme accumulates around the notochord and produces hyaline cartilage models of the vertebrae. The notochord disappears, but remnants of the notochord contribute to formation of the intervertebral discs. In the thorax region, a portion of the vertebral cartilage model splits off to form the ribs. These then become attached anteriorly to the developing cartilage model of the sternum. Growth of the cartilage models for the vertebrae, ribs, and sternum allow for enlargement of the thoracic cage during childhood and adolescence. The cartilage models gradually undergo ossification and are converted into bone.
The appendicular skeleton includes all the limb bones, plus the bones that unite each limb with the axial skeleton (Figure 11.1.1). The bones that attach each upper limb to the axial skeleton form the pectoral girdle (shoulder girdle). This consists of two bones, the scapula and clavicle (Figure 11.1.1). The clavicle (collarbone) is an S-shaped bone located on the anterior side of the shoulder. It is attached on its medial end to the sternum of the thoracic cage, which is part of the axial skeleton. The lateral end of the clavicle articulates (joins) with the scapula just above the shoulder joint. You can easily palpate, or feel with your fingers, the entire length of your clavicle.
Figure 11.1.1. Axial and appendicular skeletons. The axial skeleton forms the central axis of the body and consists of the skull, vertebral column, and thoracic cage. The appendicular skeleton consists of the pectoral and pelvic girdles, the limb bones, and the bones of the hands and feet.Figure 11.1.2. Pectoral girdle. The pectoral girdle consists of the clavicle and the scapula, which serve to attach the upper limb to the sternum of the axial skeleton.
The scapula (shoulder blade) lies on the posterior aspect of the shoulder. It is supported by the clavicle, which also articulates with the humerus (arm bone) to form the shoulder joint. The scapula is a flat, triangular-shaped bone with a prominent ridge running across its posterior surface. This ridge extends out laterally, where it forms the bony tip of the shoulder and joins with the lateral end of the clavicle. By following along the clavicle, you can palpate out to the bony tip of the shoulder, and from there, you can move back across your posterior shoulder to follow the ridge of the scapula. Move your shoulder around and feel how the clavicle and scapula move together as a unit. Both of these bones serve as important attachment sites for muscles that aid with movements of the shoulder and arm.
The right and left pectoral girdles are not joined to each other, allowing each to operate independently. In addition, the clavicle of each pectoral girdle is anchored to the axial skeleton by a single, highly mobile joint. This allows for the extensive mobility of the entire pectoral girdle, which in turn enhances movements of the shoulder and upper limb.
Clavicle
The clavicle is the only long bone that lies in a horizontal position in the body (see Figure 11.1.2). The clavicle has several important functions. First, anchored by muscles from above, it serves as a strut that extends laterally to support the scapula. This in turn holds the shoulder joint superiorly and laterally from the body trunk, allowing for maximal freedom of motion for the upper limb. The clavicle also transmits forces acting on the upper limb to the sternum and axial skeleton. Finally, it serves to protect the underlying nerves and blood vessels as they pass between the trunk of the body and the upper limb.
The clavicle has three regions: the medial end, the lateral end, and the shaft. The medial end, known as the sternal end of the clavicle, has a triangular shape and articulates with the manubrium portion of the sternum. This forms the sternoclavicular joint, which is the only bony articulation between the pectoral girdle of the upper limb and the axial skeleton. This joint allows considerable mobility, enabling the clavicle and scapula to move in upward/downward and anterior/posterior directions during shoulder movements. The sternoclavicular joint is indirectly supported by the costoclavicular ligament (costo- = “rib”), which spans the sternal end of the clavicle and the underlying first rib. The lateral or acromial end of the clavicle articulates with the acromion of the scapula, the portion of the scapula that forms the bony tip of the shoulder. There are some sex differences in the morphology of the clavicle. In women, the clavicle tends to be shorter, thinner, and less curved. In men, the clavicle is heavier and longer, and has a greater curvature and rougher surfaces where muscles attach, features that are more pronounced in manual workers.
The clavicle is the most commonly fractured bone in the body. Such breaks often occur because of the force exerted on the clavicle when a person falls onto his or her outstretched arms, or when the lateral shoulder receives a strong blow. Because the sternoclavicular joint is strong and rarely dislocated, excessive force results in the breaking of the clavicle, usually between the middle and lateral portions of the bone. If the fracture is complete, the shoulder and lateral clavicle fragment will drop due to the weight of the upper limb, causing the person to support the sagging limb with their other hand. Muscles acting across the shoulder will also pull the shoulder and lateral clavicle anteriorly and medially, causing the clavicle fragments to override. The clavicle overlies many important blood vessels and nerves for the upper limb, but fortunately, due to the anterior displacement of a broken clavicle, these structures are rarely affected when the clavicle is fractured.
Scapula
The scapula is also part of the pectoral girdle and thus plays an important role in anchoring the upper limb to the body. The scapula is located on the posterior side of the shoulder. It is surrounded by muscles on both its anterior (deep) and posterior (superficial) sides, and thus does not articulate with the ribs of the thoracic cage.
The scapula has several important landmarks (Figure 11.1.3). The three margins or borders of the scapula, named for their positions within the body, are the superior border of the scapula, the medial border of the scapula, and the lateral border of the scapula. The suprascapular notch is located lateral to the midpoint of the superior border. The corners of the triangular scapula, at either end of the medial border, are the superior angle of the scapula, located between the medial and superior borders, and the inferior angle of the scapula, located between the medial and lateral borders. The inferior angle is the most inferior portion of the scapula and is particularly important because it serves as the attachment point for several powerful muscles involved in shoulder and upper limb movements. The remaining corner of the scapula, between the superior and lateral borders, is the location of the glenoid cavity (glenoid fossa). This shallow depression articulates with the humerus bone of the arm to form the glenohumeral joint (shoulder joint). The small bony bumps located immediately above and below the glenoid cavity are the supraglenoid tubercle and the infraglenoid tubercle, respectively. These provide attachments for muscles of the arm.
Figure 11.1.3. Scapula. The isolated scapula is shown here from its anterior (deep) side and its posterior (superficial) side.
The scapula also has two prominent projections. Toward the lateral end of the superior border, between the suprascapular notch and glenoid cavity, is the hook-like coracoid process (coracoid = “shaped like a crow’s beak”). This process projects anteriorly and curves laterally. At the shoulder, the coracoid process is located inferior to the lateral end of the clavicle. It is anchored to the clavicle by a strong ligament and serves as the attachment site for muscles of the anterior chest and arm. On the posterior aspect, the spine of the scapula is a long and prominent ridge that runs across its upper portion. Extending laterally from the spine is a flattened and expanded region called the acromion or acromial process. The acromion forms the bony tip of the superior shoulder region and articulates with the lateral end of the clavicle, forming the acromioclavicular joint (see Figure 11.1.2). Together, the clavicle, acromion, and spine of the scapula form a V-shaped bony line that provides for the attachment of neck and back muscles that act on the shoulder, as well as muscles that pass across the shoulder joint to act on the arm.
The scapula has three depressions, each of which is called a fossa (plural = fossae). Two of these are found on the posterior scapula, above and below the scapular spine. Superior to the spine is the narrow supraspinous fossa, and inferior to the spine is the broad infraspinous fossa. The anterior (deep) surface of the scapula forms the broad subscapular fossa. All these fossae provide large surface areas for the attachment of muscles that cross the shoulder joint to act on the humerus.
The acromioclavicular joint transmits forces from the upper limb to the clavicle. The ligaments around this joint are relatively weak. A hard fall onto the elbow or outstretched hand can stretch or tear the acromioclavicular ligaments, resulting in a moderate injury to the joint. However, the primary support for the acromioclavicular joint comes from a very strong ligament called the coracoclavicular ligament (see Figure 11.1.2). This connective tissue band anchors the coracoid process of the scapula to the inferior surface of the acromial end of the clavicle and thus provides important indirect support for the acromioclavicular joint. Following a strong blow to the lateral shoulder, such as when a hockey player is driven into the boards, a complete dislocation of the acromioclavicular joint can result. In this case, the acromion is thrust under the acromial end of the clavicle, resulting in ruptures of both the acromioclavicular and coracoclavicular ligaments. The scapula then separates from the clavicle, with the weight of the upper limb pulling the shoulder downward. This dislocation injury of the acromioclavicular joint is known as a “shoulder separation” and is common in contact sports such as hockey, football, or martial arts.
Section Review
The pectoral girdle, consisting of the clavicle and the scapula, attaches each upper limb to the axial skeleton. The clavicle is an anterior bone whose sternal end articulates with the manubrium of the sternum at the sternoclavicular joint. The sternal end is also anchored to the first rib by the costoclavicular ligament. The acromial end of the clavicle articulates with the acromion of the scapula at the acromioclavicular joint. This end is also anchored to the coracoid process of the scapula by the coracoclavicular ligament, which provides indirect support for the acromioclavicular joint. The clavicle supports the scapula, transmits the weight and forces from the upper limb to the body trunk, and protects the underlying nerves and blood vessels.
The scapula lies on the posterior aspect of the pectoral girdle. It mediates the attachment of the upper limb to the clavicle and contributes to the formation of the glenohumeral (shoulder) joint. This triangular bone has three sides called the medial, lateral, and superior borders. The suprascapular notch is located on the superior border. The scapula also has three corners, two of which are the superior and inferior angles. The third corner is occupied by the glenoid cavity. Posteriorly, the spine separates the supraspinous and infraspinous fossae, and then extends laterally as the acromion. The subscapular fossa is located on the anterior surface of the scapula. The coracoid process projects anteriorly, passing inferior to the lateral end of the clavicle.
Identify the divisions of the upper limb and describe the bones in each region
List the bones and bony landmarks that articulate at each joint of the upper limb
The upper limb is divided into three regions. These consist of the arm, located between the shoulder and elbow joints; the forearm, which is between the elbow and wrist joints; and the hand, which is located distal to the wrist. There are 30 bones in each upper limb. The humerus is the single bone of the upper arm, and the ulna (medially) and the radius (laterally) are the paired bones of the forearm. The base of the hand contains eight bones, each called a carpal bone, and the palm of the hand is formed by five bones, each called a metacarpal bone. The fingers and thumb contain a total of 14 bones, each of which is a phalanx bone of the hand.
Humerus
The humerus is the single bone of the upper arm region (Figure 11.2.1). At its proximal end is the head of the humerus. This is the large, round, smooth region that faces medially. The head articulates with the glenoid cavity of the scapula to form the glenohumeral (shoulder) joint. The margin of the smooth area of the head is the anatomical neck of the humerus. Located on the lateral side of the proximal humerus is an expanded bony area called the greater tubercle. The smaller lesser tubercle of the humerus is found on the anterior aspect of the humerus. Both the greater and lesser tubercles serve as attachment sites for muscles that act across the shoulder joint. Passing between the greater and lesser tubercles is the narrow intertubercular groove (sulcus), which is also known as the bicipital groove because it provides passage for a tendon of the biceps brachii muscle. The surgical neck is located at the base of the expanded, proximal end of the humerus, where it joins the narrow shaft of the humerus. The surgical neck is a common site of arm fractures. The deltoid tuberosity is a roughened, V-shaped region located on the lateral side in the middle of the humerus shaft. As its name indicates, it is the site of attachment for the deltoid muscle.
Figure 11.2.1. Humorous and elbow joint. The humerus is the single bone of the upper arm region. It articulates with the radius and ulna bones of the forearm to form the elbow joint.
Distally, the humerus becomes flattened. The prominent bony projection on the medial side is the medial epicondyle of the humerus. The much smaller lateral epicondyle of the humerus is found on the lateral side of the distal humerus. The roughened ridge of bone above the lateral epicondyle is the lateral supracondylar ridge. All these areas are attachment points for muscles that act on the forearm, wrist, and hand. The powerful grasping muscles of the anterior forearm arise from the medial epicondyle, which is thus larger and more robust than the lateral epicondyle that gives rise to the weaker posterior forearm muscles.
The distal end of the humerus has two articulation areas, which join the ulna and radius bones of the forearm to form the elbow joint. The more medial of these areas is the trochlea, a spindle- or pulley-shaped region (trochlea = “pulley”), which articulates with the ulna bone. Immediately lateral to the trochlea is the capitulum (“small head”), a knob-like structure located on the anterior surface of the distal humerus. The capitulum articulates with the radius bone of the forearm. Just above these bony areas are two small depressions. These spaces accommodate the forearm bones when the elbow is fully bent (flexed). Superior to the trochlea is the coronoid fossa, which receives the coronoid process of the ulna, and above the capitulum is the radial fossa (anatomical snuff box), which receives the head of the radius when the elbow is flexed. Similarly, the posterior humerus has the olecranon fossa, a larger depression that receives the olecranon process of the ulna when the forearm is fully extended.
Ulna
The ulna is the medial bone of the forearm. It runs parallel to the radius, which is the lateral bone of the forearm (Figure 11.2.2). The proximal end of the ulna resembles a crescent wrench with its large, C-shaped trochlear notch. This region articulates with the trochlea of the humerus as part of the elbow joint. The inferior margin of the trochlear notch is formed by a prominent lip of bone called the coronoid process of the ulna. Just below this on the anterior ulna is a roughened area called the ulnar tuberosity. To the lateral side and slightly inferior to the trochlear notch is a small, smooth area called the radial notch of the ulna. This area is the site of articulation between the proximal radius and the ulna, forming the proximal radioulnar joint. The posterior and superior portions of the proximal ulna make up the olecranon process, which forms the bony tip of the elbow.
Figure 11.2.2. Ulna and radius. The ulna is located on the medial side of the forearm, and the radius is on the lateral side. These bones are attached to each other by an interosseous membrane.
More distal is the shaft of the ulna. The lateral side of the shaft forms a ridge called the interosseous border of the ulna. This is the line of attachment for the interosseous membrane of the forearm, a sheet of dense connective tissue that unites the ulna and radius bones. The small, rounded area that forms the distal end is the head of the ulna. Projecting from the posterior side of the ulnar head is the styloid process of the ulna, a short bony projection. This serves as an attachment point for a connective tissue structure that unites the distal ends of the ulna and radius.
In the anatomical position, with the elbow fully extended and the palms facing forward, the arm and forearm do not form a straight line. Instead, the forearm deviates laterally by 5–15 degrees from the line of the arm. This deviation is called the carrying angle. It allows the forearm and hand to swing freely or to carry an object without hitting the hip. The carrying angle is larger in females to accommodate their wider pelvis.
Radius
The radius runs parallel to the ulna, on the lateral (thumb) side of the forearm (see Figure 11.2.2). The head of the radius is a disc-shaped structure that forms the proximal end. The small depression on the surface of the head articulates with the capitulum of the humerus as part of the elbow joint, whereas the smooth, outer margin of the head articulates with the radial notch of the ulna at the proximal radioulnar joint. The neck of the radius is the narrowed region immediately below the expanded head. Inferior to this point on the medial side is the radial tuberosity, an oval-shaped, bony protuberance that serves as a muscle attachment point. The shaft of the radius is slightly curved and has a small ridge along its medial side. This ridge forms the interosseous border of the radius, which, like the similar border of the ulna, is the line of attachment for the interosseous membrane that unites the two forearm bones. The distal end of the radius has a smooth surface for articulation with two carpal bones to form the radiocarpal joint or wrist joint (Figure 11.2.3 and Figure 11.2.4). On the medial side of the distal radius is the ulnar notch of the radius. This shallow depression articulates with the head of the ulna, which together form the distal radioulnar joint. The lateral end of the radius has a pointed projection called the styloid process of the radius. This provides attachment for ligaments that support the lateral side of the wrist joint. Compared to the styloid process of the ulna, the styloid process of the radius projects more distally, thereby limiting the range of movement for lateral deviations of the hand at the wrist joint.
Carpal Bones
The wrist and base of the hand are formed by a series of eight small carpal bones (see Figure 11.2.3). The carpal bones are arranged in two rows, forming a proximal row of four carpal bones and a distal row of four carpal bones. The bones in the proximal row, running from the lateral (thumb) side to the medial side, are the scaphoid (“boat-shaped”), lunate (“moon-shaped”), triquetrum (“three-cornered”), and pisiform (“pea-shaped”) bones. The small, rounded pisiform bone articulates with the anterior surface of the triquetrum bone. The pisiform thus projects anteriorly, where it forms the bony bump that can be felt at the medial base of your hand. The distal bones (lateral to medial) are the trapezium (“table”), trapezoid (“resembles a table”), capitate (“head-shaped”), and hamate (“hooked bone”) bones. The hamate bone is characterised by a prominent bony extension on its anterior side called the hook of the hamate bone.
A helpful mnemonic for remembering the arrangement of the carpal bones is “So Long To Pinky, Here Comes The Thumb.” This mnemonic starts on the lateral side and names the proximal bones from lateral to medial (scaphoid, lunate, triquetrum, pisiform), then makes a U-turn to name the distal bones from medial to lateral (hamate, capitate, trapezoid, trapezium). Thus, it starts and finishes on the lateral side.
Figure 11.2.3. Bones of the wrist and hand. The eight carpal bones form the base of the hand. These are arranged into proximal and distal rows of four bones each. The metacarpal bones form the palm of the hand. The thumb and fingers consist of the phalanx bones.
The carpal bones form the base of the hand. This can be seen in the radiograph (X-ray image) of the hand that shows the relationships of the hand bones to the skin creases of the hand (see Figure 11.2.4). Within the carpal bones, the four proximal bones are united to each other by ligaments to form a unit. Only three of these bones, the scaphoid, lunate, and triquetrum, contribute to the radiocarpal joint. The scaphoid and lunate bones articulate directly with the distal end of the radius, whereas the triquetrum bone articulates with a fibrocartilaginous pad that spans the radius and styloid process of the ulna. The distal end of the ulna thus does not directly articulate with any of the carpal bones.
The four distal carpal bones are also held together as a group by ligaments. The proximal and distal rows of carpal bones articulate with each other to form the midcarpal joint (see Figure 11.2.4). Together, the radiocarpal and midcarpal joints are responsible for all movements of the hand at the wrist. The distal carpal bones also articulate with the metacarpal bones of the hand.
Figure 11.2.4. Bones of the hand posterior view. This radiograph shows the position of the bones within the hand. Note the carpal bones that form the base of the hand. (credit: modification of work by Trace Meek).
In the articulated hand, the carpal bones form a U-shaped grouping. A strong ligament called the flexor retinaculum spans the top of this U-shaped area to maintain this grouping of the carpal bones. The flexor retinaculum is attached laterally to the trapezium and scaphoid bones, and medially to the hamate and pisiform bones. Together, the carpal bones and the flexor retinaculum form a passageway called the carpal tunnel, with the carpal bones forming the walls and floor, and the flexor retinaculum forming the roof of this space. The tendons of nine muscles of the anterior forearm and an important nerve pass through this narrow tunnel to enter the hand. Overuse of the muscle tendons or wrist injury can produce inflammation and swelling within this space. This produces compression of the nerve, resulting in carpal tunnel syndrome, which is characterised by pain or numbness, and muscle weakness in those areas of the hand supplied by this nerve.
Metacarpal Bones
The palm of the hand contains five elongated metacarpal bones. These bones lie between the carpal bones of the wrist and the bones of the fingers and thumb (see Figure 11.2.3). The proximal end of each metacarpal bone articulates with one of the distal carpal bones. Each of these articulations is a carpometacarpal joint (see Figure 11.2.4). The expanded distal end of each metacarpal bone articulates at the metacarpophalangeal joint with the proximal phalanx bone of the thumb or one of the fingers. The distal end also forms the knuckles of the hand, at the base of the fingers. The metacarpal bones are numbered 1–5, beginning at the thumb.
The first metacarpal bone, at the base of the thumb, is separated from the other metacarpal bones. This allows it a freedom of motion that is independent of the other metacarpal bones, which is very important for thumb mobility. The remaining metacarpal bones are united together to form the palm of the hand. The second and third metacarpal bones are firmly anchored in place and are immobile. However, the fourth and fifth metacarpal bones have limited anterior-posterior mobility, a motion that is greater for the fifth bone. This mobility is important during power gripping with the hand (Figure 11.2.5). The anterior movement of these bones, particularly the fifth metacarpal bone, increases the strength of contact for the medial hand during gripping actions.
Figure 11.2.5. Hand during gripping. During tight gripping—compare (b) to (a)—the fourth and, particularly, the fifth metatarsal bones are pulled anteriorly. This increases the contact between the object and the medial side of the hand, thus improving the firmness of the grip.
Phalanx Bones
The fingers and thumb contain 14 bones, each of which is called a phalanx bone (plural = phalanges), named after the ancient Greek phalanx (a rectangular block of soldiers). The thumb (pollex) is digit number 1 and has two phalanges, a proximal phalanx, and a distal phalanx bone (see Figure 11.2.3). Digits 2 (index finger) through 5 (little finger) have three phalanges each, called the proximal, middle, and distal phalanx bones. An interphalangeal joint is one of the articulations between adjacent phalanges of the digits (see Figure 11.2.4).
Disorders of the Appendicular System: Fractures of Upper Limb Bones
Due to our constant use of the hands and the rest of our upper limbs, an injury to any of these areas will cause a significant loss of functional ability. Many fractures result from a hard fall onto an outstretched hand. The resulting transmission of force up the limb may result in a fracture of the humerus, radius, or scaphoid bones. These injuries are especially common in elderly people whose bones are weakened due to osteoporosis.
Falls onto the hand or elbow, or direct blows to the arm, can result in fractures of the humerus (Figure 11.2.6). Following a fall, fractures at the surgical neck, the region at which the expanded proximal end of the humerus joins with the shaft, can result in an impacted fracture, in which the distal portion of the humerus is driven into the proximal portion. Falls or blows to the arm can also produce transverse or spiral fractures of the humeral shaft.
In children, a fall onto the tip of the elbow frequently results in a distal humerus fracture. In these, the olecranon of the ulna is driven upward, resulting in a fracture across the distal humerus, above both epicondyles (supracondylar fracture), or a fracture between the epicondyles, thus separating one or both epicondyles from the body of the humerus (intercondylar fracture). With these injuries, the immediate concern is compression of the artery to the forearm due to swelling of the surrounding tissues. If compression occurs, the resulting ischemia (lack of oxygen) due to reduced blood flow can quickly produce irreparable damage to the forearm muscles. In addition, four major nerves for shoulder and upper limb muscles are closely associated with different regions of the humerus, and thus, humeral fractures may also damage these nerves.
Another frequent injury following a fall onto an outstretched hand is a Colles fracture (“col-lees”) of the distal radius (see Figure 11.2.6). This involves a complete transverse fracture across the distal radius that drives the separated distal fragment of the radius posteriorly and superiorly. This injury results in a characteristic “dinner fork” bend of the forearm just above the wrist due to the posterior displacement of the hand. This is the most frequent forearm fracture and is a common injury in persons over the age of 50, particularly in older women with osteoporosis. It also commonly occurs following a high-speed fall onto the hand during activities such as snowboarding or skating.
The most commonly fractured carpal bone is the scaphoid, often resulting from a fall onto the hand. Deep pain at the lateral wrist may yield an initial diagnosis of a wrist sprain, but a radiograph taken several weeks after the injury, after tissue swelling has subsided, will reveal the fracture. Due to the poor blood supply to the scaphoid bone, healing will be slow and there is the danger of bone necrosis and subsequent degenerative joint disease of the wrist.
Figure 11.2.6. Fractures of the humerus and radius. Falls or direct blows can result in fractures of the surgical neck or shaft of the humerus. Falls onto the elbow can fracture the distal humerus. A Colles fracture of the distal radius is the most common forearm fracture.
Section Review
Each upper limb is divided into three regions and contains a total of 30 bones. The upper arm is the region located between the shoulder and elbow joints. This area contains the humerus. The proximal humerus consists of the head, which articulates with the scapula at the glenohumeral joint, the greater and lesser tubercles separated by the intertubercular (bicipital) groove, and the anatomical and surgical necks. The humeral shaft has the roughened area of the deltoid tuberosity on its lateral side. The distal humerus is flattened, forming a lateral supracondylar ridge that terminates at the small lateral epicondyle. The medial side of the distal humerus has the large, medial epicondyle. The articulating surfaces of the distal humerus consist of the trochlea medially and the capitulum laterally. Depressions on the humerus that accommodate the forearm bones during bending (flexing) and straightening (extending) of the elbow include the coronoid fossa, the radial fossa, and the olecranon fossa.
The forearm is the region of the upper limb located between the elbow and wrist joints. This region contains two bones, the ulna medially and the radius on the lateral (thumb) side. The elbow joint is formed by the articulation between the trochlea of the humerus and the trochlear notch of the ulna, plus the articulation between the capitulum of the humerus and the head of the radius. The proximal radioulnar joint is the articulation between the head of the radius and the radial notch of the ulna. The proximal ulna also has the olecranon process, forming an expanded posterior region, and the coronoid process and ulnar tuberosity on its anterior aspect. On the proximal radius, the narrowed region below the head is the neck; distal to this is the radial tuberosity. The shaft portions of both the ulna and radius have an interosseous border, whereas the distal ends of each bone have a pointed styloid process. The distal radioulnar joint is found between the head of the ulna and the ulnar notch of the radius. The distal end of the radius articulates with the proximal carpal bones, but the ulna does not.
The base of the hand is formed by eight carpal bones. The carpal bones are united into two rows of bones. The proximal row contains (from lateral to medial) the scaphoid, lunate, triquetrum, and pisiform bones. The scaphoid, lunate, and triquetrum bones contribute to the formation of the radiocarpal joint. The distal row of carpal bones contains (from medial to lateral) the hamate, capitate, trapezoid, and trapezium bones (“So Long To Pinky, Here Comes The Thumb”). The anterior hamate has a prominent bony hook. The proximal and distal carpal rows articulate with each other at the midcarpal joint. The carpal bones, together with the flexor retinaculum, also form the carpal tunnel of the wrist.
The five metacarpal bones form the palm of the hand. The metacarpal bones are numbered 1–5, starting with the thumb side. The first metacarpal bone is freely mobile, but the other bones are united as a group. The digits are also numbered 1–5, with the thumb being number 1. The fingers and thumb contain a total of 14 phalanges (phalanx bones). The thumb contains a proximal and a distal phalanx, whereas the remaining digits each contain proximal, middle, and distal phalanges.
Define the pelvic girdle and describe the bones and ligaments of the pelvis
Explain the three regions of the hip bone and identify their bony landmarks
Describe the openings of the pelvis and the boundaries of the greater and lesser pelvis
The pelvic girdle (hip girdle) is formed by a single bone, the hip bone or coxal bone (coxal = “hip”), which serves as the attachment point for each lower limb. Each hip bone, in turn, is firmly joined to the axial skeleton via its attachment to the sacrum of the vertebral column. The right and left hip bones also converge anteriorly to attach to each other. The bony pelvis is the entire structure formed by the two hip bones, the sacrum, and, attached inferiorly to the sacrum, the coccyx (Figure 11.3.1).
Unlike the bones of the pectoral girdle, which are highly mobile to enhance the range of upper limb movements, the bones of the pelvis are strongly united to each other to form a largely immobile, weight-bearing structure. This is important for stability because it enables the weight of the body to be easily transferred laterally from the vertebral column, through the pelvic girdle and hip joints, and into either lower limb whenever the other limb is not bearing weight. Thus, the immobility of the pelvis provides a strong foundation for the upper body as it rests on top of the mobile lower limbs.
Figure 11.3.1. Pelvis. The pelvic girdle is formed by a single hip bone. The hip bone attaches the lower limb to the axial skeleton through its articulation with the sacrum. The right and left hip bones, plus the sacrum and the coccyx, together form the pelvis.
Hip Bone
The hip bone, or coxal bone, forms the pelvic girdle portion of the pelvis. The paired hip bones are the large, curved bones that form the lateral and anterior aspects of the pelvis. Each adult hip bone is formed by three separate bones that fuse together during the late teenage years. These bony components are the ilium, ischium, and pubis (Figure 11.3.2). These names are retained and used to define the three regions of the adult hip bone.
Figure 11.3.2. The hip bone. The adult hip bone consists of three regions. The ilium forms the large, fan-shaped superior portion, the ischium forms the posteroinferior portion, and the pubis forms the anteromedial portion.
The ilium is the fan-like, superior region that forms the largest part of the hip bone. It is firmly united to the sacrum at the largely immobile sacroiliac joint (see Figure 11.3.1). The ischium forms the posteroinferior region of each hip bone. It supports the body when sitting. The pubis forms the anterior portion of the hip bone. The pubis curves medially, where it joins to the pubis of the opposite hip bone at a specialised joint called the pubic symphysis.
Ilium
When you place your hands on your waist, you can feel the arching, superior margin of the ilium along your waistline (see Figure 11.3.2). This curved, superior margin of the ilium is the iliac crest. The rounded, anterior termination of the iliac crest is the anterior superior iliac spine. This important bony landmark can be felt at your anterolateral hip. Inferior to the anterior superior iliac spine is a rounded protuberance called the anterior inferior iliac spine. Both iliac spines serve as attachment points for muscles of the thigh. Posteriorly, the iliac crest curves downward to terminate as the posterior superior iliac spine. Muscles and ligaments surround but do not cover this bony landmark, thus sometimes producing a depression seen as a “dimple” located on the lower back. More inferiorly is the posterior inferior iliac spine. This is located at the inferior end of a large, roughened area called the auricular surface of the ilium. The auricular surface articulates with the auricular surface of the sacrum to form the sacroiliac joint. Both the posterior superior and posterior inferior iliac spines serve as attachment points for the muscles and very strong ligaments that support the sacroiliac joint.
The shallow depression located on the anteromedial (internal) surface of the upper ilium is called the iliac fossa. The inferior margin of this space is formed by the arcuate line of the ilium, the ridge formed by the pronounced change in curvature between the upper and lower portions of the ilium. The large, inverted U-shaped indentation located on the posterior margin of the lower ilium is called the greater sciatic notch.
Ischium
The ischium forms the posterolateral portion of the hip bone (see Figure 11.3.2). The large, roughened area of the inferior ischium is the ischial tuberosity. This serves as the attachment for the posterior thigh muscles and also carries the weight of the body when sitting. You can feel the ischial tuberosity if you wiggle your pelvis against the seat of a chair. Projecting superiorly and anteriorly from the ischial tuberosity is a narrow segment of bone called the ischial ramus. The slightly curved posterior margin of the ischium above the ischial tuberosity is the lesser sciatic notch. The bony projection separating the lesser sciatic notch and greater sciatic notch is the ischial spine.
Pubis
The pubis forms the anterior portion of the hip bone (see Figure 11.3.2). The enlarged medial portion of the pubis is the pubic body. Located superiorly on the pubic body is a small bump called the pubic tubercle. The superior pubic ramus is the segment of bone that passes laterally from the pubic body to join the ilium. The narrow ridge running along the superior margin of the superior pubic ramus is the pectineal line of the pubis.
The pubic body is joined to the pubic body of the opposite hip bone by the pubic symphysis. Extending downward and laterally from the body is the inferior pubic ramus. The pubic arch is the bony structure formed by the pubic symphysis, and the bodies and inferior pubic rami of the adjacent pubic bones. The inferior pubic ramus extends downward to join the ischial ramus. Together, these form the single ischiopubic ramus, which extends from the pubic body to the ischial tuberosity. The inverted V-shape formed as the ischiopubic rami from both sides come together at the pubic symphysis is called the subpubic angle.
Pelvis
The pelvis consists of four bones: the right and left hip bones, the sacrum, and the coccyx (see Figure 11.3.1). The pelvis has several essential functions. Its primary role is to support the weight of the upper body when sitting and to transfer this weight to the lower limbs when standing. It serves as an attachment point for trunk and lower limb muscles and protects the internal pelvic organs. When standing in the anatomical position, the pelvis is tilted anteriorly. In this position, the anterior superior iliac spines and the pubic tubercles lie in the same vertical plane, and the anterior (internal) surface of the sacrum faces forward and downward.
The three areas of each hip bone, the ilium, pubis, and ischium, converge centrally to form a deep, cup-shaped cavity called the acetabulum. This is located on the lateral side of the hip bone and is part of the hip joint. The large opening in the anteroinferior hip bone between the ischium and pubis is the obturator foramen. This space is largely filled in by a layer of connective tissue and serves for the attachment of muscles on both its internal and external surfaces.
Several ligaments unite the bones of the pelvis (Figure 11.3.3). The largely immobile sacroiliac joint is supported by a pair of strong ligaments that are attached between the sacrum and ilium portions of the hip bone. These are the anterior sacroiliac ligament on the anterior side of the joint and the posterior sacroiliac ligament on the posterior side. Also spanning the sacrum and hip bone are two additional ligaments. The sacrospinous ligament runs from the sacrum to the ischial spine, and the sacrotuberous ligament runs from the sacrum to the ischial tuberosity. These ligaments help to support and immobilize the sacrum as it carries the weight of the body.
Figure 11.3.3. Ligaments of the pelvis. The posterior sacroiliac ligament supports the sacroiliac joint. The sacrospinous ligament spans the sacrum to the ischial spine, and the sacrotuberous ligament spans the sacrum to the ischial tuberosity. The sacrospinous and sacrotuberous ligaments contribute to the formation of the greater and lesser sciatic foramens.
The sacrospinous and sacrotuberous ligaments also help to define two openings on the posterolateral sides of the pelvis through which muscles, nerves, and blood vessels for the lower limb exit. The superior opening is the greater sciatic foramen. This large opening is formed by the greater sciatic notch of the hip bone, the sacrum, and the sacrospinous ligament. The smaller, more inferior lesser sciatic foramen is formed by the lesser sciatic notch of the hip bone, together with the sacrospinous and sacrotuberous ligaments.
The space enclosed by the bony pelvis is divided into two regions. The broad, superior region, defined laterally by the large, fan-like portion of the upper hip bone, is called the greater pelvis (greater pelvic cavity; false pelvis). This broad area is occupied by portions of the small and large intestines, and because it is more closely associated with the abdominal cavity, it is sometimes referred to as the false pelvis. More inferiorly, the narrow, rounded space of the lesser pelvis (lesser pelvic cavity; true pelvis) contains the bladder and other pelvic organs, and thus is also known as the true pelvis. The pelvic brim (also known as the pelvic inlet) forms the superior margin of the lesser pelvis, separating it from the greater pelvis. The pelvic brim is defined by a line formed by the upper margin of the pubic symphysis anteriorly, and the pectineal line of the pubis, the arcuate line of the ilium, and the sacral promontory (the anterior margin of the superior sacrum) posteriorly. The inferior limit of the lesser pelvic cavity is called the pelvic outlet. This large opening is defined by the inferior margin of the pubic symphysis anteriorly, and the ischiopubic ramus, the ischial tuberosity, the sacrotuberous ligament, and the inferior tip of the coccyx posteriorly. Because of the anterior tilt of the pelvis, the lesser pelvis is also angled, giving it an anterosuperior (pelvic inlet) to posteroinferior (pelvic outlet) orientation.
Figure 11.3.4. Male and female pelvis. The female pelvis is adapted for childbirth and is broader, with a larger subpubic angle, a rounder pelvic brim, and a wider and more shallow lesser pelvic cavity than the male pelvis.
Comparison of the Female and Male Pelvis
The differences between the adult female and male pelvis relate to function and body size. In general, the bones of the male pelvis are thicker and heavier, adapted for support of the male’s heavier physical build and stronger muscles. The greater sciatic notch of the male hip bone is narrower and deeper than the broader notch of females. Because the female pelvis is adapted for childbirth, it is wider than the male pelvis, as evidenced by the distance between the anterior superior iliac spines (see Figure 11.3.4). The ischial tuberosities of females are also farther apart, which increases the size of the pelvic outlet. Because of this increased pelvic width, the subpubic angle is larger in females (greater than 80 degrees) than it is in males (less than 70 degrees). The female sacrum is wider, shorter, and less curved, and the sacral promontory projects less into the pelvic cavity, thus giving the female pelvic inlet (pelvic brim) a more rounded or oval shape compared to males. The lesser pelvic cavity of females is also wider and more shallow than the narrower, deeper, and tapering lesser pelvis of males. Because of the obvious differences between female and male hip bones, this is the one bone of the body that allows for the most accurate sex determination. Table 11.3.1 provides an overview of the general differences between the female and male pelvis.
Table 11.3.1 Overview of Differences between the Female and Male Pelvis
Female pelvis
Male pelvis
Pelvic weight
Bones of the pelvis are lighter and thinner
Bones of the pelvis are thicker and heavier
Pelvic inlet shape
Pelvic inlet has a round or oval shape
Pelvic inlet is heart-shaped
Lesser pelvic cavity shape
Lesser pelvic cavity is shorter and wider
Lesser pelvic cavity is longer and narrower
Subpubic angle
Subpubic angle is greater than 80 degrees
Subpubic angle is less than 70 degrees
Pelvic outlet shape
Pelvic outlet is rounded and larger
Pelvic outlet is smaller
Career Connection
Forensic Pathology and Forensic Anthropology
A forensic pathologist (also known as a medical examiner) is a medically trained physician who has been specifically trained in pathology to examine the bodies of the deceased to determine the cause of death. A forensic pathologist applies his or her understanding of disease as well as toxins, blood and DNA analysis, firearms and ballistics, and other factors to assess the cause and manner of death. At times, a forensic pathologist will be called to testify under oath in situations that involve a possible crime. Forensic pathology is a field that has received much media attention on television shows or following a high-profile death.
While forensic pathologists are responsible for determining whether the cause of someone’s death was natural, a suicide, accidental, or a homicide, there are times when uncovering the cause of death is more complex, and other skills are needed. Forensic anthropology brings the tools and knowledge of physical anthropology and human osteology (the study of the skeleton) to the task of investigating a death. A forensic anthropologist assists medical and legal professionals in identifying human remains. The science behind forensic anthropology involves the study of archaeological excavation; the examination of hair; an understanding of plants, insects, and footprints; the ability to determine how much time has elapsed since the person died; the analysis of past medical history and toxicology; the ability to determine whether there are any post-mortem injuries or alterations of the skeleton; and the identification of the decedent (deceased person) using skeletal and dental evidence.
Due to the extensive knowledge and understanding of excavation techniques, a forensic anthropologist is an integral and invaluable team member to have on-site when investigating a crime scene, especially when the recovery of human skeletal remains is involved. When remains are bought to a forensic anthropologist for examination, he or she must first determine whether the remains are in fact human. Once the remains have been identified as belonging to a person and not to an animal, the next step is to approximate the individual’s age, sex, race, and height. The forensic anthropologist does not determine the cause of death, but provides information to the forensic pathologist, who will use all the data collected to make a final determination regarding the cause of death.
In Australia, a completed medical practitioner qualification is required prior to specialising in pathology. Further specialisation and a Fellowship of the Royal College of Pathologists of Australia (FRCPA) is required, which comprises of a five-year specialist training in an accredited position. The average annual salary of a forensic pathologist in Australia is between $69k to $102.
Section Review
The pelvic girdle, consisting of a hip bone, serves to attach a lower limb to the axial skeleton. The hip bone articulates posteriorly at the sacroiliac joint with the sacrum, which is part of the axial skeleton. The right and left hip bones converge anteriorly and articulate with each other at the pubic symphysis. The combination of the hip bone, the sacrum, and the coccyx forms the pelvis. The pelvis has a pronounced anterior tilt. The primary function of the pelvis is to support the upper body and transfer body weight to the lower limbs. It also serves as the site of attachment for multiple muscles.
The hip bone consists of three regions: the ilium, ischium, and pubis. The ilium forms the large, fan-like region of the hip bone. The superior margin of this area is the iliac crest. Located at either end of the iliac crest are the anterior superior and posterior superior iliac spines. Inferior to these are the anterior inferior and posterior inferior iliac spines. The auricular surface of the ilium articulates with the sacrum to form the sacroiliac joint. The medial surface of the upper ilium forms the iliac fossa, with the arcuate line marking the inferior limit of this area. The posterior margin of the ilium has the large greater sciatic notch.
The posterolateral portion of the hip bone is the ischium. It has the expanded ischial tuberosity, which supports body weight when sitting. The ischial ramus projects anteriorly and superiorly. The posterior margin of the ischium has the shallow lesser sciatic notch and the ischial spine, which separates the greater and lesser sciatic notches.
The pubis forms the anterior portion of the hip bone. The body of the pubis articulates with the pubis of the opposite hip bone at the pubic symphysis. The superior margin of the pubic body has the pubic tubercle. The pubis is joined to the ilium by the superior pubic ramus, the superior surface of which forms the pectineal line. The inferior pubic ramus projects inferiorly and laterally. The pubic arch is formed by the pubic symphysis, the bodies of the adjacent pubic bones, and the two inferior pubic rami. The inferior pubic ramus joins the ischial ramus to form the ischiopubic ramus. The subpubic angle is formed by the medial convergence of the right and left ischiopubic rami.
The lateral side of the hip bone has the cup-like acetabulum, which is part of the hip joint. The large anterior opening is the obturator foramen. The sacroiliac joint is supported by the anterior and posterior sacroiliac ligaments. The sacrum is also joined to the hip bone by the sacrospinous ligament, which attaches to the ischial spine, and the sacrotuberous ligament, which attaches to the ischial tuberosity. The sacrospinous and sacrotuberous ligaments contribute to the formation of the greater and lesser sciatic foramina.
The broad space of the upper pelvis is the greater pelvis, and the narrow, inferior space is the lesser pelvis. These areas are separated by the pelvic brim (pelvic inlet). The inferior opening of the pelvis is the pelvic outlet. Compared to the male, the female pelvis is wider to accommodate childbirth, has a larger subpubic angle, and a broader greater sciatic notch.
Identify the divisions of the lower limb and describe the bones of each region
Describe the bones and bony landmarks that articulate at each joint of the lower limb
Like the upper limb, the lower limb is divided into three regions. The thigh is that portion of the lower limb located between the hip joint and knee joint. The leg is specifically the region between the knee joint and the ankle joint. Distal to the ankle is the foot. The lower limb contains 30 bones. These are the femur, patella, tibia, fibula, tarsal bones, metatarsal bones, and phalanges. The femur is the single bone of the thigh. The patella is the kneecap and articulates with the distal femur. The tibia is the larger, weight-bearing bone located on the medial side of the leg, and the fibula is the thin bone of the lateral leg. The bones of the foot are divided into three groups. The posterior portion of the foot is formed by a group of seven bones, each of which is known as a tarsal bone, whereas the mid-foot contains five elongated bones, each of which is a metatarsal bone. The toes contain 14 small bones, each of which is a phalanx bone of the foot.
Femur
The femur, or thigh bone, is the single bone of the thigh region (Figure 11.4.1). It is the longest and strongest bone of the body, and accounts for approximately one-quarter of a person’s total height. The rounded, proximal end is the head of the femur, which articulates with the acetabulum of the hip bone to form the hip joint. The fovea capitis is a minor indentation on the medial side of the femoral head that serves as the site of attachment for the ligament of the head of the femur. This ligament spans the femur and acetabulum but is weak and provides little support for the hip joint. It does, however, carry an important artery that supplies the head of the femur.
Figure 11.4.1. Femur and patella. The femur is the single bone of the thigh region. It articulates superiorly with the hip bone at the hip joint, and inferiorly with the tibia at the knee joint. The patella only articulates with the distal end of the femur.
The narrowed region below the head is the neck of the femur. This is a common area for fractures of the femur. The greater trochanter is the large, upward, bony projection located above the base of the neck. Multiple muscles that act across the hip joint attach to the greater trochanter, which, because of its projection from the femur, gives additional leverage to these muscles. The greater trochanter can be felt just under the skin on the lateral side of your upper thigh. The lesser trochanter is a small, bony prominence that lies on the medial aspect of the femur, just below the neck. A single, powerful muscle attaches to the lesser trochanter. Running between the greater and lesser trochanters on the anterior side of the femur is the roughened intertrochanteric line. The trochanters are also connected on the posterior side of the femur by the larger intertrochanteric crest.
The elongated shaft of the femur has a slight anterior bowing or curvature. At its proximal end, the posterior shaft has the gluteal tuberosity, a roughened area extending inferiorly from the greater trochanter. More inferiorly, the gluteal tuberosity becomes continuous with the linea aspera (“rough line”). This is the roughened ridge that passes distally along the posterior side of the mid-femur. Multiple muscles of the hip and thigh regions make long, thin attachments to the femur along the linea aspera.
The distal end of the femur has medial and lateral bony expansions. On the lateral side, the smooth portion that covers the distal and posterior aspects of the lateral expansion is the lateral condyle of the femur. The roughened area on the outer, lateral side of the condyle is the lateral epicondyle of the femur. Similarly, the smooth region of the distal and posterior medial femur is the medial condyle of the femur, and the irregular outer, medial side of this is the medial epicondyle of the femur. The lateral and medial condyles articulate with the tibia to form the knee joint. The epicondyles provide attachment for muscles and supporting ligaments of the knee. The adductor tubercle is a small bump located at the superior margin of the medial epicondyle. Posteriorly, the medial and lateral condyles are separated by a deep depression called the intercondylar fossa. Anteriorly, the smooth surfaces of the condyles join together to form a wide groove called the patellar surface, which provides for articulation with the patella bone. The combination of the medial and lateral condyles with the patellar surface gives the distal end of the femur a horseshoe (U) shape.
Patella
The patella (kneecap) is largest sesamoid bone of the body (see Figure 11.4.1). A sesamoid bone is a bone that is incorporated into the tendon of a muscle where that tendon crosses a joint. The sesamoid bone articulates with the underlying bones to prevent damage to the muscle tendon due to rubbing against the bones during movements of the joint. The patella is found in the tendon of the quadriceps femoris muscle, the large muscle of the anterior thigh that passes across the anterior knee to attach to the tibia. The patella articulates with the patellar surface of the femur and thus prevents rubbing of the muscle tendon against the distal femur. The patella also lifts the tendon away from the knee joint, which increases the leverage power of the quadriceps femoris muscle as it acts across the knee. The patella does not articulate with the tibia.
Homeostatic Imbalances
Runner’s Knee
Runner’s knee, also known as patellofemoral syndrome, is the most common overuse injury among runners. It is most frequent in adolescents and young adults and is more common in females. It often results from excessive running, particularly downhill, but may also occur in athletes who do a lot of knee bending, such as jumpers, skiers, cyclists, weightlifters, and soccer players. It is felt as a dull, aching pain around the front of the knee and deep to the patella. The pain may be felt when walking or running, going up or down stairs, kneeling or squatting, or after sitting with the knee bent for an extended period.
Patellofemoral syndrome may be initiated by a variety of causes, including individual variations in the shape and movement of the patella, a direct blow to the patella, or flat feet or improper shoes that cause excessive turning in or out of the feet or leg. These factors may cause in an imbalance in the muscle pull that acts on the patella, resulting in an abnormal tracking of the patella that allows it to deviate too far toward the lateral side of the patellar surface on the distal femur.
Because the hips are wider than the knee region, the femur has a diagonal orientation within the thigh, in contrast to the vertically oriented tibia of the leg (Figure 11.4.2). The Q-angle is a measure of how far the femur is angled laterally away from vertical. The Q-angle is normally 10–15 degrees, with females typically having a larger Q-angle due to their wider pelvis. During extension of the knee, the quadriceps femoris muscle pulls the patella both superiorly and laterally, with the lateral pull greater in women due to their large Q-angle. This makes women more vulnerable to developing patellofemoral syndrome than men. Normally, the large lip on the lateral side of the patellar surface of the femur compensates for the lateral pull on the patella, and thus helps to maintain its proper tracking.
However, if the pull produced by the medial and lateral sides of the quadriceps femoris muscle is not balanced, abnormal tracking of the patella toward the lateral side may occur. With continued use, this produces pain and could result in damage to the articulating surfaces of the patella and femur, and the potential future development of arthritis. Treatment involves stopping the activity that produces knee pain for a period, followed by a gradual resumption of activity. Proper strengthening of the quadriceps femoris muscle to correct for imbalances is also important to help prevent reoccurrence.
Figure 11.4.2. The Q-angle. The Q-angle is a measure of the amount of lateral deviation of the femur from the vertical line of the tibia. Adult females have a larger Q-angle due to their wider pelvis than adult males.
Tibia
The tibia (shin bone) is the medial bone of the leg and is larger than the fibula, with which it is paired (Figure 11.4.3). The tibia is the main weight-bearing bone of the lower leg and the second longest bone of the body, after the femur. The medial side of the tibia is located immediately under the skin, allowing it to be easily palpated down the entire length of the medial leg.
Figure 11.4.3. Tibia and fibula. The tibia is the larger, weight-bearing bone located on the medial side of the leg. The fibula is the slender bone of the lateral side of the leg and does not bear weight.
The proximal end of the tibia is greatly expanded. The two sides of this expansion form the medial condyle of the tibia and the lateral condyle of the tibia. The tibia does not have epicondyles. The top surface of each condyle is smooth and flattened. These areas articulate with the medial and lateral condyles of the femur to form the knee joint. Between the articulating surfaces of the tibial condyles is the intercondylar eminence, an irregular, elevated area that serves as the inferior attachment point for two supporting ligaments of the knee.
The tibial tuberosity is an elevated area on the anterior side of the tibia, near its proximal end. It is the final site of attachment for the muscle tendon associated with the patella. More inferiorly, the shaft of the tibia becomes triangular in shape. The anterior apex of
MH this triangle forms the anterior border of the tibia, which begins at the tibial tuberosity and runs inferiorly along the length of the tibia. Both the anterior border and the medial side of the triangular shaft are located immediately under the skin and can be easily palpated along the entire length of the tibia. A small ridge running down the lateral side of the tibial shaft is the interosseous border of the tibia. This is for the attachment of the interosseous membrane of the leg, the sheet of dense connective tissue that unites the tibia and fibula bones. Located on the posterior side of the tibia is the soleal line, a diagonally running, roughened ridge that begins below the base of the lateral condyle, and runs down and medially across the proximal third of the posterior tibia. Muscles of the posterior leg attach to this line.
The large expansion found on the medial side of the distal tibia is the medial malleolus (“little hammer”). This forms the large bony bump found on the medial side of the ankle region. Both the smooth surface on the inside of the medial malleolus and the smooth area at the distal end of the tibia articulate with the talus bone of the foot as part of the ankle joint. On the lateral side of the distal tibia is a wide groove called the fibular notch. This area articulates with the distal end of the fibula, forming the distal tibiofibular joint.
Fibula
The fibula is the slender bone located on the lateral side of the leg (see Figure 11.4.3). The fibula does not bear weight. It serves primarily for muscle attachments and thus is surrounded by muscles. Only the proximal and distal ends of the fibula can be palpated.
The head of the fibula is the small, knob-like, proximal end of the fibula. It articulates with the inferior aspect of the lateral tibial condyle, forming the proximal tibiofibular joint. The thin shaft of the fibula has the interosseous border of the fibula, a narrow ridge running down its medial side for the attachment of the interosseous membrane that spans the fibula and tibia. The distal end of the fibula forms the lateral malleolus, which forms the easily palpated bony bump on the lateral side of the ankle. The deep (medial) side of the lateral malleolus articulates with the talus bone of the foot as part of the ankle joint. The distal fibula also articulates with the fibular notch of the tibia.
Tarsal Bones
The posterior half of the foot is formed by seven tarsal bones (Figure 11.4.4). The most superior bone is the talus. This has a square-shaped upper surface that articulates with the tibia and fibula to form the ankle joint. Three areas of articulation form the ankle joint: The superomedial surface of the talus bone articulates with the medial malleolus of the tibia, the top of the talus articulates with the distal end of the tibia, and the lateral side of the talus articulates with the lateral malleolus of the fibula. Inferiorly, the talus articulates with the calcaneus (heel bone), the largest bone of the foot, which forms the heel. Body weight is transferred from the tibia to the talus to the calcaneus, which rests on the ground. The medial calcaneus has a prominent bony extension called the sustentaculum tali (“support for the talus”) that supports the medial side of the talus bone.
Figure 11.4.4. Bones of the foot. The bones of the foot are divided into three groups. The posterior foot is formed by the seven tarsal bones. The mid-foot has the five metatarsal bones. The toes contain the phalanges.
The cuboid bone articulates with the anterior end of the calcaneus bone. The cuboid has a deep groove running across its inferior surface, which provides passage for a muscle tendon. The talus bone articulates anteriorly with the navicular bone, which in turn articulates anteriorly with the three cuneiform (“wedge-shaped”) bones. These bones are the medial cuneiform, the intermediate cuneiform, and the lateral cuneiform. Each of these bones has a broad superior surface and a narrow inferior surface, which together produce the transverse (medial-lateral) curvature of the foot. The navicular and lateral cuneiform bones also articulate with the medial side of the cuboid bone.
Metatarsal Bones
The anterior half of the foot is formed by the five metatarsal bones, which are located between the tarsal bones of the posterior foot and the phalanges of the toes (see Figure 11.4.4). These elongated bones are numbered 1–5, starting with the medial side of the foot. The first metatarsal bone is shorter and thicker than the others. The second metatarsal is the longest. The base of the metatarsal bone is the proximal end of each metatarsal bone. These articulate with the cuboid or cuneiform bones. The base of the fifth metatarsal has a large, lateral expansion that provides for muscle attachments. This expanded base of the fifth metatarsal can be felt as a bony bump at the midpoint along the lateral border of the foot. The expanded distal end of each metatarsal is the head of the metatarsal bone. Each metatarsal bone articulates with the proximal phalanx of a toe to form a metatarsophalangeal joint. The heads of the metatarsal bones also rest on the ground and form the ball (anterior end) of the foot.
Phalanges
The toes contain a total of 14 phalanx bones (phalanges), arranged in a similar manner as the phalanges of the fingers (see Figure 11.4.4). The toes are numbered 1–5, starting with the big toe (hallux). The big toe has two phalanx bones, the proximal and distal phalanges. The remaining toes all have proximal, middle, and distal phalanges. A joint between adjacent phalanx bones is called an interphalangeal joint.
Arches of the Foot
When the foot meets the ground during walking, running, or jumping activities, the impact of the body weight puts a tremendous amount of pressure and force on the foot. During running, the force applied to each foot as it contacts the ground can be up to 2.5 times your body weight. The bones, joints, ligaments, and muscles of the foot absorb this force, thus greatly reducing the amount of shock that is passed superiorly into the lower limb and body. The arches of the foot play a key role in this shock-absorbing ability. When weight is applied to the foot, these arches will flatten, thus absorbing energy. When the weight is removed, the arch rebounds, giving “spring” to the step. The arches also serve to distribute body weight side to side and to either end of the foot.
The foot has a transverse arch, a medial longitudinal arch, and a lateral longitudinal arch (see Figure 11.4.4). The transverse arch forms the medial-lateral curvature of the mid-foot. It is formed by the wedge shapes of the cuneiform bones and bases (proximal ends) of the first to fourth metatarsal bones. This arch helps to distribute body weight from side to side within the foot, thus allowing the foot to accommodate uneven terrain.
The longitudinal arches run down the length of the foot. The lateral longitudinal arch is flat, whereas the medial longitudinal arch is larger (taller). The longitudinal arches are formed by the tarsal bones posteriorly and the metatarsal bones anteriorly. These arches are supported at either end, where they contact the ground. Posteriorly, this support is provided by the calcaneus bone and anteriorly by the heads (distal ends) of the metatarsal bones. The talus bone, which receives the weight of the body, is located at the top of the longitudinal arches. Body weight is then conveyed from the talus to the ground by the anterior and posterior ends of these arches. Strong ligaments unite the adjacent foot bones to prevent disruption of the arches during weight bearing. On the bottom of the foot, additional ligaments tie together the anterior and posterior ends of the arches. These ligaments have elasticity, which allows them to stretch during weight bearing, thus allowing the longitudinal arches to spread. The stretching of these ligaments stores energy within the foot, rather than passing these forces into the leg. Contraction of the foot muscles also plays a key role in this energy absorption. When the weight is removed, the elastic ligaments recoil and pull the ends of the arches closer together. This recovery of the arches releases the stored energy and improves the energy efficiency of walking.
Stretching of the ligaments that support the longitudinal arches can lead to pain. This can occur in overweight individuals, with people who have jobs that involve standing for extended periods of time (such as wait-staff) or walking or running long distances. If stretching of the ligaments is prolonged, excessive, or repeated, it can result in a gradual lengthening of the supporting ligaments, with subsequent depression or collapse of the longitudinal arches, particularly on the medial side of the foot. This condition is called pes planus (“flat foot” or “fallen arches”).
Section Review
The lower limb is divided into three regions. These are the thigh, located between the hip and knee joints; the leg, located between the knee and ankle joints; and distal to the ankle, the foot. There are 30 bones in each lower limb. These are the femur, patella, tibia, fibula, seven tarsal bones, five metatarsal bones, and 14 phalanges.
The femur is the single bone of the thigh. Its rounded head articulates with the acetabulum of the hip bone to form the hip joint. The head has the fovea capitis for attachment of the ligament of the head of the femur. The narrow neck joins inferiorly with the greater and lesser trochanters. Passing between these bony expansions are the intertrochanteric line on the anterior femur and the larger intertrochanteric crest on the posterior femur. On the posterior shaft of the femur is the gluteal tuberosity proximally and the linea aspera in the mid-shaft region. The expanded distal end consists of three articulating surfaces: the medial and lateral condyles, and the patellar surface. The outside margins of the condyles are the medial and lateral epicondyles. The adductor tubercle is on the superior aspect of the medial epicondyle.
The patella is a sesamoid bone located within a muscle tendon. It articulates with the patellar surface on the anterior side of the distal femur, thereby protecting the muscle tendon from rubbing against the femur.
The leg contains the large tibia on the medial side and the slender fibula on the lateral side. The tibia bears the weight of the body, whereas the fibula does not bear weight. The interosseous border of each bone is the attachment site for the interosseous membrane of the leg, the connective tissue sheet that unites the tibia and fibula.
The proximal tibia consists of the expanded medial and lateral condyles, which articulate with the medial and lateral condyles of the femur to form the knee joint. Between the tibial condyles is the intercondylar eminence. On the anterior side of the proximal tibia is the tibial tuberosity, which is continuous inferiorly with the anterior border of the tibia. On the posterior side, the proximal tibia has the curved soleal line. The bony expansion on the medial side of the distal tibia is the medial malleolus. The groove on the lateral side of the distal tibia is the fibular notch.
The head of the fibula forms the proximal end and articulates with the underside of the lateral condyle of the tibia. The distal fibula articulates with the fibular notch of the tibia. The expanded distal end of the fibula is the lateral malleolus.
The posterior foot is formed by the seven tarsal bones. The talus articulates superiorly with the distal tibia, the medial malleolus of the tibia, and the lateral malleolus of the fibula to form the ankle joint. The talus articulates inferiorly with the calcaneus bone. The sustentaculum tali of the calcaneus helps to support the talus. Anterior to the talus is the navicular bone, and anterior to this are the medial, intermediate, and lateral cuneiform bones. The cuboid bone is anterior to the calcaneus.
The five metatarsal bones form the anterior foot. The base of these bones articulates with the cuboid or cuneiform bones. The metatarsal heads, at their distal ends, articulate with the proximal phalanges of the toes. The big toe (toe number 1) has proximal and distal phalanx bones. The remaining toes have proximal, middle, and distal phalanges.
Describe the growth and development of the embryonic limb buds
Discuss the appearance of primary and secondary ossification centres
Embryologically, the appendicular skeleton arises from mesenchyme, a type of embryonic tissue that can differentiate into many types of tissues, including bone or muscle tissue. Mesenchyme gives rise to the bones of the upper and lower limbs, as well as to the pectoral and pelvic girdles. Development of the limbs begins near the end of the fourth embryonic week, with the upper limbs appearing first. Thereafter, the development of the upper and lower limbs follows similar patterns, with the lower limbs lagging the upper limbs by a few days.
Limb Growth
Figure 11.5.1. Embryo at seven weeks. Limb buds are visible in an embryo at the end of the seventh week of development (embryo derived from an ectopic pregnancy). (credit: Ed Uthman/flickr).
Each upper and lower limb initially develops as a small bulge called a limb bud, which appears on the lateral side of the early embryo. The upper limb bud appears near the end of the fourth week of development, with the lower limb bud appearing shortly after (Figure 11.5.1).
Initially, the limb buds consist of a core of mesenchyme covered by a layer of ectoderm. The ectoderm at the end of the limb bud thickens to form a narrow crest called the apical ectodermal ridge. This ridge stimulates the underlying mesenchyme to rapidly proliferate, producing the outgrowth of the developing limb. As the limb bud elongates, cells located farther from the apical ectodermal ridge slow their rates of cell division and begin to differentiate. In this way, the limb develops along a proximal-to-distal axis.
During the sixth week of development, the distal ends of the upper and lower limb buds expand and flatten into a paddle shape. This region will become the hand or foot. The wrist or ankle areas then appear as a constriction that develops at the base of the paddle. Shortly after this, a second constriction on the limb bud appears at the future site of the elbow or knee. Within the paddle, areas of tissue undergo cell death, producing separations between the growing fingers and toes. Also, during the sixth week of development, mesenchyme within the limb buds begins to differentiate into hyaline cartilage that will form models of the future limb bones.
The early outgrowth of the upper and lower limb buds initially has the limbs positioned so that the regions that will become the palm of the hand or the bottom of the foot are facing medially toward the body, with the future thumb or big toe both oriented toward the head. During the seventh week of development, the upper limb rotates laterally by 90 degrees, so that the palm of the hand faces anteriorly and the thumb points laterally. In contrast, the lower limb undergoes a 90-degree medial rotation, thus bringing the big toe to the medial side of the foot
Ossification of Appendicular Bones
All the girdle and limb bones, except for the clavicle, develop by the process of endochondral ossification. This process begins as the mesenchyme within the limb bud differentiates into hyaline cartilage to form cartilage models for future bones. By the twelfth week, a primary ossification centre will have appeared in the diaphysis (shaft) region of the long bones, initiating the process that converts the cartilage model into bone. A secondary ossification centre will appear in each epiphysis (expanded end) of these bones later, usually after birth. The primary and secondary ossification centres are separated by the epiphyseal plate, a layer of growing hyaline cartilage. This plate is located between the diaphysis and each epiphysis. It continues to grow and is responsible for the lengthening of the bone. The epiphyseal plate is retained for many years, until the bone reaches its final, adult size, at which time the epiphyseal plate disappears and the epiphysis fuses to the diaphysis. (Seek additional content on ossification in the chapter on bone tissue.)
Small bones, such as the phalanges, will develop only one secondary ossification centre and will thus have only a single epiphyseal plate. Large bones, such as the femur, will develop several secondary ossification centres, with an epiphyseal plate associated with each secondary centre. Thus, ossification of the femur begins at the end of the seventh week with the appearance of the primary ossification centre in the diaphysis, which rapidly expands to ossify the shaft of the bone prior to birth. Secondary ossification centres develop at later times. Ossification of the distal end of the femur, to form the condyles and epicondyles, begins shortly before birth. Secondary ossification centres also appear in the femoral head late in the first year after birth, in the greater trochanter during the fourth year, and in the lesser trochanter between the ages of 9 and 10 years. Once these areas have ossified, their fusion to the diaphysis and the disappearance of each epiphyseal plate follow a reversed sequence. Thus, the lesser trochanter is the first to fuse, doing so at the onset of puberty (around 11 years of age), followed by the greater trochanter approximately 1 year later. The femoral head fuses between the ages of 14–17 years, whereas the distal condyles of the femur are the last to fuse, between the ages of 16–19 years. Knowledge of the age at which different epiphyseal plates disappear is important when interpreting radiographs taken of children. Since the cartilage of an epiphyseal plate is less dense than bone, the plate will appear dark in a radiograph image. Thus, a normal epiphyseal plate may be mistaken for a bone fracture.
The clavicle is the one appendicular skeleton bone that does not develop via endochondral ossification. Instead, the clavicle develops through the process of intramembranous ossification. During this process, mesenchymal cells differentiate directly into bone-producing cells, which produce the clavicle directly, without first making a cartilage model. Because of this early production of bone, the clavicle is the first bone of the body to begin ossification, with ossification centres appearing during the fifth week of development. However, ossification of the clavicle is not complete until age 25.
Disorders of the Appendicular System: Congenital Clubfoot
Clubfoot, also known as talipes, is a congenital (present at birth) disorder of unknown cause and is the most common deformity of the lower limb. It affects the foot and ankle, causing the foot to be twisted inward at a sharp angle, like the head of a golf club (Figure 11.5.2). Clubfoot has a frequency of about 1 out of every 1,000 births worldwide and is twice as likely to occur in a male child as in a female child. In 50 percent of cases, both feet are affected. To date there is no combined Australian rate of this disease, however it has been shown that the rate is higher in the Indigenous population.
Figure 11.5.2. Clubfoot. Clubfoot is a common deformity of the ankle and foot that is present at birth. Most cases are corrected without surgery, and affected individuals will grow up to lead normal, active lives. (credit: James W. Hanson).
At birth, children with a clubfoot have the heel turned inward and the anterior foot twisted so that the lateral side of the foot is facing inferiorly, commonly due to ligaments or leg muscles attached to the foot that are shortened or abnormally tight. These pull the foot into an abnormal position, resulting in bone deformities. Other symptoms may include bending of the ankle that lifts the heel of the foot and an extremely high foot arch. Due to the limited range of motion in the affected foot, it is difficult to place the foot into the correct position. Additionally, the affected foot may be shorter than normal, and the calf muscles are usually underdeveloped on the affected side. Despite the appearance, this is not a painful condition for newborns. However, it must be treated early to avoid future pain and impaired walking ability.
Although the cause of clubfoot is idiopathic (unknown), evidence indicates that foetal position within the uterus is not a contributing factor. Genetic factors are involved, because clubfoot tends to run within families. Cigarette smoking during pregnancy has been linked to the development of clubfoot, particularly in families with a history of clubfoot.
Previously, clubfoot required extensive surgery. Today, 90 percent of cases are successfully treated without surgery using new corrective casting techniques. The best chance for a full recovery requires that clubfoot treatment begin during the first 2 weeks after birth. Corrective casting gently stretches the foot, which is followed by the application of a holding cast to keep the foot in the proper position. This stretching and casting is repeated weekly for several weeks. In severe cases, surgery may also be required, after which the foot typically remains in a cast for 6 to 8 weeks. After the cast is removed following either surgical or nonsurgical treatment, the child will be required to wear a brace part-time (at night) for up to 4 years. In addition, special exercises will be prescribed, and the child must also wear special shoes. Close monitoring by the parents and adherence to postoperative instructions are imperative in minimising the risk of relapse.
Despite these difficulties, treatment for clubfoot is usually successful, and the child will grow up to lead a normal, active life. Numerous examples of individuals born with a clubfoot who went on to successful careers include Dudley Moore (comedian and actor), Damon Wayans (comedian and actor), Troy Aikman (three-time Super Bowl-winning quarterback), Kristi Yamaguchi (Olympic gold medallist in figure skating), Mia Hamm (two-time Olympic gold medallist in soccer), and Charles Woodson (Heisman trophy and Super Bowl winner).
Section Review
The bones of the appendicular skeleton arise from embryonic mesenchyme. Limb buds appear at the end of the fourth week. The apical ectodermal ridge, located at the end of the limb bud, stimulates growth and elongation of the limb. During the sixth week, the distal end of the limb bud becomes paddle-shaped, and selective cell death separates the developing fingers and toes. At the same time, mesenchyme within the limb bud begins to differentiate into hyaline cartilage, forming models for future bones. During the seventh week, the upper limbs rotate laterally, and the lower limbs rotate medially, bringing the limbs into their final positions.
Endochondral ossification, the process that converts the hyaline cartilage model into bone, begins in most appendicular bones by the twelfth foetal week. This begins as a primary ossification centre in the diaphysis, followed by the later appearance of one or more secondary ossifications centres in the regions of the epiphyses. Each secondary ossification centre is separated from the primary ossification centre by an epiphyseal plate. Continued growth of the epiphyseal plate cartilage provides for bone lengthening. Disappearance of the epiphyseal plate is followed by fusion of the bony components to form a single, adult bone.
The clavicle develops via intramembranous ossification, in which mesenchyme is converted directly into bone tissue. Ossification within the clavicle begins during the fifth week of development and continues until 25 years of age.
Distinguish between the functional and structural classifications for joints
Describe the three functional types of joints and give an example of each
List the three types of diarthrodial joints
A joint, also called an articulation, is any place where adjacent bones or bone and cartilage come together (articulate with each other) to form a connection. Joints are classified both structurally and functionally. Structural classifications of joints consider whether the adjacent bones are strongly anchored to each other by fibrous connective tissue or cartilage, or whether the adjacent bones articulate with each other within a fluid-filled space called a joint cavity. Functional classifications describe the degree of movement available between the bones, ranging from immobile, to slightly mobile, to freely moveable joints. The amount of movement available at a particular joint of the body is related to the functional requirements for that joint. Thus, immobile or slightly moveable joints serve to protect internal organs, give stability to the body, and allow for limited body movement. In contrast, freely moveable joints allow for much more extensive movements of the body and limbs.
Structural Classification of Joints
The structural classification of joints is based on whether the articulating surfaces of the adjacent bones are directly connected by fibrous connective tissue or cartilage, or whether the articulating surfaces contact each other within a fluid-filled joint cavity. These differences serve to divide the joints of the body into three structural classifications. A fibrous joint is where the adjacent bones are united by fibrous connective tissue. At a cartilaginous joint, the bones are joined by hyaline cartilage or fibrocartilage. At a synovial joint, the articulating surfaces of the bones are not directly connected, but instead meet each other within a joint cavity that is filled with a lubricating fluid. Synovial joints allow for free movement between the bones and are the most common joints of the body.
Functional Classification of Joints
The functional classification of joints is determined by the amount of mobility found between the adjacent bones. Joints are thus functionally classified as a synarthrosis or immobile joint, an amphiarthrosis or slightly moveable joint, or as a diarthrosis, which is a freely moveable joint (arthroun = “to fasten by a joint”). Depending on their location, fibrous joints may be functionally classified as a synarthrosis (immobile joint) or an amphiarthrosis (slightly mobile joint). Cartilaginous joints are also functionally classified as either a synarthrosis or an amphiarthrosis joint. All synovial joints are functionally classified as a diarthrosis joint.
Synarthrosis
An immobile or nearly immobile joint is called a synarthrosis. The immobile nature of these joints provides for a strong union between the articulating bones. This is important at locations where the bones provide protection for internal organs. Examples include sutures, the fibrous joints between the bones of the skull that surround and protect the brain (Figure 11.6.1), and the manubriosternal joint, the cartilaginous joint that unites the manubrium and body of the sternum for protection of the heart.
Figure 11.6.1. Suture joints of skull. The suture joints of the skull are an example of a synarthrosis, an immobile or essentially immobile joint.
Amphiarthrosis
An amphiarthrosis is a joint that has limited mobility. An example of this type of joint is the cartilaginous joint that unites the bodies of adjacent vertebrae. Filling the gap between the vertebrae is a thick pad of fibrocartilage called an intervertebral disc (Figure 11.6.2). Each intervertebral disc strongly unites the vertebrae but still allows for a limited amount of movement between them. However, the small movements available between adjacent vertebrae can sum together along the length of the vertebral column to provide for large ranges of body movements.
Another example of an amphiarthrosis is the pubic symphysis of the pelvis. This is a cartilaginous joint in which the pubic regions of the right and left hip bones are strongly anchored to each other by fibrocartilage. This joint normally has little mobility. The strength of the pubic symphysis is important in conferring weight-bearing stability to the pelvis.
Figure 11.6.2. Intervertebral disc. An intervertebral disc unites the bodies of adjacent vertebrae within the vertebral column. Each disc allows for limited movement between the vertebrae and thus functionally forms an amphiarthrosis type of joint. Intervertebral discs are made of fibrocartilage and thereby structurally form a symphysis type of cartilaginous joint.
Diarthrosis
A freely mobile joint is classified as a diarthrosis. These types of joints include all synovial joints of the body, which provide most body movements. Most diarthrotic joints are found in the appendicular skeleton and thus give the limbs a wide range of motion. These joints are divided into three categories, based on the number of axes of motion provided by each. An axis in anatomy is described as the movements in reference to the three anatomical planes: transverse, frontal, and sagittal. Thus, diarthroses are classified as uniaxial (for movement in one plane), biaxial (for movement in two planes), or multiaxial joints (for movement in all three anatomical planes).
A uniaxial joint only allows for a motion in a single plane (around a single axis). The elbow joint, which only allows for bending or straightening, is an example of a uniaxial joint. A biaxial joint allows for motions within two planes. An example of a biaxial joint is a metacarpophalangeal joint (knuckle joint) of the hand. The joint allows for movement along one axis to produce bending or straightening of the finger, and movement along a second axis, which allows for spreading of the fingers away from each other and bringing them together. A joint that allows for the several directions of movement is called a multiaxial joint (polyaxial or triaxial joint). This type of diarthrotic joint allows for movement along three axes (Figure 11.6.3). The shoulder and hip joints are multiaxial joints. They allow the upper or lower limb to move in an anterior-posterior direction and a medial-lateral direction. In addition, the limb can also be rotated around its long axis. This third movement results in rotation of the limb so that its anterior surface is moved either toward or away from the midline of the body.
Figure 11.6.3. Multiaxial joint. A multiaxial joint, such as the hip joint, allows for three types of movement: anterior-posterior, medial-lateral, and rotational.
Section Review
Structural classifications of the body joints are based on how the bones are held together and articulate with each other. At fibrous joints, the adjacent bones are directly united to each other by fibrous connective tissue. Similarly, at a cartilaginous joint, the adjacent bones are united by cartilage. In contrast, at a synovial joint, the articulating bone surfaces are not directly united to each other but come together within a fluid-filled joint cavity.
The functional classification of body joints is based on the degree of movement found at each joint. A synarthrosis is a joint that is essentially immobile. This type of joint provides for a strong connection between the adjacent bones, which serves to protect internal structures such as the brain or heart. Examples include the fibrous joints of the skull sutures and the cartilaginous manubriosternal joint. A joint that allows for limited movement is an amphiarthrosis. An example is the pubic symphysis of the pelvis, the cartilaginous joint that strongly unites the right and left hip bones of the pelvis. The cartilaginous joints in which vertebrae are united by intervertebral discs provide for small movements between the adjacent vertebrae and are also an amphiarthrosis type of joint. Thus, based on their movement ability, both fibrous and cartilaginous joints are functionally classified as a synarthrosis or amphiarthrosis.
The most common type of joint is the diarthrosis, which is a freely moveable joint. All synovial joints are functionally classified as diarthroses. A uniaxial diarthrosis, such as the elbow, is a joint that only allows for movement within a single anatomical plane. Joints that allow for movements in two planes are biaxial joints, such as the metacarpophalangeal joints of the fingers. A multiaxial joint, such as the shoulder or hip joint, allows for three planes of motions.
Describe the structural features of fibrous joints
Distinguish between a suture, syndesmosis and gomphosis
Give an example of each type of fibrous joint
At a fibrous joint, the adjacent bones are directly connected to each other by fibrous connective tissue, and thus the bones do not have a joint cavity between them (Figure 11.7.1). The gap between the bones may be narrow or wide. There are three types of fibrous joints. A suture is the narrow fibrous joint found between most bones of the skull. At a syndesmosis joint, the bones are more widely separated but are held together by a narrow band of fibrous connective tissue called a ligament or a wide sheet of connective tissue called an interosseous membrane. This type of fibrous joint is found between the shaft regions of the long bones in the forearm and in the leg. Lastly, a gomphosis is the narrow fibrous joint between the roots of a tooth and the bony socket in the jaw into which the tooth fits.
Figure 11.7.1. Fibrous joints. Fibrous joints form strong connections between bones. (a) Sutures join most bones of the skull. (b) An interosseous membrane forms a syndesmosis between the radius and ulna bones of the forearm. (c) A gomphosis is a specialised fibrous joint that anchors a tooth to its socket in the jaw.
Suture
All the bones of the skull, except for the mandible, are joined to each other by a fibrous joint called a suture. The fibrous connective tissue found at a suture (“to bind or sew”) strongly unites the adjacent skull bones and thus helps to protect the brain and form the face. In adults, the skull bones are closely apposed and fibrous connective tissue fills the narrow gap between the bones. The suture is frequently convoluted, forming a tight union that prevents most movement between the bones. (See Figure 11.7.1a). Thus, skull sutures are functionally classified as a synarthrosis, although some sutures may allow for slight movements between the cranial bones.
In newborns and infants, the areas of connective tissue between the bones are much wider, especially in those areas on the top and sides of the skull that will become the sagittal, coronal, squamous, and lambdoid sutures. These broad areas of connective tissue are called fontanelles (Figure 11.7.2). During birth, the fontanelles provide flexibility to the skull, allowing the bones to push closer together or to overlap slightly, thus aiding movement of the infant’s head through the birth canal. After birth, these expanded regions of connective tissue allow for rapid growth of the skull and enlargement of the brain. The fontanelles decrease in width during the first year after birth as the skull bones enlarge. When the connective tissue between the adjacent bones is reduced to a narrow layer, these fibrous joints are now called sutures. At some sutures, the connective tissue will ossify and be converted into bone, causing the adjacent bones to fuse to each other. This fusion between bones is called a synostosis (“joined by bone”). Examples of synostosis fusions between cranial bones are found both early and late in life. At the time of birth, the frontal and maxillary bones consist of right and left halves joined together by sutures, which disappear by the eighth year as the halves fuse together to form a single bone. Late in life, the sagittal, coronal, and lambdoid sutures of the skull will begin to ossify and fuse, causing the suture line to gradually disappear.
Figure 11.7.2. The newborn skull. The fontanelles of a newborn’s skull are broad areas of fibrous connective tissue that form fibrous joints between the bones of the skull.
Syndesmosis
A syndesmosis (“fastened with a band”) is a type of fibrous joint in which two parallel bones are united to each other by fibrous connective tissue. The gap between the bones may be narrow, with the bones joined by ligaments, or the gap may be wide and filled in by a broad sheet of connective tissue called an interosseous membrane.
In the forearm, the wide gap between the shaft portions of the radius and ulna bones are strongly united by an interosseous membrane (see Figure 11.7.1b). Similarly, in the leg, the shafts of the tibia and fibula are also united by an interosseous membrane. In addition, at the distal tibiofibular joint, the articulating surfaces of the bones lack cartilage and the narrow gap between the bones is anchored by fibrous connective tissue and ligaments on both the anterior and posterior aspects of the joint. Together, the interosseous membrane and these ligaments form the tibiofibular syndesmosis.
The syndesmoses found in the forearm and leg serve to unite parallel bones and prevent their separation. However, a syndesmosis does not prevent all movement between the bones, and thus this type of fibrous joint is functionally classified as an amphiarthrosis. In the leg, the syndesmosis between the tibia and fibula strongly unites the bones, allows for little movement, and firmly locks the talus bone in place between the tibia and fibula at the ankle joint. This provides strength and stability to the leg and ankle, which are important during weight bearing. In the forearm, the interosseous membrane is flexible enough to allow for rotation of the radius bone during forearm movements. Thus, in contrast to the stability provided by the tibiofibular syndesmosis, the flexibility of the antebrachial interosseous membrane allows for the much greater mobility of the forearm.
The interosseous membranes of the leg and forearm also provide areas for muscle attachment. Damage to a syndesmotic joint, which usually results from a fracture of the bone with an accompanying tear of the interosseous membrane, will produce pain, loss of stability of the bones, and may damage the muscles attached to the interosseous membrane. If the fracture site is not properly immobilised with a cast or splint, contractile activity by these muscles can cause improper alignment of the broken bones during healing.
Gomphosis
A gomphosis (“fastened with bolts”) is the specialised fibrous joint that anchors the root of a tooth into its bony socket within the maxillary bone (upper jaw) or mandible bone (lower jaw) of the skull. A gomphosis is also known as a peg-and-socket joint. Spanning between the bony walls of the socket and the root of the tooth are numerous short bands of dense connective tissue, each of which is called a periodontal ligament (see Figure 11.7.1c). Due to the immobility of a gomphosis, this type of joint is functionally classified as a synarthrosis.
Section Review
Fibrous joints are where adjacent bones are strongly united by fibrous connective tissue. The gap filled by connective tissue may be narrow or wide. The three types of fibrous joints are sutures, gomphoses, and syndesmoses. A suture is the narrow fibrous joint that unites most bones of the skull. At a gomphosis, the root of a tooth is anchored across a narrow gap by periodontal ligaments to the walls of its socket in the bony jaw. A syndesmosis is the type of fibrous joint found between parallel bones. The gap between the bones may be wide and filled with a fibrous interosseous membrane, or it may narrow with ligaments spanning between the bones. Syndesmoses are found between the bones of the forearm (radius and ulna) and the leg (tibia and fibula). Fibrous joints strongly unite adjacent bones and thus serve to provide protection for internal organs, strength to body regions, or weight-bearing stability.
Describe the structural features of cartilaginous joints
Distinguish between a synchondrosis and symphysis
Give an example of each type of cartilaginous joint
As the name indicates, at a cartilaginous joint, the adjacent bones are united by cartilage, a tough but flexible type of connective tissue. These types of joints lack a joint cavity and involve bones that are joined together by either hyaline cartilage or fibrocartilage (Figure 11.8.1). There are two types of cartilaginous joints. A synchondrosis is a cartilaginous joint where the bones are joined by hyaline cartilage. Also classified as a synchondrosis are places where bone is united to a cartilage structure, such as between the anterior end of a rib and the costal cartilage of the thoracic cage. The second type of cartilaginous joint is a symphysis, where the bones are joined by fibrocartilage.
Figure 11.8.1. Cartilaginous joints. At cartilaginous joints, bones are united by hyaline cartilage to form a synchondrosis or by fibrocartilage to form a symphysis. (a) The hyaline cartilage of the epiphyseal plate (growth plate) forms a synchondrosis that unites the shaft (diaphysis) and end (epiphysis) of a long bone and allows the bone to grow in length. (b) The pubic portions of the right and left hip bones of the pelvis are joined together by fibrocartilage, forming the pubic symphysis.
Synchondrosis
A synchondrosis (“joined by cartilage”) is a cartilaginous joint where bones are joined together by hyaline cartilage, or where bone is united to hyaline cartilage. A synchondrosis may be temporary or permanent. A temporary synchondrosis is the epiphyseal plate (growth plate) of a growing long bone. The epiphyseal plate is the region of growing hyaline cartilage that unites the diaphysis (shaft) of the bone to the epiphysis (end of the bone). Bone lengthening involves growth of the epiphyseal plate cartilage and its replacement by bone, which adds to the diaphysis. For many years during childhood growth, the rates of cartilage growth and bone formation are equal and thus the epiphyseal plate does not change in overall thickness as the bone lengthens. During the late teens and early 20s, growth of the cartilage slows and eventually stops. The epiphyseal plate is then completely replaced by bone, and the diaphysis and epiphysis portions of the bone fuse together to form a single adult bone. This fusion of the diaphysis and epiphysis is a synostosis. Once this occurs, bone lengthening ceases. For this reason, the epiphyseal plate is a temporary synchondrosis. Because cartilage is softer than bone tissue, injury to a growing long bone can damage the epiphyseal plate cartilage, thus stopping bone growth and preventing additional bone lengthening.
Growing layers of cartilage also form synchondroses that join together the ilium, ischium, and pubic portions of the hip bone during childhood and adolescence. When body growth stops, the cartilage disappears and is replaced by bone, forming synostoses and fusing the bony components together into the single hip bone of the adult. Similarly, synostoses unite the sacral vertebrae that fuse together to form the adult sacrum.
Examples of permanent synchondroses are found in the thoracic cage. One example is the first sternocostal joint, where the first rib is anchored to the manubrium by its costal cartilage. (The articulations of the remaining costal cartilages to the sternum are all synovial joints.) Additional synchondroses are formed where the anterior end of the other 11 ribs is joined to its costal cartilage. Unlike the temporary synchondroses of the epiphyseal plate, these permanent synchondroses retain their hyaline cartilage and thus do not ossify with age. Due to the lack of movement between the bone and cartilage, both temporary and permanent synchondroses are functionally classified as a synarthrosis.
Symphysis
A cartilaginous joint where the bones are joined by fibrocartilage is called a symphysis (“growing together”). Fibrocartilage is extraordinarily strong because it contains numerous bundles of thick collagen fibres, thus giving it a much greater ability to resist pulling and bending forces when compared with hyaline cartilage. This gives symphyses the ability to strongly unite the adjacent bones but can still allow for limited movement to occur. Thus, a symphysis is functionally classified as an amphiarthrosis.
The gap separating the bones at a symphysis may be narrow or wide. Examples in which the gap between the bones is narrow include the pubic symphysis and the manubriosternal joint. At the pubic symphysis, the pubic portions of the right and left hip bones of the pelvis are joined together by fibrocartilage across a narrow gap. Similarly, at the manubriosternal joint, fibrocartilage unites the manubrium and body portions of the sternum.
The intervertebral symphysis is a wide symphysis located between the bodies of adjacent vertebrae of the vertebral column. Here a thick pad of fibrocartilage called an intervertebral disc strongly unites the adjacent vertebrae by filling the gap between them. The width of the intervertebral symphysis is important because it allows for small movements between the adjacent vertebrae. In addition, the thick intervertebral disc provides cushioning between the vertebrae, which is important when carrying heavy objects or during high-impact activities such as running or jumping.
Section Review
There are two types of cartilaginous joints. A synchondrosis is formed when the adjacent bones are united by hyaline cartilage. A temporary synchondrosis is formed by the epiphyseal plate of a growing long bone, which is lost when the epiphyseal plate ossifies as the bone reaches maturity. The synchondrosis is thus replaced by a synostosis. Permanent synchondroses that do not ossify are found at the first sternocostal joint and between the anterior ends of the bony ribs and the junction with their costal cartilage. A symphysis is where the bones are joined by fibrocartilage and the gap between the bones may be narrow or wide. A narrow symphysis is found at the manubriosternal joint and at the pubic symphysis. A wide symphysis is the intervertebral symphysis in which the bodies of adjacent vertebrae are united by an intervertebral disc.
Describe the structural features of a synovial joint
Discuss the function of additional structures associated with synovial joints
List the six types of synovial joints and give an example of each
Synovial joints are the most common type of joint in the body (Figure 11.9.1). A key structural characteristic for a synovial joint that is not seen at fibrous or cartilaginous joints is the presence of a joint cavity. This fluid-filled space is the site at which the articulating surfaces of the bones contact each other. Unlike fibrous or cartilaginous joints, the articulating bone surfaces at a synovial joint are not directly connected to each other with fibrous connective tissue or cartilage. This gives the bones of a synovial joint the ability to move smoothly against each other, allowing for increased joint mobility.
Figure 11.9.1. Synovial joints. Synovial joints allow for smooth movements between the adjacent bones. The joint is surrounded by an articular capsule that defines a joint cavity filled with synovial fluid. The articulating surfaces of the bones are covered by a thin layer of articular cartilage. Ligaments support the joint by holding the bones together and resisting excess or abnormal joint motions.
Structural Features of Synovial Joints
Synovial joints are characterised by the presence of a joint cavity. The walls of this space are formed by the articular capsule, a fibrous connective tissue structure that is attached to each bone just outside the area of the bone’s articulating surface. The bones of the joint articulate with each other within the joint cavity.
Friction between the bones at a synovial joint is prevented by the presence of the articular cartilage, a thin layer of hyaline cartilage that covers the entire articulating surface of each bone. However, unlike at a cartilaginous joint, the articular cartilages of each bone are not continuous with each other. Instead, the articular cartilage acts like a Teflon® coating over the bone surface, allowing the articulating bones to move smoothly against each other without damaging the underlying bone tissue. Lining the inner surface of the articular capsule is a thin synovial membrane. The cells of this membrane secrete synovial fluid (synovia = “a thick fluid”), a thick, slimy fluid that provides lubrication to further reduce friction between the bones of the joint. This fluid also provides nourishment to the articular cartilage, which does not contain blood vessels. The ability of the bones to move smoothly against each other within the joint cavity, and the freedom of joint movement this provides, means that each synovial joint is functionally classified as a diarthrosis.
Outside of their articulating surfaces, the bones are connected together by ligaments, which are strong bands of fibrous connective tissue. These strengthen and support the joint by anchoring the bones together and preventing their separation. Ligaments allow for normal movements at a joint, but limit the range of these motions, thus preventing excessive or abnormal joint movements. Ligaments are classified based on their relationship to the fibrous articular capsule. An extrinsic ligament is located outside of the articular capsule, an intrinsic ligament is fused to or incorporated into the wall of the articular capsule, and an intracapsular ligament is located inside of the articular capsule.
At many synovial joints, additional support is provided by the muscles and their tendons that act across the joint. A tendon is the dense connective tissue structure that attaches a muscle to bone. As forces acting on a joint increase, the body will automatically increase the overall strength of contraction of the muscles crossing that joint, thus allowing the muscle and its tendon to serve as a “dynamic ligament” to resist forces and support the joint. This type of indirect support by muscles is particularly important at the shoulder joint, for example, where the ligaments are weak.
Additional Structures Associated with Synovial Joints
A few synovial joints of the body have a fibrocartilage structure located between the articulating bones. This is called an articular disc, which is small and oval-shaped, or a meniscus, which is larger and C-shaped. These structures can serve several functions, depending on the specific joint. In some places, an articular disc may act to strongly unite the bones of the joint to each other. Examples of this include the articular discs found at the sternoclavicular joint or between the distal ends of the radius and ulna bones. At other synovial joints, the disc can provide shock absorption and cushioning between the bones, which is the function of each meniscus within the knee joint. Finally, an articular disc can serve to smooth the movements between the articulating bones, as seen at the temporomandibular joint. Some synovial joints also have a fat pad, which can serve as a cushion between the bones.
Additional structures located outside of a synovial joint serve to prevent friction between the bones of the joint and the overlying muscle tendons or skin. A bursa (plural = bursae) is a thin connective tissue sac filled with lubricating liquid. They are located in regions where skin, ligaments, muscles, or muscle tendons can rub against each other, usually near a body joint (Figure 11.9.2). Bursae reduce friction by separating the adjacent structures, preventing them from rubbing directly against each other. Bursae are classified by their location. A subcutaneous bursa is located between the skin and an underlying bone. It allows skin to move smoothly over the bone. Examples include the prepatellar bursa located over the kneecap and the olecranon bursa at the tip of the elbow. A submuscular bursa is found between a muscle and an underlying bone, or between adjacent muscles. These prevent rubbing of the muscle during movements. A large submuscular bursa, the trochanteric bursa, is found at the lateral hip, between the greater trochanter of the femur and the overlying gluteus maximus muscle. A subtendinous bursa is found between a tendon and a bone. Examples include the subacromial bursa that protects the tendon of shoulder muscle as it passes under the acromion of the scapula, and the suprapatellar bursa that separates the tendon of the large anterior thigh muscle from the distal femur just above the knee.
Figure 11.9.2. Bursae. Bursae are fluid-filled sacs that serve to prevent friction between skin, muscle, or tendon and an underlying bone. Three major bursae and a fat pad are part of the complex joint that unites the femur and tibia of the leg.
A tendon sheath is similar in structure to a bursa, but smaller. It is a connective tissue sac that surrounds a muscle tendon at places where the tendon crosses a joint. It contains a lubricating fluid that allows for smooth motions of the tendon during muscle contraction and joint movements.
Homeostatic Imbalances
Bursitis
Bursitis is the inflammation of a bursa near a joint. This will cause pain, swelling, or tenderness of the bursa and surrounding area, and may also result in joint stiffness. Bursitis is most associated with the bursae found at or near the shoulder, hip, knee, or elbow joints. At the shoulder, subacromial bursitis may occur in the bursa that separates the acromion of the scapula from the tendon of a shoulder muscle as it passes deep to the acromion. In the hip region, trochanteric bursitis can occur in the bursa that overlies the greater trochanter of the femur, just below the lateral side of the hip. Ischial bursitis occurs in the bursa that separates the skin from the ischial tuberosity of the pelvis, the bony structure that is weight bearing when sitting. At the knee, inflammation and swelling of the bursa located between the skin and patella bone is prepatellar bursitis (“housemaid’s knee”), a condition more commonly seen today in roofers or floor and carpet installers who do not use knee pads. At the elbow, olecranon bursitis is inflammation of the bursa between the skin and olecranon process of the ulna. The olecranon forms the bony tip of the elbow, and bursitis here is also known as “student’s elbow.”
Bursitis can be either acute (lasting only a few days) or chronic. It can arise from muscle overuse, trauma, excessive or prolonged pressure on the skin, rheumatoid arthritis, gout, or infection of the joint. Repeated acute episodes of bursitis can result in a chronic condition. Treatments for the disorder include antibiotics if the bursitis is caused by an infection, or anti-inflammatory agents, such as nonsteroidal anti-inflammatory drugs (NSAIDs) or corticosteroids if the bursitis is due to trauma or overuse. Chronic bursitis may require that fluid be drained, but additional surgery is usually not required.
Types of Synovial Joints
Synovial joints are subdivided based on the shapes of the articulating surfaces of the bones that form each joint. The six types of synovial joints are pivot, hinge, condyloid, saddle, plane, and ball-and socket-joints (Figure 11.9.3).
Figure 11.9.3. Types of synovial joints. The six types of synovial joints allow the body to move in a variety of ways. (a) Pivot joints allow for rotation around an axis, such as between the first and second cervical vertebrae, which allows for side-to-side rotation of the head. (b) The hinge joint of the elbow works like a door hinge. (c) The articulation between the trapezium carpal bone and the first metacarpal bone at the base of the thumb is a saddle joint. (d) Plane joints, such as those between the tarsal bones of the foot, allow for limited gliding movements between bones. (e) The radiocarpal joint of the wrist is a condyloid joint. (f) The hip and shoulder joints are the only ball-and-socket joints of the body.
Pivot Point
At a pivot joint, a rounded portion of a bone is enclosed within a ring formed partially by the articulation with another bone and partially by a ligament (see Figure 11.9.3a). The bone rotates within this ring. Since the rotation is around a single axis, pivot joints are functionally classified as a uniaxial diarthrosis type of joint. An example of a pivot joint is the atlantoaxial joint, found between the C1 (atlas) and C2 (axis) vertebrae. Here, the upward projecting dens of the axis articulates with the inner aspect of the atlas, where it is held in place by a ligament. Rotation at this joint allows you to turn your head from side to side. A second pivot joint is found at the proximal radioulnar joint. Here, the head of the radius is encircled by a ligament that holds it in place as it articulates with the radial notch of the ulna. Rotation of the radius allows for forearm movements.
Hinge Joint
In a hinge joint, the convex end of one bone articulates with the concave end of the adjoining bone (see Figure 11.9.3b). This type of joint allows only for bending and straightening motions along a single axis, and thus hinge joints are functionally classified as uniaxial joints. A good example is the elbow joint, with the articulation between the trochlea of the humerus and the trochlear notch of the ulna. Other hinge joints of the body include the knee, ankle, and interphalangeal joints between the phalanx bones of the fingers and toes.
Condyloid Joint
At a condyloid joint (ellipsoid joint), the shallow depression at the end of one bone articulates with a rounded structure from an adjacent bone or bones (see Figure 11.9.3e). The knuckle (metacarpophalangeal) joints of the hand between the distal end of a metacarpal bone and the proximal phalanx bone are condyloid joints. Another example is the radiocarpal joint of the wrist, between the shallow depression at the distal end of the radius bone and the rounded scaphoid, lunate, and triquetrum carpal bones. In this case, the articulation area has a more oval (elliptical) shape. Functionally, condyloid joints are biaxial joints that allow for two planes of movement. One movement involves the bending and straightening of the fingers or the anterior-posterior movements of the hand. The second movement is a side-to-side movement, which allows you to spread your fingers apart and bring them together, or to move your hand in a medial-going or lateral-going direction.
Saddle Joint
At a saddle joint, both of the articulating surfaces for the bones have a saddle shape, which is concave in one direction and convex in the other (see Figure 11.9.3c). This allows the two bones to fit together like a rider sitting on a saddle. Saddle joints are functionally classified as biaxial joints. The primary example is the first carpometacarpal joint, between the trapezium (a carpal bone) and the first metacarpal bone at the base of the thumb. This joint provides the thumb the ability to move away from the palm of the hand along two planes. Thus, the thumb can move within the same plane as the palm of the hand, or it can jut out anteriorly, perpendicular to the palm. This movement of the first carpometacarpal joint is what gives humans their distinctive “opposable” thumbs. The sternoclavicular joint is also classified as a saddle joint.
Plane Joint
At a plane joint (gliding joint), the articulating surfaces of the bones are flat or slightly curved and of approximately the same size, which allows the bones to slide against each other (see Figure 11.9.3d). The motion at this type of joint is usually small and tightly constrained by surrounding ligaments. Based only on their shape, plane joints can allow multiple movements, including rotation. Thus, plane joints can be functionally classified as a multiaxial joint. However, not all these movements are available to every plane joint due to limitations placed on it by ligaments or neighbouring bones. Thus, depending upon the specific joint of the body, a plane joint may exhibit only a single type of movement or several movements. Plane joints are found between the carpal bones (intercarpal joints) of the wrist or tarsal bones (intertarsal joints) of the foot, between the clavicle and acromion of the scapula (acromioclavicular joint), and between the superior and inferior articular processes of adjacent vertebrae (zygapophysial joints).
Ball-and-Socket Joint
The joint with the greatest range of motion is the ball-and-socket joint. At these joints, the rounded head of one bone (the ball) fits into the concave articulation (the socket) of the adjacent bone (see Figure 11.9.3f). The hip joint and the glenohumeral (shoulder) joint are the only ball-and-socket joints of the body. At the hip joint, the head of the femur articulates with the acetabulum of the hip bone, and at the shoulder joint, the head of the humerus articulates with the glenoid cavity of the scapula.
Ball-and-socket joints are classified functionally as multiaxial joints. The femur and the humerus are able to move in both anterior-posterior and medial-lateral directions and they can also rotate around their long axis. The shallow socket formed by the glenoid cavity allows the shoulder joint an extensive range of motion. In contrast, the deep socket of the acetabulum and the strong supporting ligaments of the hip joint serve to constrain movements of the femur, reflecting the need for stability and weight-bearing ability at the hip.
Ageing and the Joints
Arthritis is a common disorder of synovial joints that involves inflammation of the joint. This often results in significant joint pain, along with swelling, stiffness and reduced joint mobility. There are more than 100 different forms of arthritis. Arthritis may arise from ageing, damage to the articular cartilage, autoimmune diseases, bacterial or viral infections, or unknown (probably genetic) causes.
The most common type of arthritis is osteoarthritis, which is associated with ageing and “wear and tear” of the articular cartilage (Figure 11.9.4). Risk factors that may lead to osteoarthritis later in life include injury to a joint; jobs that involve physical labour; sports with running, twisting, or throwing actions; and being overweight. These factors put stress on the articular cartilage that covers the surfaces of bones at synovial joints, causing the cartilage to gradually become thinner. As the articular cartilage layer wears down, more pressure is placed on the bones. The joint responds by increasing production of the lubricating synovial fluid, but this can lead to swelling of the joint cavity, causing pain and joint stiffness as the articular capsule is stretched. The bone tissue underlying the damaged articular cartilage also responds by thickening, producing irregularities and causing the articulating surface of the bone to become rough or bumpy. Joint movement then results in pain and inflammation. In its initial stages, symptoms of osteoarthritis may be reduced by mild activity that “warms up” the joint, but the symptoms may worsen following exercise. In individuals with more advanced osteoarthritis, the affected joints can become more painful and therefore are difficult to use effectively, resulting in increased immobility. There is no cure for osteoarthritis, but several treatments can help alleviate the pain. Treatments may include lifestyle changes, such as weight loss and low-impact exercise, and over-the-counter or prescription medications that help to alleviate the pain and inflammation. For severe cases, joint replacement surgery (arthroplasty) may be required.
Joint replacement is a very invasive procedure, so other treatments are always tried before surgery. However, arthroplasty can provide relief from chronic pain and can enhance mobility within a few months following the surgery. This type of surgery involves replacing the articular surfaces of the bones with prosthesis (artificial components). For example, in hip arthroplasty, the worn or damaged parts of the hip joint, including the head and neck of the femur and the acetabulum of the pelvis, are removed, and replaced with artificial joint components. The replacement head for the femur consists of a rounded ball attached to the end of a shaft that is inserted inside the diaphysis of the femur. The acetabulum of the pelvis is reshaped, and a replacement socket is fitted into its place. The parts, which are always built-in advance of the surgery, are sometimes custom made to produce the best possible fit for a patient.
Gout is a form of arthritis that results from the deposition of uric acid crystals within a body joint. Usually only one or a few joints are affected, such as the big toe, knee, or ankle. The attack may only last a few days but may return to the same or another joint. Gout occurs when the body makes too much uric acid or the kidneys do not properly excrete it. A diet with excessive fructose has been implicated in raising the chances of a susceptible individual developing gout.
Other forms of arthritis are associated with various autoimmune diseases, bacterial infections of the joint, or unknown genetic causes. Autoimmune diseases, including rheumatoid arthritis, scleroderma, or systemic lupus erythematosus, produce arthritis because the immune system of the body attacks the body joints. In rheumatoid arthritis, the joint capsule and synovial membrane become inflamed. As the disease progresses, the articular cartilage is severely damaged or destroyed, resulting in joint deformation, loss of movement, and severe disability. The most involved joints are the hands, feet, and cervical spine, with corresponding joints on both sides of the body usually affected, though not always to the same extent. Rheumatoid arthritis is also associated with lung fibrosis, vasculitis (inflammation of blood vessels), coronary heart disease, and premature mortality. With no known cure, treatments are aimed at alleviating symptoms. Exercise, anti-inflammatory and pain medications, various specific disease-modifying anti-rheumatic drugs, or surgery are used to treat rheumatoid arthritis.
Figure 11.9.4. Osteoarthritis. Osteoarthritis of a synovial joint results from ageing or prolonged joint wear and tear. These cause erosion and loss of the articular cartilage covering the surfaces of the bones, resulting in inflammation that causes joint stiffness and pain.
Section Review
Synovial joints are the most common type of joints in the body. They are characterised by the presence of a joint cavity, inside of which the bones of the joint articulate with each other. The articulating surfaces of the bones at a synovial joint are not directly connected to each other by connective tissue or cartilage, which allows the bones to move freely against each other. The walls of the joint cavity are formed by the articular capsule. Friction between the bones is reduced by a thin layer of articular cartilage covering the surfaces of the bones, and by a lubricating synovial fluid, which is secreted by the synovial membrane.
Synovial joints are strengthened by the presence of ligaments, which hold the bones together and resist excessive or abnormal movements of the joint. Ligaments are classified as extrinsic ligaments if they are located outside of the articular capsule, intrinsic ligaments if they are fused to the wall of the articular capsule, or intracapsular ligaments if they are located inside the articular capsule. Some synovial joints also have an articular disc (meniscus), which can provide padding between the bones, smooth their movements, or strongly join the bones together to strengthen the joint. Muscles and their tendons acting across a joint can also increase their contractile strength when needed, thus providing indirect support for the joint.
Bursae contain a lubricating fluid that serves to reduce friction between structures. Subcutaneous bursae prevent friction between the skin and an underlying bone, submuscular bursae protect muscles from rubbing against a bone or another muscle, and a subtendinous bursa prevents friction between bone and a muscle tendon. Tendon sheaths contain a lubricating fluid and surround tendons to allow for smooth movement of the tendon as it crosses a joint.
Based on the shape of the articulating bone surfaces and the types of movement allowed, synovial joints are classified into six types. At a pivot joint, one bone is held within a ring by a ligament and its articulation with a second bone. Pivot joints only allow for rotation around a single axis. These are found at the articulation between the C1 (atlas) and the dens of the C2 (axis) vertebrae, which provides the side-to-side rotation of the head, or at the proximal radioulnar joint between the head of the radius and the radial notch of the ulna, which allows for rotation of the radius during forearm movements. Hinge joints, such as at the elbow, knee, ankle, or interphalangeal joints between phalanx bones of the fingers and toes, allow only for bending and straightening of the joint. Pivot and hinge joints are functionally classified as uniaxial joints.
Condyloid joints are found where the shallow depression of one bone receives a rounded bony area formed by one or two bones. Condyloid joints are found at the base of the fingers (metacarpophalangeal joints) and at the wrist (radiocarpal joint). At a saddle joint, the articulating bones fit together like a rider and a saddle. An example is the first carpometacarpal joint located at the base of the thumb. Both condyloid and saddle joints are functionally classified as biaxial joints.
Plane joints are formed between the small, flattened surfaces of adjacent bones. These joints allow the bones to slide or rotate against each other, but the range of motion is usually slight and tightly limited by ligaments or surrounding bones. This type of joint is found between the articular processes of adjacent vertebrae, at the acromioclavicular joint, or at the intercarpal joints of the hand and intertarsal joints of the foot. Ball-and-socket joints, in which the rounded head of a bone fits into a large depression or socket, are found at the shoulder and hip joints. Both plane and ball-and-sockets joints are classified functionally as multiaxial joints. However, ball-and-socket joints allow for large movements, while the motions between bones at a plane joint are small.
Synovial joints allow the body a tremendous range of movements. Each movement at a synovial joint result from the contraction or relaxation of the muscles that are attached to the bones on either side of the articulation. The type of movement that can be produced at a synovial joint is determined by its structural type. While the ball-and-socket joint gives the greatest range of movement at an individual joint, in other regions of the body, several joints may work together to produce a particular movement. Overall, each type of synovial joint is necessary to provide the body with its great flexibility and mobility. There are many types of movement that can occur at synovial joints (Table 11.10.1). Movement types are paired, with one being the opposite of the other. Body movements are always described in relation to the anatomical position of the body: upright stance, with upper limbs to the side of body and palms facing forward. Refer to Figure 11.10.1 as you go through this section.
Figure 11.10.1. Movements of the body, part 1. Synovial joints give the body many ways in which to move. (a)–(b) Flexion and extension motions are in the sagittal (anterior–posterior) plane of motion. These movements take place at the shoulder, hip, elbow, knee, wrist, metacarpophalangeal, metatarsophalangeal, and interphalangeal joints. (c)–(d) Anterior bending of the head or vertebral column is flexion, while any posterior-going movement is extension. (e) Abduction and adduction are motions of the limbs, hand, fingers, or toes in the coronal (medial–lateral) plane of movement. Moving the limb or hand laterally away from the body, or spreading the fingers or toes, is abduction. Adduction brings the limb or hand toward or across the midline of the body or brings the fingers or toes together. Circumduction is the movement of the limb, hand, or fingers in a circular pattern, using the sequential combination of flexion, adduction, extension, and abduction motions. Adduction/abduction and circumduction take place at the shoulder, hip, wrist, metacarpophalangeal, and metatarsophalangeal joints. (f) Turning of the head side to side or twisting of the body is rotation. Medial and lateral rotation of the upper limb at the shoulder or lower limb at the hip involves turning the anterior surface of the limb toward the midline of the body (medial or internal rotation) or away from the midline (lateral or external rotation).Figure 11.10.2. Movements of the body, part 2. (g) Supination of the forearm turns the hand to the palm forward position in which the radius and ulna are parallel, while forearm pronation turns the hand to the palm backward position in which the radius crosses over the ulna to form an “X.” (h) Dorsiflexion of the foot at the ankle joint moves the top of the foot toward the leg, while plantar flexion lifts the heel and points the toes. (i) Eversion of the foot moves the bottom (sole) of the foot away from the midline of the body, while foot inversion faces the sole toward the midline. (j) Protraction of the mandible pushes the chin forward, and retraction pulls the chin back. (k) Depression of the mandible opens the mouth, while elevation closes it. (l) Opposition of the thumb brings the tip of the thumb into contact with the tip of the fingers of the same hand and reposition brings the thumb back next to the index finger.
Flexion and Extension
Flexion and extension are movements that take place within the sagittal plane and involve anterior or posterior movements of the body or limbs. For the vertebral column, flexion (anterior flexion) is an anterior (forward) bending of the neck or body, while extension involves a posterior-directed motion, such as straightening from a flexed position or bending backward. Lateral flexion is the bending of the neck or body toward the right or left side. These movements of the vertebral column involve both the symphysis joint formed by each intervertebral disc, as well as the plane type of synovial joint formed between the inferior articular processes of one vertebra and the superior articular processes of the next lower vertebra.
In the limbs, flexion decreases the angle between the bones (bending of the joint), while extension increases the angle and straightens the joint. For the upper limb, all anterior-going motions are flexion and all posterior-going motions are extension. These include anterior-posterior movements of the arm at the shoulder, the forearm at the elbow, the hand at the wrist, and the fingers at the metacarpophalangeal and interphalangeal joints. For the thumb, extension moves the thumb away from the palm of the hand, within the same plane as the palm, while flexion brings the thumb back against the index finger or into the palm. These motions take place at the first carpometacarpal joint. In the lower limb, bringing the thigh forward and upward is flexion at the hip joint, while any posterior-going motion of the thigh is extension. Note that extension of the thigh beyond the anatomical (standing) position is limited by the ligaments that support the hip joint. Knee flexion is the bending of the knee to bring the foot toward the posterior thigh, and extension is the straightening of the knee. Flexion and extension movements are seen at the hinge, condyloid, saddle, and ball-and-socket joints of the limbs (see Figure 11.10.1a-d).
Hyperextension is the abnormal or excessive extension of a joint beyond its normal range of motion, thus resulting in injury. Similarly, hyperflexion is excessive flexion at a joint. Hyperextension injuries are common at hinge joints such as the knee or elbow. In cases of “whiplash” in which the head is suddenly moved backward and then forward, a patient may experience both hyperextension and hyperflexion of the cervical region.
Abduction and Adduction
Abduction and adduction motions occur within the coronal plane and involve medial-lateral motions of the limbs, fingers, toes, or thumb. Abduction moves the limb laterally away from the midline of the body, while adduction is the opposing movement that brings the limb toward the body or across the midline. For example, abduction is raising the arm at the shoulder joint, moving it laterally away from the body, while adduction brings the arm down to the side of the body. Similarly, abduction and adduction at the wrist moves the hand away from or toward the midline of the body. Spreading the fingers or toes apart is also abduction, while bringing the fingers or toes together is adduction. For the thumb, abduction is the anterior movement that brings the thumb to a 90° perpendicular position, pointing straight out from the palm. Adduction moves the thumb back to the anatomical position, next to the index finger. Abduction and adduction movements are seen at condyloid, saddle, and ball-and-socket joints (see Figure 11.10.1e).
Circumduction
Circumduction is the movement of a body region in a circular manner, in which one end of the body region being moved stays stationary while the other end describes a circle. It involves the sequential combination of flexion, adduction, extension, and abduction at a joint. This type of motion is found at biaxial condyloid and saddle joints, and at multiaxial ball-and-sockets joints (see Figure 11.10.1e).
Rotation
Rotation can occur within the vertebral column, at a pivot joint, or at a ball-and-socket joint. Rotation of the neck or body is the twisting movement produced by the summation of the small rotational movements available between adjacent vertebrae. At a pivot joint, one bone rotates in relation to another bone. This is a uniaxial joint, and thus rotation is the only motion allowed at a pivot joint. For example, at the atlantoaxial joint, the first cervical (C1) vertebra (atlas) rotates around the dens, the upward projection from the second cervical (C2) vertebra (axis). This allows the head to rotate from side to side as when shaking the head “no.” The proximal radioulnar joint is a pivot joint formed by the head of the radius and its articulation with the ulna. This joint allows for the radius to rotate along its length during pronation and supination movements of the forearm.
Rotation can also occur at the ball-and-socket joints of the shoulder and hip. Here, the humerus and femur rotate around their long axis, which moves the anterior surface of the arm or thigh either toward or away from the midline of the body. Movement that brings the anterior surface of the limb toward the midline of the body is called medial (internal) rotation. Conversely, rotation of the limb so that the anterior surface moves away from the midline is lateral (external) rotation (see Figure 11.10.1f). Be sure to distinguish medial and lateral rotation, which can only occur at the multiaxial shoulder and hip joints, from circumduction, which can occur at either biaxial or multiaxial joints.
Supination and Pronation
Supination and pronation are movements of the forearm. In the anatomical position, the upper limb is held next to the body with the palm facing forward. This is the supinated position of the forearm. In this position, the radius and ulna are parallel to each other. When the palm of the hand faces backward, the forearm is in the pronated position, and the radius and ulna form an X-shape.
Supination and pronation are the movements of the forearm that go between these two positions. Pronation is the motion that moves the forearm from the supinated (anatomical) position to the pronated (palm backward) position. This motion is produced by rotation of the radius at the proximal radioulnar joint, accompanied by movement of the radius at the distal radioulnar joint. The proximal radioulnar joint is a pivot joint that allows for rotation of the head of the radius. Because of the slight curvature of the shaft of the radius, this rotation causes the distal end of the radius to cross over the distal ulna at the distal radioulnar joint. This crossing over brings the radius and ulna into an X-shape position. Supination is the opposite motion, in which rotation of the radius returns the bones to their parallel positions and moves the palm to the anterior facing (supinated) position. It helps to remember that supination is the motion you use when scooping up soup with a spoon (see Figure 11.10.2g).
Dorsiflexion and Plantar Flexion
Dorsiflexion and plantar flexion are movements at the ankle joint, which is a hinge joint. Lifting the front of the foot, so that the top of the foot moves toward the anterior leg is dorsiflexion, while lifting the heel of the foot from the ground or pointing the toes downward is plantar flexion. These are the only movements available at the ankle joint (see Figure 11.10.2h).
Inversion and Eversion
Inversion and eversion are complex movements that involve the multiple plane joints among the tarsal bones of the posterior foot (intertarsal joints) and thus are not motions that take place at the ankle joint. Inversion is the turning of the foot to angle the bottom of the foot toward the midline, while eversion turns the bottom of the foot away from the midline. The foot has a greater range of inversion than eversion motion. These are important motions that help to stabilise the foot when walking or running on an uneven surface and aid in the quick side-to-side changes in direction used during active sports such as basketball, racquetball, or soccer (see Figure 11.10.2i).
Protraction and Reaction
Protraction and retraction are anterior-posterior movements of the scapula or mandible. Protraction of the scapula occurs when the shoulder is moved forward, as when pushing against something or throwing a ball. Retraction is the opposite motion, with the scapula being pulled posteriorly and medially, toward the vertebral column. For the mandible, protraction occurs when the lower jaw is pushed forward, to stick out the chin, while retraction pulls the lower jaw backward. (See Figure 11.10.2j.)
Depression and Elevation
Depression and elevation are downward and upward movements of the scapula or mandible. The upward movement of the scapula and shoulder is elevation, while a downward movement is depression. These movements are used to shrug your shoulders. Similarly, elevation of the mandible is the upward movement of the lower jaw used to close the mouth or bite on something, and depression is the downward movement that produces opening of the mouth (see Figure 11.10.2k).
Excursion
Excursion is the side-to-side movement of the mandible. Lateral excursion moves the mandible away from the midline, toward either the right or left side. Medial excursion returns the mandible to its resting position at the midline.
Superior Rotation and Inferior Rotation
Superior and inferior rotation are movements of the scapula and are defined by the direction of movement of the glenoid cavity. These motions involve rotation of the scapula around a point inferior to the scapular spine and are produced by combinations of muscles acting on the scapula. During superior rotation, the glenoid cavity moves upward as the medial end of the scapular spine moves downward. This is a particularly important motion that contributes to upper limb abduction. Without superior rotation of the scapula, the greater tubercle of the humerus would hit the acromion of the scapula, thus preventing any abduction of the arm above shoulder height. Superior rotation of the scapula is thus required for full abduction of the upper limb. Superior rotation is also used without arm abduction when carrying a heavy load with your hand or on your shoulder. You can feel this rotation when you pick up a load, such as a heavy book bag and carry it on only one shoulder. To increase its weight-bearing support for the bag, the shoulder lifts as the scapula superiorly rotates. Inferior rotation occurs during limb adduction and involves the downward motion of the glenoid cavity with upward movement of the medial end of the scapular spine.
Opposition and Reposition
Opposition is the thumb movement that brings the tip of the thumb in contact with the tip of a finger. This movement is produced at the first carpometacarpal joint, which is a saddle joint formed between the trapezium carpal bone and the first metacarpal bone. Thumb opposition is produced by a combination of flexion and abduction of the thumb at this joint. Returning the thumb to its anatomical position next to the index finger is called reposition (see Figure 11.10.2l).
Knee; elbow; ankle; interphalangeal joints of fingers and toes
Condyloid
Biaxial joint; allows flexion/extension, abduction/adduction, and circumduction movements
Metacarpophalangeal (knuckle) joints of fingers; radiocarpal joint of wrists; metatarsophalangeal joints for toes
Saddle
Biaxial joint; allows flexion/extension, abduction/adduction, and circumduction movements
First carpometacarpal joint of the thumb; sternoclavicular joint
Plane
Multiaxial joint; allows inversion and eversion of foot, or flexion, extension, and lateral flexion of the vertebral column
Intertarsal joints of foot; superior-inferior articular process articulations between vertebrae
Ball-and-socket
Multiaxial joint; allows flexion/extension, abduction/adduction, circumduction, and medial/lateral rotation movements
Shoulder and hip joints
Section Review
The variety of movements provided by the several types of synovial joints allows for a broad range of body motions and gives you tremendous mobility. These movements allow you to flex or extend your body or limbs, medially rotate, and adduct your arms and flex your elbows to hold a heavy object against your chest, raise your arms above your head, rotate or shake your head, and bend to touch the toes (with or without bending your knees).
Each of the different structural types of synovial joints also allow for specific motions. The atlantoaxial pivot joint provides side-to-side rotation of the head, while the proximal radioulnar articulation allows for rotation of the radius during pronation and supination of the forearm. Hinge joints, such as at the knee and elbow, allow only for flexion and extension. Similarly, the hinge joint of the ankle only allows for dorsiflexion and plantar flexion of the foot.
Condyloid and saddle joints are biaxial. These allow for flexion and extension, and abduction and adduction. The sequential combination of flexion, adduction, extension, and abduction produces circumduction. Multiaxial plane joints provide for only small motions, but these can add together over several adjacent joints to produce body movement, such as inversion and eversion of the foot. Similarly, plane joints allow for flexion, extension, and lateral flexion movements of the vertebral column. The multiaxial ball and socket joints allow for flexion-extension, abduction-adduction, and circumduction. In addition, these also allow for medial (internal) and lateral (external) rotation. Ball-and-socket joints have the greatest range of motion of all synovial joints.
Describe the bones that articulate together to form selected synovial joints
Discuss the movements available at each joint
Describe the structures that support and prevent excess movements at each joint
Each synovial joint of the body is specialised to perform certain movements. The movements that are allowed are determined by the structural classification for each joint. For example, a multiaxial ball-and-socket joint has much more mobility than a uniaxial hinge joint. However, the ligaments and muscles that support a joint may place restrictions on the total range of motion available. Thus, the ball-and-socket joint of the shoulder has little in the way of ligament support, which gives the shoulder an exceptionally large range of motion. In contrast, movements at the hip joint are restricted by strong ligaments, which reduce its range of motion but confer stability during standing and weight bearing.
This section will examine the anatomy of selected synovial joints of the body. Anatomical names for most joints are derived from the names of the bones that articulate at that joint, although some joints, such as the elbow, hip, and knee joints are exceptions to this general naming scheme.
Articulations of the Vertebral Column
In addition to being held together by the intervertebral discs, adjacent vertebrae also articulate with each other at synovial joints formed between the superior and inferior articular processes called zygapophysial joints (facet joints). These are plane joints that provide for only limited motions between the vertebrae. The orientation of the articular processes at these joints varies in different regions of the vertebral column and serves to determine the types of motions available in each vertebral region. The cervical and lumbar regions have the greatest ranges of motions.
In the neck, the articular processes of cervical vertebrae are flattened and face upward or downward. This orientation provides the cervical vertebral column with extensive ranges of motion for flexion, extension, lateral flexion, and rotation. In the thoracic region, the downward projecting and overlapping spinous processes, along with the attached thoracic cage, limit flexion, extension, and lateral flexion. However, the flattened and vertically positioned thoracic articular processes allow for the greatest range of rotation within the vertebral column. The lumbar region allows for considerable extension, flexion, and lateral flexion, but the orientation of the articular processes prohibits rotation.
The articulations formed between the skull, the atlas (C1 vertebra), and the axis (C2 vertebra) differ from the articulations in other vertebral areas and play important roles in movement of the head. The atlanto-occipital joint is formed by the articulations between the superior articular processes of the atlas and the occipital condyles on the base of the skull. This articulation has a pronounced U-shaped curvature, oriented along the anterior-posterior axis. This allows the skull to rock forward and backward, producing flexion and extension of the head. This moves the head up and down, as when shaking your head “yes.”
The atlantoaxial joint, between the atlas and axis, consists of three articulations. The paired superior articular processes of the axis articulate with the inferior articular processes of the atlas. These articulating surfaces are flat and oriented horizontally. The third articulation is the pivot joint formed between the dens, which projects upward from the body of the axis, and the inner aspect of the anterior arch of the atlas (Figure 11.11.1). A strong ligament passes posterior to the dens to hold it in position against the anterior arch. These articulations allow the atlas to rotate on top of the axis, moving the head toward the right or left, as when shaking your head “no.”
Figure 11.11.1. Atlantoaxial joint. The atlantoaxial joint is a pivot type of joint between the dens portion of the axis (C2 vertebra) and the anterior arch of the atlas (C1 vertebra), with the dens held in place by a ligament.
Temporomandibular Joint
The temporomandibular joint (TMJ) is the joint that allows for opening (mandibular depression) and closing (mandibular elevation) of the mouth, as well as side-to-side and protraction/retraction motions of the lower jaw. This joint involves the articulation between the mandibular fossa and articular tubercle of the temporal bone, with the condyle (head) of the mandible. Located between these bony structures, filling the gap between the skull and mandible, is a flexible articular disc (Figure 11.11.2). This disc serves to smooth the movements between the temporal bone and mandibular condyle.
Movement at the TMJ during opening and closing of the mouth involves both gliding and hinge motions of the mandible. With the mouth closed, the mandibular condyle and articular disc are located within the mandibular fossa of the temporal bone. During opening of the mouth, the mandible hinges downward and at the same time is pulled anteriorly, causing both the condyle and the articular disc to glide forward from the mandibular fossa onto the downward projecting articular tubercle. The net result is a forward and downward motion of the condyle and mandibular depression. The temporomandibular joint is supported by an extrinsic ligament that anchors the mandible to the skull. This ligament spans the distance between the base of the skull and the lingula on the medial side of the mandibular ramus.
Dislocation of the TMJ may occur when opening the mouth too wide (such as when taking a large bite) or following a blow to the jaw, resulting in the mandibular condyle moving beyond (anterior to) the articular tubercle. In this case, the individual would not be able to close his or her mouth. Temporomandibular joint disorder is a painful condition that may arise due to arthritis, wearing of the articular cartilage covering the bony surfaces of the joint, muscle fatigue from overuse or grinding of the teeth, damage to the articular disc within the joint, or jaw injury. Temporomandibular joint disorders can also cause headache, difficulty chewing, or even the inability to move the jaw (lock jaw). Pharmacologic agents for pain or other therapies, including bite guards, are used as treatments.
Figure 11.11.2. Temporomandibular joint. The temporomandibular joint is the articulation between the temporal bone of the skull and the condyle of the mandible, with an articular disc located between these bones. During depression of the mandible (opening of the mouth), the mandibular condyle moves both forward and hinges downward as it travels from the mandibular fossa onto the articular tubercle.
Shoulder Joint
The shoulder joint is called the glenohumeral joint. This is a ball-and-socket joint formed by the articulation between the head of the humerus and the glenoid cavity of the scapula (Figure 11.11.3). This joint has the largest range of motion of any joint in the body. However, this freedom of movement is due to the lack of structural support and thus the enhanced mobility is offset by a loss of stability.
Figure 11.11.3. Glenohumeral joint. The glenohumeral (shoulder) joint is a ball-and-socket joint that provides the widest range of motions. It has a loose articular capsule and is supported by ligaments and the rotator cuff muscles.
The broad range of motions at the shoulder joint is provided by the articulation of the large, rounded humeral head with the small and shallow glenoid cavity, which is only about one third of the size of the humeral head. The socket formed by the glenoid cavity is deepened slightly by a small lip of fibrocartilage called the glenoid labrum, which extends around the outer margin of the cavity. The articular capsule that surrounds the glenohumeral joint is thin and loose to allow for large motions of the upper limb. Some structural support for the joint is provided by thickenings of the articular capsule wall that form weak intrinsic ligaments. These include the coracohumeral ligament, running from the coracoid process of the scapula to the anterior humerus, and three ligaments, each called a glenohumeral ligament, located on the anterior side of the articular capsule. These ligaments help to strengthen the superior and anterior capsule walls.
However, the primary support for the shoulder joint is provided by muscles crossing the joint, particularly the four rotator cuff muscles. These muscles (supraspinatus, infraspinatus, teres minor, and subscapularis) arise from the scapula and attach to the greater or lesser tubercles of the humerus. As these muscles cross the shoulder joint, their tendons encircle the head of the humerus and become fused to the anterior, superior, and posterior walls of the articular capsule. The thickening of the capsule formed by the fusion of these four muscle tendons is called the rotator cuff. Two bursae, the subacromial bursa and the subscapular bursa, help to prevent friction between the rotator cuff muscle tendons and the scapula as these tendons cross the glenohumeral joint. In addition to their individual actions of moving the upper limb, the rotator cuff muscles also serve to hold the head of the humerus in position within the glenoid cavity. By constantly adjusting their strength of contraction to resist forces acting on the shoulder, these muscles serve as “dynamic ligaments” and thus provide the primary structural support for the glenohumeral joint.
Injuries to the shoulder joint are common. Repetitive use of the upper limb, particularly in abduction such as during throwing, swimming, or racquet sports, may lead to acute or chronic inflammation of the bursa or muscle tendons, a tear of the glenoid labrum, or degeneration or tears of the rotator cuff. Because the humeral head is strongly supported by muscles and ligaments around its anterior, superior, and posterior aspects, most dislocations of the humerus occur in an inferior direction. This can occur when force is applied to the humerus when the upper limb is fully abducted, as when diving to catch a baseball and landing on your hand or elbow. Inflammatory responses to any shoulder injury can lead to the formation of scar tissue between the articular capsule and surrounding structures, thus reducing shoulder mobility, a condition called adhesive capsulitis (“frozen shoulder”).
Elbow Joint
The elbow joint is a uniaxial hinge joint formed by the humeroulnar joint, the articulation between the trochlea of the humerus and the trochlear notch of the ulna. Also associated with the elbow are the humeroradial joint and the proximal radioulnar joint. All three of these joints are enclosed within a single articular capsule (Figure 11.11.4).
The articular capsule of the elbow is thin on its anterior and posterior aspects but is thickened along its outside margins by strong intrinsic ligaments. These ligaments prevent side-to-side movements and hyperextension. On the medial side is the triangular ulnar collateral ligament. This arises from the medial epicondyle of the humerus and attaches to the medial side of the proximal ulna. The strongest part of this ligament is the anterior portion, which resists hyperextension of the elbow. The ulnar collateral ligament may be injured by frequent, forceful extensions of the forearm, as is seen in baseball pitchers. Reconstructive surgical repair of this ligament is referred to as Tommy John surgery, named for the former major league pitcher who was the first person to have this treatment.
The lateral side of the elbow is supported by the radial collateral ligament. This arises from the lateral epicondyle of the humerus and then blends into the lateral side of the annular ligament. The annular ligament encircles the head of the radius. This ligament supports the head of the radius as it articulates with the radial notch of the ulna at the proximal radioulnar joint. This is a pivot joint that allows for rotation of the radius during supination and pronation of the forearm.
Figure 11.11.4. Elbow joint. (a) The elbow is a hinge joint that allows only for flexion and extension of the forearm. (b) It is supported by the ulnar and radial collateral ligaments. (c) The annular ligament supports the head of the radius at the proximal radioulnar joint, the pivot joint that allows for rotation of the radius.
Hip Joint
The hip joint is a multiaxial ball-and-socket joint between the head of the femur and the acetabulum of the hip bone (Figure 11.11.5). The hip carries the weight of the body and thus requires strength and stability during standing and walking. For these reasons, its range of motion is more limited than at the shoulder joint.
The acetabulum is the socket portion of the hip joint. This space is deep and has a large articulation area for the femoral head, thus giving stability and weight bearing ability to the joint. The acetabulum is further deepened by the acetabular labrum, a fibrocartilage lip attached to the outer margin of the acetabulum. The surrounding articular capsule is strong, with several thickened areas forming intrinsic ligaments. These ligaments arise from the hip bone, at the margins of the acetabulum, and attach to the femur at the base of the neck. The ligaments are the iliofemoral ligament, pubofemoral ligament, and ischiofemoral ligament, all of which spiral around the head and neck of the femur. The ligaments are tightened by extension at the hip, thus pulling the head of the femur tightly into the acetabulum when in the upright, standing position. Little additional extension of the thigh is permitted beyond this vertical position. These ligaments thus stabilise the hip joint and allow you to maintain an upright standing position with only minimal muscle contraction. Inside of the articular capsule, the ligament of the head of the femur (ligamentum teres) spans between the acetabulum and femoral head. This intracapsular ligament is normally slack and does not provide any significant joint support, but it does provide a pathway for an important artery that supplies the head of the femur.
The hip is prone to osteoarthritis, and thus was the first joint for which a replacement prosthesis was developed. A common injury in elderly individuals, particularly those with weakened bones due to osteoporosis, is a “broken hip,” which is a fracture of the femoral neck. This may result from a fall, or it may cause the fall. This can happen as one lower limb is taking a step and all the body weight is placed on the other limb, causing the femoral neck to break, and producing a fall. Any accompanying disruption of the blood supply to the femoral neck or head can lead to necrosis of these areas, resulting in bone and cartilage death. Femoral fractures usually require surgical treatment, after which the patient will need mobility assistance for a prolonged period, either from family members or in a long-term care facility. Consequentially, the associated health care costs of “broken hips” are substantial. In addition, hip fractures are associated with increased rates of morbidity (incidences of disease) and mortality (death). Surgery for a hip fracture followed by prolonged bed rest may lead to life-threatening complications, including pneumonia, infection of pressure ulcers (bedsores), and thrombophlebitis (deep vein thrombosis; blood clot formation) that can result in a pulmonary embolism (blood clot within the lung).
Figure 11.11.5. Hip joint. (a) The ball-and-socket joint of the hip is a multiaxial joint that provides both stability and a wide range of motion. (b–c) When standing, the supporting ligaments are tight, pulling the head of the femur into the acetabulum.
Knee Joint
The knee joint is the largest joint of the body (Figure 11.11.6). It consists of three articulations. The femoropatellar joint is found between the patella and the distal femur. The medial tibiofemoral joint and lateral tibiofemoral joint are located between the medial and lateral condyles of the femur and the medial and lateral condyles of the tibia. All these articulations are enclosed within a single articular capsule. The knee functions as a hinge joint, allowing flexion and extension of the leg. This action is generated by both rolling and gliding motions of the femur on the tibia. In addition, some rotation of the leg is available when the knee is flexed, but not when extended. The knee is well constructed for weight bearing in its extended position, but is vulnerable to injuries associated with hyperextension, twisting, or blows to the medial or lateral side of the joint, particularly while weight bearing.
At the femoropatellar joint, the patella slides vertically within a groove on the distal femur. The patella is a sesamoid bone incorporated into the tendon of the quadriceps femoris muscle, the large muscle of the anterior thigh. The patella serves to protect the quadriceps tendon from friction against the distal femur. Continuing from the patella to the anterior tibia just below the knee is the patellar ligament. Acting via the patella and patellar ligament, the quadriceps femoris is a powerful muscle that acts to extend the leg at the knee. It also serves as a “dynamic ligament” to provide important support and stabilisation for the knee joint.
The medial and lateral tibiofemoral joints are the articulations between the rounded condyles of the femur and the flat condyles of the tibia. During flexion and extension motions, the condyles of the femur both roll and glide over the surfaces of the tibia. The rolling action produces flexion or extension, while the gliding action serves to maintain the femoral condyles centred over the tibial condyles, thus ensuring maximal bony, weight-bearing support for the femur in all knee positions. As the knee comes into full extension, the femur undergoes a slight medial rotation in relation to tibia. The rotation results because the lateral condyle of the femur is slightly smaller than the medial condyle. Thus, the lateral condyle finishes its rolling motion first, followed by the medial condyle. The resulting small medial rotation of the femur serves to “lock” the knee into its fully extended and most stable position. Flexion of the knee is initiated by a slight lateral rotation of the femur on the tibia, which “unlocks” the knee. This lateral rotation motion is produced by the popliteus muscle of the posterior leg.
Located between the articulating surfaces of the femur and tibia are two articular discs, the medial meniscus and lateral meniscus (see Figure 11.11.6b). Each is a C-shaped fibrocartilage structure that is thin along its inside margin and thick along the outer margin. They are attached to their tibial condyles, but do not attach to the femur. While both menisci are free to move during knee motions, the medial meniscus shows less movement because it is anchored at its outer margin to the articular capsule and tibial collateral ligament. The menisci provide padding between the bones and help to fill the gap between the round femoral condyles and flattened tibial condyles. Some areas of each meniscus lack an arterial blood supply and thus these areas heal poorly if damaged.
The knee joint has multiple ligaments that provide support, particularly in the extended position (see Figure 11.11.6c). Outside of the articular capsule, located at the sides of the knee, are two extrinsic ligaments. The fibular collateral ligament (lateral collateral ligament) is on the lateral side and spans from the lateral epicondyle of the femur to the head of the fibula. The tibial collateral ligament (medial collateral ligament) of the medial knee runs from the medial epicondyle of the femur to the medial tibia. As it crosses the knee, the tibial collateral ligament is attached on its deep side to the articular capsule and to the medial meniscus, a crucial factor when considering knee injuries. In the fully extended knee position, both collateral ligaments are taut (tight), thus serving to stabilise and support the extended knee and preventing side-to-side or rotational motions between the femur and tibia.
The articular capsule of the posterior knee is thickened by intrinsic ligaments that help to resist knee hyperextension. Inside the knee are two intracapsular ligaments, the anterior cruciate ligament and posterior cruciate ligament. These ligaments are anchored inferiorly to the tibia at the intercondylar eminence, the roughened area between the tibial condyles. The cruciate ligaments are named for whether they are attached anteriorly or posteriorly to this tibial region. Each ligament runs diagonally upward to attach to the inner aspect of a femoral condyle. The cruciate ligaments are named for the X-shape formed as they pass each other (cruciate means “cross”). The posterior cruciate ligament is the stronger ligament. It serves to support the knee when it is flexed and weight bearing, as when walking downhill. In this position, the posterior cruciate ligament prevents the femur from sliding anteriorly off the top of the tibia. The anterior cruciate ligament becomes tight when the knee is extended, and thus resists hyperextension.
Figure 11.11.6. Knee joint. (a) The knee joint is the largest joint of the body. (b)–(c) It is supported by the tibial and fibular collateral ligaments located on the sides of the knee outside of the articular capsule, and the anterior and posterior cruciate ligaments found inside the capsule. The medial and lateral menisci provide padding and support between the femoral condyles and tibial condyles.
Disorders of the Joints
Injuries to the knee are common. Since this joint is primarily supported by muscles and ligaments, injuries to any of these structures will result in pain or knee instability. Injury to the posterior cruciate ligament occurs when the knee is flexed and the tibia is driven posteriorly, such as falling and landing on the tibial tuberosity or hitting the tibia on the dashboard when not wearing a seatbelt during an automobile accident. More commonly, injuries occur when forces are applied to the extended knee, particularly when the foot is planted and unable to move. Anterior cruciate ligament injuries can result with a forceful blow to the anterior knee, producing hyperextension, or when a runner makes a quick change of direction that produces both twisting and hyperextension of the knee.
A worse combination of injuries can occur with a hit to the lateral side of the extended knee (Figure 11.11.7). A moderate blow to the lateral knee will cause the medial side of the joint to open, resulting in stretching or damage to the tibial collateral ligament. Because the medial meniscus is attached to the tibial collateral ligament, a stronger blow can tear the ligament and also damage the medial meniscus. This is one reason that the medial meniscus is 20 times more likely to be injured than the lateral meniscus. A powerful blow to the lateral knee produces a “terrible triad” injury, in which there is a sequential injury to the tibial collateral ligament, medial meniscus, and anterior cruciate ligament.
Arthroscopic surgery has improved the surgical treatment of knee injuries and reduced subsequent recovery times. This procedure involves a small incision and the insertion into the joint of an arthroscope, a pencil-thin instrument that allows for visualisation of the joint interior. Small surgical instruments are also inserted via additional incisions. These tools allow a surgeon to remove or repair a torn meniscus or to reconstruct a ruptured cruciate ligament. The current method for anterior cruciate ligament replacement involves using a portion of the patellar ligament. Holes are drilled into the cruciate ligament attachment points on the tibia and femur, and the patellar ligament graft, with small areas of attached bone still intact at each end, is inserted into these holes. The bone-to-bone sites at each end of the graft heal rapidly and strongly, thus enabling a rapid recovery.
Knee Injury
A strong blow to the lateral side of the extended knee will cause three injuries, in sequence: tearing of the tibial collateral ligament, damage to the medial meniscus, and rupture of the anterior cruciate ligament.
Figure 11.11.7. Knee injury. A strong blow to the lateral side of the extended knee will cause three injuries, in sequence: tearing of the tibial collateral ligament, damage to the medial meniscus, and rupture of the anterior cruciate ligament.
Ankle and Foot Joints
The ankle is formed by the talocrural joint (Figure 11.11.8). It consists of the articulations between the talus bone of the foot and the distal ends of the tibia and fibula of the leg (crural = “leg”). The superior aspect of the talus bone is square-shaped and has three areas of articulation. The top of the talus articulates with the inferior tibia. This is the portion of the ankle joint that carries the body weight between the leg and foot. The sides of the talus are firmly held in position by the articulations with the medial malleolus of the tibia and the lateral malleolus of the fibula, which prevent any side-to-side motion of the talus. The ankle is thus a uniaxial hinge joint that allows only for dorsiflexion and plantar flexion of the foot.
Additional joints between the tarsal bones of the posterior foot allow for the movements of foot inversion and eversion. Most important for these movements is the subtalar joint, located between the talus and calcaneus bones. The joints between the talus and navicular bones and the calcaneus and cuboid bones are also important contributors to these movements. All the joints between tarsal bones are plane joints. Together, the small motions that take place at these joints all contribute to the production of inversion and eversion foot motions.
Like the hinge joints of the elbow and knee, the talocrural joint of the ankle is supported by several strong ligaments located on the sides of the joint. These ligaments extend from the medial malleolus of the tibia or lateral malleolus of the fibula and anchor to the talus and calcaneus bones. Since they are located on the sides of the ankle joint, they allow for dorsiflexion and plantar flexion of the foot. They also prevent abnormal side-to-side and twisting movements of the talus and calcaneus bones during eversion and inversion of the foot. On the medial side is the broad deltoid ligament. The deltoid ligament supports the ankle joint and resists excessive eversion of the foot. The lateral side of the ankle has several smaller ligaments. These include the anterior talofibular ligament and the posterior talofibular ligament, both of which span between the talus bone and the lateral malleolus of the fibula, and the calcaneofibular ligament, located between the calcaneus bone and fibula. These ligaments support the ankle and resist excess inversion of the foot.
Figure 11.11.8. Ankle joint. The talocrural (ankle) joint is a uniaxial hinge joint that only allows for dorsiflexion or plantar flexion of the foot. Movements at the subtalar joint, between the talus and calcaneus bones, combined with motions at other intertarsal joints, enables eversion/inversion movements of the foot. Ligaments that unite the medial or lateral malleolus with the talus and calcaneus bones serve to support the talocrural joint and to resist excess eversion or inversion of the foot.
Disorders of the Joints
The ankle is the most frequently injured joint in the body, with the most common injury being an inversion ankle sprain. A sprain is the stretching or tearing of the supporting ligaments. Excess inversion causes the talus bone to tilt laterally, thus damaging the ligaments on the lateral side of the ankle. The anterior talofibular ligament is most commonly injured, followed by the calcaneofibular ligament. In severe inversion injuries, the forceful lateral movement of the talus not only ruptures the lateral ankle ligaments, but also fractures the distal fibula.
Less common are eversion sprains of the ankle, which involve stretching of the deltoid ligament on the medial side of the ankle. Forcible eversion of the foot, for example, with an awkward landing from a jump or when a football player has a foot planted and is hit on the lateral ankle, can result in a Pott’s fracture and dislocation of the ankle joint. In this injury, the very strong deltoid ligament does not tear, but instead shears off the medial malleolus of the tibia. This frees the talus, which moves laterally and fractures the distal fibula. In extreme cases, the posterior margin of the tibia may also be sheared off.
Above the ankle, the distal ends of the tibia and fibula are united by a strong syndesmosis formed by the interosseous membrane and ligaments at the distal tibiofibular joint. These connections prevent separation between the distal ends of the tibia and fibula and maintain the talus locked into position between the medial malleolus and lateral malleolus. Injuries that produce a lateral twisting of the leg on top of the planted foot can result in stretching or tearing of the tibiofibular ligaments, producing a syndesmotic ankle sprain or “high ankle sprain.”
Most ankle sprains can be treated using the RICE technique: Rest, Ice, Compression, and Elevation. Reducing joint mobility using a brace or cast may be required for a period of time. More severe injuries involving ligament tears or bone fractures may require surgery.
Section Review
Although synovial joints share many common features, each joint of the body is specialised for certain movements and activities. The joints of the upper limb provide for large ranges of motion, which give the upper limb great mobility, thus enabling actions such as the throwing of a ball or typing on a keyboard. The joints of the lower limb are more robust, giving them greater strength and the stability needed to support the body weight during running, jumping, or kicking activities.
The joints of the vertebral column include the symphysis joints formed by each intervertebral disc and the plane synovial joints between the superior and inferior articular processes of adjacent vertebrae. Each of these joints provide for limited motions, but these sum together to produce flexion, extension, lateral flexion, and rotation of the neck and body. The range of motions available in each region of the vertebral column varies, with all these motions available in the cervical region. Only rotation is allowed in the thoracic region, while the lumbar region has considerable extension, flexion, and lateral flexion, but rotation is prevented. The atlanto-occipital joint allows for flexion and extension of the head, while the atlantoaxial joint is a pivot joint that provides for rotation of the head.
The temporomandibular joint is the articulation between the condyle of the mandible and the mandibular fossa and articular tubercle of the skull temporal bone. An articular disc is located between the bony components of this joint. A combination of gliding and hinge motions of the mandibular condyle allows for elevation/depression, protraction/retraction, and side-to-side motions of the lower jaw.
The glenohumeral (shoulder) joint is a multiaxial ball-and-socket joint that provides flexion/extension, abduction/adduction, circumduction, and medial/lateral rotation of the humerus. The head of the humerus articulates with the glenoid cavity of the scapula. The glenoid labrum extends around the margin of the glenoid cavity. Intrinsic ligaments, including the coracohumeral ligament and glenohumeral ligaments, provide some support for the shoulder joint. However, the primary support comes from muscles crossing the joint whose tendons form the rotator cuff. These muscle tendons are protected from friction against the scapula by the subacromial bursa and subscapular bursa.
The elbow is a uniaxial hinge joint that allows for flexion/extension of the forearm. It includes the humeroulnar joint and the humeroradial joint. The medial elbow is supported by the ulnar collateral ligament and the radial collateral ligament supports the lateral side. These ligaments prevent side-to-side movements and resist hyperextension of the elbow. The proximal radioulnar joint is a pivot joint that allows for rotation of the radius during pronation/supination of the forearm. The annular ligament surrounds the head of the radius to hold it in place at this joint.
The hip joint is a ball-and-socket joint whose motions are more restricted than at the shoulder to provide greater stability during weight bearing. The hip joint is the articulation between the head of the femur and the acetabulum of the hip bone. The acetabulum is deepened by the acetabular labrum. The iliofemoral, pubofemoral, and ischiofemoral ligaments strongly support the hip joint in the upright, standing position. The ligament of the head of the femur provides little support but carries an important artery that supplies the femur.
The knee includes three articulations. The femoropatellar joint is between the patella and distal femur. The patella, a sesamoid bone incorporated into the tendon of the quadriceps femoris muscle of the anterior thigh, serves to protect this tendon from rubbing against the distal femur during knee movements. The medial and lateral tibiofemoral joints, between the condyles of the femur and condyles of the tibia, are modified hinge joints that allow for knee extension and flexion. During these movements, the condyles of the femur both roll and glide over the surface of the tibia. As the knee comes into full extension, a slight medial rotation of the femur serves to “lock” the knee into its most stable, weight-bearing position. The reverse motion, a small lateral rotation of the femur, is required to initiate knee flexion. When the knee is flexed, some rotation of the leg is available.
Two extrinsic ligaments, the tibial collateral ligament on the medial side and the fibular collateral ligament on the lateral side, serve to resist hyperextension or rotation of the extended knee joint. Two intracapsular ligaments, the anterior cruciate ligament and posterior cruciate ligament, span between the tibia and the inner aspects of the femoral condyles. The anterior cruciate ligament resists hyperextension of the knee, while the posterior cruciate ligament prevents anterior sliding of the femur, thus supporting the knee when it is flexed and weight bearing. The medial and lateral menisci, located between the femoral and tibial condyles, are articular discs that provide padding and improve the fit between the bones.
The talocrural joint forms the ankle. It consists of the articulation between the talus bone and the medial malleolus of the tibia, the distal end of the tibia, and the lateral malleolus of the fibula. This is a uniaxial hinge joint that allows only dorsiflexion and plantar flexion of the foot. Gliding motions at the subtalar and intertarsal joints of the foot allow for inversion/eversion of the foot. The ankle joint is supported on the medial side by the deltoid ligament, which prevents side-to-side motions of the talus at the talocrural joint and resists excessive eversion of the foot. The lateral ankle is supported by the anterior and posterior talofibular ligaments and the calcaneofibular ligament. These support the ankle joint and resist excess inversion of the foot. An inversion ankle sprain, a common injury, will result in injury to one or more of these lateral ankle ligaments.
Describe the two processes by which mesenchyme can give rise to bone
Discuss the process by which joints of the limbs are formed
Joints form during embryonic development in conjunction with the formation and growth of the associated bones. The embryonic tissue that gives rise to all bones, cartilages, and connective tissues of the body is called mesenchyme. In the head, mesenchyme will accumulate at those areas that will become the bones that form the top and sides of the skull. The mesenchyme in these areas will develop directly into bone through the process of intramembranous ossification, in which mesenchymal cells differentiate into bone-producing cells that then generate bone tissue. The mesenchyme between the areas of bone production will become the fibrous connective tissue that fills the spaces between the developing bones. Initially, the connective tissue-filled gaps between the bones are wide and are called fontanelles. After birth, as the skull bones grow and enlarge, the gaps between them decrease in width and the fontanelles are reduced to suture joints in which the bones are united by a narrow layer of fibrous connective tissue.
The bones that form the base and facial regions of the skull develop through the process of endochondral ossification. In this process, mesenchyme accumulates and differentiates into hyaline cartilage, which forms a model of the future bone. The hyaline cartilage model is then gradually, over a period of many years, displaced by bone. The mesenchyme between these developing bones becomes the fibrous connective tissue of the suture joints between the bones in these regions of the skull.
A similar process of endochondral ossification gives rises to the bones and joints of the limbs. The limbs initially develop as small limb buds that appear on the sides of the embryo around the end of the fourth week of development. Starting during the sixth week, as each limb bud continues to grow and elongate, areas of mesenchyme within the bud begin to differentiate into the hyaline cartilage that will form models for of each of the future bones. The synovial joints will form between the adjacent cartilage models, in an area called the joint interzone. Cells at the centre of this interzone region undergo cell death to form the joint cavity, while surrounding mesenchyme cells will form the articular capsule and supporting ligaments. The process of endochondral ossification, which converts the cartilage models into bone, begins by the twelfth week of embryonic development. At birth, ossification of much of the bone has occurred, but the hyaline cartilage of the epiphyseal plate will remain throughout childhood and adolescence to allow for bone lengthening. Hyaline cartilage is also retained as the articular cartilage that covers the surfaces of the bones at synovial joints.
Section Review
During embryonic growth, bones and joints develop from mesenchyme, an embryonic tissue that gives rise to bone, cartilage, and fibrous connective tissues. In the skull, the bones develop either directly from mesenchyme through the process of intramembranous ossification, or indirectly through endochondral ossification, which initially forms a hyaline cartilage model of the future bone, which is later converted into bone. In both cases, the mesenchyme between the developing bones differentiates into fibrous connective tissue that will unite the skull bones at suture joints. In the limbs, mesenchyme accumulations within the growing limb bud will become a hyaline cartilage model for each of the limb bones. A joint interzone will develop between these areas of cartilage. Mesenchyme cells at the margins of the interzone will give rise to the articular capsule, while cell death at the centre forms the space that will become the joint cavity of the future synovial joint. The hyaline cartilage model of each limb bone will eventually be converted into bone via the process of endochondral ossification. However, hyaline cartilage will remain, covering the ends of the adult bone as the articular cartilage.
Identify the organs of the gastrointestinal tract (alimentary canal) from proximal to distal, and briefly state their function
Identify the accessory digestive organs and briefly state their function
Describe the four fundamental tissue layers of the gastrointestinal tract
Contrast the contributions of the enteric and autonomic nervous systems to digestive system functioning
Explain how the peritoneum anchors the digestive organs
The function of the digestive system is to break down the foods you eat, release their nutrients, and absorb those nutrients into the body. Although the small intestine is the workhorse of the system, where the majority of digestion occurs, and where most of the released nutrients are absorbed into the blood or lymph, each of the digestive system organs makes a vital contribution to this process (Figure 12.1.1).
Figure 12.1.1. Components of the digestive system. All digestive organs play integral roles in the life-sustaining process of digestion.
As is the case with all body systems, the digestive system does not work in isolation; it functions cooperatively with the other systems of the body. Consider for example, the interrelationship between the digestive and cardiovascular systems. Arteries supply the digestive organs with oxygen and processed nutrients, and veins drain the digestive tract. These intestinal veins, constituting the hepatic portal system, are unique; they do not return blood directly to the heart. Rather, this blood is diverted to the liver where its nutrients are off-loaded for processing before blood completes its circuit back to the heart. At the same time, the digestive system provides nutrients to the heart muscle and vascular tissue to support their functioning. The interrelationship of the digestive and endocrine systems is also critical. Hormones secreted by several endocrine glands, as well as endocrine cells of the pancreas, the stomach, and the small intestine, contribute to the control of digestion and nutrient metabolism. In turn, the digestive system provides the nutrients to fuel endocrine function. Table 12.1.1 gives a quick glimpse at how these other systems contribute to the functioning of the digestive system.
Table 12.1.1. Contribution of Other Body Systems to the Digestive System
Body system
Benefits received by the digestive system
Cardiovascular
Blood supplies digestive organs with oxygen and processed nutrients
Endocrine
Endocrine hormones help regulate secretion in digestive glands and accessory organs
Integumentary
Skin helps protect digestive organs and synthesises vitamin D for calcium absorption
Lymphatic
Mucosa-associated lymphoid tissue and other lymphatic tissue defend against entry of pathogens; lacteal absorb lipids; and lymphatic vessels transport lipids to bloodstream
Muscular
Skeletal muscles support and protect abdominal organs
Nervous
Sensory and motor neurons help regulate secretions and muscle contractions in the digestive tract
Respiratory
Respiratory organs provide oxygen and remove carbon dioxide
Skeletal
Bones help protect and support digestive organs
Urinary
Kidneys convert vitamin D into its active form, allowing calcium absorption in the small intestine
Digestive System Organs
The easiest way to understand the digestive system is to divide its organs into two main categories. The first group is the organs that make up the gastrointestinal tract. Accessory digestive organs comprise the second group and are critical for orchestrating the breakdown of food and the assimilation of its nutrients into the body. Accessory digestive organs, despite their name, are critical to the function of the digestive system.
Gastrointestinal Tract Organs
Also called the gastrointestinal (GI) tract or gut, the gastrointestinal tract (aliment- = “to nourish”) is a hollow, muscular, one-way tube about 7.62 metres in length during life and closer to 10.67 metres in length when measured after death, once smooth muscle tone is lost. The main function of the organs of the gastrointestinal tract is to nourish the body. This tube begins at the mouth and terminates at the anus. Between those two points, the canal is modified as the pharynx, oesophagus, stomach, and small and large intestines to fit the functional needs of the body. Both the mouth and anus are open to the external environment; thus, food and wastes within the gastrointestinal tract are technically considered to be outside the body. Only through the process of absorption do the nutrients in food enter into and nourish the body’s “inner space.”
Accessory Structures
Each accessory digestive organ aids in the breakdown of food (Figure 12.1.2). Within the mouth, the teeth and tongue begin mechanical digestion, whereas the salivary glands begin chemical digestion. Once food products enter the small intestine, the gallbladder, liver, and pancreas release secretions—such as bile and enzymes—essential for digestion to continue. Together, these are called accessory organs because they sprout from the lining cells of the developing gut (mucosa) and augment its function; indeed, you could not live without their vital contributions, and many significant diseases result from their malfunction. Even after development is complete, they maintain a connection to the gut by way of ducts.
Histology of the Gastrointestinal Tract
Throughout its length, the alimentary tract is composed of the same four tissue layers; the details of their structural arrangements vary to fit their specific functions. Starting from the lumen and moving outwards, these layers are the mucosa, submucosa, muscularis, and serosa, which is continuous with the mesentery (see Figure 12.1.2).
Figure 12.1.2. Layers of the gastrointestinal tract. The wall of the gastrointestinal tract has four basic tissue layers: the mucosa, submucosa, muscularis and serosa.
The mucosa is referred to as a mucous membrane, because mucus production is a characteristic feature of gut epithelium. The membrane consists of epithelium, which is in direct contact with ingested food, and the lamina propria, a layer of connective tissue analogous to the dermis. In addition, the mucosa has a thin, smooth muscle layer, called the muscularis mucosa (not to be confused with the muscularis layer, described below).
Epithelium—In the mouth, pharynx, oesophagus, and anal canal, the epithelium is primarily a non-keratinised, stratified squamous epithelium. In the stomach and intestines, it is a simple columnar epithelium. Notice that the epithelium is in direct contact with the lumen, the space inside the gastrointestinal tract. Interspersed among its epithelial cells are goblet cells, which secrete mucus and fluid into the lumen, and enteroendocrine cells, which secrete hormones into the interstitial spaces between cells. Epithelial cells have a very brief lifespan, averaging from only a couple of days (in the mouth) to about a week (in the gut). This process of rapid renewal helps preserve the health of the gastrointestinal tract, despite the wear and tear resulting from continued contact with foodstuffs.
Lamina propria—In addition to loose connective tissue, the lamina propria contains numerous blood and lymphatic vessels that transport nutrients absorbed through the gastrointestinal tract to other parts of the body. The lamina propria also serves an immune function by housing clusters of lymphocytes, making up the mucosa-associated lymphoid tissue (MALT). These lymphocyte clusters are particularly substantial in the distal ileum where they are known as Peyer’s patches. When you consider that the gastrointestinal tract is exposed to foodborne bacteria and other foreign matter, it is not hard to appreciate why the immune system has evolved a means of defending against the pathogens encountered within it.
Muscularis mucosa—This thin layer of smooth muscle is in a constant state of tension, pulling the mucosa of the stomach and small intestine into undulating folds. These folds dramatically increase the surface area available for digestion and absorption.
As its name implies, the submucosa lies immediately beneath the mucosa. A broad layer of dense connective tissue, it connects the overlying mucosa to the underlying muscularis. It includes blood and lymphatic vessels (which transport absorbed nutrients), and a scattering of submucosal glands that release digestive secretions. Additionally, it serves as a conduit for a dense branching network of nerves, the submucosal plexus, which functions as described below.
The third layer of the gastrointestinal tract is the muscularis (also called the muscularis externa). The muscularis in the small intestine is made up of a double layer of smooth muscle: an inner circular layer and an outer longitudinal layer. The contractions of these layers promote mechanical digestion, expose more of the food to digestive chemicals, and move the food along the canal. In the most proximal and distal regions of the gastrointestinal tract, including the mouth, pharynx, anterior part of the oesophagus, and external anal sphincter, the muscularis is made up of skeletal muscle, which gives you voluntary control over swallowing and defecation. The basic two-layer structure found in the small intestine is modified in the organs proximal and distal to it. The stomach is equipped for its churning function by the addition of a third layer, the oblique muscle. While the colon has two layers like the small intestine, its longitudinal layer is segregated into three narrow parallel bands, the taeniae coli, which make it look like a series of pouches rather than a simple tube.
The serosa is the portion of the gastrointestinal tract superficial to the muscularis. Present only in the region of the gastrointestinal tract within the abdominal cavity, it consists of a layer of visceral peritoneum overlying a layer of loose connective tissue. Instead of serosa, the mouth, pharynx, and oesophagus have a dense sheath of collagen fibres called the adventitia. These tissues serve to hold the gastrointestinal tract in place near the ventral surface of the vertebral column.
Nerve Supply
As soon as food enters the mouth, it is detected by receptors that send impulses along the sensory neurons of cranial nerves. Without these nerves, not only would your food be without taste, but you would also be unable to feel either the food or the structures of your mouth, and you would be unable to avoid biting yourself as you chew, an action enabled by the motor branches of cranial nerves.
Intrinsic innervation of much of the gastrointestinal tract is provided by the enteric nervous system, which runs from the oesophagus to the anus, and contains approximately 100 million motor, sensory, and interneurons (unique to this system compared to all other parts of the peripheral nervous system). These enteric neurons are grouped into two plexuses. The myenteric plexus (plexus of Auerbach) lies in the muscularis layer of the gastrointestinal tract and is responsible for motility, especially the rhythm and force of the contractions of the muscularis. The submucosal plexus (plexus of Meissner) lies in the submucosal layer and is responsible for regulating digestive secretions and reacting to the presence of food.
Extrinsic innervations of the gastrointestinal tract are provided by the autonomic nervous system, which includes both sympathetic and parasympathetic nerves. In general, sympathetic activation (the fight-or-flight response) restricts the activity of enteric neurons, thereby decreasing GI secretion and motility. In contrast, parasympathetic activation (the rest-and-digest response) increases GI secretion and motility by stimulating neurons of the enteric nervous system.
Blood Supply
The blood vessels serving the digestive system have two functions. They transport the protein and carbohydrate nutrients absorbed by mucosal cells after food is digested in the lumen. Lipids are absorbed via lacteals, tiny structures of the lymphatic system. The blood vessels’ second function is to supply the organs of the gastrointestinal tract with the nutrients and oxygen needed to drive their cellular processes.
Specifically, the more anterior parts of the gastrointestinal tract are supplied with blood by arteries branching off the aortic arch and thoracic aorta. Below this point, the gastrointestinal tract is supplied with blood by arteries branching from the abdominal aorta. The celiac trunk services the liver, stomach, and duodenum, whereas the superior and inferior mesenteric arteries supply blood to the remaining small and large intestines.
The veins that collect nutrient-rich blood from the small intestine (where most absorption occurs) empty into the hepatic portal system. This venous network takes the blood into the liver where the nutrients are either processed or stored for later use. Only then does the blood drained from the gastrointestinal tract viscera circulate back to the heart. To appreciate just how demanding the digestive process is on the cardiovascular system, consider that while you are “resting and digesting,” about one-fourth of the blood pumped with each heartbeat enters arteries serving the intestines.
The Peritoneum
The digestive organs within the abdominal cavity are held in place by the peritoneum, a broad serous membranous sac made up of squamous epithelial tissue surrounded by connective tissue. It is composed of two different regions: the parietal peritoneum, which lines the abdominal wall, and the visceral peritoneum, which envelopes the abdominal organs (Figure 12.1.3). The peritoneal cavity is the space bounded by the visceral and parietal peritoneal surfaces. A few millilitres of watery fluid act as a lubricant to minimise friction between the serosal surfaces of the peritoneum.
Figure 12.1.3. The peritoneum. A cross-section of the abdomen shows the relationship between abdominal organs and the peritoneum (darker lines).
Disorders of the Digestive System: Peritonitis
Inflammation of the peritoneum is called peritonitis. Chemical peritonitis can develop any time the wall of the gastrointestinal tract is breached, allowing the contents of the lumen entry into the peritoneal cavity, for example, when an ulcer perforates the stomach wall, gastric juices spill into the peritoneal cavity. Haemorrhagic peritonitis occurs after a ruptured tubal pregnancy or traumatic injury to the liver or spleen fills the peritoneal cavity with blood. Even more severe peritonitis is associated with bacterial infections seen with appendicitis, colonic diverticulitis, and pelvic inflammatory disease (infection of uterine tubes, usually by sexually transmitted bacteria). Peritonitis is life threatening and often results in emergency surgery to correct the underlying problem and intensive antibiotic therapy. When your great grandparents and even your parents were young, the mortality from peritonitis was high. Aggressive surgery, improvements in anaesthesia safety, the advance of critical care expertise, and antibiotics have greatly improved the mortality rate from this condition. Even so, the mortality rate still ranges from 30 to 40 percent.
The visceral peritoneum includes multiple large folds that envelope various abdominal organs, holding them to the dorsal surface of the body wall. Within these folds are blood vessels, lymphatic vessels, and nerves that innervate the organs with which they are in contact, supplying their adjacent organs. The five major peritoneal folds are described in Table 12.1.2. Note that during foetal development, certain digestive structures, including the first portion of the small intestine (called the duodenum), the pancreas, and portions of the large intestine (the ascending and descending colon, and the rectum) remain completely or partially posterior to the peritoneum. Thus, the location of these organs is described as retroperitoneal.
Table 12.1.2. The Five Major Peritoneal Folds
Fold
Description
Greater omentum
Apron-like structure that lies superficial to the small intestine and transverse colon; a site of fat deposition in people who are overweight
Falciform ligament
Anchors the liver to the anterior abdominal wall and inferior border of the diaphragm
Lesser omentum
Suspends the stomach from the inferior border of the liver; provides a pathway for structures connecting to the liver
Mesentery
Vertical band of tissue anterior to the lumbar vertebrae and anchoring all of the small intestine except the initial portion (the duodenum)
Mesocolon
Attaches two portions of the large intestine (the transverse and sigmoid colon) to the posterior abdominal wall
Section Review
The digestive system includes the organs of the gastrointestinal tract and accessory structures. The gastrointestinal tract forms a continuous tube that is open to the outside environment at both ends. The organs of the gastrointestinal tract are the mouth, pharynx, oesophagus, stomach, small intestine, and large intestine. The accessory digestive structures include the teeth, tongue, salivary glands, liver, pancreas and gallbladder. The wall of the gastrointestinal tract is composed of four basic tissue layers: mucosa, submucosa, muscularis and serosa. The enteric nervous system provides intrinsic innervation, and the autonomic nervous system provides extrinsic innervation.
Discuss six fundamental activities of the digestive system, giving an example of each
Compare and contrast the neural and hormonal controls involved in digestion
The digestive system uses mechanical and chemical activities to break food down into absorbable substances during its journey through the digestive system. Table 12.2.1 provides an overview of the basic functions of the digestive organs.
Table 12.2.1. Functions of the Digestive Organs
Organ
Major functions
Other functions
Mouth
Ingests food
Chews and mixes food
Begins chemical breakdown of carbohydrates
Moves food into the pharynx
Begins breakdown of lipids via lingual lipase
Moistens and dissolves food, allowing you to taste it
Cleans and lubricates the teeth and oral cavity
Has some antimicrobial activity
Pharynx
Propels food from the oral cavity to the oesophagus
Lubricates food and passageways
Oesophagus
Propels food to the stomach
Lubricates food and passageways
Stomach
Mixes and churns food with gastric juices to form chyme
Begins chemical breakdown of proteins
Releases food into the duodenum as chyme
Absorbs some fat-soluble substances (for example alcohol, aspirin)
Possesses antimicrobial functions
Stimulates protein-digesting enzymes
Secretes intrinsic factor required for vitamin B12 absorption in small intestine
Small intestine
Mixes chyme with digestive juices
Propels food at a rate slow enough for digestion and absorption
Absorbs breakdown products of carbohydrates, proteins, lipids, and nucleic acids, along with vitamins, minerals, and water
Performs physical digestion via segmentation
Provides optimal medium for enzymatic activity
Accessory organs
Liver: produces bile salts, which emulsify lipids, aiding their digestion and absorption
Gallbladder: stores, concentrates and releases bile
Pancreas: produces digestive enzymes and bicarbonate
Bicarbonate-rich pancreatic secretions (juices) help neutralise acidic chyme and provide optimal environment for enzymatic activity
Large intestine
Further breaks down food residues
Absorbs most residual water, electrolytes, and vitamins produced by enteric bacteria
Propels faeces towards rectums
Eliminates faeces
Food residue is concentrated and temporarily stored prior to defecation
Mucus eases passage of faeces through colon
Digestive Processes
The processes of digestion include six activities: ingestion, propulsion, mechanical or physical digestion, chemical digestion, absorption and defaecation.
The first of these processes, ingestion, refers to the entry of food into the gastrointestinal tract through the mouth. There, the food is chewed and mixed with saliva, which contains enzymes that begin breaking down the carbohydrates in the food plus some lipid digestion via lingual lipase. Chewing increases the surface area of the food and allows an appropriately sized bolus to be produced.
Food leaves the mouth when the tongue and pharyngeal muscles propel it into the oesophagus. This act of swallowing, the last voluntary act until defaecation, is an example of propulsion, which refers to the movement of food through the digestive tract. It includes both the voluntary process of swallowing and the involuntary process of peristalsis. Peristalsis consists of sequential, alternating waves of contraction and relaxation of alimentary wall smooth muscles, which act to propel food along (Figure 12.2.1). These waves also play a role in mixing food with digestive secretions. Peristalsis is so powerful that foods and liquids you swallow enter your stomach even if you are standing on your head.
Figure 12.2.1. Peristalsis. Peristalsis moves food through the digestive tract with alternating waves of muscle contraction and relaxation.
Digestion includes both mechanical and chemical processes. Mechanical digestion is a purely physical process that does not change the chemical nature of the food. Instead, it makes the food smaller to increase both surface area and mobility. It includes mastication, or chewing, as well as tongue movements that help break food into smaller bits and mix food with saliva. Although there may be a tendency to think that mechanical digestion is limited to the first steps of the digestive process, it occurs after the food leaves the mouth, as well. The mechanical churning of food in the stomach serves to further break it apart and expose more of its surface area to digestive secretions, creating an acidic “soup” called chyme. Segmentation, which occurs mainly in the small intestine, consists of localised contractions of circular muscle of the muscularis layer of the gastrointestinal tract. These contractions isolate small sections of the intestine, moving their contents back and forth while continuously subdividing, breaking up, and mixing the contents. By moving food back and forth in the intestinal lumen, segmentation mixes food with digestive secretions and facilitates absorption.
In chemical digestion, starting in the mouth, digestive secretions break down complex food molecules into their chemical building blocks (for example, proteins into separate amino acids). These secretions vary in composition, but typically contain water, various enzymes, acids, and salts. The process is completed in the small intestine.
Food that has been broken down is of no value to the body unless it enters the bloodstream, and its nutrients are put to work. This occurs through the process of absorption, which takes place primarily within the small intestine. There, most nutrients are absorbed from the lumen of the gastrointestinal tract into the bloodstream through the epithelial cells that make up the mucosa. Lipids are absorbed into lacteals and are transported via the lymphatic vessels to the bloodstream (the subclavian veins near the heart). The details of these processes will be discussed later.
In defaecation, the final step in digestion, undigested materials are removed from the body as faeces.
Ageing and the Digestive System: From Appetite Suppression to Constipation
Age-related changes in the digestive system begin in the mouth and can affect virtually every aspect of the digestive system. Taste buds become less sensitive, so food is not as appetising as it once was. A slice of pizza is a challenge, not a treat, when you have lost teeth, your gums are diseased, and your salivary glands are not producing enough saliva. Swallowing can be difficult, and ingested food moves slowly through the gastrointestinal tract because of reduced strength and tone of muscular tissue. Neurosensory feedback is also dampened, slowing the transmission of messages that stimulate the release of enzymes and hormones.
Pathologies that affect the digestive organs—such as hiatal hernia, gastritis, and peptic ulcer disease—can occur at greater frequencies as you age. Problems in the small intestine may include duodenal ulcers, maldigestion, and malabsorption. Problems in the large intestine include haemorrhoids, diverticular disease, and constipation. Conditions that affect the function of accessory organs—and their abilities to deliver pancreatic enzymes and bile to the small intestine—include jaundice, acute pancreatitis, cirrhosis, and gallstones.
In some cases, a single organ oversees a digestive process, for example, ingestion occurs only in the mouth and defaecation only in the anus. However, most digestive processes involve the interaction of several organs and occur gradually as food moves through the gastrointestinal tract (Figure 12.2.2).
Figure 12.2.2. Digestive processes. The digestive processes are ingestion, propulsion, mechanical digestion, chemical digestion, absorption and defaecation.
Some chemical digestion occurs in the mouth. Some absorption can occur in the mouth and stomach, for example, alcohol and aspirin.
Regulatory Mechanisms
Neural and endocrine regulatory mechanisms work to maintain the optimal conditions in the lumen needed for digestion and absorption. These regulatory mechanisms, which stimulate digestive activity through mechanical and chemical activity, are controlled both extrinsically and intrinsically.
Neural Controls
The walls of the gastrointestinal tract contain a variety of sensors that help regulate digestive functions. These include mechanoreceptors, chemoreceptors (including taste receptors), and osmoreceptors, which are capable of detecting mechanical, chemical, and osmotic stimuli, respectively. , These receptors can sense when the presence of food has caused the stomach to expand, whether food particles have been sufficiently broken down, how much liquid is present, and the type of nutrients in the food (lipids, carbohydrates, and/or proteins). Stimulation of these receptors provokes an appropriate reflex that furthers the process of digestion. This may entail sending a message that activates the glands that secrete digestive secretions into the lumen, or it may mean the stimulation of muscles within the gastrointestinal tract, thereby activating peristalsis and segmentation that move food along the intestinal tract.
The walls of the entire gastrointestinal tract are embedded with nerve plexuses that interact with the central nervous system and other nerve plexuses—either within the same digestive organ or in different ones. These interactions prompt several types of reflexes. Extrinsic nerve plexuses orchestrate long reflexes, which involve the central and autonomic nervous systems and work in response to stimuli from outside the digestive system. Short reflexes, on the other hand, are orchestrated by intrinsic nerve plexuses within the gastrointestinal tract wall. These two plexuses and their connections were introduced earlier as the enteric nervous system. Short reflexes regulate activities in one area of the digestive tract and may coordinate local peristaltic movements and stimulate digestive secretions, for example, the sight, smell, and taste of food initiate long reflexes that begin with a sensory neuron delivering a signal to the medulla oblongata. The response to the signal is to stimulate cells in the stomach to begin secreting digestive secretions in preparation for incoming food. In contrast, food that distends the stomach initiates short reflexes that cause cells in the stomach wall to increase their secretion of digestive secretions.
Hormonal Controls
A variety of hormones are involved in the digestive process. The main digestive hormone of the stomach is gastrin, which is secreted in response to the presence of food. Gastrin stimulates the secretion of gastric acid by the parietal cells of the stomach mucosa. Other GI hormones are produced and act upon the gut and its accessory organs. Hormones produced by the duodenum include secretin, which stimulates a watery secretion of bicarbonate by the pancreas; cholecystokinin (CCK), which stimulates the secretion of pancreatic enzymes and bile from the liver and release of bile from the gallbladder; and gastric inhibitory peptide, which inhibits gastric secretion and slows gastric emptying and motility. These GI hormones are secreted by specialised epithelial cells, called enteroendocrine cells, located in the mucosal epithelium of the stomach and small intestine. These hormones then enter the bloodstream, through which they can reach their target organs.
Section Review
The digestive system ingests and digests food, absorbs released nutrients, and excretes food components that are indigestible. The six activities involved in this process are ingestion, motility, mechanical digestion, chemical digestion, absorption and defaecation. These processes are regulated by neural and hormonal mechanisms.
Describe the structures of the mouth, including its three accessory digestive organs
Group the 32 adult teeth according to name, location, and function
Describe the process of swallowing, including the roles of the tongue, upper oesophageal sphincter and epiglottis
Trace the pathway food follows from ingestion into the mouth through release into the stomach
In this section, you will examine the anatomy and functions of the three main organs of the upper gastrointestinal tract—the mouth, pharynx, and oesophagus—as well as three associated accessory organs—the tongue, salivary glands and teeth.
The Mouth
The cheeks, tongue, and palate frame the mouth, which is also called the oral cavity (or buccal cavity). The structures of the mouth are illustrated in Figure 12.3.1.
At the entrance to the mouth are the lips, or labia (singular = labium). Their outer covering is skin, which transitions to a mucous membrane in the mouth proper. Lips are very vascular with a thin layer of keratin; hence, the reason they are “red.” They have a huge representation on the cerebral cortex, which probably explains the human fascination with kissing! The lips cover the orbicularis oris muscle, which regulates what comes in and goes out of the mouth. The labial frenulum is a midline fold of mucous membrane that attaches the inner surface of each lip to the gum. The cheeks make up the oral cavity’s sidewalls. While their outer covering is skin, their inner covering is mucous membrane. This membrane is made up of non-keratinised, stratified squamous epithelium. Between the skin and mucous membranes are connective tissue and buccinator muscles. The next time you eat some food, notice how the buccinator muscles in your cheeks and the orbicularis oris muscle in your lips contract, helping you keep the food from falling out of your mouth. Additionally, notice how these muscles work when you are speaking.
The pocket-like part of the mouth that is framed on the inside by the gums and teeth, and on the outside by the cheeks and lips is called the oral vestibule. Moving farther into the mouth, the opening between the oral cavity and throat (oropharynx) is called the fauces (like the kitchen “faucet”). The main open area of the mouth, or oral cavity proper, runs from the gums and teeth to the fauces.
When you are chewing, you do not find it difficult to breathe simultaneously. The next time you have food in your mouth, notice how the arched shape of the roof of your mouth allows you to handle both digestion and respiration at the same time. This arch is called the palate. The anterior region of the palate serves as a wall (or septum) between the oral and nasal cavities as well as a rigid shelf against which the tongue can push food. It is created by the maxillary and palatine bones of the skull and, given its bony structure, is known as the hard palate. If you run your tongue along the roof of your mouth, you’ll notice that the hard palate ends in the posterior oral cavity, and the tissue becomes fleshier. This part of the palate, known as the soft palate, is composed mainly of skeletal muscle. You can therefore manipulate, subconsciously, the soft palate—for instance, to yawn, swallow, or sing (see Figure 12.3.1).
Figure 12.3.1. Mouth. The mouth includes the lips, tongue, palate, gums, and teeth.
A fleshy bead of tissue called the uvula drops down from the centre of the posterior edge of the soft palate. Although some have suggested that the uvula is a vestigial organ, it serves an important purpose. When you swallow, the soft palate and uvula move upward, helping to keep foods and liquid from entering the nasal cavity. Unfortunately, it can also contribute to the sound produced by snoring. Two muscular folds extend downward from the soft palate, on either side of the uvula. Toward the front, the palatoglossal arch lies next to the base of the tongue; behind it, the palatopharyngeal arch forms the superior and lateral margins of the fauces. Between these two arches are the palatine tonsils, clusters of lymphoid tissue that protect the pharynx. The lingual tonsils are located at the base of the tongue.
The Tongue
Perhaps you have heard it said that the tongue is the strongest muscle in the body. Those who stake this claim cite its strength proportionate to its size. Although it is difficult to quantify the relative strength of different muscles, it remains indisputable that the tongue is a workhorse, facilitating ingestion, mechanical digestion, chemical digestion (lingual lipase), sensation (of taste, texture, and temperature of food), swallowing, and vocalisation.
The tongue is attached to the mandible, the styloid processes of the temporal bones, and the hyoid bone. The hyoid is unique in that it only distantly/indirectly articulates with other bones. The tongue is positioned over the floor of the oral cavity. A medial septum extends the entire length of the tongue, dividing it into symmetrical halves.
Beneath its mucous membrane covering, each half of the tongue is composed of the same number and type of intrinsic and extrinsic skeletal muscles. The intrinsic muscles (those within the tongue) are the longitudinalis inferior, longitudinalis superior, transversus linguae, and verticalis linguae muscles. These allow you to change the size and shape of your tongue, as well as to stick it out, if you wish. Having such a flexible tongue facilitates both swallowing and speech.
As you learned in your study of the muscular system, the extrinsic muscles of the tongue are the mylohyoid, hyoglossus, styloglossus, and genioglossus muscles. These muscles originate outside the tongue and insert into connective tissues within the tongue. The mylohyoid is responsible for raising the tongue, the hyoglossus pulls it down and back, the styloglossus pulls it up and back, and the genioglossus pulls it forward. Working in concert, these muscles perform three important digestive functions in the mouth: (1) position food for optimal chewing, (2) gather food into a bolus (rounded mass), and (3) position food so it can be swallowed.
The top and sides of the tongue are studded with papillae, extensions of lamina propria of the mucosa, which are covered in stratified squamous epithelium (Figure 12.3.2). Fungiform papillae, which are mushroom shaped, cover a large area of the tongue; they tend to be larger toward the rear of the tongue and smaller on the tip and sides. In contrast, filiform papillae are long and thin. Fungiform papillae contain taste buds, and filiform papillae have touch receptors that help the tongue move food around in the mouth. The filiform papillae create an abrasive surface that performs mechanically, much like a cat’s rough tongue that is used for grooming. Lingual glands in the lamina propria of the tongue secrete mucus and a watery serous fluid that contains the enzyme lingual lipase, which plays a minor role in breaking down triglycerides but does not begin working until it is activated in the stomach. A fold of mucous membrane on the underside of the tongue, the lingual frenulum, tethers the tongue to the floor of the mouth. People with the congenital anomaly ankyloglossia, also known by the non-medical term “tongue tie,” have a lingual frenulum that is too short or otherwise malformed. Severe ankyloglossia can impair speech and must be corrected with surgery.
Figure 12.3.2. Tongue. This superior view of the tongue shows the locations and types of lingual papillae.
The Salivary Glands
Many small salivary glands are housed within the mucous membranes of the mouth and tongue. These minor exocrine glands are constantly secreting saliva, either directly into the oral cavity or indirectly through ducts, even while you sleep. In fact, an average of 1 to 1.5 litres of saliva is secreted each day. Usually just enough saliva is present to moisten the mouth and teeth. Secretion increases when you eat, because saliva is essential to moisten food and initiate the chemical breakdown of carbohydrates. Small amounts of saliva are also secreted by the labial glands in the lips. In addition, the buccal glands in the cheeks, palatal glands in the palate, and lingual glands in the tongue help ensure that all areas of the mouth are supplied with adequate saliva.
The Major Salivary Glands
Outside the oral mucosa are three pairs of major salivary glands, which secrete the majority of saliva into ducts that open into the mouth:
The submandibular glands, which are in the floor of the mouth, secrete saliva into the mouth through the submandibular ducts.
The sublingual glands, which lie below the tongue, use the lesser sublingual ducts to secrete saliva into the oral cavity.
The parotid glands lie between the skin and the masseter muscle, near the ears. They secrete saliva into the mouth through the parotid duct, which is located near the second upper molar tooth (Figure 12.3.3).
Saliva
Saliva is essentially (95.5 percent) water. The remaining 4.5 percent is a complex mixture of ions, glycoproteins, enzymes, growth factors, and waste products. Perhaps the most important ingredient in salvia from the perspective of digestion is the enzyme salivary amylase, which initiates the breakdown of carbohydrates. Food does not spend enough time in the mouth to allow all the carbohydrates to break down, but salivary amylase continues acting until it is inactivated by stomach acids. Bicarbonate and phosphate ions function as chemical buffers, maintaining saliva at a pH between 6.35 and 6.85. Salivary mucus helps lubricate food, facilitating movement in the mouth, bolus formation, and swallowing. Saliva contains immunoglobulin A, which prevents microbes from penetrating the epithelium, and lysozyme, which makes saliva antimicrobial. Saliva also contains epidermal growth factor, which might have given rise to the adage “a mother’s kiss can heal a wound.”
Each of the major salivary glands secretes a unique formulation of saliva according to its cellular makeup, for example the parotid glands secrete a watery solution that contains salivary amylase. The submandibular glands have cells like those of the parotid glands, as well as mucus-secreting cells. Therefore, saliva secreted by the submandibular glands also contains amylase but in a liquid thickened with mucus. The sublingual glands contain mostly mucous cells, and they secrete the thickest saliva with the least amount of salivary amylase.
Figure 12.3.3. Salivary glands. The major salivary glands are located outside the oral mucosa and deliver saliva into the mouth through ducts.
Homeostatic Imbalances
The Parotid Glands: Mumps
Infections of the nasal passages and pharynx can attack any salivary gland. The parotid glands are the usual site of infection with the virus that causes mumps (paramyxovirus). Mumps manifests by enlargement and inflammation of the parotid glands, causing a characteristic swelling between the ears and the jaw. Symptoms include fever and throat pain, which can be severe when swallowing acidic substances such as orange juice.
In about one-third of men who are past puberty, mumps also causes testicular inflammation, typically affecting only one testis and rarely resulting in sterility. With the increasing use and effectiveness of mumps vaccines, the incidence of mumps worldwide has decreased dramatically. Since the foundation of the vaccine in 1983, there have been no recorded childhood deaths due to mumps. Almost 95% of Australian toddlers (24 months old) were fully vaccinated against mumps in 2017, however the reported incidence rate has been gradually increasing in Australia. Majority of the outbreaks have been recorded in people 18-24 years of age, and it is speculated that this group missed their vaccination as children as they were born in a time when the disease rate was low. These outbreaks have been most commonly seen in the Aboriginal and Torres Straight Island populations.
Regulation of Salivation
The autonomic nervous system regulates salivation (the secretion of saliva). In the absence of food, parasympathetic stimulation keeps saliva flowing at just the right level for comfort as you speak, swallow, sleep, and generally go about life. Over-salivation can occur, for example, if you are stimulated by the smell of food, but that food is not available for you to eat. Drooling is an extreme instance of the overproduction of saliva. During times of stress, such as before speaking in public, sympathetic stimulation takes over, reducing salivation and producing the symptom of dry mouth often associated with anxiety. When you are dehydrated, salivation is reduced, causing the mouth to feel dry and prompting you to take action to quench your thirst.
Salivation can be stimulated by the sight, smell, and taste of food. It can even be stimulated by thinking about food. You might notice whether reading about food and salivation right now has had any effect on your production of saliva.
How does the salivation process work while you are eating? Food contains chemicals that stimulate taste receptors on the tongue, which send impulses to the superior and inferior salivatory nuclei in the brain stem. These two nuclei then send back parasympathetic impulses through fibres in the glossopharyngeal and facial nerves, which stimulate salivation. Even after you swallow food, salivation is increased to cleanse the mouth and to water down and neutralise any irritating chemical remnants, such as that hot sauce in your burrito. Most saliva is swallowed along with food and is reabsorbed, so that fluid is not lost.
The Teeth
The teeth, or dentes (singular = dens), are organs like bones that you use to tear, grind, and otherwise mechanically break down food.
Types of Teeth
During the course of your lifetime, you have two sets of teeth (one set of teeth is a dentition). Your 20 deciduous teeth, or baby teeth, first begin to appear at about 6 months of age. Between approximately age 6 and 12, these teeth are replaced by 32 permanent teeth. Moving from the centre of the mouth toward the side, these are as follows (Figure 12.3.4):
The eight incisors, four top and four bottom, are the sharp front teeth you use for biting into food.
The four cuspids (or canines) flank the incisors and have a pointed edge (cusp) to tear up food. These fang-like teeth are superb for piercing tough or fleshy foods.
Posterior to the cuspids are the eight premolars (or bicuspids), which have an overall flatter shape with two rounded cusps useful for mashing foods.
The most posterior and largest are the 12 molars, which have several pointed cusps used to crush food so it is ready for swallowing. The third members of each set of three molars, top and bottom, are commonly referred to as the wisdom teeth, because their eruption is commonly delayed until early adulthood. It is not uncommon for wisdom teeth to fail to erupt; that is, they remain impacted. In these cases, the teeth are typically removed by orthodontic surgery.
Figure 12.3.4. Permanent and deciduous teeth. This figure of two human dentitions shows the arrangement of teeth in the maxilla and mandible, and the relationship between the deciduous and permanent teeth.
Anatomy of a Tooth
The teeth are secured in the alveolar processes (sockets) of the maxilla and the mandible, with this joint known as a gomphosis. Gingivae (commonly called the gums) are soft tissues that line the alveolar processes and surround the necks of the teeth. Teeth are also held in their sockets by a connective tissue called the periodontal ligament.
The two main parts of a tooth are the crown, which is the portion projecting above the gum line, and the root, which is embedded within the maxilla and mandible. Both parts contain an inner pulp cavity, containing loose connective tissue through which run nerves and blood vessels. The region of the pulp cavity that runs through the root of the tooth is called the root canal. Surrounding the pulp cavity is dentin, a bone-like tissue. In the root of each tooth, the dentin is covered by an even harder bone-like layer called cementum. In the crown of each tooth, the dentin is covered by an outer layer of enamel, the hardest substance in the body (Figure 12.3.5).
Although enamel protects the underlying dentin and pulp cavity, it is still nonetheless susceptible to mechanical and chemical erosion, or what is known as tooth decay. The most common form, dental caries (cavities) develops when colonies of bacteria feeding on sugars in the mouth release acids that cause soft tissue inflammation and degradation of the calcium crystals of the enamel. The digestive functions of the mouth are summarised in Table 12.3.1.
Figure 12.3.5. The Structure of the tooth. This longitudinal section through a molar in its alveolar socket shows the relationships between enamel, dentin, and pulp.
Table 12.3.1. Digestive Functions of the Mouth
Structure
Action
Outcome
Lips and cheeks
Confine food between teeth
Food is chewed evenly during mastication
Salivary glands
Secrete saliva
Moisten and lubricate the lining of the mouth and pharynx
Moisten, soften and dissolve food
Clean the mouth and teeth
Salivary amylase breaks down starch
Tongue’s extrinsic muscles
Move tongue sideways, and in and out
Manipulate food for chewing
Shape food into a bolus
Manipulate food for swallowing
Tongue’s intrinsic muscles
Change tongue shape
Manipulate food for swallowing
Taste buds
Sense food in mouth and sense taste
Nerve impulses from taste buds are conducted to salivary nuclei in the brain stem and then to salivary glands, stimulating saliva secretion
Lingual glands
Secrete lingual lipase
Activated in the stomach
Breakdown triglycerides into fatty acids and diglycerides
Teeth
Shred and crush food
Break down solid food into smaller particles for deglutition
The Pharynx
The pharynx (throat) is involved in both digestion and respiration. It receives food and air from the mouth, and air from the nasal cavities. When food enters the pharynx, involuntary muscle contractions close off the air passageways.
A short tube of skeletal muscle lined with a mucous membrane, the pharynx runs from the posterior oral and nasal cavities to the opening of the oesophagus and larynx. It has three subdivisions. The most superior, the nasopharynx, is involved only in breathing and speech. The other two subdivisions, the oropharynx and the laryngopharynx, are used for both breathing and digestion. The oropharynx begins inferior to the nasopharynx and is continuous below with the laryngopharynx (Figure 12.3.6). The inferior border of the laryngopharynx connects to the oesophagus, whereas the anterior portion connects to the larynx, allowing air to flow into the bronchial tree.
Figure 12.3.6. Pharynx. The pharynx runs from the nostrils to the oesophagus and the larynx.
Histologically, the wall of the oropharynx is like that of the oral cavity. The mucosa includes a stratified squamous epithelium that is endowed with mucus-producing glands. During swallowing, the elevator skeletal muscles of the pharynx contract, raising and expanding the pharynx to receive the bolus of food. Once received, these muscles relax and the constrictor muscles of the pharynx contract, forcing the bolus into the oesophagus and initiating peristalsis.
Usually during swallowing, the soft palate and uvula rise reflexively to close off the entrance to the nasopharynx. At the same time, the larynx is pulled superiorly and the cartilaginous epiglottis, its most superior structure, folds inferiorly, covering the glottis (the opening to the larynx); this process effectively blocks access to the trachea and bronchi. When the food “goes down the wrong way,” it goes into the trachea. When food enters the trachea, the reaction is to cough, which usually forces the food up and out of the trachea, and back into the pharynx.
The Oesophagus
The oesophagus is a muscular tube that connects the pharynx to the stomach. It is approximately 25.4 cm in length, located posterior to the trachea, and remains in a collapsed form when not engaged in swallowing. As you can see in Figure 12.3.7, the oesophagus runs a mainly straight route through the mediastinum of the thorax. To enter the abdomen, the oesophagus penetrates the diaphragm through an opening called the oesophagus hiatus.
Passage of Food Through the Oesophagus
The upper oesophagus sphincter, which is continuous with the inferior pharyngeal constrictor, controls the movement of food from the pharynx into the oesophagus. The upper two-thirds of the oesophagus consists of both smooth and skeletal muscle fibres, with the latter fading out in the bottom third of the oesophagus. Rhythmic waves of peristalsis, which begin in the upper oesophagus, propel the bolus of food toward the stomach. Meanwhile, secretions from the oesophageal mucosa lubricate the oesophagus and food. Food passes from the oesophagus into the stomach at the lower oesophageal sphincter (also called the gastroesophageal or cardiac sphincter). Recall that sphincters are muscles that surround tubes and serve as valves, closing the tube when the sphincters contract and opening it when they relax. The lower oesophageal sphincter relaxes to let food pass into the stomach, and then contracts to prevent stomach acids from backing up into the oesophagus. Surrounding this sphincter is the muscular diaphragm, which helps close off the sphincter when no food is being swallowed. When the lower oesophageal sphincter does not completely close, the stomach’s contents can reflux (that is, back up into the oesophagus), causing heartburn or gastroesophageal reflux disease (GERD).
Figure 12.3.7. Oesophagus. The upper oesophageal sphincter controls the movement of food from the pharynx to the oesophagus. The lower oesophageal sphincter controls the movement of food from the oesophagus to the stomach.
Histology of the Oesophagus
The mucosa of the oesophagus is made up of an epithelial lining that contains non-keratinised, stratified squamous epithelium, with a layer of basal and parabasal cells. This epithelium protects against erosion from food particles. The mucosa’s lamina propria contains mucus-secreting glands. The muscularis layer changes according to location: In the upper third of the oesophagus, the muscularis is skeletal muscle. In the middle third, it is both skeletal and smooth muscle. In the lower third, it is smooth muscle. As mentioned previously, the most superficial layer of the oesophagus is called the adventitia, not the serosa. In contrast to the stomach and intestines, the loose connective tissue of the adventitia is not covered by a fold of visceral peritoneum. The digestive functions of the oesophagus are identified in Table 12.3.2.
Table 12.3.2. Digestive Functions of the Oesophagus
Action
Outcome
Upper oesophageal
sphincter relaxation
Allows the bolus to move from the laryngopharynx to the oesophagus
Peristalsis
Propels the bolus through the oesophagus
Lower oesophageal sphincter relaxation
Allows the bolus to move from the oesophagus into the stomach and prevents chime from entering the oesophagus
Mucus secretion
Lubricates the oesophagus, allowing easy passage of the bolus
Deglutition
Deglutition is another word for swallowing—the movement of food from the mouth to the stomach. The entire process takes about four to eight seconds for solid or semisolid food, and about one second for very soft food and liquids. Although this sounds quick and effortless, deglutition is, in fact, a complex process that involves both the skeletal muscle of the tongue and the muscles of the pharynx and oesophagus. It is aided by the presence of mucus and saliva. There are three stages in deglutition: the voluntary phase, the pharyngeal phase and the oesophageal phase (Figure 12.3.8). The autonomic nervous system controls the latter two phases.
Figure 12.3.8. Deglutition. Deglutition includes the voluntary phase and two involuntary phases: the pharyngeal phase and the oesophageal phase.
The Voluntary Phase
The voluntary phase of deglutition (also known as the oral or buccal phase) is so called because you can control when you swallow food. In this phase, chewing has been completed and swallowing is set in motion. The tongue moves upward and backward against the palate, pushing the bolus to the back of the oral cavity and into the oropharynx. Other muscles keep the mouth closed and prevent food from falling out. At this point, the two involuntary phases of swallowing begin.
The Pharyngeal Phase
In the pharyngeal phase, stimulation of receptors in the oropharynx sends impulses to the deglutition centre (a collection of neurons that controls swallowing) in the medulla oblongata. Impulses are then sent back to the uvula and soft palate, causing them to move upward and close off the nasopharynx. The laryngeal muscles also constrict to prevent aspiration of food into the trachea. At this point, deglutition apnoea takes place, which means that breathing ceases for a very brief time. Contractions of the pharyngeal constrictor muscles move the bolus through the oropharynx and laryngopharynx. Relaxation of the upper oesophageal sphincter then allows food to enter the oesophagus.
The Oesophageal Phase
The entry of food into the oesophagus marks the beginning of the oesophageal phase of deglutition and the initiation of peristalsis. As in the previous phase, the complex neuromuscular actions are controlled by the medulla oblongata. Peristalsis propels the bolus through the oesophagus and toward the stomach. The circular muscle layer of the muscularis contracts, pinching the oesophageal wall and forcing the bolus forward. At the same time, the longitudinal muscle layer of the muscularis also contracts, shortening this area and pushing out its walls to receive the bolus. In this way, a series of contractions keeps moving food toward the stomach. When the bolus nears the stomach, distention of the oesophagus initiates a short reflex relaxation of the lower oesophageal sphincter that allows the bolus to pass into the stomach. During the oesophageal phase, oesophageal glands secrete mucus that lubricates the bolus and minimises friction.
Section Review
In the mouth, the tongue and the teeth begin mechanical digestion, and saliva begins chemical digestion. The pharynx, which plays roles in breathing and vocalisation as well as digestion, runs from the nasal and oral cavities superiorly to the oesophagus inferiorly (for digestion) and to the larynx anteriorly (for respiration). During deglutition (swallowing), the soft palate rises to close off the nasopharynx, the larynx elevates, and the epiglottis folds over the glottis. The oesophagus includes an upper oesophageal sphincter made of skeletal muscle, which regulates the movement of food from the pharynx to the oesophagus. It also has a lower oesophageal sphincter, made of smooth muscle, which controls the passage of food from the oesophagus to the stomach. Cells in the oesophageal wall secrete mucus that eases the passage of the food bolus.
Label on a diagram the four main regions of the stomach, its curvatures and its sphincters
Identify the four main types of secreting cells in gastric glands and their important products
Explain why the stomach does not digest itself
Describe the mechanical and chemical digestion of food entering the stomach
Although a minimal amount of carbohydrate digestion occurs in the mouth, chemical digestion really gets underway in the stomach. An expansion of the gastrointestinal tract that lies immediately inferior to the oesophagus, the stomach links the oesophagus to the first part of the small intestine (the duodenum) and is fixed in place at its oesophageal and duodenal ends. In between, however, it can be a highly active structure, contracting and continually changing position and size. These contractions provide mechanical assistance to digestion. The empty stomach is only about the size of your fist, but can stretch to hold as much as 4 litres of food and fluid, or more than 75 times its empty volume, and then return to its resting size when empty. Although you might think that the size of a person’s stomach is related to how much food that individual consumes, body weight does not correlate with stomach size. Rather, when you eat greater quantities of food—such as at holiday dinner—you stretch the stomach more than when you eat less.
Popular culture tends to refer to the stomach as the location where all digestion takes place. Of course, this is not true. An important function of the stomach is to serve as a temporary holding chamber. You can ingest a meal far more quickly than it can be digested and absorbed by the small intestine. Thus, the stomach holds food and parses only small amounts into the small intestine at a time. Foods are not processed in the order they are eaten; rather, they are mixed together with digestive secretions in the stomach until they are converted into chyme, which is released into the small intestine.
As you will see in the sections that follow, the stomach plays several important roles in chemical digestion, including the continued digestion of carbohydrates and the initial digestion of proteins and triglycerides. Little if any nutrient absorption occurs in the stomach, except for the negligible amount of nutrients in alcohol.
Structure
There are four main regions in the stomach: the cardia, fundus, body, and pylorus (Figure 12.4.1). The cardia (or cardiac region) is the point where the oesophagus connects to the stomach and through which food passes into the stomach. Located inferior to the diaphragm, above and to the left of the cardia, is the dome-shaped fundus. Below the fundus is the body, the main part of the stomach. The funnel-shaped pylorus connects the stomach to the duodenum. The wider end of the funnel, the pyloric antrum, connects to the body of the stomach. The narrower end is called the pyloric canal, which connects to the duodenum. The smooth muscle pyloric sphincter is located at this latter point of connection and controls stomach emptying. In the absence of food, the stomach deflates inward, and its mucosa and submucosa fall into a large fold called a ruga (rugae – plural)
Figure 12.4.1. Stomach. The stomach has four major regions: the cardia, fundus, body, and pylorus. The addition of an inner oblique smooth muscle layer gives the muscularis the ability to vigorously churn and mix food.
The convex lateral surface of the stomach is called the greater curvature; the concave medial border is the lesser curvature. The stomach is held in place by the lesser omentum, which extends from the liver to the lesser curvature, and the greater omentum, which runs from the greater curvature to the posterior abdominal wall.
Histology
The wall of the stomach is made of the same four layers as most of the rest of the gastrointestinal tract, but with adaptations to the mucosa and muscularis for the unique functions of this organ. In addition to the typical circular and longitudinal smooth muscle layers, the muscularis has an inner oblique smooth muscle layer (Figure 12.4.2). As a result, in addition to moving food through the tract, the stomach can vigorously churn food, mechanically breaking it down into smaller particles.
Figure 12.4.2. Histology of the stomach. The stomach wall is adapted for the functions of the stomach. In the epithelium, gastric pits lead to gastric glands that secrete gastric juice. The gastric glands (one gland is shown enlarged on the right) contain diverse types of cells that secrete a variety of enzymes, including hydrochloride acid, which activates the protein-digesting enzyme pepsin.
The stomach mucosa’s epithelial lining consists only of surface mucus cells, which secrete a protective coat of alkaline mucus. A vast number of gastric pits dot the surface of the epithelium, giving it the appearance of a well-used pincushion, and mark the entry to each gastric gland, which secretes a complex digestive fluid referred to as gastric juice.
Although the walls of the gastric pits are made up primarily of mucus cells, the gastric glands are made up of diverse types of cells. The glands of the cardia and pylorus are composed primarily of mucus-secreting cells. Cells that make up the pyloric antrum secrete mucus and several hormones, including the majority of the stimulatory hormone, gastrin. The much larger glands of the fundus and body of the stomach, the site of most chemical digestion, produce most of the gastric secretions. These glands are made up of a variety of secretory cells. These include parietal cells, chief cells, mucous neck cells, and enteroendocrine cells.
Parietal cells—Located primarily in the middle region of the gastric glands are parietal cells, which are among the most highly differentiated of the body’s epithelial cells. These large cells produce both hydrochloric acid (HCl) and intrinsic factor. HCl is responsible for the high acidity (pH 1.5 to 3.5) of the stomach contents and is needed to activate the protein-digesting enzyme, pepsin. The acidity also kills much of the bacteria you ingest with food and helps to denature proteins, making them more available for enzymatic digestion. Intrinsic factor is a glycoprotein necessary for the absorption of vitamin B12 in the small intestine.
Chief cells—Located primarily in the basal regions of gastric glands are chief cells, which secrete pepsinogen, the inactive proenzyme form of pepsin. HCl is necessary for the conversion of pepsinogen to pepsin.
Mucous neck cells—Gastric glands in the upper part of the stomach contain mucous neck cells that secrete thin, acidic mucus that is much different from the mucus secreted by the goblet cells of the surface epithelium. The role of this mucus is not currently known.
Enteroendocrine cells—Finally, enteroendocrine cells found in the gastric glands secrete various hormones into the interstitial fluid of the lamina propria. These include gastrin, which is released mainly by enteroendocrine G cells.
Table 12.4.1 describes the digestive functions of important hormones secreted by the stomach.
Table 12.4.1. Hormones secreted by stomach
Hormone
Production site
Production stimulus
Target organ
Action
Gastrin
Stomach mucosa, mainly G cells of the pyloric antrum
Presence of peptides and amino acids in stomach
Stomach
Increases secretion by gastric glands; promotes gastric emptying
Stomach mucosa, mainly G cells of the pyloric antrum
Presence of peptides and amino acids in stomach
Ileocaecal valve
Relaxes valve
Stomach mucosa, mainly G cells of the pyloric antrum
Presence of peptides and amino acids in stomach
Large intestine
Triggers mass movements
Ghrelin
Stomach mucosa, mainly fundus
Fasting state (levels increase just prior to meals)
Hypothalamus
Regulates food intake, primarily by stimulating hunger and satiety
Histamine
Stomach mucosa
Presence of food in stomach
Stomach
Stimulates parietal cells to release
Somatostatin
Mucosa of stomach, especially pyloric antrum; also duodenum
Presence of food in the stomach; sympathetic axon stimulation
Stomach
Restricts all gastric secretions, gastric motility, and emptying
Mucosa of stomach, especially pyloric antrum; also duodenum
Presence of food in stomach; sympathetic axon stimulation
Pancreas
Restricts pancreatic secretions
Mucosa of stomach, especially pyloric antrum; also duodenum
Presence of food in the stomach; sympathetic axon stimulation
Small intestine
Reduces intestinal absorption by reducing blood flow
Gastric Secretion
The secretion of gastric juice is controlled by both nerves and hormones. Stimuli in the brain, stomach, and small intestine activate or inhibit gastric juice production. This is why the three phases of gastric secretion are called the cephalic, gastric, and intestinal phases (Figure 12.4.3). However, once gastric secretion begins, all three phases can occur simultaneously.
Figure 12.4.3. The three phases of gastric secretion. Gastric secretion occurs in three phases: cephalic, gastric and intestinal. During each phase, the secretion of gastric juice can be stimulated or inhibited.
The cephalic phase (reflex phase) of gastric secretion, which is brief, takes place before food enters the stomach. The smell, taste, sight, or thought of food triggers this phase. For example, when you bring a piece of sushi to your lips, impulses from receptors in your taste buds or the nose are relayed to your brain, which returns signals that increase gastric secretion to prepare your stomach for digestion. This enhanced secretion is a conditioned reflex, meaning it occurs only if you like or want a particular food. Depression and loss of appetite can suppress the cephalic reflex.
The gastric phase of secretion lasts 3 to 4 hours and is set in motion by local neural and hormonal mechanisms triggered by the entry of food into the stomach. For example, when your sushi reaches the stomach, it creates distention that activates the stretch receptors. This stimulates parasympathetic neurons to release acetylcholine, which then provokes increased secretion of gastric secretion (‘juice’). Partially digested proteins, caffeine, and rising pH stimulate the release of gastrin from enteroendocrine G cells, which in turn induces parietal cells to increase their production of HCl, which is needed to create an acidic environment for the conversion of pepsinogen to pepsin, and protein digestion. Additionally, the release of gastrin activates vigorous smooth muscle contractions. However, it should be noted that the stomach does have a natural means of avoiding excessive acid secretion and potential heartburn. Whenever pH levels drop too low, cells in the stomach react by suspending HCl secretion and increasing mucous secretions.
The intestinal phase of gastric secretion has both excitatory and inhibitory elements. The duodenum has a key role in regulating the stomach and its emptying. When partially digested food fills the duodenum, intestinal mucosal cells release a hormone called intestinal (enteric) gastrin, which further excites gastric secretions. This stimulatory activity is brief, however, because when the intestine distends with chyme, the enterogastric reflex inhibits secretion. One of the effects of this reflex is to close the pyloric sphincter, which blocks additional acidic chyme from entering the duodenum.
The Mucosal Barrier
The mucosa of the stomach is exposed to the highly corrosive acidity of gastric juice. Gastric enzymes that can digest protein can also digest the stomach itself. The stomach is protected from self-digestion by the mucosal barrier. This barrier has several components. First, the stomach wall is covered by a thick coating of bicarbonate-rich mucus. This mucus forms a physical barrier, and its bicarbonate ions neutralise acid. Second, the epithelial cells of the stomach’s mucosa meet at tight junctions, which block gastric juice from penetrating the underlying tissue layers. Finally, stem cells located where gastric glands join the gastric pits quickly replace damaged epithelial mucosal cells, when the epithelial cells are shed. In fact, the surface epithelium of the stomach is completely replaced every 3 to 6 days.
Homeostatic Imbalances
Ulcers: When the Mucosal Barrier Breaks Down
As effective as the mucosal barrier is, it is not a “fail-safe” mechanism. Sometimes, gastric juice eats away at the superficial lining of the stomach mucosa, creating erosions, which mostly heal on their own. Deeper and larger erosions are called ulcers.
Why does the mucosal barrier break down? Several factors can interfere with its ability to protect the stomach lining. The majority of all ulcers are caused by either excessive intake of non-steroidal anti-inflammatory drugs (NSAIDs), including aspirin, or more commonly infection with Helicobacter pylori. Spicy foods and stress do not cause gastric ulcers but could worsen the symptoms.
Antacids help relieve symptoms of ulcers such as “burning” pain and indigestion. When ulcers are caused by NSAID use, switching to other classes of pain relievers allows healing. When caused by H. pylori infection, antibiotics are effective.
A potential complication of ulcers is perforation: Perforated ulcers create a hole in the stomach wall, resulting in peritonitis (inflammation of the peritoneum). These ulcers must be repaired surgically.
Digestive Functions of the Stomach
The stomach participates in all the digestive activities except for ingestion and defaecation. Although almost all absorption takes place in the small intestine, the stomach does absorb some nonpolar substances, such as alcohol and aspirin.
Mechanical Digestion
Within a few moments after food after enters your stomach, mixing waves begin to occur at intervals of approximately 20 seconds. A mixing wave is a unique type of peristalsis that mixes and softens the food with gastric secretions to create chyme. The initial mixing waves are gentle, but these are followed by more intense waves, starting at the body of the stomach and increasing in force as they reach the pylorus. It is fair to say that long before your sushi exits through the pyloric sphincter, it bears little resemblance to the sushi you ate.
The pylorus, which holds around 30 mL of chyme, acts as a filter, permitting only liquids and small food particles to pass through the mostly, but not fully, closed pyloric sphincter. In a process called gastric emptying, rhythmic mixing waves force about 3 mL of chyme at a time through the pyloric sphincter and into the duodenum. Release of a greater amount of chyme at one time would overwhelm the capacity of the small intestine to handle it. The rest of the chyme is pushed back into the body of the stomach, where it continues mixing. This process is repeated when the next mixing waves force more chyme into the duodenum.
Gastric emptying is regulated by both the stomach and the duodenum. The presence of chyme in the duodenum activates receptors that inhibit gastric secretion. This prevents additional chyme from being released by the stomach before the duodenum is ready to process it.
Chemical Digestion
The fundus plays a key role, because it stores both undigested food and gases that are released during the process of chemical digestion. Food may sit in the fundus of the stomach for a while before being mixed with the chyme. While the food is in the fundus, the digestive activities of salivary amylase continue until the food begins mixing with the acidic chyme. Ultimately, mixing waves incorporate this food with the chyme, the acidity of which inactivates salivary amylase and activates lingual lipase. Lingual lipase then begins breaking down triglycerides into free fatty acids, and mono- and diglycerides.
The breakdown of protein begins in the stomach through the actions of HCl and the enzyme pepsin. During infancy, gastric glands also produce rennin, an enzyme that helps digest milk protein.
Its numerous digestive functions notwithstanding, there is only one stomach function necessary to life: the production of intrinsic factor. The intestinal absorption of vitamin B12, which is necessary for both the production of mature red blood cells and normal neurological functioning, cannot occur without intrinsic factor. People who undergo total gastrectomy (stomach removal)—for life-threatening stomach cancer, for example—can survive with minimal digestive dysfunction if they receive vitamin B12 injections.
The contents of the stomach are completely emptied into the duodenum within 2 to 4 hours after you eat a meal. Different types of food take different amounts of time to process. Foods heavy in carbohydrates empty fastest, followed by high-protein foods. Meals with a high triglyceride content remain in the stomach the longest. Since enzymes in the small intestine digest fats slowly, food can stay in the stomach for 6 hours or longer when the duodenum is processing fatty chyme. However, note that this is still a fraction of the 24 to 72 hours that full digestion typically takes from start to finish.
Section Review
The stomach participates in all digestive activities except ingestion and defaecation. It vigorously churns food. It secretes gastric secretions that break down food and absorbs certain drugs, including aspirin and some alcohol. The stomach begins the digestion of protein and continues the digestion of carbohydrates and fats. It stores food as an acidic liquid called chyme and releases it gradually into the small intestine through the pyloric sphincter.
Compare and contrast the location and gross anatomy of the small and large intestines
Identify three main adaptations of the small intestine wall that increase its absorptive capacity
Describe the mechanical and chemical digestion of chyme upon its release into the small intestine
List three features unique to the wall of the large intestine and identify their contributions to its function
Identify the beneficial roles of the microbiome in digestive system functioning
Trace the pathway of food waste from its point of entry into the large intestine through its exit from the body as faeces
The word intestine is derived from a Latin root meaning “internal,” and indeed, the two organs together nearly fill the interior of the abdominal cavity. In addition, called the small and large bowel, or colloquially the “guts,” they constitute the greatest mass and length of the gastrointestinal tract and, except for ingestion, perform all digestive system functions.
The Small Intestine
Chyme released from the stomach enters the small intestine, which is the primary digestive organ in the body. Not only is this where most digestion occurs, it is also where practically all absorption occurs. The longest part of the gastrointestinal tract, the small intestine is about 3.05 metres long in a living person (but about twice as long in a cadaver due to the loss of muscle tone). Since this makes it about five times longer than the large intestine, you might wonder why it is called “small.” In fact, its name derives from its relatively smaller diameter of only about 2.54 cm, compared with 7.62 cm for the large intestine. As we will see shortly, in addition to its length, the folds and projections of the lining of the small intestine work to give it an enormous surface area, which is approximately 200 m2, more than 100 times the surface area of your skin. This large surface area is necessary for complex processes of digestion and absorption that occur within it.
Structure
The coiled tube of the small intestine is subdivided into three regions. From proximal (at the stomach) to distal, these are the duodenum, jejunum and ileum (Figure 12.5.1).
The shortest region is the 25.4-cm duodenum, which begins at the pyloric sphincter. Just past the pyloric sphincter, it bends posteriorly behind the peritoneum, becoming retroperitoneal, and then makes a C-shaped curve around the head of the pancreas before ascending anteriorly again to return to the peritoneal cavity and join the jejunum. The duodenum can therefore be subdivided into four segments: the superior, descending, horizontal, and ascending duodenum.
Of particular interest is the hepatopancreatic ampulla (ampulla of Vater). Located in the duodenal wall, the ampulla marks the transition from the anterior portion of the gastrointestinal tract to the mid-region, and is where the bile duct (through which bile passes from the liver) and the main pancreatic duct (through which pancreatic juice passes from the pancreas) join. This ampulla opens into the duodenum at a tiny volcano-shaped structure called the major duodenal papilla. The hepatopancreatic sphincter (sphincter of Oddi) regulates the flow of both bile and pancreatic secretions from the ampulla into the duodenum.
Figure 12.5.1. Small intestine. The three regions of the small intestine are the duodenum, jejunum, and ileum.
The jejunum is about 0.9 metres long (in the living adult) and runs from the duodenum to the ileum. Jejunum means “empty” in Latin and supposedly was so named by the ancient Greeks who noticed it was always empty at death. No clear demarcation exists between the jejunum and the final segment of the small intestine, the ileum.
The ileum is the longest part of the small intestine, measuring about 1.8 metres in length. It is thicker, more vascular, and has more developed mucosal folds than the jejunum. The ileum joins the caecum, the first portion of the large intestine, at the ileocaecal sphincter (or valve). The jejunum and ileum are tethered to the posterior abdominal wall by the mesentery. The large intestine frames these three parts of the small intestine.
Parasympathetic nerve fibres from the vagus nerve and sympathetic nerve fibres from the thoracic splanchnic nerve provide extrinsic innervation to the small intestine. The superior mesenteric artery is its main arterial supply. Veins run parallel to the arteries and drain into the superior mesenteric vein. Nutrient-rich blood from the small intestine is then carried to the liver via the hepatic portal vein.
Histology
The wall of the small intestine is composed of the same four layers typically present in the gastrointestinal system. However, three features of the mucosa and submucosa are unique. These features, which increase the absorptive surface area of the small intestine more than 600-fold, include circular folds, villi, and microvilli (Figure 12.5.2). These adaptations are most abundant in the proximal two-thirds of the small intestine, where most of the absorption occurs.
Also called plicae circulares, a circular fold resulting in a deep ridge in the mucosa and submucosa. Beginning near the proximal part of the duodenum and ending near the middle of the ileum, these folds facilitate absorption. Their shape causes the chyme to spiral, rather than move in a straight line, through the small intestine. Spiralling slows the movement of chyme and provides the time needed for nutrients to be fully absorbed.
Villi
Within the circular folds are small (0.5–1 mm long) hairlike vascularised projections called villi (singular = villus) that give the mucosa a furry texture. There are about 20 to 40 villi per square millimetre, increasing the surface area of the epithelium tremendously. The mucosal epithelium, primarily composed of absorptive cells, covers the villi. In addition to muscle and connective tissue to support its structure, each villus contains a capillary bed composed of one arteriole and one venule, as well as a lymphatic capillary called a lacteal. The breakdown products of carbohydrates and proteins (sugars and amino acids) can enter the bloodstream directly, but lipid breakdown products are absorbed by the lacteals and transported to the bloodstream via the lymphatic system.
Microvilli
As their name suggests, microvilli (singular = microvillus) are much smaller (1 µm) than villi. They are cylindrical apical surface extensions of the plasma membrane of the mucosa’s epithelial cells and are supported by microfilaments within those cells. Although their small size makes it difficult to see each microvillus, their combined microscopic appearance suggests a mass of bristles, which is termed the brush border. Fixed to the surface of the microvilli membranes are enzymes that finish digesting carbohydrates and proteins. There are an estimated 200 million microvilli per square millimetre of small intestine, expanding the surface area of the plasma membrane and thus greatly enhancing absorption.
Intestinal Glands
In addition to the three specialised absorptive features just discussed, the mucosa between the villi is dotted with deep crevices that each lead into a tubular intestinal gland (crypt of Lieberkühn), which is formed by cells that line the crevices (see Figure 12.5.2). These produce intestinal juice, a slightly alkaline (pH 7.4 to 7.8) mixture of water and mucus. Each day, about 0.95 to 1.9 litres are secreted in response to the distention of the small intestine or the irritating effects of chyme on the intestinal mucosa.
The submucosa of the duodenum is the only site of the complex mucus-secreting duodenal glands (Brunner’s glands), which produce a bicarbonate-rich alkaline mucus that buffers the acidic chyme as it enters from the stomach.
The roles of the cells in the small intestinal mucosa are detailed in Table 12.5.1.
Table 12.5.1. Cells of the small intestinal mucosa
Cell type
Location in the mucosa
Function
Absorptive
Epithelium/intestinal glands
Digestion and absorption of nutrients in chyme
Goblet
Epithelium/intestinal glands
Secretion of mucus
Paneth
Intestinal glands
Secretion of the bactericidal enzyme lysosome; phagocytosis
G cells
Intestinal glands of duodenum
Secretion of the hormone intestinal gastrin
I cells
Intestinal glands of duodenum
Secretion of the hormone cholecystokinin, which stimulates release of pancreatic secretions and bile
K cells
Intestinal glands
Secretion of the hormone glucose-dependent insulinotropic peptide, which stimulates the release of insulin
M cells
Intestinal glands of duodenum and jejunum
Secretion of the hormone motilin, which accelerates gastric emptying, stimulates intestinal peristalsis, and stimulates the production of pepsin
The lamina propria of the small intestine mucosa is studded with MALT. In addition to solitary lymphatic nodules, aggregations of intestinal MALT, which are typically referred to as Peyer’s patches, are concentrated in the distal ileum, and serve to keep bacteria from entering the bloodstream. Peyer’s patches are most prominent in young people and become less distinct as you age, which coincides with the general activity of our immune system.
Mechanical Digestion in the Small Intestine
The movement of intestinal smooth muscles includes both segmentation and a form of peristalsis called migrating motility complexes. The kind of peristaltic mixing waves seen in the stomach are not observed here.
If you could see into the small intestine when it was going through segmentation, it would look as if the contents were being shoved incrementally back and forth, as the rings of smooth muscle repeatedly contract and then relax. Segmentation in the small intestine does not force chyme through the tract. Instead, it combines the chyme with digestive secretions and pushes food particles against the mucosa to be absorbed. The duodenum is where the most rapid segmentation occurs, at a rate of about 12 times per minute. In the ileum, segmentations are only about eight times per minute (Figure 12.5.3).
Figure 12.5.3. Segmentation. Segmentation separates chyme and then pushes it back together, mixing it and providing time for digestion and absorption.
When most of the chyme has been absorbed, the small intestinal wall becomes less distended. At this point, the localised segmentation process is replaced by transport movements. The duodenal mucosa secretes the hormone motilin, which initiates peristalsis in the form of a migrating motility complex. These complexes, which begin in the duodenum, force chyme through a short section of the small intestine and then stop. The next contraction begins a little bit farther down than the first, forces chyme a bit farther through the small intestine, then stops. These complexes move slowly down the small intestine, forcing chyme on the way, taking around 90 to 120 minutes to finally reach the end of the ileum. At this point, the process is repeated, starting in the duodenum.
The ileocaecal valve, a sphincter, is usually in a constricted state, but when motility in the ileum increases, this sphincter relaxes, allowing food residue to enter the first portion of the large intestine, the cecum. Relaxation of the ileocaecal sphincter is controlled by both nerves and hormones. First, digestive activity in the stomach provokes the gastroileal reflex, which increases the force of ileal segmentation. Second, the stomach releases the hormone gastrin, which enhances ileal motility, thus relaxing the ileocaecal sphincter. After chyme passes through, backward pressure helps close the sphincter, preventing backflow into the ileum. Because of this reflex, your lunch is completely emptied from your stomach and small intestine by the time you eat your dinner. It takes about three to five hours for all chyme to leave the small intestine.
Chemical Digestion in the Small Intestine
The digestion of proteins and carbohydrates, which partially occurs in the stomach, is completed in the small intestine with the aid of intestinal and pancreatic secretions. Lipids arrive in the intestine largely undigested, so much of the focus here is on lipid digestion, which is facilitated by bile and the enzyme pancreatic lipase.
Moreover, intestinal juice combines with pancreatic juice to provide a liquid medium that facilitates absorption. The intestine is also where most water is absorbed, via osmosis. The small intestine’s absorptive cells also synthesise digestive enzymes and then place them in the plasma membranes of the microvilli. This distinguishes the small intestine from the stomach; that is, enzymatic digestion occurs not only in the lumen, but also on the luminal surfaces of the mucosal cells.
For optimal chemical digestion, chyme must be delivered from the stomach slowly and in small amounts. This is because chyme from the stomach is typically hypertonic, and if large quantities were forced all at once into the small intestine, the resulting osmotic water loss from the blood into the intestinal lumen would result in potentially life-threatening low blood volume. In addition, continued digestion requires an upward adjustment of the low pH of stomach chyme, along with rigorous mixing of the chyme with bile and pancreatic secretions. Both processes take time, so the pumping action of the pylorus must be carefully controlled to prevent the duodenum from being overwhelmed with chyme.
Disorders of the Small Intestine: Lactose Intolerance
Lactose intolerance is a condition characterised by indigestion caused by dairy products. It occurs when the absorptive cells of the small intestine do not produce enough lactase, the enzyme that digests the milk sugar lactose. In most mammals, lactose intolerance increases with age. In contrast, some human populations, most notably Caucasians, can maintain the ability to produce lactase as adults.
In people with lactose intolerance, the lactose in chyme is not digested. Bacteria in the large intestine ferment the undigested lactose, a process that produces gas. In addition to gas, symptoms include abdominal cramps, bloating, and diarrhea. Symptom severity ranges from mild discomfort to severe pain; however, symptoms resolve once the lactose is eliminated in faeces.
The hydrogen breath test is used to help diagnose lactose intolerance. Lactose-tolerant people have very little hydrogen in their breath. Those with lactose intolerance exhale hydrogen, which is one of the gases produced by the bacterial fermentation of lactose in the colon. After the hydrogen is absorbed from the intestine, it is transported through blood vessels into the lungs. There are a number of lactose-free dairy products available in grocery stores. In addition, dietary supplements are available. Taken with food, they provide lactase to help digest lactose.
The Large Intestine
The large intestine is the terminal part of the gastrointestinal tract. The primary function of this organ is to finish absorption of nutrients and water, synthesise certain vitamins, form faeces, and eliminate faeces from the body.
Structure
The large intestine runs from the appendix to the anus. It frames the small intestine on three sides. Despite its being about one-half as long as the small intestine, it is called large because it is more than twice the diameter of the small intestine.
Subdivisions
The large intestine is subdivided into four main regions: the caecum, the colon, the rectum, and the anus. The ileocaecal valve, located at the opening between the ileum and the large intestine, controls the flow of chyme from the small intestine to the large intestine.
Caecum
The first part of the large intestine is the caecum, a sac-like structure that is suspended inferior to the ileocaecal valve. It is about 6 cm long, receives the contents of the ileum, and continues the absorption of water and salts. The appendix (or vermiform appendix) is a winding tube that attaches to the caecum. Although the 7.6-cm long appendix contains lymphoid tissue, suggesting an immunologic function, this organ is considered vestigial. However, at least one recent report postulates a survival advantage conferred by the appendix: In diarrhoeal illness, the appendix may serve as a bacterial reservoir to repopulate the enteric bacteria for those surviving the initial phases of the illness. Moreover, its twisted anatomy provides a haven for the accumulation and multiplication of enteric bacteria. The mesoappendix, the mesentery of the appendix, tethers it to the mesentery of the ileum.
Colon
The caecum blends seamlessly with the colon. Upon entering the colon, the food residue first travels up the ascending colon on the right side of the abdomen. At the inferior surface of the liver, the colon bends to form the right colic flexure (hepatic flexure) and becomes the transverse colon. The region defined as hindgut begins with the last third of the transverse colon and continues on. Food residue passing through the transverse colon travels across to the left side of the abdomen, where the colon angles sharply immediately inferior to the spleen, at the left colic flexure (splenic flexure). From there, food residue passes through the descending colon, which runs down the left side of the posterior abdominal wall. After entering the pelvis inferiorly, it becomes the s-shaped sigmoid colon, which extends medially to the midline (Figure 12.5.4). The ascending and descending colon, and the rectum (discussed next) are located in the retroperitoneum. The transverse and sigmoid colon are tethered to the posterior abdominal wall by the mesocolon.
Figure 12.5.4. Large intestine. The large intestine includes the cecum, colon, and rectum.
Homeostatic Imbalances
Colorectal Cancer
Colorectal cancer is one of the most prevalent cancers in the western world. In 2019, almost 16,400 new cases were recorded in Australia, with over 5,500 deaths. In Australia, the 5-year relative survival of patients suffering from colorectal cancer is approximately 70%. People with a family history of colorectal cancer are at increased risk. Smoking, excessive alcohol consumption and a diet high in animal fat and protein also increase the risk. Despite popular opinion to the contrary, studies support the conclusion that dietary fibre and calcium do not reduce the risk of colorectal cancer.
Colorectal cancer may be signalled by constipation or diarrhoea, cramping, abdominal pain, and rectal bleeding. Bleeding from the rectum may be either obvious or occult (hidden in faeces). Since most colon cancers arise from benign mucosal growths called polyps, cancer prevention is focused on identifying these polyps. The colonoscopy is both diagnostic and therapeutic. Colonoscopy not only allows identification of precancerous polyps, the procedure also enables them to be removed before they become malignant. Screening for faecal occult blood tests and colonoscopy is recommended for those over 50 years of age.
Rectum
Food residue leaving the sigmoid colon enters the rectum in the pelvis, near the third sacral vertebra. The final 20.3 cm of the gastrointestinal tract, the rectum extends anterior to the sacrum and coccyx. Even though rectum is Latin for “straight,” this structure follows the curved contour of the sacrum and has three lateral bends that create a trio of internal transverse folds called the rectal valves. These valves help separate the faeces from gas to prevent the simultaneous passage of faeces and gas.
Anal Canal
Finally, food residue reaches the last part of the large intestine, the anal canal, which is located in the perineum, completely outside of the abdominopelvic cavity. This 3.8–5 cm long structure opens to the exterior of the body at the anus. The anal canal includes two sphincters. The internal anal sphincter is made of smooth muscle, and its contractions are involuntary. The external anal sphincter is made of skeletal muscle, which is under voluntary control. Except when defaecating, both usually remain closed.
Histology
There are several notable differences between the walls of the large and small intestines (Figure 12.5.5, for example, few enzyme-secreting cells are found in the wall of the large intestine, and there are no circular folds or villi. Other than in the anal canal, the mucosa of the colon is simple columnar epithelium made mostly of enterocytes (absorptive cells) and goblet cells. In addition, the wall of the large intestine has far more intestinal glands, which contain a vast population of enterocytes and goblet cells. These goblet cells secrete mucus that eases the movement of faeces and protects the intestine from the effects of the acids and gases produced by enteric microorganisms. The enterocytes absorb water and salts as well as vitamins produced by your intestinal microbiome.
Three features are unique to the large intestine: teniae coli, haustra, and epiploic appendages (Figure 12.5. 6). The teniae coli are three bands of smooth muscle that make up the longitudinal muscle layer of the muscularis of the large intestine, except at its terminal end. Tonic contractions of the teniae coli bunch up the colon into a succession of pouches called haustra (singular = hostrum), which are responsible for the wrinkled appearance of the colon. Attached to the teniae coli are small, fat-filled sacs of visceral peritoneum called epiploic appendages. The purpose of these is unknown. Although the rectum and anal canal have neither teniae coli nor haustra, they do have well-developed layers of muscularis that create the strong contractions needed for defaecation.
Figure 12.5. 6. Teniae coli, haustra, and epiploic appendages.
The stratified squamous epithelial mucosa of the anal canal connects to the skin on the outside of the anus. This mucosa varies from that of the rest of the colon to accommodate the high level of abrasion as faeces pass through. The anal canal’s mucous membrane is organised into longitudinal folds, each called an anal column, which house a grid of arteries and veins. Two superficial venous plexuses are found in the anal canal: one within the anal columns and one at the anus.
Depressions between the anal columns, each called an anal sinus, secrete mucus that facilitates defaecation. The pectinate line (or dentate line) is a horizontal, jagged band that runs circumferentially just below the level of the anal sinuses and represents the junction between the hindgut and external skin. The mucosa above this line is fairly insensitive, whereas the area below is very sensitive. The resulting difference in pain threshold is due to the fact that the upper region is innervated by visceral sensory fibres, and the lower region is innervated by somatic sensory fibres.
Microbiome
Most microorganisms that enter the gastrointestinal tract are killed by lysozyme, defensins, HCl, or protein-digesting enzymes. However, trillions of microorganisms live within the large intestine and are referred to as the microbiome. Most of the more than 700 species of these microorganisms are non-pathogenic commensal organisms that cause no harm if they stay in the gut lumen. In fact, many facilitate chemical digestion and absorption, and some synthesis e certain vitamins, mainly biotin, pantothenic acid and vitamin K. Some are linked to increased immune response. A refined system prevents these bacteria from crossing the mucosal barrier. First, peptidoglycan, a component of bacterial cell walls, activates the release of chemicals by the mucosa’s epithelial cells, which draft immune cells, especially dendritic cells, into the mucosa. Dendritic cells open the tight junctions between epithelial cells and extend probes into the lumen to evaluate the microbial antigens. The dendritic cells with antigens then travel to neighbouring lymphoid follicles in the mucosa where T cells inspect for antigens. This process triggers an IgA-mediated response, if warranted, in the lumen that blocks the commensal organisms from infiltrating the mucosa and setting off a far greater, widespread systematic reaction.
Digestive Functions of the Large Intestine
The residue of chyme that enters the large intestine contains few nutrients except water, which is reabsorbed as the residue lingers in the large intestine, typically for 12 to 24 hours. Thus, it may not surprise you that the large intestine can be completely removed without significantly affecting digestive functioning. In severe cases of inflammatory bowel disease, the large intestine can be removed by a procedure known as a colectomy. Often, a new faecal pouch can be crafted from the small intestine and sutured to the anus, but if not, an ileostomy can be created by bringing the distal ileum through the abdominal wall, allowing the watery chyme to be collected in a bag-like adhesive appliance.
Mechanical Digestion
In the large intestine, mechanical digestion begins when chyme moves from the ileum into the caecum, an activity regulated by the ileocaecal sphincter. Right after you eat, peristalsis in the ileum forces chyme into the cecum. When the cecum is distended with chyme, contractions of the ileocaecal sphincter strengthen. Once chyme enters the caecum, colon movements begin.
Mechanical digestion in the large intestine includes a combination of three types of movements. The presence of food residues in the colon stimulates a slow-moving haustral contraction. This type of movement involves sluggish segmentation, primarily in the transverse and descending colons. When a haustrum is distended with chyme, its muscle contracts, pushing the residue into the next haustrum. These contractions occur about every 30 minutes, and each last about one minute. These movements also mix the food residue, which helps the large intestine absorb water. The second type of movement is peristalsis, which, in the large intestine, is slower than in the more proximal portions of the gastrointestinal tract. The third type is a mass movement. These strong waves start midway through the transverse colon and quickly force the contents toward the rectum. Mass movements usually occur three or four times per day, either while you eat or immediately afterward. Distension in the stomach and the breakdown products of digestion in the small intestine provoke the gastrocolic reflex, which increases motility, including mass movements, in the colon. Fibre in the diet both softens the stool and increases the power of colonic contractions, optimising the activities of the colon.
Chemical Digestion
Although the glands of the large intestine secrete mucus, they do not secrete digestive enzymes. Therefore, chemical digestion in the large intestine occurs exclusively because of microbiota of the colon. Through the process of saccharolytic fermentation, microorganisms break down some of the remaining carbohydrates. This results in the discharge of hydrogen, carbon dioxide, and methane gases that create flatus (gas) in the colon; flatulence is excessive flatus. Each day, up to 1500 mL of flatus is produced in the colon. More is produced when you eat foods such as beans, which are rich in otherwise indigestible sugars and complex carbohydrates like soluble dietary fibre.
Absorption, Faeces Formation and Defaecation
The small intestine absorbs about 90 percent of the water you ingest (either as liquid or within solid food). The large intestine absorbs most of the remaining water, a process that converts the liquid chyme residue into semisolid faeces (“stool”). Faeces is composed of undigested food residues, unabsorbed digested substances, millions of bacteria and other microorganisms, old epithelial cells from the GI mucosa, inorganic salts, and enough water to let it pass smoothly out of the body. Of every 500 mL of food residue that enters the caecum each day, about 150 mL become faeces.
Faeces are eliminated through contractions of the rectal muscles. You help this process by a voluntary procedure called Valsalva’s manoeuvre, in which you increase intra-abdominal pressure by contracting your diaphragm and abdominal wall muscles and closing your glottis.
The process of defaecation begins when mass movements force faeces from the colon into the rectum, stretching the rectal wall and provoking the defaecation reflex, which eliminates faeces from the rectum. This parasympathetic reflex is mediated by the spinal cord. It contracts the sigmoid colon and rectum, relaxes the internal anal sphincter, and initially contracts the external anal sphincter. The presence of faeces in the anal canal sends a signal to the brain, which gives you the choice of voluntarily opening the external anal sphincter (defecating) or keeping it temporarily closed. If you decide to delay defecation, it takes a few seconds for the reflex contractions to stop and the rectal walls to relax. The next mass movement will trigger additional defecation reflexes until you defecate.
If defaecation is delayed for an extended time, additional water is absorbed, making the faeces firmer and potentially leading to constipation. On the other hand, if the waste matter moves too quickly through the intestines, not enough water is absorbed, and diarrhoea can result. This can be caused by the ingestion of foodborne pathogens. In general, diet, health, and stress determine the frequency of bowel movements. The number of bowel movements varies greatly between individuals, ranging from two or three per day to three or four per week.
Section Review
The three main regions of the small intestine are the duodenum, the jejunum, and the ileum. The small intestine is where digestion is completed and virtually all absorption occurs. These two activities are facilitated by structural adaptations that increase the mucosal surface area by 600-fold, including circular folds, villi, and microvilli. There are around 200 million microvilli per square millimetre of small intestine, which contain brush border enzymes that complete the digestion of carbohydrates and proteins. Combined with pancreatic juice, intestinal juice provides the liquid medium needed to further digest and absorb substances from chyme. The small intestine is also the site of unique mechanical digestive movements. Segmentation moves the chyme back and forth, increasing mixing and opportunities for absorption. Migrating motility complexes propel the residual chyme toward the large intestine.
The main regions of the large intestine are the caecum, the colon, and the rectum. The large intestine absorbs water and forms faeces and is responsible for defaecation. The microbiome aids in the breakdown of additional carbohydrate residue and synthesise certain vitamins. The mucosa of the large intestinal wall is generously endowed with goblet cells, which secrete mucus that eases the passage of faeces. The entry of faeces into the rectum activates the defaecation reflex.
12.6 Accessory Organs in Digestion: the Liver, Pancreas and Gallbladder
Learning Objectives
By the end of this section, you will be able to:
State the main digestive roles of the liver, pancreas and gallbladder
Identify three main features of liver histology that are critical to its function
Discuss the composition and function of bile
Identify the major types of enzymes and buffers present in pancreatic secretion
Chemical digestion in the small intestine relies on the activities of three accessory digestive organs: the liver, pancreas, and gallbladder (Figure 12.6.1). The digestive role of the liver is to produce bile and export it to the duodenum. The gallbladder primarily stores, concentrates and releases bile. The pancreas produces pancreatic secretion, which contains digestive enzymes and bicarbonate ions and delivers it to the duodenum.
Figure 12.6.1. Accessory organs. The liver, pancreas, and gallbladder are considered accessory digestive organs, but their roles in the digestive system are vital.
The Liver
The liver is the largest gland in the body, weighing about 1.4 kilograms (three pounds) in an adult. It is also one of the most important organs. In addition to being an accessory digestive organ, it plays several roles in metabolism and regulation. The liver lies inferior to the diaphragm in the right upper quadrant of the abdominal cavity and receives protection from the surrounding ribs.
The liver is divided into two primary lobes: a large right lobe and a much smaller left lobe. In the right lobe, some anatomists also identify an inferior quadrate lobe and a posterior caudate lobe, which are defined by internal features. The liver is connected to the abdominal wall and diaphragm by five peritoneal folds referred to as ligaments. These are the falciform ligament, the coronary ligament, two lateral ligaments, and the ligamentum teres hepatis. The falciform ligament and ligamentum teres hepatis are remnants of the umbilical vein and separate the right and left lobes anteriorly. The lesser omentum tethers the liver to the lesser curvature of the stomach.
The porta hepatis (“gate to the liver”) is where the hepatic artery and hepatic portal vein enter the liver. These two vessels, along with the common hepatic duct, run behind the lateral border of the lesser omentum on the way to their destinations. As shown in Figure 12.6.2, the hepatic artery delivers oxygenated blood from the heart to the liver. The hepatic portal vein delivers partially deoxygenated blood containing nutrients absorbed from the small intestine and supplies more oxygen to the liver than do the much smaller hepatic arteries. In addition to nutrients, drugs and toxins are also absorbed. After processing the bloodborne nutrients and toxins, the liver releases nutrients needed by other cells back into the blood, which drains into the central vein and then through the hepatic vein to the inferior vena cava. With this hepatic portal circulation, all blood from the gastrointestinal tract passes through the liver. This explains why the liver is the most common site for the metastasis of cancers that originate in the gastrointestinal tract.
Figure 12.6.2. Microscopic anatomy of the liver. The liver receives oxygenated blood from the hepatic artery and nutrient-rich deoxygenated blood from the hepatic portal vein.
Histology
The liver has three main components: hepatocytes, bile canaliculi, and hepatic sinusoids. A hepatocyte is the liver’s main cell type, accounting for around 80 percent of the liver’s volume. These cells play a role in a wide variety of secretory, metabolic, and endocrine functions. Plates of hepatocytes called hepatic laminae radiate outward from the portal vein in each hepatic lobule.
Between adjacent hepatocytes, grooves in the cell membranes provide room for each bile canaliculus (plural = canaliculi). These small ducts accumulate the bile produced by hepatocytes. From here, bile flows first into bile ductules and then into bile ducts. The bile ducts unite to form the larger right and left hepatic ducts, which themselves merge and exit the liver as the common hepatic duct. This duct then joins with the cystic duct from the gallbladder, forming the common bile duct through which bile flows into the small intestine.
A hepatic sinusoid is an open, porous blood space formed by fenestrated capillaries from nutrient-rich hepatic portal veins and oxygen-rich hepatic arteries. Hepatocytes are tightly packed around the fenestrated endothelium of these spaces, giving them easy access to the blood. From their central position, hepatocytes process the nutrients, toxins, and waste materials carried by the blood. Materials such as bilirubin are processed and excreted into the bile canaliculi. Other materials including proteins, lipids, and carbohydrates are processed and secreted into the sinusoids or just stored in the cells until called upon. The hepatic sinusoids combine and send blood to a central vein. Blood then flows through a hepatic vein into the inferior vena cava. This means that blood and bile flow in opposite directions. The hepatic sinusoids also contain star-shaped reticuloendothelial cells (Kupffer cells), phagocytes that remove dead red and white blood cells, bacteria, and other foreign material that enter the sinusoids. The portal triad is a distinctive arrangement around the perimeter of hepatic lobules, consisting of three basic structures: a bile duct, a hepatic artery branch, and a hepatic portal vein branch.
Bile
Recall that lipids are hydrophobic, that is, they do not dissolve in water. Thus, before they can be digested in the watery environment of the small intestine, large lipid globules must be broken down into smaller lipid globules, a process called emulsification. Bile is a mixture secreted by the liver to accomplish the emulsification of lipids in the small intestine.
Hepatocytes secrete about one litre of bile each day. A yellow-brown or yellow-green alkaline solution (pH 7.6 to 8.6), bile is a mixture of water, bile salts, bile pigments, phospholipids (such as lecithin), electrolytes, cholesterol, and triglycerides. The components most critical to emulsification are bile salts and phospholipids, which have a nonpolar (hydrophobic) region as well as a polar (hydrophilic) region. The hydrophobic region interacts with the large lipid molecules, whereas the hydrophilic region interacts with the watery chyme in the intestine. This results in the large lipid globules being pulled apart into many tiny lipid fragments of about 1 µm in diameter. This change dramatically increases the surface area available for lipid-digesting enzyme activity. This is the same way dish soap works on fats mixed with water.
Bile salts act as emulsifying agents, so they are also important for the absorption of digested lipids. While most constituents of bile are eliminated in faeces, bile salts are reclaimed by the enterohepatic circulation. Once bile salts reach the ileum, they are absorbed and returned to the liver in the hepatic portal blood. The hepatocytes then excrete the bile salts into newly formed bile. Thus, this precious resource is recycled.
Bilirubin, the main bile pigment, is a waste product produced when the spleen removes old or damaged red blood cells from the circulation. These breakdown products, including proteins, iron, and toxic bilirubin, are transported to the liver via the splenic vein of the hepatic portal system. In the liver, proteins and iron are recycled, whereas bilirubin is excreted in the bile. It accounts for the green colour of bile. Bilirubin is eventually transformed by intestinal bacteria into stercobilin, a brown pigment that gives your stool its characteristic colour! In some disease states, bile does not enter the intestine, resulting in white (‘acholic’) stool with a high fat content, since virtually no fats are broken down or absorbed.
Hepatocytes work non-stop, but bile production increases when fatty chyme enters the duodenum and stimulates the secretion of the gut hormone secretin. Between meals, bile is produced but conserved. The valve-like hepatopancreatic ampulla closes, allowing bile to divert to the gallbladder, where it is concentrated and stored until the next meal.
The Pancreas
The soft, oblong, glandular pancreas lies transversely in the retroperitoneum behind the stomach. Its head is nestled into the “c-shaped” curvature of the duodenum with the body extending to the left about 15.2 cm and ending as a tapering tail in the hilum of the spleen. It is a curious mix of exocrine (secreting digestive enzymes) and endocrine (releasing hormones into the blood) functions (Figure 12.6.3).
Figure 12.6.3. Exocrine and endocrine pancreas. The pancreas has a head, a body, and a tail. It delivers pancreatic secretion to the duodenum through the pancreatic duct.
The exocrine part of the pancreas arises as little grape-like cell clusters, each called an acinus (plural = acini), located at the terminal ends of pancreatic ducts. These acinar cells secrete enzyme-rich pancreatic secretion into tiny merging ducts that form two dominant ducts. The larger duct fuses with the common bile duct (carrying bile from the liver and gallbladder) just before entering the duodenum via a common opening (the hepatopancreatic ampulla). The smooth muscle sphincter of the hepatopancreatic ampulla controls the release of pancreatic secretion and bile into the small intestine. The second and smaller pancreatic duct, the accessory duct (duct of Santorini), runs from the pancreas directly into the duodenum, approximately 2.5 cm above the hepatopancreatic ampulla. When present, it is a persistent remnant of pancreatic development.
Scattered through the sea of exocrine acini are small islands of endocrine cells, the islets of Langerhans. These vital cells produce the hormones pancreatic polypeptide, insulin, glucagon, and somatostatin among others.
Pancreatic Secretion
The pancreas produces over a litre of pancreatic secretion each day. Unlike bile, it is clear and composed mostly of water along with some salts, sodium bicarbonate, and several digestive enzymes. Sodium bicarbonate is responsible for the slight alkalinity of pancreatic secretion (pH 7.1 to 8.2), which serves to buffer the acidic gastric secretion in chyme, inactivate pepsin from the stomach, and create an optimal environment for the activity of pH-sensitive digestive enzymes in the small intestine. Pancreatic enzymes are active in the digestion of sugars, proteins, and fats.
The pancreas produces protein-digesting enzymes in their inactive forms. These enzymes are activated in the duodenum. If produced in an active form, they would digest the pancreas (which is exactly what occurs in the disease, pancreatitis). The intestinal brush border enzyme enteropeptidase stimulates the activation of trypsin from trypsinogen of the pancreas, which in turn changes the pancreatic enzymes procarboxypeptidase and chymotrypsinogen into their active forms, carboxypeptidase and chymotrypsin.
The enzymes that digest starch (amylase), fat (lipase), and nucleic acids (nuclease) are secreted in their active forms, since they do not attack the pancreas as do the protein-digesting enzymes.
Regulation of pancreatic secretion is the job of hormones and the parasympathetic nervous system. The entry of acidic chyme into the duodenum stimulates the release of secretin, which in turn causes the duct cells to release bicarbonate-rich pancreatic secretion. The presence of proteins and fats in the duodenum stimulates the secretion of CCK, which then stimulates the acini to secrete enzyme-rich pancreatic secretion and enhances the activity of secretin. Parasympathetic regulation occurs during the cephalic and gastric phases of gastric secretion, when vagal stimulation prompts the secretion of pancreatic secretion.
Usually, the pancreas secretes just enough bicarbonate to counterbalance the amount of HCl produced in the stomach. Hydrogen ions enter the blood when bicarbonate is secreted by the pancreas. Thus, the acidic blood draining from the pancreas neutralises the alkaline blood draining from the stomach, maintaining the pH of the venous blood that flows to the liver.
The Gallbladder
The gallbladder is 8–10 cm long and is nested in a shallow area on the posterior aspect of the right lobe of the liver. This muscular sac stores, concentrates, and, when stimulated, propels the bile into the duodenum via the common bile duct. It is divided into three regions. The fundus is the widest portion and tapers medially into the body, which in turn narrows to become the neck. The neck angles slightly superiorly as it approaches the hepatic duct. The cystic duct is 1–2 cm long and turns inferiorly as it bridges the neck and hepatic duct.
The simple columnar epithelium of the gallbladder mucosa is organised in rugae, like those of the stomach. There is no submucosa in the gallbladder wall. The wall’s middle, muscular coat is made of smooth muscle fibres. When these fibres contract, the gallbladder’s contents are ejected through the cystic duct and into the bile duct (Figure 12.6.4). Visceral peritoneum reflected from the liver capsule holds the gallbladder against the liver and forms the outer coat of the gallbladder. The gallbladder’s mucosa absorbs water and ions from bile, concentrating it by up to 10-fold.
Figure 12.6.4. Gallbladder. The gallbladder stores and concentrates bile and releases it into the two-way cystic duct when it is needed by the small intestine.
Section Review
Chemical digestion in the small intestine cannot occur without the help of the liver and pancreas. The liver produces bile and delivers it to the common hepatic duct. Bile contains bile salts and phospholipids, which emulsify large lipid globules into tiny lipid droplets, a necessary step in lipid digestion and absorption. The gallbladder stores and concentrates bile, releasing it when it is needed by the small intestine.
The pancreas produces the enzyme- and bicarbonate-rich pancreatic secretion and delivers it to the small intestine through ducts. Pancreatic secretion buffers the acidic gastric secretion in chyme, inactivates pepsin from the stomach, and enables the optimal functioning of digestive enzymes in the small intestine.
Identify the locations and primary secretions involved in the chemical digestion of carbohydrates, proteins, lipids and nucleic acids
Compare and contrast absorption of the hydrophilic and hydrophobic nutrients
As you have learned, the process of mechanical digestion is relatively simple. It involves the physical breakdown of food but does not alter its chemical makeup. Chemical digestion, on the other hand, is a complex process that reduces food into its chemical building blocks, which are then absorbed to nourish the cells of the body (Figure 12.7.1). In this section, you will look more closely at the processes of chemical digestion and absorption.
Figure 12.7.1. Digestion and absorption. Digestion begins in the mouth and continues as food travels through the small intestine. Most absorption occurs in the small intestine.
Chemical Digestion
Large food molecules (for example, proteins, lipids, nucleic acids, and starches) must be broken down into subunits that are small enough to be absorbed by the lining of the gastrointestinal tract. This is accomplished by enzymes through hydrolysis. The many enzymes involved in chemical digestion are summarised in Table 12.7.1.
Table 12.7.1. The digestive enzymes
Enzyme category
Enzyme name
Source
Substrate
Product
Salivary enzymes
Lingual lipase
Lingual glands
Triglycerides
Free fatty acids, and mono-and diglycerides
Salivary amylase
Salivary glands
Polysaccharides
Disaccharides and trisaccharides
Gastric enzymes
Gastric lipase
Chief cells
Triglycerides
Fatty acids and monoacylglycerides
Pepsin*
Chief cells
Proteins
Peptides
Brush border enzymes
α-Dextrinase
Small intestine
α-Dextrins
Glucose
Enteropeptidase
Small intestine
Trypsinogen
Trypsin
Lactase
Small intestine
Lactose
Glucose and galactose
Maltase
Small intestine
Maltose
Glucose
Nucleosidases and phosphatases
Small intestine
Nucleotides
Phosphates, nitrogenous bases, and pentoses
Peptidases
Small intestine
Aminopeptidase: amino acids at the amino end of peptides
Triglycerides that have been emulsified by bile salts
Fatty acids and monoacylglycerides
Trypsin*
Pancreatic acinar cells
Proteins
Peptides
*These enzymes have been activated by other substances.
Carbohydrate Digestion
The average American diet is about 50 percent carbohydrates, which may be classified according to the number of monomers they contain of simple sugars (monosaccharides and disaccharides) and/or complex sugars (polysaccharides). Glucose, galactose, and fructose are the three monosaccharides that are commonly consumed and are readily absorbed. Your digestive system is also able to break down the disaccharide sucrose (regular table sugar: glucose + fructose), lactose (milk sugar: glucose + galactose), and maltose (grain sugar: glucose + glucose), and the polysaccharides glycogen and starch (chains of monosaccharides). Your bodies do not produce enzymes that can break down most fibrous polysaccharides, such as cellulose. While indigestible polysaccharides do not provide any nutritional value, they do provide dietary fibre, which helps propel food through the gastrointestinal tract.
The chemical digestion of starches begins in the mouth and has been reviewed above.
In the small intestine, pancreatic amylase does the ‘heavy lifting’ for starch and carbohydrate digestion (Figure 12.7.2). After amylases break down starch into smaller fragments, the brush border enzyme α-dextrinase starts working on α-dextrin, breaking off one glucose unit at a time. Three brush border enzymes hydrolyse sucrose, lactose, and maltose into monosaccharides. Sucrase splits sucrose into one molecule of fructose and one molecule of glucose; maltase breaks down maltose and maltotriose into two and three glucose molecules, respectively; and lactase breaks down lactose into one molecule of glucose and one molecule of galactose. Insufficient lactase can lead to lactose intolerance.
Figure 12.7.2. Carbohydrate digestion flow chart. Carbohydrates are broken down into their monomers in a series of steps.
Protein Digestion
Proteins are polymers composed of amino acids linked by peptide bonds to form long chains. Digestion reduces them to their constituent amino acids. You usually consume about 15 to 20 percent of your total calorie intake as protein.
The digestion of protein starts in the stomach, where HCl and pepsin break proteins into smaller polypeptides, which then travel to the small intestine (Figure 12.7.3). Chemical digestion in the small intestine is continued by pancreatic enzymes, including chymotrypsin and trypsin, each of which act on specific bonds in amino acid sequences. At the same time, the cells of the brush border secrete enzymes such as aminopeptidase and dipeptidase, which further break down peptide chains. This results in molecules small enough to enter the bloodstream (Figure 12.7.4).
Figure 12.7.3. Digestion of protein. The digestion of protein begins in the stomach and is completed in the small intestine.
Figure 12.7.4. Digestion of protein flow chart. Proteins are successively broken down into their amino acid components.
Lipid Digestion
A healthy diet limits lipid intake to 35 percent of total calorie intake. The most common dietary lipids are triglycerides, which are made up of a glycerol molecule bound to three fatty acid chains. Small amounts of dietary cholesterol and phospholipids are also consumed.
The three lipases responsible for lipid digestion are lingual lipase, gastric lipase, and pancreatic lipase. However, because the pancreas is the only consequential source of lipase, virtually all lipid digestion occurs in the small intestine. Pancreatic lipase breaks down each triglyceride into two free fatty acids and a monoglyceride. The fatty acids include both short-chain (less than 10 to 12 carbons) and long-chain fatty acids.
Nucleic Acid Digestion
The nucleic acids DNA and RNA are found in most of the foods you eat. Two types of pancreatic nuclease are responsible for their digestion: deoxyribonuclease, which digests DNA, and ribonuclease, which digests RNA. The nucleotides produced by this digestion are further broken down by two intestinal brush border enzymes (nucleosidase and phosphatase) into pentoses, phosphates, and nitrogenous bases, which can be absorbed through the gastrointestinal tract wall. The large food molecules that must be broken down into subunits are summarised Table 12.7.2.
Table 12.7.2. Absorbable food substances
Source
Substance
Carbohydrates
Monosaccharides: glucose, galactose and fructose
Proteins
Single amino acids, dipeptides and tripeptides
Triglycerides
Monoacylglycerides, glycerol and free fatty acids
Nucleic acids
Pentose sugars, phosphates and nitrogenous bases
Absorption
The mechanical and digestive processes have one goal: to convert food into molecules small enough to be absorbed by the epithelial cells of the intestinal villi. The absorptive capacity of the gastrointestinal tract is almost endless. Each day, the gastrointestinal tract processes up to 10 litres of food, liquids, and GI secretions, yet less than one litre enters the large intestine. Almost all ingested food, 80 percent of electrolytes, and 90 percent of water are absorbed in the small intestine. Although the entire small intestine is involved in the absorption of water and lipids, most absorption of carbohydrates and proteins occurs in the jejunum. Notably, bile salts and vitamin B12 are absorbed in the terminal ileum. By the time chyme passes from the ileum into the large intestine, it is essentially indigestible food residue (mainly plant fibres like cellulose), some water and millions of bacteria and other microorganisms (Figure 12.7.5).
Figure 12.7.5. Digestive secretions and absorption of water. Absorption is a complex process, in which nutrients from digested food are harvested.
Absorption can occur through five mechanisms: (1) active transport, (2) passive diffusion, (3) facilitated diffusion, (4) co-transport (or secondary active transport), and (5) endocytosis. Active transport refers to the movement of a substance across a cell membrane going from an area of lower concentration to an area of higher concentration (up the concentration gradient). In this type of transport, proteins within the cell membrane act as “pumps,” using cellular energy (ATP) to move the substance. Passive diffusion refers to the movement of substances from an area of higher concentration to an area of lower concentration, while facilitated diffusion refers to the movement of substances from an area of higher to an area of lower concentration using a carrier protein in the cell membrane. Co-transport uses the movement of one molecule through the membrane from higher to lower concentration to power the movement of another from lower to higher. Finally, endocytosis is a transportation process in which the cell membrane engulfs material. It requires energy, generally in the form of ATP.
Because the cell’s plasma membrane is made up of hydrophobic phospholipids, water-soluble nutrients must use transport molecules embedded in the membrane to enter cells. Moreover, substances cannot pass between the epithelial cells of the intestinal mucosa because these cells are bound together by tight junctions. Thus, substances can only enter blood capillaries by passing through the apical surfaces of epithelial cells and into the interstitial fluid. Water-soluble nutrients enter the capillary blood in the villi and travel to the liver via the hepatic portal vein.
In contrast to the water-soluble nutrients, lipid-soluble nutrients can diffuse through the plasma membrane. Once inside the cell, they are packaged for transport via the base of the cell and then enter the lacteals of the villi to be transported by lymphatic vessels to the systemic circulation via the thoracic duct. The absorption of most nutrients through the mucosa of the intestinal villi requires active transport fuelled by ATP. The routes of absorption for each food category are summarised in Table 12.7.3.
Table 12.7.3. Absorption in the gastrointestinal tract
Food
Breakdown products
Absorption mechanisms
Entry to bloodstream
Destination
Carbohydrates
Glucose
Co-transport with sodium ions
Capillary blood in villi
Liver via hepatic portal vein
Galactose
Co-transports with sodium ions
Capillary blood via villi
Liver via hepatic portal vein
Fructose
Facilitated diffusion
Capillary blood in villi
Liver via hepatic portal vein
Protein
Amino acids
Co-transported with sodium ions
Capillary blood in villi
Liver in hepatic portal vein
Lipids
Long-chain fatty acids
Diffusion into intestinal cells where they are combined with proteins to create chylomicrons
Lacteals of villi
Systemic circulation via lymph entering thoracic duct
Monoacylglycerides
Diffusion into intestinal cells, where they are combined with proteins to create chylomicrons
Lacteals of villi
Systemic circulation via lymph entering the thoracic duct
Short-chain fatty acids
Simple diffusion
Capillary blood in villi
Liver via hepatic portal vein
Glycerol
Simple diffusion
Capillary blood in villi
Liver via hepatic portal vein
Nucleic acid digestion products
Active transport via membrane carriers
Capillary blood in villi
Liver via hepatic portal vein
Carbohydrate Absorption
All carbohydrates are absorbed in the form of monosaccharides. The small intestine is highly efficient at this, absorbing monosaccharides at an estimated rate of 120 grams per hour. All normally digested dietary carbohydrates are absorbed; indigestible fibres are eliminated in the faeces. The monosaccharides glucose and galactose are transported into the epithelial cells by common protein carriers via secondary active transport (that is, co-transport with sodium ions). The monosaccharides leave these cells via facilitated diffusion and enter the capillaries through intercellular clefts. The monosaccharide fructose (which is in fruit) is absorbed and transported by facilitated diffusion alone. The monosaccharides combine with the transport proteins immediately after the disaccharides are broken down.
Protein Absorption
Active transport mechanisms, primarily in the duodenum and jejunum, absorb most proteins as their breakdown products, amino acids. Almost all (95 to 98 percent) protein is digested and absorbed in the small intestine. The type of carrier that transports an amino acid varies. Most carriers are linked to the active transport of sodium. Short chains of two amino acids (dipeptides) or three amino acids (tripeptides) are also transported actively. However, after they enter the absorptive epithelial cells, they are broken down into their amino acids before leaving the cell and entering the capillary blood via diffusion.
Lipid Absorption
About 95 percent of lipids are absorbed in the small intestine. Bile salts not only speed up lipid digestion, they are also essential for the absorption of the end products of lipid digestion. Short-chain fatty acids are water soluble and can enter the absorptive cells (enterocytes) directly. Despite being hydrophobic, the small size of short-chain fatty acids enables them to be absorbed by enterocytes via simple diffusion, and then take the same path as monosaccharides and amino acids into the blood capillary of a villus.
The large and hydrophobic long-chain fatty acids and monoacylglycerides are not so easily suspended in the watery intestinal chyme. However, bile salts and lecithin resolve this issue by enclosing them in a micelle, which is a tiny sphere with polar (hydrophilic) ends facing the watery environment and hydrophobic tails turned to the interior, creating a receptive environment for the long-chain fatty acids. The core also includes cholesterol and fat-soluble vitamins. Without micelles, lipids would sit on the surface of chyme and never come in contact with the absorptive surfaces of the epithelial cells. Micelles can easily squeeze between microvilli and get very near the luminal cell surface. At this point, lipid substances exit the micelle and are absorbed via simple diffusion.
The free fatty acids and monoacylglycerides that enter the epithelial cells are reincorporated into triglycerides. The triglycerides are mixed with phospholipids and cholesterol and surrounded with a protein coat. This new complex, called a chylomicron, is a water-soluble lipoprotein. After being processed by the Golgi apparatus, chylomicrons are released from the cell (Figure 12.7.6). Too big to pass through the basement membranes of blood capillaries, chylomicrons instead enter the large pores of lacteals. The lacteals come together to form the lymphatic vessels. The chylomicrons are transported in the lymphatic vessels and empty through the thoracic duct into the subclavian vein of the circulatory system. Once in the bloodstream, the enzyme lipoprotein lipase breaks down the triglycerides of the chylomicrons into free fatty acids and glycerol. These breakdown products then pass through capillary walls to be used for energy by cells or stored in adipose tissue as fat. Liver cells combine the remaining chylomicron remnants with proteins, forming lipoproteins that transport cholesterol in the blood.
Figure 12.7.6. Lipid absorption. Unlike amino acids and simple sugars, lipids are transformed as they are absorbed through epithelial cells.
Nucleic Acid Absorption
The products of nucleic acid digestion—pentose sugars, nitrogenous bases, and phosphate ions—are transported by carriers across the villus epithelium via active transport. These products then enter the bloodstream.
Mineral Absorption
The electrolytes absorbed by the small intestine are from both GI secretions and ingested foods. Since electrolytes dissociate into ions in water, most are absorbed via active transport throughout the entire small intestine. During absorption, co-transport mechanisms result in the accumulation of sodium ions inside the cells, whereas anti-port mechanisms reduce the potassium ion concentration inside the cells. To restore the sodium-potassium gradient across the cell membrane, a sodium-potassium pump requiring ATP pumps sodium out and potassium in.
In general, all minerals that enter the intestine are absorbed, whether you need them or not. Iron and calcium are exceptions; they are absorbed in the duodenum in amounts that meet the body’s current requirements, as follows:
Iron—The ionic iron needed for the production of haemoglobin is absorbed into mucosal cells via active transport. Once inside mucosal cells, ionic iron binds to the protein ferritin, creating iron-ferritin complexes that store iron until needed. When the body has enough iron, most of the stored iron is lost when worn-out epithelial cells slough off. When the body needs iron because, for example, it is lost during acute or chronic bleeding, there is increased uptake of iron from the intestine and accelerated release of iron into the bloodstream. Since women experience significant iron loss during menstruation, they have around four times as many iron transport proteins in their intestinal epithelial cells as do men.
Calcium—Blood levels of ionic calcium determine the absorption of dietary calcium. When blood levels of ionic calcium drop, parathyroid hormone (PTH) secreted by the parathyroid glands stimulates the release of calcium ions from bone matrices and increases the reabsorption of calcium by the kidneys. PTH also upregulates the activation of vitamin D in the kidney, which then facilitates intestinal calcium ion absorption.
Vitamin Absorption
The small intestine absorbs the vitamins that occur naturally in food and supplements. Fat-soluble vitamins (A, D, E, and K) are absorbed along with dietary lipids in micelles via simple diffusion. This is why you are advised to eat some fatty foods when you take fat-soluble vitamin supplements. Most water-soluble vitamins (including most B vitamins and vitamin C) also are absorbed by simple diffusion. An exception is vitamin B12, which is a very large molecule. Intrinsic factor secreted in the stomach binds to vitamin B12, preventing its digestion and creating a complex that binds to mucosal receptors in the terminal ileum, where it is taken up by endocytosis.
Water Absorption
Each day, about nine litres of fluid enter the small intestine. About 2.3 litres are ingested in foods and beverages, and the rest is from GI secretions. About 90 percent of this water is absorbed in the small intestine. Water absorption is driven by the concentration gradient of the water: The concentration of water is higher in chyme than it is in epithelial cells. Thus, water moves down its concentration gradient from the chyme into cells. As noted earlier, much of the remaining water is then absorbed in the colon.
Section Review
The small intestine is the site of most chemical digestion and almost all absorption. Chemical digestion breaks large food molecules down into their chemical building blocks, which can then be absorbed through the intestinal wall and into the general circulation. Intestinal brush border enzymes and pancreatic enzymes are responsible for most of the chemical digestion. The breakdown of fat also requires bile.
Most nutrients are absorbed by transport mechanisms at the apical surface of enterocytes. Exceptions include lipids, fat-soluble vitamins, and most water-soluble vitamins. With the help of bile salts and lecithin, the dietary fats are emulsified to form micelles, which can carry the fat particles to the surface of the enterocytes. There, the micelles release their fats to diffuse across the cell membrane. The fats are then reassembled into triglycerides and mixed with other lipids and proteins into chylomicrons that can pass into lacteals. Other absorbed monomers travel from blood capillaries in the villus to the hepatic portal vein and then to the liver.
13.1 Basic Structure and Function of the Nervous System
Learning Objectives
By the end of this section, you will be able to:
Identify the anatomical and functional divisions of the nervous system
Relate the functional and structural differences between grey matter and white matter structures of the nervous system to the structure of neurons
List the basic functions of the nervous system
The picture you have in your mind of the nervous system probably includes the brain, the nervous tissue contained within the cranium, and the spinal cord, the extension of nervous tissue within the vertebral column. That suggests it is made of two organs—and you may not even think of the spinal cord as an organ—but the nervous system is a very complex structure. Within the brain, many different and separate regions are responsible for many different and separate functions. It is as if the nervous system is composed of many organs that all look similar and can only be differentiated using tools such as the microscope or electrophysiology. In comparison, it is easy to see that the stomach is different than the oesophagus or the liver, so you can imagine the digestive system as a collection of specific organs.
The Central and Peripheral Nervous System
The nervous system can be divided into two major regions: the central and peripheral nervous systems. The central nervous system (CNS) is the brain and spinal cord, and the peripheral nervous system (PNS) is everything else (Figure 13.1.1). The brain is contained within the cranial cavity of the skull, and the spinal cord is contained within the vertebral cavity of the vertebral column. It is a bit of an oversimplification to say that the CNS is what is inside these two cavities and the peripheral nervous system is outside of them, but that is one way to start to think about it. There are some elements of the peripheral nervous system that are within the cranial or vertebral cavities. The peripheral nervous system is so named because it is on the periphery—meaning beyond the brain and spinal cord. Depending on different aspects of the nervous system, the dividing line between central and peripheral is not necessarily universal.
Figure 13.1.1 Central and peripheral nervous system. The structures of the PNS are referred to as ganglia and nerves, which can be seen as distinct structures. The equivalent structures in the CNS are not obvious from this overall perspective and are best examined in prepared tissue under the microscope.
Nervous tissue, present in both the CNS and PNS, contains two basic types of cells: neurons and glial cells. A glial cell is one of a variety of cells that provide a framework of tissue that supports the neurons and their activities. The neuron is the more functionally important of the two, in terms of the communicative function of the nervous system. To describe the functional divisions of the nervous system, it is important to understand the structure of a neuron. Neurons are cells and therefore have a soma, or cell body, but they also have extensions of the cell; each extension is generally referred to as a process. There is one important process that every neuron has called an axon, which is the fibre that connects a neuron with its target. Another type of process that branches off from the soma is the dendrite. Dendrites are responsible for receiving most of the input from other neurons. Looking at nervous tissue, there are regions that predominantly contain cell bodies and regions that are largely composed of just axons. These two regions within nervous system structures are often referred to as grey matter (the regions with many cell bodies and dendrites) or white matter (the regions with many axons). Figure 13.1.2 demonstrates the appearance of these regions in the brain and spinal cord. The colours ascribed to these regions are what would be seen in “fresh,” or unstained, nervous tissue. Grey matter is not necessarily grey. It can be pinkish because of blood content, or even slightly tan, depending on how long the tissue has been preserved. But white matter is white because axons are insulated by a lipid-rich substance called myelin. Lipids can appear as white (“fatty”) material, much like the fat on a raw piece of chicken or beef. Actually, grey matter may have that colour ascribed to it because next to the white matter, it is just darker—hence, grey.
The distinction between grey matter and white matter is most often applied to central nervous tissue, which has large regions that can be seen with the unaided eye. When looking at peripheral structures, often a microscope is used, and the tissue is stained with artificial colours. That is not to say that central nervous tissue cannot be stained and viewed under a microscope, but unstained tissue is most likely from the CNS—for example, a frontal section of the brain or cross section of the spinal cord.
Figure 13.1.2. Grey matter and white matter. A brain removed during an autopsy, with a partial section removed, shows white matter surrounded by grey matter. Grey matter makes up the outer cortex of the brain. (credit: modification of work by “Suseno”/Wikimedia Commons).
Regardless of the appearance of stained or unstained tissue, the cell bodies of neurons or axons can be located in discrete anatomical structures that need to be named. Those names are specific to whether the structure is central or peripheral. A localised collection of neuron cell bodies in the CNS is referred to as a nucleus. In the PNS, a cluster of neuron cell bodies is referred to as a ganglion. Figure 13.1.3 indicates how the term nucleus has a few different meanings within anatomy and physiology. It is the centre of an atom, where protons and neutrons are found; it is the centre of a cell, where the DNA is found; and it is a centre of some function in the CNS. There is also a potentially confusing use of the word ganglion (plural = ganglia) that has a historical explanation. In the central nervous system, there is a group of nuclei that are connected and were once called the basal ganglia before “ganglion” became accepted as a description for a peripheral structure. Some sources refer to this group of nuclei as the “basal nuclei” to avoid confusion.
Figure 13.1.3. What is a nucleus? (a) The nucleus of an atom contains its protons and neutrons. (b) The nucleus of a cell is the organelle that contains DNA. (c) A nucleus in the CNS is a localised centre of function with the cell bodies of several neurons, shown here circled in red. (credit c: “Was a bee”/Wikimedia Commons).
Terminology applied to bundles of axons also differs depending on location. A bundle of axons, or fibres, found in the CNS is called a tract whereas the same thing in the PNS would be called a nerve. There is an important point to make about these terms, which is that they can both be used to refer to the same bundle of axons. When those axons are in the PNS, the term is nerve, but if they are CNS, the term is tract. The most obvious example of this is the axons that project from the retina into the brain. Those axons are called the optic nerve as they leave the eye, but when they are inside the cranium, they are referred to as the optic tract. There is a specific place where the name changes, which is the optic chiasm, but they are still the same axons (Figure 13.1.4). A similar situation outside of science can be described for some roads. Imagine a road called “Broad Street” in a town called “Anyville.” The road leaves Anyville and goes to the next town over, called “Hometown.” When the road crosses the line between the two towns and is in Hometown, its name changes to “Main Street.” That is the idea behind the naming of the retinal axons. In the PNS, they are called the optic nerve, and in the CNS, they are the optic tract. Table 13.1.1 helps to clarify which of these terms apply to the central or peripheral nervous systems.
Figure 13.1.4. Optic nerve versus optic tract. This drawing of the connections of the eye to the brain shows the optic nerve extending from the eye to the chiasm, where the structure continues as the optic tract. The same axons extend from the eye to the brain through these two bundles of fibres, but the chiasm represents the border between peripheral and central.
Table 13.1.1. Structures of the CNS and PNS
Structure
CNS
PNS
Group of neuron cell bodies
(i.e., grey matter)
Nucleus
Ganglion
Bundle of Axons
(i.e., white matter)
Tract
Nerve
Functional Divisions of the Nervous System
The nervous system can also be divided based on its functions, but anatomical divisions and functional divisions are different. The CNS and the PNS both contribute to the same functions, but those functions can be attributed to different regions of the brain (such as the cerebral cortex or the hypothalamus) or to different ganglia in the periphery. The problem with trying to fit functional differences into anatomical divisions is that sometimes the same structure can be part of several functions. For example, the optic nerve carries signals from the retina that are either used for the conscious perception of visual stimuli, which takes place in the cerebral cortex, or for the reflexive responses of smooth muscle tissue that are processed through the hypothalamus.
There are two ways to consider how the nervous system is divided functionally. First, the basic functions of the nervous system are sensation, integration, and response. Secondly, control of the body can be somatic or autonomic—divisions that are defined by the structures that are involved in the response. There is also a region of the peripheral nervous system that is called the enteric nervous system that is responsible for a specific set of the functions within the realm of autonomic control related to gastrointestinal functions.
Basic Functions
The nervous system is involved in receiving information about the environment around us (sensation) and generating responses to that information (motor responses). The nervous system can be divided into regions that are responsible for sensation (sensory functions) and for the response (motor functions). But there is a third function that needs to be included. Sensory input needs to be integrated with other sensations, as well as with memories, emotional state, or learning (cognition). Some regions of the nervous system are termed integration or association areas. The process of integration combines sensory perceptions and higher cognitive functions such as memories, learning, and emotion to produce a response.
Sensation. The first major function of the nervous system is sensation—receiving information about the environment to gain input about what is happening outside the body (or, sometimes, within the body). The sensory functions of the nervous system register the presence of a change from homeostasis or a particular event in the environment, known as a stimulus. The senses we think of most are the “big five”: taste, smell, touch, sight, and hearing. The stimuli for taste and smell are both chemical substances (molecules, compounds, ions, etc.), touch is physical or mechanical stimuli that interact with the skin, sight is light stimuli, and hearing is the perception of sound, which is a physical stimulus similar to some aspects of touch. There are more senses than just those, but that list represents the major senses. Those five are all senses that receive stimuli from the outside world, and of which there is conscious perception. Additional sensory stimuli might be from the internal environment (inside the body), such as the stretch of an organ wall or the concentration of certain ions in the blood.
Response. The nervous system produces a response on the basis of the stimuli perceived by sensory structures. An obvious response would be the movement of muscles, such as withdrawing a hand from a hot stove, but there are broader uses of the term. The nervous system can cause the contraction of all three types of muscle tissue, for example, skeletal muscle contracts to move the skeleton, cardiac muscle is influenced as heart rate increases during exercise, and smooth muscle contracts as the digestive system moves food along the digestive tract. Responses also include the neural control of glands in the body as well, such as the production and secretion of sweat by the eccrine and merocrine sweat glands found in the skin to lower body temperature.
Responses can be divided into those that are voluntary or conscious (contraction of skeletal muscle) and those that are involuntary (contraction of smooth muscles, regulation of cardiac muscle, activation of glands). Voluntary responses are governed by the somatic nervous system and involuntary responses are governed by the autonomic nervous system, which are discussed in the next section.
Integration. Stimuli that are received by sensory structures are communicated to the nervous system where that information is processed. This is called integration. Stimuli are compared with, or integrated with, other stimuli, memories of previous stimuli, or the state of a person at a particular time. This leads to the specific response that will be generated. Seeing a baseball pitched to a batter will not automatically cause the batter to swing. The trajectory of the ball and its speed will need to be considered. Maybe the count is three balls and one strike, and the batter wants to let this pitch go by in the hope of getting a walk to first base. Or maybe the batter’s team is so far ahead, it would be fun to just swing away.
Controlling the Body
The nervous system can be divided into two parts mostly on the basis of a functional difference in responses. The somatic nervous system (SNS) is responsible for conscious perception and voluntary motor responses. Voluntary motor response means the contraction of skeletal muscle, but those contractions are not always voluntary in the sense that you must want to perform them. Some somatic motor responses are reflexes, and often happen without a conscious decision to perform them. If your friend jumps out from behind a corner and yells “Boo!” you will be startled and you might scream or leap back. You did not decide to do that, and you may not have wanted to give your friend a reason to laugh at your expense, but it is a reflex involving skeletal muscle contractions. Other motor responses become automatic (in other words, unconscious) as a person learns motor skills (referred to as “habit learning” or “procedural memory”).
The autonomic nervous system (ANS) is responsible for involuntary control of the body, usually for the sake of homeostasis (regulation of the internal environment). Sensory input for autonomic functions can be from sensory structures tuned to external or internal environmental stimuli. The motor output extends to smooth and cardiac muscle as well as glandular tissue. The role of the autonomic system is to regulate the organ systems of the body, which usually means to control homeostasis. Sweat glands, for example, are controlled by the autonomic system. When you are hot, sweating helps cool your body down. That is a homeostatic mechanism. But when you are nervous, you might start sweating also. That is not homeostatic, it is the physiological response to an emotional state.
There is another division of the nervous system that describes functional responses. The enteric nervous system (ENS) is responsible for controlling the smooth muscle and glandular tissue in your digestive system. It is a large part of the PNS and is not dependent on the CNS. It is sometimes valid, however, to consider the enteric system to be a part of the autonomic system because the neural structures that make up the enteric system are a component of the autonomic output that regulates digestion. There are some differences between the two, but for our purposes here there will be a good bit of overlap. See Figure 13.1.5 for examples of where these divisions of the nervous system can be found.
Figure 13.1.5. Somatic, autonomic, and enteric structures of the nervous system. Somatic structures include the spinal nerves, both motor and sensory fibres, as well as the sensory ganglia (posterior root ganglia and cranial nerve ganglia). Autonomic structures are found in the nerves also but include the sympathetic and parasympathetic ganglia. The enteric nervous system includes the nervous tissue within the organs of the digestive tract.
Everyday Connection
How Much of Your Brain Do You Use?
Figure 13.1.6. fMRI. This fMRI shows activation of the visual cortex in response to visual stimuli. (credit: “Superborsuk”/Wikimedia Commons).
Have you ever heard the claim that humans only use 10 percent of their brains? Maybe you have seen an advertisement on a website saying that there is a secret to unlocking the full potential of your mind—as if there were 90 percent of your brain sitting idle, just waiting for you to use it. If you see an ad like that, do not click. It is not true.
An easy way to see how much of the brain a person uses is to take measurements of brain activity while performing a task. An example of this kind of measurement is functional magnetic resonance imaging (fMRI), which generates a map of the most active areas and can be generated and presented in three dimensions (Figure 13.1.6). This procedure is different from the standard MRI technique because it is measuring changes in the tissue in time with an experimental condition or event.
The underlying assumption is that active nervous tissue will have greater blood flow. By having the subject perform a visual task, activity all over the brain can be measured. Consider this experiment: the subject is told to look at a screen with a black dot in the middle (a fixation point). A photograph of a face is projected on the screen away from the centre. The subject must look at the photograph and decipher what it is. The subject has been instructed to push a button if the photograph is of someone they recognise. The photograph might be of a celebrity, so the subject would press the button, or it might be of a random person unknown to the subject, so the subject would not press the button.
In this task, visual sensory areas would be active, integrating areas would be active, motor areas responsible for moving the eyes would be active, and motor areas for pressing the button with a finger would be active. Those areas are distributed all around the brain and the fMRI images would show activity in more than just ten percent of the brain (some evidence suggests that about 80 percent of the brain is using energy—based on blood flow to the tissue—during well-defined tasks like the one suggested above). This task does not even include all the functions the brain performs. There is no language response, the body is mostly lying still in the MRI machine and it does not consider the autonomic functions that would be ongoing in the background.
Section Review
The nervous system can be separated into divisions based on anatomy and physiology. The anatomical divisions are the central and peripheral nervous systems. The CNS is the brain and spinal cord. The PNS is everything else. Functionally, the nervous system can be divided into those regions that are responsible for sensation, those that are responsible for integration, and those that are responsible for generating responses. All these functional areas are found in both the central and peripheral anatomy.
Considering the anatomical regions of the nervous system, there are specific names for the structures within each division. A localised collection of neuron cell bodies is referred to as a nucleus in the CNS and as a ganglion in the PNS. A bundle of axons is referred to as a tract in the CNS and as a nerve in the PNS. Whereas nuclei and ganglia are specifically in the central or peripheral divisions, axons can cross the boundary between the two. A single axon can be part of a nerve and a tract. The name for that specific structure depends on its location.
Nervous tissue can also be described as grey matter and white matter on the basis of its appearance in unstained tissue. These descriptions are more often used in the CNS. Grey matter is where nuclei are found and white matter is where tracts are found. In the PNS, ganglia are basically grey matter and nerves are white matter.
The nervous system can also be divided on the basis of how it controls the body. The somatic nervous system (SNS) is responsible for functions that result in moving skeletal muscles. Any sensory or integrative functions that result in the movement of skeletal muscle would be considered somatic. The autonomic nervous system (ANS) is responsible for functions that affect cardiac or smooth muscle tissue, or that cause glands to produce their secretions. Autonomic functions are distributed between central and peripheral regions of the nervous system. The sensations that lead to autonomic functions can be the same sensations that are part of initiating somatic responses. Somatic and autonomic integrative functions may overlap as well.
A special division of the nervous system is the enteric nervous system, which is responsible for controlling the digestive organs. Parts of the autonomic nervous system overlap with the enteric nervous system. The enteric nervous system is exclusively found in the periphery because it is the nervous tissue in the organs of the digestive system.
Identify the different types of neurons on the basis of polarity
List the glial cells of the CNS and describe their function
List the glial cells of the PNS and describe their function
Nervous tissue is composed of two types of cells, neurons and glial cells. Neurons are the primary type of cell that most anyone associates with the nervous system. They are responsible for the computation and communication that the nervous system provides. They are electrically active and release chemical signals to target cells. Glial cells, or glia, are known to play a supporting role for nervous tissue. Ongoing research pursues an expanded role that glial cells might play in signalling, but neurons are still considered the basis of this function. Neurons are important, but without glial support they would not be able to perform their function.
Neurons
Neurons are the cells considered to be the basis of nervous tissue. They are responsible for the electrical signals that communicate information about sensations, and that produce movements in response to those stimuli, along with inducing thought processes within the brain. An important part of the function of neurons is in their structure, or shape. The three-dimensional shape of these cells makes the immense numbers of connections within the nervous system possible.
Parts of the Neurons
As you learned in the first section, the main part of a neuron is the cell body, which is also known as the soma (soma = “body”). The cell body contains the nucleus and most of the major organelles. But what makes neurons special is that they have many extensions of their cell membranes, which are generally referred to as processes. Neurons are usually described as having one, and only one, axon—a fibre that emerges from the cell body and projects to target cells. That single axon can branch repeatedly to communicate with many target cells. It is the axon that propagates the nerve impulse, which is communicated to one or more cells. The other processes of the neuron are dendrites, which receive information from other neurons at specialised areas of contact called synapses. The dendrites are usually highly branched processes, providing locations for other neurons to communicate with the cell body. Information flows through a neuron from the dendrites, across the cell body, and down the axon. This gives the neuron a polarity—meaning that information flows in this one direction. Figure 13.2.1 shows the relationship of these parts to one another.
Figure 13.2.1. Parts of a neuron. The major parts of the neuron are labelled on a multipolar neuron from the CNS.
Where the axon emerges from the cell body, there is a special region referred to as the axon hillock. This is a tapering of the cell body toward the axon fibre. Within the axon hillock, the cytoplasm changes to a solution of limited components called axoplasm. Because the axon hillock represents the beginning of the axon, it is also referred to as the initial segment.
Many axons are wrapped by an insulating substance called myelin, which is made from glial cells. Myelin acts as insulation much like the plastic or rubber that is used to insulate electrical wires. A key difference between myelin and the insulation on a wire is that there are gaps in the myelin covering of an axon. Each gap is called a node of Ranvier and is important to the way that electrical signals travel down the axon. The length of the axon between each gap, which is wrapped in myelin, is referred to as an axon segment. At the end of the axon is the axon terminal, where there are usually several branches extending toward the target cell, each of which ends in an enlargement called a synaptic end bulb. These bulbs are what make the connection with the target cell at the synapse.
Types of Neurons
There are many neurons in the nervous system—a number in the trillions. And there are many different types of neurons. They can be classified by many different criteria. The first way to classify them is by the number of processes attached to the cell body. Using the standard model of neurons, one of these processes is the axon, and the rest are dendrites. Because information flows through the neuron from dendrites or cell bodies toward the axon, these names are based on the neuron’s polarity (Figure 13.2.2).
Figure 13.2.2. Neuron classification by shape. Unipolar cells have one process that includes both the axon and dendrite. Bipolar cells have two processes, the axon and a dendrite. Multipolar cells have more than two processes, the axon and two or more dendrites.
Unipolar cells have only one process emerging from the cell. True unipolar cells are only found in invertebrate animals, so the unipolar cells in humans are more appropriately called “pseudo-unipolar” cells. Invertebrate unipolar cells do not have dendrites. Human unipolar cells have an axon that emerges from the cell body, but it splits so that the axon can extend along a very long distance. At one end of the axon are dendrites, and at the other end, the axon forms synaptic connections with a target. Unipolar cells are exclusively sensory neurons and have two unique characteristics. First, their dendrites are receiving sensory information, sometimes directly from the stimulus itself. Secondly, the cell bodies of unipolar neurons are always found in ganglia. Sensory reception is a peripheral function (those dendrites are in the periphery, in the skin) so the cell body is in the periphery, though closer to the CNS in a ganglion. The axon projects from the dendrite endings, past the cell body in a ganglion, and into the central nervous system.
Bipolar cells have two processes, which extend from each end of the cell body, opposite to each other. One is the axon and one the dendrite. Bipolar cells are not very common. They are found in the olfactory epithelium (where smell stimuli are sensed), and as part of the retina.
Multipolar neurons are all the neurons that are not unipolar or bipolar. They have one axon and two or more dendrites (usually many more). With the exception of the unipolar sensory ganglion cells, and the two specific bipolar cells mentioned above, all other neurons are multipolar. Some cutting-edge research suggests that certain neurons in the CNS do not conform to the standard model of “one, and only one” axon. Some sources describe a fourth type of neuron, called an anaxonic neuron. The name suggests that it has no axon (an- = “without”), but this is not accurate. Anaxonic neurons are very small, and if you look through a microscope at the standard resolution used in histology (approximately 400X to 1000X total magnification), you will not be able to distinguish any process specifically as an axon or a dendrite. Any of those processes can function as an axon depending on the conditions at any given time. Nevertheless, even if they cannot be easily seen, and one specific process is definitively the axon, these neurons have multiple processes and are therefore multipolar.
Neurons can also be classified on the basis of where they are found, who found them, what they do, or even what chemicals they use to communicate with each other. Some neurons referred to in this section on the nervous system are named on the basis of those sorts of classifications (Figure 13.2.3). For example, a multipolar neuron that has a very important role to play in a part of the brain called the cerebellum is known as a Purkinje (commonly pronounced per-KIN-gee) cell. It is named after the anatomist who discovered it (Jan Evangilista Purkinje, 1787–1869).
Figure 13.2.3. Other neuron classifications. Three examples of neurons that are classified on the basis of other criteria. (a) The pyramidal cell is a multipolar cell with a cell body that is shaped something like a pyramid. (b) The Purkinje cell in the cerebellum was named after the scientist who originally described it. (c) Olfactory neurons are named for the functional group with which they belong.
Glial Cells
Glial cells, or neuroglia or simply glia, are the other type of cell found in nervous tissue. They are considered to be supporting cells, and many functions are directed at helping neurons complete their function for communication. The name glia comes from the Greek word that means “glue,” and was coined by the German pathologist Rudolph Virchow, who wrote in 1856: “This connective substance, which is in the brain, the spinal cord, and the special sense nerves, is a kind of glue (neuroglia) in which the nervous elements are planted.” Today, research into nervous tissue has shown that there are many deeper roles that these cells play. And research may find much more about them in the future.
There are six types of glial cells. Four of them are found in the CNS and two are found in the PNS. Table 13.2.1 outlines some common characteristics and functions.
Table 13.2.1. Glial cell types by location and basic function
CNS glia
PNS glia
Basic function
Astrocyte
Satellite cell
Support
Oligodendrocyte
Schwann cell
Insulation, myelination
Microglia
–
Immune surveillance and phagocytosis
Ependymal cell
–
Creating CSF
Glial Cells of the CNS
One cell providing support to neurons of the CNS is the astrocyte, so named because it appears to be star-shaped under the microscope (astro- = “star”). Astrocytes have many processes extending from their main cell body (not axons or dendrites like neurons, just cell extensions). Those processes extend to interact with neurons, blood vessels, or the connective tissue covering the CNS that is called the pia mater (Figure 13.2.4). They are supporting cells for the neurons in the central nervous system. Some ways in which they support neurons in the central nervous system are by maintaining the concentration of chemicals in the extracellular space, removing excess signalling molecules, reacting to tissue damage, and contributing to the blood-brain barrier (BBB). The blood-brain barrier is a physiological barrier that keeps many substances that circulate in the rest of the body from getting into the central nervous system, restricting what can cross from circulating blood into the CNS. Nutrient molecules, such as glucose or amino acids, can pass through the BBB, but other molecules cannot. This causes problems with drug delivery to the CNS. Pharmaceutical companies are challenged to design drugs that can cross the BBB as well as have an effect on the nervous system.
Figure 13.2.4. Glial cells of the CNS. The CNS has astrocytes, oligodendrocytes, microglia, and ependymal cells that support the neurons of the CNS in several ways.
Like a few other parts of the body, the brain has a privileged blood supply. Very little can pass through by diffusion. Most substances that cross the wall of a blood vessel into the CNS must do so through an active transport process. Because of this, only specific types of molecules can enter the CNS. Glucose—the primary energy source—is allowed, as are amino acids. Water and some other small particles, like gases and ions, can enter. But most everything else cannot, including white blood cells, which are one of the body’s main lines of defence. While this barrier protects the CNS from exposure to toxic or pathogenic substances, it also keeps out the cells that could protect the brain and spinal cord from disease and damage. The BBB also makes it harder for pharmaceuticals to be developed that can affect the nervous system. Aside from finding efficacious substances, the means of delivery is also crucial.
Also found in CNS tissue is the oligodendrocyte, sometimes called just “oligo,” which is the glial cell type that insulates axons in the CNS. The name means “cell of a few branches” (oligo- = “few”; dendro- = “branches”; -cyte = “cell”). There are a few processes that extend from the cell body. Each one reaches out and surrounds an axon to insulate it in myelin. One oligodendrocyte will provide the myelin for multiple axon segments, either for the same axon or for separate axons. The function of myelin will be discussed below.
Microglia are, as the name implies, smaller than most of the other glial cells. Ongoing research into these cells, although not entirely conclusive, suggests that they may originate as white blood cells, called macrophages, that become part of the CNS during early development. While their origin is not conclusively determined, their function is related to what macrophages do in the rest of the body. When macrophages encounter diseased or damaged cells in the rest of the body, they ingest and digest those cells or the pathogens that cause disease. Microglia are the cells in the CNS that can do this in normal, healthy tissue, and they are therefore also referred to as CNS-resident macrophages.
The ependymal cell is a glial cell that filters blood to make cerebrospinal fluid (CSF), the fluid that circulates through the CNS. Because of the privileged blood supply inherent in the BBB, the extracellular space in nervous tissue does not easily exchange components with the blood. Ependymal cells line each ventricle, one of four central cavities that are remnants of the hollow centre of the neural tube formed during the embryonic development of the brain. The choroid plexus is a specialised structure in the ventricles where ependymal cells come in contact with blood vessels and filter and absorb components of the blood to produce cerebrospinal fluid. Because of this, ependymal cells can be considered a component of the BBB, or a place where the BBB breaks down. These glial cells appear similar to epithelial cells, making a single layer of cells with little intracellular space and tight connections between adjacent cells. They also have cilia on their apical surface to help move the CSF through the ventricular space. The relationship of these glial cells to the structure of the CNS is seen in Figure 13.2.4.
Glial Cells of the PNS
One of the two types of glial cells found in the PNS is the satellite cell. Satellite cells are found in sensory and autonomic ganglia, where they surround the cell bodies of neurons. This accounts for the name, based on their appearance under the microscope. They provide support, performing similar functions in the periphery as astrocytes do in the CNS—except, of course, for establishing the BBB.
The second type of glial cell is the Schwann cell, which insulate axons with myelin in the periphery. Schwann cells are different than oligodendrocytes, in that a Schwann cell wraps around a portion of only one axon segment and no others. Oligodendrocytes have processes that reach out to multiple axon segments, whereas the entire Schwann cell surrounds just one axon segment. The nucleus and cytoplasm of the Schwann cell are on the edge of the myelin sheath. The relationship of these two types of glial cells to ganglia and nerves in the PNS is seen in Figure 13.2.5.
Figure 13.2.5. Glial cells of the PNS. The PNS has satellite cells and Schwann cells.
Myelin
The insulation for axons in the nervous system is provided by glial cells, oligodendrocytes in the CNS, and Schwann cells in the PNS. Whereas the manner in which either cell is associated with the axon segment, or segments, that it insulates is different, the means of myelinating an axon segment is mostly the same in the two situations. Myelin is a lipid-rich sheath that surrounds the axon and by doing so creates a myelin sheath that facilitates the transmission of electrical signals along the axon. The lipids are the phospholipids of the glial cell membrane. Myelin, however, is more than just the membrane of the glial cell. It also includes important proteins that are integral to that membrane. Some of the proteins help to hold the layers of the glial cell membrane closely together.
The appearance of the myelin sheath can be thought of as similar to the pastry wrapped around a hot dog for “pigs in a blanket” or a similar food. The glial cell is wrapped around the axon several times with little to no cytoplasm between the glial cell layers. For oligodendrocytes, the rest of the cell is separate from the myelin sheath as a cell process extends back toward the cell body. A few other processes provide the same insulation for other axon segments in the area. For Schwann cells, the outermost layer of the cell membrane contains cytoplasm and the nucleus of the cell as a bulge on one side of the myelin sheath. During development, the glial cell is loosely or incompletely wrapped around the axon (Figure 13.2.6a). The edges of this loose enclosure extend toward each other, and one end tucks under the other. The inner edge wraps around the axon, creating several layers, and the other edge closes around the outside so that the axon is completely enclosed.
Myelin sheaths can extend for one or two millimetres, depending on the diameter of the axon. Axon diameters can be as small as 1 to 20 micrometres. Because a micrometre is 1/1000 of a millimetre, this means that the length of a myelin sheath can be 100–1000 times the diameter of the axon. Figure 13.2.1, Figure 13.2.4 and Figure 13.2.5 show the myelin sheath surrounding an axon segment, but are not to scale. If the myelin sheath were drawn to scale, the neuron would have to be immense—covering an entire wall of the room in which you are sitting.
Several diseases can result from the demyelination of axons. The causes of these diseases are not the same; some have genetic causes, some are caused by pathogens, and others are the result of autoimmune disorders. Though the causes are varied, the results are similar. The myelin insulation of axons is compromised, making electrical signalling slower.
Multiple sclerosis (MS) is one such disease. It is an example of an autoimmune disease. The antibodies produced by lymphocytes (a type of white blood cell) mark myelin as something that should not be in the body. This causes inflammation and the destruction of the myelin in the central nervous system. As the insulation around the axons is destroyed by the disease, scarring becomes obvious. This is where the name of the disease comes from; sclerosis means hardening of tissue, which is what a scar is. Multiple scars are found in the white matter of the brain and spinal cord. The symptoms of MS include both somatic and autonomic deficits. Control of the musculature is compromised, as is control of organs such as the bladder.
Guillain-Barré (pronounced gee-YAN bah-RAY) syndrome is an example of a demyelinating disease of the peripheral nervous system. It is also the result of an autoimmune reaction, but the inflammation is in peripheral nerves. Sensory symptoms or motor deficits are common, and autonomic failures can lead to changes in the heart rhythm or a drop in blood pressure, especially when standing, which causes dizziness.
Section Review
Nervous tissue contains two major cell types, neurons and glial cells. Neurons are the cells responsible for communication through electrical signals. Glial cells are supporting cells, maintaining the environment around the neurons.
Neurons are polarised cells, based on the flow of electrical signals along their membrane. Signals are received at the dendrites, are passed along the cell body, and propagate along the axon towards the target, which may be another neuron, muscle tissue, or a gland. Many axons are insulated by a lipid-rich substance called myelin. Specific types of glial cells provide this insulation.
Several types of glial cells are found in the nervous system, and they can be categorised by the anatomical division in which they are found. In the CNS, astrocytes, oligodendrocytes, microglia, and ependymal cells are found. Astrocytes are important for maintaining the chemical environment around the neuron and are crucial for regulating the blood-brain barrier. Oligodendrocytes are the myelinating glia in the CNS. Microglia act as phagocytes and play a role in immune surveillance. Ependymal cells are responsible for filtering the blood to produce cerebrospinal fluid, which is a circulatory fluid that performs some of the functions of blood in the brain and spinal cord because of the BBB. In the PNS, satellite cells are supporting cells for the neurons, and Schwann cells insulate peripheral axons.
Distinguish the major functions of the nervous system: sensation, integration, and response
List the sequence of events in a simple sensory receptor–motor response pathway
Having looked at the components of nervous tissue, and the basic anatomy of the nervous system, next comes an understanding of how nervous tissue is capable of communicating within the nervous system. Before getting to the nuts and bolts of how this works, an illustration of how the components come together will be helpful. An example is summarised in Figure 13.3.1.
Figure 13.3.1. Testing the water. (1) The sensory neuron has endings in the skin that sense a stimulus such as water temperature. The strength of the signal that starts here is dependent on the strength of the stimulus. (2) The graded potential from the sensory endings, if strong enough, will initiate an action potential at the initial segment of the axon (which is immediately adjacent to the sensory endings in the skin). (3) The axon of the peripheral sensory neuron enters the spinal cord and contacts another neuron in the grey matter. The contact is a synapse where another graded potential is caused by the release of a chemical signal from the axon terminals. (4) An action potential is initiated at the initial segment of this neuron and travels up the sensory pathway to a region of the brain called the thalamus. Another synapse passes the information along to the next neuron. (5) The sensory pathway ends when the signal reaches the cerebral cortex. (6) After integration with neurons in other parts of the cerebral cortex, a motor command is sent from the precentral gyrus of the frontal cortex. (7) The upper motor neuron sends an action potential down to the spinal cord. The target of the upper motor neuron is the dendrites of the lower motor neuron in the grey matter of the spinal cord. (8) The axon of the lower motor neuron emerges from the spinal cord in a nerve and connects to a muscle through a neuromuscular junction to cause contraction of the target muscle.
Imagine you are about to take a shower in the morning before going to school. You have turned on the faucet to start the water as you prepare to get in the shower. After a few minutes, you expect the water to be a temperature that will be comfortable to enter, so you put your hand out into the spray of water. What happens next depends on how your nervous system interacts with the stimulus of the water temperature and what you do in response to that stimulus.
Found in the skin of your fingers or toes is a type of sensory receptor that is sensitive to temperature, called a thermoreceptor. When you place your hand under the shower (Figure 13.3.2), the cell membrane of the thermoreceptors changes its electrical state (voltage). The amount of change is dependent on the strength of the stimulus (how hot the water is). This is called a graded potential. If the stimulus is strong, the voltage of the cell membrane will change enough to generate an electrical signal that will travel down the axon. You have learned about this type of signalling before, with respect to the interaction of nerves and muscles at the neuromuscular junction. The voltage at which such a signal is generated is called the threshold, and the resulting electrical signal is called an action potential. In this example, the action potential travels—a process known as propagation—along the axon from the axon hillock to the axon terminals and into the synaptic end bulbs. When this signal reaches the end bulbs, it causes the release of a signalling molecule called a neurotransmitter.
Figure 13.3.2. The sensory input. Receptors in the skin sense the temperature of the water.
The neurotransmitter diffuses across the short distance of the synapse and binds to a receptor protein of the target neuron. When the molecular signal binds to the receptor, the cell membrane of the target neuron changes its electrical state and a new graded potential begins. If that graded potential is strong enough to reach threshold, the second neuron generates an action potential at its axon hillock. The target of this neuron is another neuron in the thalamus of the brain, the part of the CNS that acts as a relay for sensory information. At another synapse, neurotransmitter is released and binds to its receptor. The thalamus then sends the sensory information to the cerebral cortex, the outermost layer of grey matter in the brain, where conscious perception of that water temperature begins.
Within the cerebral cortex, information is processed among many neurons, integrating the stimulus of the water temperature with other sensory stimuli, with your emotional state (you just aren’t ready to wake up; the bed is calling to you), memories (of the lab notes you must study before a quiz). Finally, a plan is developed about what to do, whether that is to turn the temperature up, turn the whole shower off and go back to bed, or step into the shower. To do any of these things, the cerebral cortex must send a command out to your body to move muscles (Figure 13.3.3).
Figure 13.3.3. The motor response. On the basis of the sensory input and the integration in the CNS, a motor response is formulated and executed.
A region of the cortex is specialised for sending signals down to the spinal cord for movement. The upper motor neuron is in this region, called the precentral gyrus of the frontal cortex, which has an axon that extends all the way down the spinal cord. At the level of the spinal cord at which this axon makes a synapse, a graded potential occurs in the cell membrane of a lower motor neuron. This second motor neuron is responsible for causing muscle fibres to contract. In the manner described in the chapter on muscle tissue, an action potential travels along the motor neuron axon into the periphery. The axon terminates on muscle fibres at the neuromuscular junction. Acetylcholine is released at this specialised synapse, which causes the muscle action potential to begin, following a large potential known as an end plate potential. When the lower motor neuron excites the muscle fibre, it contracts. All of this occurs in a fraction of a second, but this story is the basis of how the nervous system functions.
Career Connections
Neurophysiologist
Understanding how the nervous system works could be a driving force in your career. Studying neurophysiology is a very rewarding path to follow. It means that there is a lot of work to do, but the rewards are worth the effort.
There are several ways in which a person can become a registered psychologist in Australia. The first step is to complete an undergraduate psychology course (e.g., bachelor’s degree, majoring in Psychology) which takes 3 years full-time to complete. The next step is to complete an additional year specialising in psychology (e.g., Honours or postgraduate diploma). The third step is to gain a provisional registration. This can be done in several ways. A person can either undergo ‘4 + 2 internship’ which includes a two year supervised practice, or a ‘5 + 1 internship’ which includes a fifth year study of psychology through a graduate diploma or a masters of professional psychology with one year of supervised practice. Both streams end with a general registration. There are also research streams which can be undertaken. A person can complete a two-year professional master’s degree, followed by a two-year supervised practice post professional master’s program, with 80 hours supervision and 80 hours continuing professional development (CPD) (PsyBA registrar program). Alternatively, someone can carry out a four-year combined professional masters and Doctor of Philosophy Course, followed by a 1.5 year of supervised practice and 60 hours of supervision plus 60 hours (CPD) (PsyBA registrar program). Finally, a person can complete a 3-4-year professional doctorate followed by one-year supervised practice with 40 hours supervision plus 40 hours CPD (PsyBA registrar program). All three of these final streams practice endorsement in a relevant stream.
Section Review
Sensation starts with the activation of a sensory ending, such as the thermoreceptor in the skin sensing the temperature of the water. The sensory endings in the skin initiate an electrical signal that travels along the sensory axon within a nerve into the spinal cord, where it synapses with a neuron in the grey matter of the spinal cord. The temperature information represented in that electrical signal is passed to the next neuron by a chemical signal that diffuses across the small gap of the synapse and initiates a new electrical signal in the target cell. That signal travels through the sensory pathway to the brain, passing through the thalamus, where conscious perception of the water temperature is made possible by the cerebral cortex. Following integration of that information with other cognitive processes and sensory information, the brain sends a command back down to the spinal cord to initiate a motor response by controlling a skeletal muscle. The motor pathway is composed of two cells, the upper motor neuron and the lower motor neuron. The upper motor neuron has its cell body in the cerebral cortex and synapses on a cell in the grey matter of the spinal cord. The lower motor neuron is that cell in the grey matter of the spinal cord and its axon extends into the periphery where it synapses with a skeletal muscle in a neuromuscular junction.
Describe the components of the membrane that establish the resting membrane potential
Describe the changes that occur to the membrane that result in the action potential
The functions of the nervous system—sensation, integration, and response—depend on the functions of the neurons underlying these pathways. To understand how neurons can communicate, it is necessary to describe the role of an excitable membrane in generating these signals. The basis of this communication is the action potential, which demonstrates how changes in the membrane can constitute a signal. Looking at the way these signals work in more variable circumstances involves a look at graded potentials, which will be covered in the next section.
Electrically Active Cell Membranes
Most cells in the body make use of charged particles, ions, to build up a charge across the cell membrane. Previously, this was shown to be a part of how muscle cells work. For skeletal muscles to contract, based on excitation–contraction coupling, requires input from a neuron. Both cells make use of the cell membrane to regulate ion movement between the extracellular fluid and cytosol.
As you learned in the chapter on cells, the cell membrane is primarily responsible for regulating what can cross the membrane and what stays on only one side. The cell membrane is a phospholipid bilayer, so only substances that can pass directly through the hydrophobic core can diffuse through unaided. Charged particles, which are hydrophilic, cannot pass through the cell membrane without assistance (Figure 13.4.1). Transmembrane proteins, specifically channel proteins, make this possible. Several passive transport channels, as well as active transport pumps, are necessary to generate a transmembrane potential and an action potential. Of special interest is the carrier protein referred to as the sodium/potassium pump that moves sodium ions (Na+) out of a cell and potassium ions (K+) into a cell, thus regulating ion concentration on both sides of the cell membrane.
Figure 13.4.1. Cell membrane and transmembrane proteins. The cell membrane is composed of a phospholipid bilayer and has many transmembrane proteins, including different types of channel proteins that serve as ion channels.
The sodium/potassium pump requires energy in the form of adenosine triphosphate (ATP), so it is also referred to as an ATPase. As was explained in the cell chapter, the concentration of Na+ is higher outside the cell than inside, and the concentration of K+ is higher inside the cell than outside. That means that this pump is moving the ions against the concentration gradients for sodium and potassium, which is why it requires energy. In fact, the pump basically maintains those concentration gradients.
Ion channels are pores that allow specific charged particles to cross the membrane in response to an existing concentration gradient. Proteins can span the cell membrane, including its hydrophobic core, and can interact with the charge of ions because of the varied properties of amino acids found within specific domains or regions of the protein channel. Hydrophobic amino acids are found in the domains that are apposed to the hydrocarbon tails of the phospholipids. Hydrophilic amino acids are exposed to the fluid environments of the extracellular fluid and cytosol. Additionally, the ions will interact with the hydrophilic amino acids, which will be selective for the charge of the ion. Channels for cations (positive ions) will have negatively charged side chains in the pore. Channels for anions (negative ions) will have positively charged side chains in the pore. This is called electrochemical exclusion, meaning that the channel pore is charge-specific.
Ion channels can also be specified by the diameter of the pore. The distance between the amino acids will be specific for the diameter of the ion when it dissociates from the water molecules surrounding it. Because of the surrounding water molecules, larger pores are not ideal for smaller ions because the water molecules will interact, by hydrogen bonds, more readily than the amino acid side chains. This is called size exclusion. Some ion channels are selective for charge but not necessarily for size, and thus are called a nonspecific channel. These nonspecific channels allow cations—particularly Na+, K+, and Ca2+—to cross the membrane, but exclude anions.
Ion channels do not always freely allow ions to diffuse across the membrane. Some are opened by certain events, meaning the channels are gated, so another way that channels can be categorised is based on how they are gated. Although these classes of ion channels are found primarily in the cells of nervous or muscular tissue, they also can be found in the cells of epithelial and connective tissues.
A ligand-gated channel opens because a signalling molecule, a ligand, binds to the extracellular region of the channel. This type of channel is also known as an ionotropic receptor because when the ligand, known as a neurotransmitter in the nervous system, binds to the protein, ions cross the membrane changing its charge (Figure 13.4.2).
Figure 13.4.2. Ligand-gated channels. When the ligand, in this case the neurotransmitter acetylcholine, binds to a specific location on the extracellular surface of the channel protein, the pore opens to allow select ions through. The ions, in this case, are cations of sodium, calcium, and potassium.
A mechanically gated channel opens because of a physical distortion of the cell membrane. Many channels associated with the sense of touch (somatosensation) are mechanically gated. For example, as pressure is applied to the skin, these channels open and allow ions to enter the cell. Similar to this type of channel would be the channel that opens based on temperature changes, as in testing the water in the shower (Figure 13.4.3).
Figure 13.4.3. Mechanically gated channels. When a mechanical change occurs in the surrounding tissue, such as pressure or touch, the channel is physically opened. Thermoreceptors work on a similar principle. When the local tissue temperature changes, the protein reacts by physically opening the channel.
A voltage-gated channel is a channel that responds to changes in the electrical properties of the membrane in which it is embedded. Normally, the inner portion of the membrane is at a negative voltage. When that voltage becomes less negative, the channel begins to allow ions to cross the membrane (Figure 13.4.4).
Figure 13.4.4. Voltage-gated channels. Voltage-gated channels open when the transmembrane voltage changes around them. Amino acids in the structure of the protein are sensitive to charge and cause the pore to open to the selected ion.
A leakage channel is randomly gated, meaning that it opens and closes at random, hence the reference to leaking. There is no actual event that opens the channel; instead, it has an intrinsic rate of switching between the open and closed states. Leakage channels contribute to the resting transmembrane voltage of the excitable membrane (Figure 13.4.5).
Figure 13.4.5. Leakage channels. In certain situations, ions need to move across the membrane randomly. The particular electrical properties of certain cells are modified by the presence of this type of channel.
The Membrane Potential
The electrical state of the cell membrane can have several variations. These are all variations in the membrane potential. A potential is a distribution of charge across the cell membrane, measured in millivolts (mV). The standard is to compare the inside of the cell relative to the outside, so the membrane potential is a value representing the charge on the intracellular side of the membrane based on the outside being zero, relatively speaking (Figure 13.4.6).
Figure 13.4.6. Measuring charge across a membrane with a voltmeter. A recording electrode is inserted into the cell and a reference electrode is outside the cell. By comparing the charge measured by these two electrodes, the transmembrane voltage is determined. It is conventional to express that value for the cytosol relative to the outside.
The concentration of ions in extracellular and intracellular fluids is balanced, with a net neutral charge. However, a slight difference in charge occurs right at the membrane surface, both internally and externally. It is the difference in this very limited region that has all the power in neurons (and muscle cells) to generate electrical signals, including action potentials.
Before these electrical signals can be described, the resting state of the membrane must be explained. When the cell is at rest, and the ion channels are closed (except for leakage channels which randomly open), ions are distributed across the membrane in a very predictable way. The concentration of Na+ outside the cell is 10 times greater than the concentration inside. Also, the concentration of K+ inside the cell is greater than outside. The cytosol contains a high concentration of anions, in the form of phosphate ions and negatively charged proteins. Large anions are a component of the inner cell membrane, including specialised phospholipids and proteins associated with the inner leaflet of the membrane (leaflet is a term used for one side of the lipid bilayer membrane). The negative charge is localised in the large anions.
With the ions distributed across the membrane at these concentrations, the difference in charge is measured at -70 mV, the value described as the resting membrane potential. The exact value measured for the resting membrane potential varies between cells, but -70 mV is most commonly used as this value. This voltage would be much lower except for the contributions of some important proteins in the membrane. Leakage channels allow Na+ to slowly move into the cell or K+ to slowly move out, and the Na+ /K+ pump restores them. This may appear to be a waste of energy, but each has a role in maintaining the membrane potential.
The Action Potential
Resting membrane potential describes the steady state of the cell, which is a dynamic process that is balanced by ion leakage and ion pumping. Without any outside influence, it will not change. To get an electrical signal started, the membrane potential must change.
This starts with a channel opening for Na+ in the membrane. Because the concentration of Na+ is higher outside the cell than inside the cell by a factor of 10, ions will rush into the cell that are driven largely by the concentration gradient. Because sodium is a positively charged ion, it will change the relative voltage immediately inside the cell relative to immediately outside. The resting potential is the state of the membrane at a voltage of -70 mV, so the sodium cation entering the cell will cause it to become less negative. This is known as depolarisation, meaning the membrane potential moves toward zero.
The concentration gradient for Na+ is so strong that it will continue to enter the cell even after the membrane potential has become zero, so that the voltage immediately around the pore begins to become positive. The electrical gradient also plays a role, as negative proteins below the membrane attract the sodium ion. The membrane potential will reach +30 mV by the time sodium has entered the cell.
As the membrane potential reaches +30 mV, other voltage-gated channels are opening in the membrane. These channels are specific for the potassium ion. A concentration gradient acts on K+, as well. As K+ starts to leave the cell, taking a positive charge with it, the membrane potential begins to move back toward its resting voltage. This is called repolarisation, meaning that the membrane voltage moves back toward the -70mV value of the resting membrane potential.
Repolarisation returns the membrane potential to the -70mV value that indicates the resting potential, but it actually overshoots that value. Potassium ions reach equilibrium when the membrane voltage is below -70 mV, so a period of hyperpolarisation occurs while the K+ channels are open. Those K+ channels are slightly delayed in closing, accounting for this short overshoot.
What has been described here is the action potential, which is presented as a graph of voltage over time in Figure 13.4.7. It is the electrical signal that nervous tissue generates for communication. The change in the membrane voltage from -70 mV at rest to +30 mV at the end of depolarisation is a 100-mV change. That can also be written as a 0.1-V change. To put that value in perspective, think about a battery. An AA battery that you might find in a television remote has a voltage of 1.5 V, or a 9-V battery (the rectangular battery with two posts on one end) is, obviously, 9 V. The change seen in the action potential is one or two orders of magnitude less than the charge in these batteries. In fact, the membrane potential can be described as a battery. A charge is stored across the membrane that can be released under the correct conditions. A battery in your remote has stored a charge that is “released” when you push a button.
Figure 13.4.7. Graph of action potential. Plotting voltage measured across the cell membrane against time, the action potential begins with depolarisation, followed by repolarisation, which goes past the resting potential into hyperpolarisation, and finally the membrane returns to rest.
The question is, now, what initiates the action potential? The description above conveniently glosses over that point. But it is vital to understanding what is happening. The membrane potential will stay at the resting voltage until something changes. The description above just says that a Na+ channel opens. Now, to say “a channel opens” does not mean that one individual transmembrane protein changes. Instead, it means that one kind of channel opens. There are a few different types of channels that allow Na+ to cross the membrane. A ligand-gated Na+ channel will open when a neurotransmitter binds to it and a mechanically gated Na+ channel will open when a physical stimulus affects a sensory receptor (like pressure applied to the skin compresses a touch receptor). Whether it is a neurotransmitter binding to its receptor protein or a sensory stimulus activating a sensory receptor cell, some stimulus gets the process started. Sodium starts to enter the cell and the membrane becomes less negative.
A third type of channel that is an important part of depolarisation in the action potential is the voltage-gated Na+ channel. The channels that start depolarising the membrane because of a stimulus help the cell to depolarise from -70 mV to -55 mV. Once the membrane reaches that voltage, the voltage-gated Na+ channels open. This is what is known as the threshold. Any depolarisation that does not change the membrane potential to -55 mV or higher will not reach threshold and thus will not result in an action potential. Also, any stimulus that depolarises the membrane to -55 mV or beyond will cause a large number of channels to open and an action potential will be initiated.
Because of the threshold, the action potential can be likened to a digital event—it either happens or it does not. If the threshold is not reached, then no action potential occurs. If depolarisation reaches -55 mV, then the action potential continues and runs all the way to +30 mV, at which K+ causes repolarisation, including the hyperpolarising overshoot. Also, those changes are the same for every action potential, which means that once the threshold is reached, the exact same thing happens. A stronger stimulus, which might depolarise the membrane well past threshold, will not make a “bigger” action potential. Action potentials are “all or none.” Either the membrane reaches the threshold and everything occurs as described above, or the membrane does not reach the threshold and nothing else happens. All action potentials peak at the same voltage (+30 mV), so one action potential is not bigger than another. Stronger stimuli will initiate multiple action potentials more quickly, but the individual signals are not bigger. Thus, for example, you will not feel a greater sensation of pain, or have a stronger muscle contraction, because of the size of the action potential because they are not different sizes.
As we have seen, the depolarisation and repolarisation of an action potential are dependent on two types of channels (the voltage-gated Na+ channel and the voltage-gated K+ channel). The voltage-gated Na+ channel actually has two gates. One is the activation gate, which opens when the membrane potential crosses -55 mV. The other gate is the inactivation gate, which closes after a specific period of time—on the order of a fraction of a millisecond. When a cell is at rest, the activation gate is closed, and the inactivation gate is open. However, when the threshold is reached, the activation gate opens, allowing Na+ to rush into the cell. Timed with the peak of depolarisation, the inactivation gate closes. During repolarisation, no more sodium can enter the cell. When the membrane potential passes -55 mV again, the activation gate closes. After that, the inactivation gate re-opens, making the channel ready to start the whole process over again.
The voltage-gated K+ channel has only one gate, which is sensitive to a membrane voltage of -50 mV. However, it does not open as quickly as the voltage-gated Na+ channel does. It might take a fraction of a millisecond for the channel to open once that voltage has been reached. The timing of this coincides exactly with when the Na+ flow peaks, so voltage-gated K+ channels open just as the voltage-gated Na+ channels are being inactivated. As the membrane potential repolarises and the voltage passes -50 mV again, the channel closes—again, with a little delay. Potassium continues to leave the cell for a short while and the membrane potential becomes more negative, resulting in the hyperpolarising overshoot. Then the channel closes again, and the membrane can return to the resting potential because of the ongoing activity of the non-gated channels and the Na+ /K+ pump.
All of this takes place within approximately two milliseconds (Figure 13.4.8). While an action potential is in progress, another one cannot be initiated. That effect is referred to as the refractory period. There are two phases of the refractory period: the absolute refractory period and the relative refractory period. During the absolute phase, another action potential will not start. This is because of the inactivation gate of the voltage-gated Na+ channel. Once that channel is back to its resting conformation (less than -55 mV), a new action potential could be started, but only by a stronger stimulus than the one that initiated the current action potential. This is because of the flow of K+ out of the cell. Because that ion is rushing out, any Na+ that tries to enter will not depolarise the cell but will only keep the cell from hyperpolarising.
Figure 13.4.8. Stages of an action potential. Plotting voltage measured across the cell membrane against time, the events of the action potential can be related to specific changes in the membrane voltage. (1) At rest, the membrane voltage is -70 mV. (2) The membrane begins to depolarise when an external stimulus is applied. (3) The membrane voltage begins a rapid rise toward +30 mV. (4) The membrane voltage starts to return to a negative value. (5) Repolarisation continues past the resting membrane voltage, resulting in hyperpolarisation. (6) The membrane voltage returns to the resting value shortly after hyperpolarisation.
Propagation of the Action Potential
The action potential is initiated at the beginning of the axon, at what is called the initial segment. There is a high density of voltage-gated Na+ channels so that rapid depolarisation can take place here. Going down the length of the axon, the action potential is propagated because more voltage-gated Na+ channels are opened as the depolarisation spreads. This spreading occurs because Na+ enters through the channel and moves along the inside of the cell membrane. As the Na+ moves, or flows, a short distance along the cell membrane, its positive charge depolarises a little more of the cell membrane. As that depolarisation spreads, new voltage-gated Na+ channels open and more ions rush into the cell, spreading the depolarisation a little farther.
Because voltage-gated Na+ channels are inactivated at the peak of the depolarisation, they cannot be opened again for a brief time—the absolute refractory period. Because of this, depolarisation spreading back toward previously opened channels has no effect. The action potential must propagate toward the axon terminals; as a result, the polarity of the neuron is maintained, as mentioned above.
Propagation, as described above, applies to unmyelinated axons. When myelination is present, the action potential propagates differently. Sodium ions that enter the cell at the initial segment start to spread along the length of the axon segment, but there are no voltage-gated Na+ channels until the first node of Ranvier. Because there is not constant opening of these channels along the axon segment, the depolarisation spreads at an optimal speed. The distance between nodes is the optimal distance to keep the membrane still depolarised above threshold at the next node. As Na+ spreads along the inside of the membrane of the axon segment, the charge starts to dissipate. If the node were any farther down the axon, that depolarisation would have fallen off too much for voltage-gated Na+ channels to be activated at the next node of Ranvier. If the nodes were any closer together, the speed of propagation would be slower.
Propagation along an unmyelinated axon is referred to as continuous conduction; along the length of a myelinated axon, it is saltatory conduction. Continuous conduction is slow because there are always voltage-gated Na+ channels opening, and more and more Na+ is rushing into the cell. Saltatory conduction is faster because the action potential basically jumps from one node to the next (saltare = “to leap”), and the new influx of Na+ renews the depolarised membrane. Along with the myelination of the axon, the diameter of the axon can influence the speed of conduction. Much as water runs faster in a wide river than in a narrow creek, Na+ -based depolarisation spreads faster down a wide axon than down a narrow one. This concept is known as resistance and is generally true for electrical wires or plumbing, just as it is true for axons, although the specific conditions are different at the scales of electrons or ions versus water in a river.
Homeostatic Imbalances
Potassium Concentration
Glial cells, especially astrocytes, are responsible for maintaining the chemical environment of the CNS tissue. The concentrations of ions in the extracellular fluid are the basis for how the membrane potential is established and changes in electrochemical signalling. If the balance of ions is upset, drastic outcomes are possible.
Normally the concentration of K+ is higher inside the neuron than outside. After the repolarising phase of the action potential, K+ leakage channels and the Na+ /K+ pump ensure that the ions return to their original locations. Following a stroke or other ischemic event, extracellular K+ levels are elevated. The astrocytes in the area are equipped to clear excess K+ to aid the pump. But when the level is far out of balance, the effects can be irreversible.
Section Review
The nervous system is characterised by electrical signals that are sent from one area to another. Whether those areas are close or very far apart, the signal must travel along an axon. The basis of the electrical signal is the controlled distribution of ions across the membrane. Transmembrane ion channels regulate when ions can move in or out of the cell, so that a precise signal is generated. This signal is the action potential which has a very characteristic shape based on voltage changes across the membrane in a given time period.
The membrane is normally at rest with established Na+ and K+ concentrations on either side. A stimulus will start the depolarisation of the membrane, and voltage-gated channels will result in further depolarisation followed by repolarisation of the membrane. A slight overshoot of hyperpolarisation marks the end of the action potential. While an action potential is in progress, another cannot be generated under the same conditions. While the voltage-gated Na+ channel is inactivated, absolutely no action potentials can be generated. Once that channel has returned to its resting state, a new action potential is possible, but it must be started by a relatively stronger stimulus to overcome the K+ leaving the cell.
The action potential travels down the axon as voltage-gated ion channels are opened by the spreading depolarisation. In unmyelinated axons, this happens in a continuous fashion because there are voltage-gated channels throughout the membrane. In myelinated axons, propagation is described as saltatory because voltage-gated channels are only found at the nodes of Ranvier and the electrical events seem to “jump” from one node to the next. Saltatory conduction is faster than continuous conduction, meaning that myelinated axons propagate their signals faster. The diameter of the axon also makes a difference as ions diffusing within the cell have less resistance in a wider space.
Explain the differences between the types of graded potentials
Categorise the major neurotransmitters by chemical type and effect
The electrical changes taking place within a neuron, as described in the previous section, are similar to a light switch being turned on. A stimulus starts the depolarisation, but the action potential runs on its own once a threshold has been reached. The question is now, “What flips the light switch on?” Temporary changes to the cell membrane voltage can result from neurons receiving information from the environment, or from the action of one neuron on another. These special types of potentials influence a neuron and determine whether an action potential will occur or not. Many of these transient signals originate at the synapse.
Graded Potentials
Local changes in the membrane potential are called graded potentials and are usually associated with the dendrites of a neuron. The amount of change in the membrane potential is determined by the size of the stimulus that causes it. In the example of testing the temperature of the shower, slightly warm water would only initiate a small change in a thermoreceptor, whereas hot water would cause a large amount of change in the membrane potential.
Graded potentials can be of two sorts, either they are depolarising or hyperpolarising (Figure 13.5.1). For a membrane at the resting potential, a graded potential represents a change in that voltage either above -70 mV or below -70 mV. Depolarising graded potentials are often the result of Na+ or Ca 2+ entering the cell. Both ions have higher concentrations outside the cell than inside; because they have a positive charge, they will move into the cell causing it to become less negative relative to the outside. Hyperpolarising graded potentials can be caused by K+ leaving the cell or Cl– entering the cell. If a positive charge moves out of a cell, the cell becomes more negative; if a negative charge enters the cell, the same thing happens.
Figure 13.5.1. Graded potentials. Graded potentials are temporary changes in the membrane voltage, the characteristics of which depend on the size of the stimulus. Some types of stimuli cause depolarisation of the membrane, whereas others cause hyperpolarisation. It depends on the specific ion channels that are activated in the cell membrane.
Types of Graded Potentials
For the unipolar cells of sensory neurons—both those with free nerve endings and those within encapsulations—graded potentials develop in the dendrites that influence the generation of an action potential in the axon of the same cell. This is called a generator potential. For other sensory receptor cells, such as taste cells or photoreceptors of the retina, graded potentials in their membranes result in the release of neurotransmitters at synapses with sensory neurons. This is called a receptor potential.
A postsynaptic potential (PSP) is the graded potential in the dendrites of a neuron that is receiving synapses from other cells. Postsynaptic potentials can be depolarising or hyperpolarising. Depolarisation in a postsynaptic potential is called an excitatory postsynaptic potential (EPSP) because it causes the membrane potential to move toward threshold. Hyperpolarisation in a postsynaptic potential is an inhibitory postsynaptic potential (IPSP) because it causes the membrane potential to move away from threshold.
Summation
All types of graded potentials will result in small changes of either depolarisation or hyperpolarisation in the voltage of a membrane. These changes can lead to the neuron reaching threshold if the changes add together, or summate. The combined effects of different types of graded potentials are illustrated in Figure 13.5.2. If the total change in voltage in the membrane is a positive 15 mV, meaning that the membrane depolarises from -70 mV to -55 mV, then the graded potentials will result in the membrane reaching threshold.
For receptor potentials, threshold is not a factor because the change in membrane potential for receptor cells directly causes neurotransmitter release. However, generator potentials can initiate action potentials in the sensory neuron axon, and postsynaptic potentials can initiate an action potential in the axon of other neurons. Graded potentials summate at a specific location at the beginning of the axon to initiate the action potential, namely the initial segment. For sensory neurons, which do not have a cell body between the dendrites and the axon, the initial segment is directly adjacent to the dendritic endings. For all other neurons, the axon hillock is essentially the initial segment of the axon, and it is where summation takes place. These locations have a high density of voltage-gated Na+ channels that initiate the depolarising phase of the action potential.
Summation can be spatial or temporal, meaning it can be the result of multiple graded potentials at different locations on the neuron, or all at the same place but separated in time. Spatial summation is related to associating the activity of multiple inputs to a neuron with each other. Temporal summation is the relationship of multiple action potentials from a single cell resulting in a significant change in the membrane potential. Spatial and temporal summation can act together, as well.
Figure 13.5.2. Postsynaptic potential summation. The result of summation of postsynaptic potentials is the overall change in the membrane potential. At point A, several different excitatory postsynaptic potentials add up to a large depolarisation. At point B, a mix of excitatory and inhibitory postsynaptic potentials result in a different end result for the membrane potential.
Synapses
There are two types of connections between electrically active cells, chemical synapses and electrical synapses. In a chemical synapse, a chemical signal—namely, a neurotransmitter—is released from one cell and it affects the other cell. In an electrical synapse, there is a direct connection between the two cells so that ions can pass directly from one cell to the next. If one cell is depolarised in an electrical synapse, the joined cell also depolarises because the ions pass between the cells. Chemical synapses involve the transmission of chemical information from one cell to the next. This section will concentrate on the chemical type of synapse.
An example of a chemical synapse is the neuromuscular junction (NMJ) described in the chapter on muscle tissue. In the nervous system, there are many more synapses that are essentially the same as the NMJ. All synapses have common characteristics, which can be summarised in this list:
presynaptic element
neurotransmitter (packaged in vesicles)
synaptic cleft
receptor proteins
postsynaptic element
neurotransmitter elimination or re-uptake
For the NMJ, these characteristics are as follows: the presynaptic element is the motor neuron’s axon terminals, the neurotransmitter is acetylcholine, the synaptic cleft is the space between the cells where the neurotransmitter diffuses, the receptor protein is the nicotinic acetylcholine receptor, the postsynaptic element is the sarcolemma of the muscle cell, and the neurotransmitter is eliminated by acetylcholinesterase. Other synapses are similar to this, and the specifics are different, but they all contain the same characteristics.
Neurotransmitter Release
When an action potential reaches the axon terminals, voltage-gated Ca 2+ channels in the membrane of the synaptic end bulb open. The concentration of Ca 2+ increases inside the end bulb, and the Ca 2+ ion associates with proteins in the outer surface of neurotransmitter vesicles. The Ca2+ + facilitates the merging of the vesicle with the presynaptic membrane so that the neurotransmitter is released through exocytosis into the small gap between the cells, known as the synaptic cleft.
Once in the synaptic cleft, the neurotransmitter diffuses the short distance to the postsynaptic membrane and can interact with neurotransmitter receptors. Receptors are specific for the neurotransmitter, and the two fit together like a key and lock. One neurotransmitter binds to its receptor and will not bind to receptors for other neurotransmitters, making the binding a specific chemical event (Figure 13.5.3).
Figure 13.5.3. The synapse. The synapse is a connection between a neuron and its target cell (which is not necessarily a neuron). The presynaptic element is the synaptic end bulb of the axon where Ca2+ enters the bulb to cause vesicle fusion and neurotransmitter release. The neurotransmitter diffuses across the synaptic cleft to bind to its receptor. The neurotransmitter is cleared from the synapse either by enzymatic degradation, neuronal reuptake, or glial reuptake.
Neurotransmitter Systems
There are several systems of neurotransmitters that are found at various synapses in the nervous system. These groups refer to the chemicals that are the neurotransmitters, and within the groups are specific systems.
The first group, which is a neurotransmitter system of its own, is the cholinergic system. It is the system based on acetylcholine. This includes the NMJ as an example of a cholinergic synapse, but cholinergic synapses are found in other parts of the nervous system. They are in the autonomic nervous system, as well as distributed throughout the brain.
The cholinergic system has two types of receptors, the nicotinic receptor is found in the NMJ as well as other synapses. There is also an acetylcholine receptor known as the muscarinic receptor. Both receptors are named for drugs that interact with the receptor in addition to acetylcholine. Nicotine will bind to the nicotinic receptor and activate it similar to acetylcholine. Muscarine, a product of certain mushrooms, will bind to the muscarinic receptor. However, nicotine will not bind to the muscarinic receptor and muscarine will not bind to the nicotinic receptor.
Another group of neurotransmitters are amino acids. This includes glutamate (Glu), GABA (gamma-aminobutyric acid, a derivative of glutamate), and glycine (Gly). These amino acids have an amino group and a carboxyl group in their chemical structures. Glutamate is one of the 20 amino acids that are used to make proteins. Each amino acid neurotransmitter would be part of its own system, namely the glutamatergic, GABAergic, and glycinergic systems. They each have their own receptors and do not interact with each other. Amino acid neurotransmitters are eliminated from the synapse by reuptake. A pump in the cell membrane of the presynaptic element, or sometimes a neighbouring glial cell, will clear the amino acid from the synaptic cleft so that it can be recycled, repackaged in vesicles, and released again.
Another class of neurotransmitter is the biogenic amine, a group of neurotransmitters that are enzymatically made from amino acids. They have amino groups in them, but no longer have carboxyl groups and are therefore no longer classified as amino acids. Serotonin is made from tryptophan. It is the basis of the serotonergic system, which has its own specific receptors. Serotonin is transported back into the presynaptic cell for repackaging.
Other biogenic amines are made from tyrosine, and include dopamine, adrenaline [renal = “kidney”] (US: epinephrine [epi- = “on”; “-nephrine” = kidney]) and noradrenaline (US: norepinephrine). Dopamine is part of its own system, the dopaminergic system, which has dopamine receptors. Dopamine is removed from the synapse by transport proteins in the presynaptic cell membrane. Noradrenaline and adrenaline belong to the adrenergic neurotransmitter system. The two molecules are very similar and bind to the same receptors, which are referred to as alpha and beta receptors. Noradrenaline and adrenaline are also transported back into the presynaptic cell. The adrenal gland produces adrenaline and noradrenaline to be released into the blood stream as hormones.
A neuropeptide is a neurotransmitter molecule made up of chains of amino acids connected by peptide bonds. This is what a protein is, but the term protein implies a certain length to the molecule. Some neuropeptides are quite short, such as met-enkephalin, which is five amino acids long. Others are long, such as beta-endorphin, which is 31 amino acids long. Neuropeptides are often released at synapses in combination with another neurotransmitter, and they often act as hormones in other systems of the body, such as vasoactive intestinal peptide (VIP) or substance P.
The effect of a neurotransmitter on the postsynaptic element is entirely dependent on the receptor protein. First, if there is no receptor protein in the membrane of the postsynaptic element, then the neurotransmitter has no effect. The depolarising or hyperpolarising effect is also dependent on the receptor. When acetylcholine binds to the nicotinic receptor, the postsynaptic cell is depolarised. This is because the receptor is a cation channel and positively charged Na+ will rush into the cell. However, when acetylcholine binds to the muscarinic receptor, of which there are several variants, it might cause depolarisation or hyperpolarisation of the target cell.
The amino acid neurotransmitters, glutamate, glycine, and GABA are almost exclusively associated with just one effect. Glutamate is considered an excitatory amino acid, but only because Glu receptors in the adult cause depolarisation of the postsynaptic cell. Glycine and GABA are considered inhibitory amino acids, again because their receptors cause hyperpolarisation.
The biogenic amines have mixed effects, for example, the dopamine receptors that are classified as D1 receptors are excitatory whereas D2-type receptors are inhibitory. Biogenic amine receptors and neuropeptide receptors can have even more complex effects because some may not directly affect the membrane potential, but rather have an effect on gene transcription or other metabolic processes in the neuron. The characteristics of the various neurotransmitter systems presented in this section are organised in Table 13.5.1.
The important thing to remember about neurotransmitters, and signalling chemicals in general, is that the effect is entirely dependent on the receptor. Neurotransmitters bind to one of two classes of receptors at the cell surface, ionotropic or metabotropic (Figure 13.5.4). Ionotropic receptors are ligand-gated ion channels, such as the nicotinic receptor for acetylcholine or the glycine receptor. A metabotropic receptor involves a complex of proteins that result in metabolic changes within the cell. The receptor complex includes the transmembrane receptor protein, a G protein, and an effector protein. The neurotransmitter, referred to as the first messenger, binds to the receptor protein on the extracellular surface of the cell, and the intracellular side of the protein initiates activity of the G protein. The G protein is a guanosine triphosphate (GTP) hydrolase that physically moves from the receptor protein to the effector protein to activate the latter. An effector protein is an enzyme that catalyses the generation of a new molecule, which acts as the intracellular mediator of the signal that binds to the receptor. This intracellular mediator is called the second messenger.
Different receptors use different second messengers. Two common examples of second messengers are cyclic adenosine monophosphate (cAMP) and inositol triphosphate (IP3). The enzyme adenylate cyclase (an example of an effector protein) makes cAMP, and phospholipase C is the enzyme that makes IP3. Second messengers, after they are produced by the effector protein, cause metabolic changes within the cell. These changes are most likely the activation of other enzymes in the cell. In neurons, they often modify ion channels, either opening or closing them. These enzymes can also cause changes in the cell, such as the activation of genes in the nucleus, and therefore the increased synthesis of proteins. In neurons, these kinds of changes are often the basis of stronger connections between cells at the synapse and may be the basis of learning and memory.
Figure 13.5.4. Receptor types. (a) An ionotropic receptor is a channel that opens when the neurotransmitter binds to it. (b) A metabotropic receptor is a complex that causes metabolic changes in the cell when the neurotransmitter binds to it (1). After binding, the G protein hydrolyses GTP and moves to the effector protein (2). When the G protein contacts the effector protein, a second messenger is generated, such as cAMP (3). The second messenger can then go on to cause changes in the neuron, such as opening or closing ion channels, metabolic changes, and changes in gene transcription.
Table 13.5.1. Characteristics of neurotransmitter system
5-HT receptors, D1 and D2 receptors, α-adrenergic and β-adrenergic receptors
Receptors are too numerous to list, but are specific to peptides
Elimination
Degradation by acetylcholinesterase
Reuptake by neurons or glia
Reuptake by neurons
Degradation by enzymes called peptidases
Postsynaptic effect
Muscarinic receptors can cause both depolarisation or hyperpolarisation depending on the subtype
Glu receptors cause depolarisation.
Gly and GABA receptors cause hyperpolarisation
Depolarisation or hyperpolarisation depends on the specific receptor, for example, D1 receptors cause depolarisation and D2 receptors cause hyperpolarisation
Depolarisation or hyperpolarisation depends on the specific receptor
Disorders of the Nervous System
The underlying cause of some neurodegenerative diseases, such as Alzheimer’s and Parkinson’s, appears to be related to proteins—specifically, to proteins behaving badly. One of the strongest theories of what causes Alzheimer’s disease is based on the accumulation of beta-amyloid plaques, dense conglomerations of a protein that is not functioning correctly. Parkinson’s disease is linked to an increase in a protein known as alpha-synuclein that is toxic to the cells of the substantia nigra nucleus in the midbrain.
For proteins to function correctly, they are dependent on their three-dimensional shape. The linear sequence of amino acids folds into a three-dimensional shape that is based on the interactions between and among those amino acids. When the folding is disturbed, and proteins take on a different shape, they stop functioning correctly. But the disease is not necessarily the result of functional loss of these proteins; rather, these altered proteins start to accumulate and may become toxic. In Alzheimer’s, the hallmark of the disease is the accumulation of these amyloid plaques in the cerebral cortex. The term coined to describe this sort of disease is “proteopathy” and it includes other diseases. Creutzfeld-Jacob disease, the human variant of the prion disease known as mad cow disease in the bovine, also involves the accumulation of amyloid plaques, similar to Alzheimer’s. Diseases of other organ systems can fall into this group as well, such as cystic fibrosis or type 2 diabetes. Recognising the relationship between these diseases has suggested new therapeutic possibilities. Interfering with the accumulation of the proteins, and possibly as early as their original production within the cell, may unlock new ways to alleviate these devastating diseases.
Section Review
The basis of the electrical signal within a neuron is the action potential that propagates down the axon. For a neuron to generate an action potential, it needs to receive input from another source, either another neuron or a sensory stimulus. That input will result in opening ion channels in the neuron, resulting in a graded potential based on the strength of the stimulus. Graded potentials can be depolarising or hyperpolarising and can summate to affect the probability of the neuron reaching threshold.
Graded potentials can be the result of sensory stimuli. If the sensory stimulus is received by the dendrites of a unipolar sensory neuron, such as the sensory neuron ending in the skin, the graded potential is called a generator potential because it can directly generate the action potential in the initial segment of the axon. If the sensory stimulus is received by a specialised sensory receptor cell, the graded potential is called a receptor potential. Graded potentials produced by interactions between neurons at synapses are called postsynaptic potentials (PSPs). A depolarising graded potential at a synapse is called an excitatory PSP, and a hyperpolarising graded potential at a synapse is called an inhibitory PSP.
Synapses are the contacts between neurons, which can either be chemical or electrical in nature. Chemical synapses are far more common. At a chemical synapse, neurotransmitter is released from the presynaptic element and diffuses across the synaptic cleft. The neurotransmitter binds to a receptor protein and causes a change in the postsynaptic membrane (the PSP). The neurotransmitter must be inactivated or removed from the synaptic cleft so that the stimulus is limited in time.
The particular characteristics of a synapse vary based on the neurotransmitter system produced by that neuron. The cholinergic system is found at the neuromuscular junction and in certain places within the nervous system. Amino acids, such as glutamate, glycine, and gamma-aminobutyric acid (GABA) are used as neurotransmitters. Other neurotransmitters are the result of amino acids being enzymatically changed, as in the biogenic amines, or being covalently bonded together, as in the neuropeptides.
Describe the growth and differentiation of the neural tube
Relate the different stages of development to the adult structures of the central nervous system
Explain the expansion of the ventricular system of the adult brain from the central canal of the neural tube
Describe the connections of the diencephalon and cerebellum based on patterns of embryonic development
The brain is a complex organ composed of grey parts and white matter, which can be hard to distinguish. Starting from an embryologic perspective allows you to understand more easily how the parts relate to each other. The embryonic nervous system begins as a very simple structure—just a straight line, which then gets increasingly complex. Looking at the development of the nervous system with a couple of early snapshots makes it easier to understand the whole complex system.
Many structures that appear to be adjacent in the adult brain are not connected, and the connections that exist may seem arbitrary. But there is an underlying order to the system that comes from how different parts develop. By following the developmental pattern, it is possible to learn what the major regions of the nervous system are.
The Neural Tube
To begin, a sperm cell and an egg cell fuse to become a fertilised egg. The fertilised egg cell, or zygote, starts dividing to generate the cells that make up an entire organism. Sixteen days after fertilisation, the developing embryo’s cells belong to one of three germ layers that give rise to the different tissues in the body. The endoderm, or inner tissue, is responsible for generating the lining tissues of various spaces within the body, such as the mucosae of the digestive and respiratory systems. The mesoderm, or middle tissue, gives rise to most of the muscle and connective tissues. Finally, the ectoderm, or outer tissue, develops into the integumentary system (the skin) and the nervous system. It is probably not difficult to see that the outer tissue of the embryo becomes the outer covering of the body. But how is it responsible for the nervous system?
As the embryo develops, a portion of the ectoderm differentiates into a specialised region of neuroectoderm, which is the precursor for the tissue of the nervous system. Molecular signals induce cells in this region to differentiate into the neuroepithelium, forming a neural plate. The cells then begin to change shape, causing the tissue to buckle and fold inward (Figure 13.6.1). A neural groove forms, visible as a line along the dorsal surface of the embryo. The ridge-like edge on either side of the neural groove is referred as the neural fold. As the neural folds come together and converge, the underlying structure forms into a tube just beneath the ectoderm called the neural tube. Cells from the neural folds then separate from the ectoderm to form a cluster of cells referred to as the neural crest, which runs lateral to the neural tube. The neural crest migrates away from the nascent, or embryonic, central nervous system (CNS) that will form along the neural groove and develops into several parts of the peripheral nervous system (PNS), including the enteric nervous tissue. Many tissues that are not part of the nervous system also arise from the neural crest, such as craniofacial cartilage and bone, and melanocytes.
Figure 13.6.1. Early embryonic development of nervous system. The neuroectoderm begins to fold inward to form the neural groove. As the two sides of the neural groove converge, they form the neural tube, which lies beneath the ectoderm. The anterior end of the neural tube will develop into the brain, and the posterior portion will become the spinal cord. The neural crest develops into peripheral structures.
At this point, the early nervous system is a simple, hollow tube. It runs from the anterior end of the embryo to the posterior end. Beginning at 25 days, the anterior end develops into the brain, and the posterior portion becomes the spinal cord. This is the most basic arrangement of tissue in the nervous system, and it gives rise to the more complex structures by the fourth week of development.
Primary Vesicles
As the anterior end of the neural tube starts to develop into the brain, it undergoes a couple of enlargements; the result is the production of sac-like vesicles. Similar to a child’s balloon animal, the long, straight neural tube begins to take on a new shape. Three vesicles form at the first stage, which are called primary vesicles. These vesicles are given names that are based on Greek words, the main root word being enkephalon, which means “brain” (en- = “inside”; kephalon = “head”). The prefix to each corresponds to its position along the length of the developing nervous system.
The mesencephalon (mes- = “middle”) is the next vesicle, which can be called the midbrain. The third vesicle at this stage is the rhombencephalon. The first part of this word is also the root of the word rhombus, which is a geometrical figure with four sides of equal length (a square is a rhombus with 90° angles). Whereas prosencephalon and mesencephalon translate into the English words forebrain and midbrain, there is not a word for “four-sided-figure-brain.” However, the third vesicle can be called the hindbrain. One way of thinking about how the brain is arranged is to use these three regions—forebrain, midbrain, and hindbrain—which are based on the primary vesicle stage of development (Figure 13.6.2 a).
Secondary Vesicles
The brain continues to develop, and the vesicles differentiate further (see Figure 13.6.2b). The three primary vesicles become five secondary vesicles. The prosencephalon enlarges into two new vesicles called the telencephalon and the diencephalon. The telecephalon will become the cerebrum. The diencephalon gives rise to several adult structures; two that will be important are the thalamus and the hypothalamus. In the embryonic diencephalon, a structure known as the eye cup develops, which will eventually become the retina, the nervous tissue of the eye called the retina. This is a rare example of nervous tissue developing as part of the CNS structures in the embryo but becoming a peripheral structure in the fully formed nervous system.
The mesencephalon does not differentiate into any finer divisions. The midbrain is an established region of the brain at the primary vesicle stage of development and remains that way. The rest of the brain develops around it and constitutes a large percentage of the mass of the brain. Dividing the brain into forebrain, midbrain, and hindbrain is useful in considering its developmental pattern, but the midbrain is a small proportion of the entire brain, relatively speaking.
The rhombencephalon develops into the metencephalon and myelencephalon. The metencephalon corresponds to the adult structure known as the pons and gives rise to the cerebellum. The cerebellum (from the Latin meaning “little brain”) accounts for about 10 percent of the mass of the brain and is an important structure. The most significant connection between the cerebellum and the rest of the brain is at the pons because the pons and cerebellum develop out of the same vesicle. The myelencephalon corresponds to the adult structure known as the medulla oblongata. The structures that come from the mesencephalon and rhombencephalon, except for the cerebellum, are collectively considered the brain stem, which specifically includes the midbrain, pons, and medulla.
Figure 13.6.2. Primary and secondary vesicle stages of development. The embryonic brain develops complexity through enlargements of the neural tube called vesicles; (a) The primary vesicle stage has three regions, and (b) the secondary vesicle stage has five regions.
Spinal Cord Development
While the brain is developing from the anterior neural tube, the spinal cord is developing from the posterior neural tube. However, its structure does not differ from the basic layout of the neural tube. It is a long, straight cord with a small, hollow space down the centre. The neural tube is defined in terms of its anterior versus posterior portions, but it also has a dorsal–ventral dimension. As the neural tube separates from the rest of the ectoderm, the side closest to the surface is dorsal, and the deeper side is ventral.
As the spinal cord develops, the cells making up the wall of the neural tube proliferate and differentiate into the neurons and glia of the spinal cord. The dorsal tissues will be associated with sensory functions, and the ventral tissues will be associated with motor functions.
Relating Embryonic Development to the Adult Brain
Embryonic development can help in understanding the structure of the adult brain because it establishes a framework on which more complex structures can be built. First, the neural tube establishes the anterior–posterior dimension of the nervous system, which is called the neuraxis. The embryonic nervous system in mammals can be said to have a standard arrangement. Humans (and other primates, to some degree) make this complicated by standing up and walking on two legs. The anterior–posterior dimension of the neuraxis overlays the superior–inferior dimension of the body. However, there is a major curve between the brain stem and forebrain, which is called the cephalic flexure. Because of this, the neuraxis starts in an inferior position—the end of the spinal cord—and ends in an anterior position, the front of the cerebrum. If this is confusing, just imagine a four-legged animal standing up on two legs. Without the flexure in the brain stem, and at the top of the neck, that animal would be looking straight up instead of straight in front (Figure 13.6.3).
Figure 13.6.3. Human neuraxis. The mammalian nervous system is arranged with the neural tube running along an anterior to posterior axis, from nose to tail for a four-legged animal like a dog. Humans, as two-legged animals, have a bend in the neuraxis between the brain stem and the diencephalon, along with a bend in the neck, so that the eyes and the face are oriented forward.
In summary, the primary vesicles help to establish the basic regions of the nervous system: forebrain, midbrain, and hindbrain. These divisions are useful in certain situations, but they are not equivalent regions. The midbrain is small compared with the hindbrain and particularly the forebrain. The secondary vesicles go on to establish the major regions of the adult nervous system that will be followed in this text. The telencephalon is the cerebrum, which is the major portion of the human brain. The diencephalon continues to be referred to by this Greek name, because there is no better term for it (dia- = “through”). The diencephalon is between the cerebrum and the rest of the nervous system and can be described as the region through which all projections have to pass between the cerebrum and everything else. The brain stem includes the midbrain, pons, and medulla, which correspond to the mesencephalon, metencephalon, and myelencephalon. The cerebellum, being a large portion of the brain, is considered a separate region. Table 13.6.1 connects the different stages of development to the adult structures of the CNS.
One other benefit of considering embryonic development is that certain connections are more obvious because of how these adult structures are related. The retina, which began as part of the diencephalon, is primarily connected to the diencephalon. The eyes are just inferior to the anterior-most part of the cerebrum, but the optic nerve extends back to the thalamus as the optic tract, with branches into a region of the hypothalamus. There is also a connection of the optic tract to the midbrain, but the mesencephalon is adjacent to the diencephalon, so that is not difficult to imagine. The cerebellum originates out of the metencephalon, and its largest white matter connection is to the pons, also from the metencephalon. There are connections between the cerebellum and both the medulla and midbrain, which are adjacent structures in the secondary vesicle stage of development. In the adult brain, the cerebellum seems close to the cerebrum, but there is no direct connection between them.
Another aspect of the adult CNS structures that relates to embryonic development is the ventricles—open spaces within the CNS where cerebrospinal fluid circulates. They are the remnant of the hollow centre of the neural tube. The four ventricles and the tubular spaces associated with them can be linked back to the hollow centre of the embryonic brain (see Table 13.6.1).
Table 13.6.1. Stages of embryonic development
Neural tube
Primary vesicle stage
Secondary vesicle stage
Adult structures
Ventricles
Anterior neural tube
Prosencephalon
Telencephalon
Cerebrum
Lateral ventricles
Prosencephalon
Diencephalon
Diencephalon
Third ventricle
Mesencephalon
Mesencephalon
Midbrain
Cerebral aqueduct
Rhombencephalon
Metencephalon
Pons, cerebellum
Fourth ventricle
Rhombencephalon
Myelencephalon
Medulla
Fourth ventricle
Posterior neural tube
Spinal cord
Central canal
Disorders of the Nervous System
Early formation of the nervous system depends on the formation of the neural tube. A groove forms along the dorsal surface of the embryo, which becomes deeper until its edges meet and close off to form the tube. If this fails to happen, especially in the posterior region where the spinal cord forms, a developmental defect called spina bifida occurs. The closing of the neural tube is important for more than just the proper formation of the nervous system. The surrounding tissues are dependent on the correct development of the tube. The connective tissues surrounding the CNS can be involved as well.
There are three classes of this disorder: occulta, meningocele, and myelomeningocele (Figure 13.6.4). The first type, spina bifida occulta, is the mildest because the vertebral bones do not fully surround the spinal cord, but the spinal cord itself is not affected. No functional differences may be noticed, which is what the word occulta means; it is hidden spina bifida. The other two types both involve the formation of a cyst—a fluid-filled sac of the connective tissues that cover the spinal cord called the meninges. “Meningocele” means that the meninges protrude through the spinal column, but nerves may not be involved, and few symptoms are present, though complications may arise later in life. “Myelomeningocele” means that the meninges protrude, and spinal nerves are involved, and therefore severe neurological symptoms can be present.
Often surgery to close the opening or to remove the cyst is necessary. The earlier that surgery can be performed, the better the chances of controlling or limiting further damage or infection at the opening. For many children with meningocele, surgery will alleviate the pain, although they may experience some functional loss. Because the myelomeningocele form of spina bifida involves more extensive damage to the nervous tissue, neurological damage may persist, but symptoms can often be handled. Complications of the spinal cord may present later in life, but overall life expectancy is not reduced.
Figure 13.6.4. Spinal bifida. (a) Spina bifida is a birth defect of the spinal cord caused when the neural tube does not completely close, but the rest of development continues. The result is the emergence of meninges and neural tissue through the vertebral column. (b) Foetal myelomeningocele is evident in this ultrasound taken at 21 weeks.
Section Review
The development of the nervous system starts early in embryonic development. The outer layer of the embryo, the ectoderm, gives rise to the skin and the nervous system. A specialised region of this layer, the neuroectoderm, becomes a groove that folds in and becomes the neural tube beneath the dorsal surface of the embryo. The anterior end of the neural tube develops into the brain, and the posterior region becomes the spinal cord. Tissues at the edges of the neural groove, when it closes off, are called the neural crest and migrate through the embryo to give rise to PNS structures as well as some non-nervous tissues.
The brain develops from this early tube structure and gives rise to specific regions of the adult brain. As the neural tube grows and differentiates, it enlarges into three vesicles that correspond to the forebrain, midbrain, and hindbrain regions of the adult brain. Later in development, two of these three vesicles differentiate further, resulting in five vesicles. Those five vesicles can be aligned with the four major regions of the adult brain. The cerebrum is formed directly from the telencephalon. The diencephalon is the only region that keeps its embryonic name. The mesencephalon, metencephalon, and myelencephalon become the brain stem. The cerebellum also develops from the metencephalon and is a separate region of the adult brain.
The spinal cord develops out of the rest of the neural tube and retains the tube structure, with the nervous tissue thickening and the hollow centre becoming a very small central canal through the cord. The rest of the hollow centre of the neural tube corresponds to open spaces within the brain called the ventricles, where cerebrospinal fluid is found.
Describe the connections between the cerebrum and brain stem through the diencephalon, and from those regions into the spinal cord
Recognise the complex connections within the subcortical structures of the basal nuclei
Explain the arrangement of grey and white matter in the spinal cord
The brain and the spinal cord are the central nervous system, and they represent the main organs of the nervous system. The spinal cord is a single structure, whereas the adult brain is described in terms of four major regions: the cerebrum, the diencephalon, the brain stem, and the cerebellum. A person’s conscious experiences are based on neural activity in the brain. The regulation of homeostasis is governed by a specialised region in the brain. The coordination of reflexes depends on the integration of sensory and motor pathways in the spinal cord.
The Cerebrum
The iconic grey mantle of the human brain, which appears to make up most of the mass of the brain, is the cerebrum (Figure 13.7.1). The wrinkled portion is the cerebral cortex, and the rest of the structure is beneath that outer covering. There is a large separation between the two sides of the cerebrum called the longitudinal fissure. It separates the cerebrum into two distinct halves, a right and left cerebral hemisphere. Deep within the cerebrum, the white matter of the corpus callosum provides the major pathway for communication between the two hemispheres of the cerebral cortex.
Figure 13.7.1. The cerebrum. The cerebrum is a large component of the CNS in humans, and the most obvious aspect of it is the folded surface called the cerebral cortex.
Many of the higher neurological functions, such as memory, emotion, and consciousness, are the result of cerebral function. The complexity of the cerebrum is different across vertebrate species. The cerebrum of the most primitive vertebrates is not much more than the connection for the sense of smell. In mammals, the cerebrum comprises the outer grey matter that is the cortex (from the Latin word meaning “bark of a tree”) and several deep nuclei that belong to three important functional groups. The basal nuclei are responsible for cognitive processing, the most important function being that associated with planning movements. The basal forebrain contains nuclei that are important in learning and memory. The limbic cortex is the region of the cerebral cortex that is part of the limbic system, a collection of structures involved in emotion, memory, and behaviour.
Cerebral Cortex
The cerebrum is covered by a continuous layer of grey matter that wraps around either side of the forebrain—the cerebral cortex. This thin, extensive region of wrinkled grey matter is responsible for the higher functions of the nervous system. A gyrus (plural = gyri) is the ridge of one of those wrinkles, and a sulcus (plural = sulci) is the groove between two gyri. The pattern of these folds of tissue indicates specific regions of the cerebral cortex.
The head is limited by the size of the birth canal, and the brain must fit inside the cranial cavity of the skull. Extensive folding in the cerebral cortex enables more grey matter to fit into this limited space. If the grey matter of the cortex were peeled off of the cerebrum and laid out flat, its surface area would be roughly equal to one square meter.
The folding of the cortex maximises the amount of grey matter in the cranial cavity. During embryonic development, as the telencephalon expands within the skull, the brain goes through a regular course of growth that results in everyone’s brain having a similar pattern of folds. The surface of the brain can be mapped on the basis of the locations of large gyri and sulci. Using these landmarks, the cortex can be separated into four major regions, or lobes (Figure 13.7.2). The lateral sulcus that separates the temporal lobe from the other regions is one such landmark. Superior to the lateral sulcus are the parietal lobe and frontal lobe, which are separated from each other by the central sulcus. The posterior region of the cortex is the occipital lobe, which has no obvious anatomical border between it and the parietal or temporal lobes on the lateral surface of the brain. From the medial surface, an obvious landmark separating the parietal and occipital lobes is called the parieto-occipital sulcus. The fact that there is no obvious anatomical border between these lobes is consistent with the functions of these regions being interrelated.
Figure 13.7.2. Lobes of the cerebral cortex. The cerebral cortex is divided into four lobes. Extensive folding increases the surface area available for cerebral functions.
Different regions of the cerebral cortex can be associated with particular functions, a concept known as localisation of function. In the early 1900s, a German neuroscientist named Korbinian Brodmann performed an extensive study of the microscopic anatomy—the cytoarchitecture—of the cerebral cortex and divided the cortex into 52 separate regions on the basis of the histology of the cortex. His work resulted in a system of classification known as Brodmann’s areas, which is still used today to describe the anatomical distinctions within the cortex (Figure 13.7.3). The results from Brodmann’s work on the anatomy align very well with the functional differences within the cortex. Areas 17 and 18 in the occipital lobe are responsible for primary visual perception. That visual information is complex, so it is processed in the temporal and parietal lobes as well.
The temporal lobe is associated with primary auditory sensation, known as Brodmann’s areas 41 and 42 in the superior temporal lobe. Because regions of the temporal lobe are part of the limbic system, memory is an important function associated with that lobe. Memory is essentially a sensory function; memories are recalled sensations such as the smell of Mom’s baking or the sound of a barking dog. Even memories of movement are really the memory of sensory feedback from those movements, such as stretching muscles or the movement of the skin around a joint. Structures in the temporal lobe are responsible for establishing long-term memory, but the ultimate location of those memories is usually in the region in which the sensory perception was processed.
The main sensation associated with the parietal lobe is somatosensation, meaning the general sensations associated with the body. Posterior to the central sulcus is the postcentral gyrus, the primary somatosensory cortex, which is identified as Brodmann’s areas 1, 2, and 3. All the tactile senses are processed in this area, including touch, pressure, tickle, pain, itch, and vibration, as well as more general senses of the body such as proprioception and kinaesthesia, which are the senses of body position and movement, respectively.
Anterior to the central sulcus is the frontal lobe, which is primarily associated with motor functions. The precentral gyrus is the primary motor cortex. Cells from this region of the cerebral cortex are the upper motor neurons that instruct cells in the spinal cord to move skeletal muscles. Anterior to this region are a few areas that are associated with planned movements. The premotor area is responsible for thinking of a movement to be made. The frontal eye fields are important in eliciting eye movements and in attending to visual stimuli. Broca’s area is responsible for the production of language or controlling movements responsible for speech; in most people, it is located only on the left side. Anterior to these regions is the prefrontal lobe, which serves cognitive functions that can be the basis of personality, short-term memory, and consciousness. The prefrontal lobotomy is an outdated mode of treatment for personality disorders (psychiatric conditions) that profoundly affected the personality of the patient.
Figure 13.7.3. Brodmann’s areas of the cerebral cortex. Brodmann mapping of functionally distinct regions of the cortex was based on its cytoarchitecture at a microscopic level.
Subcortical Structures
Beneath the cerebral cortex are sets of nuclei known as subcortical nuclei that augment cortical processes. The nuclei of the basal forebrain serve as the primary location for acetylcholine production, which modulates the overall activity of the cortex, leading to greater attention to sensory stimuli. Alzheimer’s disease is associated with a loss of neurons in the basal forebrain. The hippocampus and amygdala are medial-lobe structures that, along with the adjacent cortex, are involved in long-term memory formation and emotional responses. The basal nuclei are a set of nuclei in the cerebrum responsible for comparing cortical processing with the general state of activity in the nervous system to influence the likelihood of movement taking place, for example, while a student is sitting in a classroom listening to a lecture, the basal nuclei will keep the urge to jump up and scream from happening. (The basal nuclei are also referred to as the basal ganglia, although that is potentially confusing because the term ganglia is typically used for peripheral structures).
The major structures of the basal nuclei that control movement are the caudate, putamen, and globus pallidus, which are located deep in the cerebrum. The caudate is a long nucleus that follows the basic C-shape of the cerebrum from the frontal lobe, through the parietal and occipital lobes, into the temporal lobe. The putamen is mostly deep in the anterior regions of the frontal and parietal lobes. Together, the caudate and putamen are called the striatum. The globus pallidus is a layered nucleus that lies just medial to the putamen; they are called the lenticular nuclei because they look like curved pieces fitting together like lenses. The globus pallidus has two subdivisions, the external and internal segments, which are lateral and medial, respectively. These nuclei are depicted in a frontal section of the brain in Figure 13.7.4.
Figure 13.7.4. Frontal section of cerebral cortex and basal nuclei. The major components of the basal nuclei, shown in a frontal section of the brain, are the caudate (just lateral to the lateral ventricle), the putamen (inferior to the caudate and separated by the large white-matter structure called the internal capsule), and the globus pallidus (medial to the putamen).
The basal nuclei in the cerebrum are connected with a few more nuclei in the brain stem that together act as a functional group that forms a motor pathway. Two streams of information processing take place in the basal nuclei. All input to the basal nuclei is from the cortex into the striatum (Figure 13.7.5). The direct pathway is the projection of axons from the striatum to the globus pallidus internal segment (GPi) and the substantia nigra pars reticulata (SNr). The GPi/SNr then projects to the thalamus, which projects back to the cortex. The indirect pathway is the projection of axons from the striatum to the globus pallidus external segment (GPe), then to the subthalamic nucleus (STN), and finally to GPi/SNr. The two streams both target the GPi/SNr, but one has a direct projection and the other goes through a few intervening nuclei. The direct pathway causes the disinhibition of the thalamus (inhibition of one cell on a target cell that then inhibits the first cell), whereas the indirect pathway causes, or reinforces, the normal inhibition of the thalamus. The thalamus then can either excite the cortex (because of the direct pathway) or fail to excite the cortex (because of the indirect pathway).
Figure 13.7.5. Connections of basal nuclei. Input to the basal nuclei is from the cerebral cortex, which is an excitatory connection releasing glutamate as a neurotransmitter. This input is to the striatum, or the caudate and putamen. In the direct pathway, the striatum projects to the internal segment of the globus pallidus and the substantia nigra pars reticulata (GPi/SNr). This is an inhibitory pathway, in which GABA is released at the synapse, and the target cells are hyperpolarised and less likely to fire. The output from the basal nuclei is to the thalamus, which is an inhibitory projection using GABA.
The switch between the two pathways is the substantia nigra pars compacta, which projects to the striatum and releases the neurotransmitter dopamine. Dopamine receptors are either excitatory (D1-type receptors) or inhibitory (D2-type receptors). The direct pathway is activated by dopamine, and the indirect pathway is inhibited by dopamine. When the substantia nigra pars compacta is firing, it signals to the basal nuclei that the body is in an active state, and movement will be more likely. When the substantia nigra pars compacta is silent, the body is in a passive state, and movement is inhibited. To illustrate this situation, while a student is sitting listening to a lecture, the substantia nigra pars compacta would be silent and the student less likely to get up and walk around. Likewise, while the professor is lecturing, and walking around at the front of the classroom, the professor’s substantia nigra pars compacta would be active, in keeping with his or her activity level.
Everyday Connections
The Myth of Left Brain/Right Brain
There is a persistent myth that people are “right-brained” or “left-brained,” which is an oversimplification of an important concept about the cerebral hemispheres. There is some lateralisation of function, in which the left side of the brain is devoted to language function and the right side is devoted to spatial and nonverbal reasoning. Whereas these functions are associated with those sides of the brain, there is no monopoly by either side on these functions. Many pervasive functions, such as language, are distributed globally around the cerebrum.
Some of the support for this misconception has come from studies of split brains. A drastic way to deal with a rare and devastating neurological condition (intractable epilepsy) is to separate the two hemispheres of the brain. After sectioning the corpus callosum, a split-brained patient will have trouble producing verbal responses based on sensory information processed on the right side of the cerebrum, leading to the idea that the left side is responsible for language function.
However, there are well-documented cases of language functions lost from damage to the right side of the brain. The deficits seen in damage to the left side of the brain are classified as aphasia, a loss of speech function; damage on the right side can affect the use of language. Right-side damage can result in a loss of ability to understand figurative aspects of speech, such as jokes, irony, or metaphors. Nonverbal aspects of speech can be affected by damage to the right side, such as facial expression or body language, and right-side damage can lead to a “flat affect” in speech, or a loss of emotional expression in speech—sounding like a robot when talking.
The Diencephalon
The diencephalon is the one region of the adult brain that retains its name from embryologic development. The etymology of the word diencephalon translates to “through brain.” It is the connection between the cerebrum and the rest of the nervous system, with one exception. The rest of the brain, the spinal cord, and the PNS all send information to the cerebrum through the diencephalon. Output from the cerebrum passes through the diencephalon. The single exception is the system associated with olfaction, or the sense of smell, which connects directly with the cerebrum. In the earliest vertebrate species, the cerebrum was not much more than olfactory bulbs that received peripheral information about the chemical environment (to call it smell in these organisms is imprecise because they lived in the ocean).
The diencephalon is deep beneath the cerebrum and constitutes the walls of the third ventricle. The diencephalon can be described as any region of the brain with “thalamus” in its name. The two major regions of the diencephalon are the thalamus itself and the hypothalamus (Figure 13.7.6). There are other structures, such as the epithalamus, which contains the pineal gland, or the subthalamus, which includes the subthalamic nucleus that is part of the basal nuclei.
Thalamus
The thalamus is a collection of nuclei that relay information between the cerebral cortex and the periphery, spinal cord, or brain stem. All sensory information, except for the sense of smell, passes through the thalamus before processing by the cortex. Axons from the peripheral sensory organs, or intermediate nuclei, synapse in the thalamus, and thalamic neurons project directly to the cerebrum. It is a requisite synapse in any sensory pathway, except for olfaction. The thalamus does not just pass the information on, it also processes that information, for example, the portion of the thalamus that receives visual information will influence what visual stimuli are important, or what receives attention.
The cerebrum also sends information down to the thalamus, which usually communicates motor commands. This involves interactions with the cerebellum and other nuclei in the brain stem. The cerebrum interacts with the basal nuclei, which involves connections with the thalamus. The primary output of the basal nuclei is to the thalamus, which relays that output to the cerebral cortex. The cortex also sends information to the thalamus that will then influence the effects of the basal nuclei.
Hypothalamus
Inferior and slightly anterior to the thalamus is the hypothalamus, the other major region of the diencephalon. The hypothalamus is a collection of nuclei that are involved in regulating homeostasis. The hypothalamus is the executive region in charge of the autonomic nervous system and the endocrine system through its regulation of the anterior pituitary gland. Other parts of the hypothalamus are involved in memory and emotion as part of the limbic system.
Figure 13.7.6. The diencephalon. The diencephalon is composed primarily of the thalamus and hypothalamus, which together define the walls of the third ventricle. The thalami are two elongated, ovoid structures on either side of the midline that make contact in the middle. The hypothalamus is inferior and anterior to the thalamus, culminating in a sharp angle to which the pituitary gland is attached.
Brain Stem
The midbrain and hindbrain (composed of the pons and the medulla) are collectively referred to as the brain stem (Figure 13.7.7). The structure emerges from the ventral surface of the forebrain as a tapering cone that connects the brain to the spinal cord. Attached to the brain stem, but considered a separate region of the adult brain, is the cerebellum. The midbrain coordinates sensory representations of the visual, auditory, and somatosensory perceptual spaces. The pons is the main connection with the cerebellum. The pons and the medulla regulate several crucial functions, including the cardiovascular and respiratory systems and rates.
The cranial nerves connect through the brain stem and provide the brain with the sensory input and motor output associated with the head and neck, including most of the special senses. The major ascending and descending pathways between the spinal cord and brain, specifically the cerebrum, pass through the brain stem.
Figure 13.7.7. The brain stem. The brain stem comprises three regions: the midbrain, the pons, and the medulla.
Midbrain
One of the original regions of the embryonic brain, the midbrain is a small region between the thalamus and pons. It is separated into the tectum and tegmentum, from the Latin words for roof and floor, respectively. The cerebral aqueduct passes through the centre of the midbrain, such that these regions are the roof and floor of that canal.
The tectum is composed of four bumps known as the colliculi (singular = colliculus), which means “little hill” in Latin. The inferior colliculus is the inferior pair of these enlargements and is part of the auditory brain stem pathway. Neurons of the inferior colliculus project to the thalamus, which then sends auditory information to the cerebrum for the conscious perception of sound. The superior colliculus is the superior pair and combines sensory information about visual space, auditory space, and somatosensory space. Activity in the superior colliculus is related to orienting the eyes to a sound or touch stimulus. If you are walking along the sidewalk on campus and you hear chirping, the superior colliculus coordinates that information with your awareness of the visual location of the tree right above you. That is the correlation of auditory and visual maps. If you suddenly feel something wet fall on your head, your superior colliculus integrates that with the auditory and visual maps and you know that the chirping bird just relieved itself on you. You want to look up to see the culprit, but do not.
The tegmentum is continuous with the grey matter of the rest of the brain stem. Throughout the midbrain, pons, and medulla, the tegmentum contains the nuclei that receive and send information through the cranial nerves, as well as regions that regulate important functions such as those of the cardiovascular and respiratory systems.
Pons
The word pons comes from the Latin word for bridge. It is visible on the anterior surface of the brain stem as the thick bundle of white matter attached to the cerebellum. The pons is the main connection between the cerebellum and the brain stem. The bridge-like white matter is only the anterior surface of the pons; the grey matter beneath that is a continuation of the tegmentum from the midbrain. Grey matter in the tegmentum region of the pons contains neurons receiving descending input from the forebrain that is sent to the cerebellum.
Medulla
The medulla is the region known as the myelencephalon in the embryonic brain. The initial portion of the name, “myel,” refers to the significant white matter found in this region—especially on its exterior, which is continuous with the white matter of the spinal cord. The tegmentum of the midbrain and pons continues into the medulla because this grey matter is responsible for processing cranial nerve information. A diffuse region of grey matter throughout the brain stem, known as the reticular formation, is related to sleep and wakefulness, such as general brain activity and attention.
The Cerebellum
The cerebellum, as the name suggests, is the “little brain.” It is covered in gyri and sulci like the cerebrum and looks like a miniature version of that part of the brain (Figure 13.7.8). The cerebellum is responsible for comparing information from the cerebrum with sensory feedback from the periphery through the spinal cord. It accounts for approximately 10 percent of the mass of the brain.
Figure 13.7.8. The cerebellum. The cerebellum is situated on the posterior surface of the brain stem. Descending input from the cerebellum enters through the large white matter structure of the pons. Ascending input from the periphery and spinal cord enters through the fibres of the inferior olive. Output goes to the midbrain, which sends a descending signal to the spinal cord.
Descending fibres from the cerebrum have branches that connect to neurons in the pons. Those neurons project into the cerebellum, providing a copy of motor commands sent to the spinal cord. Sensory information from the periphery, which enters through spinal or cranial nerves, is copied to a nucleus in the medulla known as the inferior olive. Fibres from this nucleus enter the cerebellum and are compared with the descending commands from the cerebrum. If the primary motor cortex of the frontal lobe sends a command down to the spinal cord to initiate walking, a copy of that instruction is sent to the cerebellum. Sensory feedback from the muscles and joints, proprioceptive information about the movements of walking, and sensations of balance are sent to the cerebellum through the inferior olive and the cerebellum compares them. If walking is not coordinated, perhaps because the ground is uneven or a strong wind is blowing, then the cerebellum sends out a corrective command to compensate for the difference between the original cortical command and the sensory feedback. The output of the cerebellum is into the midbrain, which then sends a descending input to the spinal cord to correct the messages going to skeletal muscles.
The Spinal Cord
The description of the CNS is concentrated on the structures of the brain, but the spinal cord is another major organ of the system. Whereas the brain develops out of expansions of the neural tube into primary and then secondary vesicles, the spinal cord maintains the tube structure and is only specialised into certain regions. As the spinal cord continues to develop in the newborn, anatomical features mark its surface. The anterior midline is marked by the anterior median fissure, and the posterior midline is marked by the posterior median sulcus. Axons enter the posterior side through the dorsal (posterior) nerve root, which marks the posterolateral sulcus on either side. The axons emerging from the anterior side do so through the ventral (anterior) nerve root. Note that it is common to see the terms dorsal (dorsal = “back”) and ventral (ventral = “belly”) used interchangeably with posterior and anterior, particularly in reference to nerves and the structures of the spinal cord. You should learn to be comfortable with both.
Overall, the posterior regions are responsible for sensory functions and the anterior regions are associated with motor functions. This comes from the initial development of the spinal cord, which is divided into the basal plate and the alar plate. The basal plate is closest to the ventral midline of the neural tube, which will become the anterior face of the spinal cord and gives rise to motor neurons. The alar plate is on the dorsal side of the neural tube and gives rise to neurons that will receive sensory input from the periphery.
The length of the spinal cord is divided into regions that correspond to the regions of the vertebral column. The name of a spinal cord region corresponds to the level at which spinal nerves pass through the intervertebral foramina. Immediately adjacent to the brain stem is the cervical region, followed by the thoracic, then the lumbar, and finally the sacral region. The spinal cord is not the full length of the vertebral column because the spinal cord does not grow significantly longer after the first or second year, but the skeleton continues to grow. The nerves that emerge from the spinal cord pass through the intervertebral formina at the respective levels. As the vertebral column grows, these nerves grow with it and result in a long bundle of nerves that resembles a horse’s tail and is named the cauda equina. The sacral spinal cord is at the level of the upper lumbar vertebral bones. The spinal nerves extend from their various levels to the proper level of the vertebral column.
Grey Horns
In cross-section, the grey matter of the spinal cord has the appearance of an ink-blot test, with the spread of the grey matter on one side replicated on the other—a shape reminiscent of a bulbous capital “H.” As shown in Figure 13.7.9, the grey matter is subdivided into regions that are referred to as horns. The posterior horn is responsible for sensory processing. The anterior horn sends out motor signals to the skeletal muscles. The lateral horn, which is only found in the thoracic, upper lumbar, and sacral regions, is the central component of the sympathetic division of the autonomic nervous system.
Some of the largest neurons of the spinal cord are the multipolar motor neurons in the anterior horn. The fibres that cause contraction of skeletal muscles are the axons of these neurons. The motor neuron that causes contraction of the big toe, for example, is located in the sacral spinal cord. The axon that must reach all the way to the belly of that muscle may be a meter in length. The neuronal cell body that maintains that long fibre must be quite large, possibly several hundred micrometres in diameter, making it one of the largest cells in the body.
Just as the grey matter is separated into horns, the white matter of the spinal cord is separated into columns. Ascending tracts of nervous system fibres in these columns carry sensory information up to the brain, whereas descending tracts carry motor commands from the brain. Looking at the spinal cord longitudinally, the columns extend along its length as continuous bands of white matter. Between the two posterior horns of grey matter are the posterior columns. Between the two anterior horns, and bounded by the axons of motor neurons emerging from that grey matter area, are the anterior columns. The white matter on either side of the spinal cord, between the posterior horn and the axons of the anterior horn neurons, are the lateral columns. The posterior columns are composed of axons of ascending tracts. The anterior and lateral columns are composed of many different groups of axons of both ascending and descending tracts—the latter carrying motor commands down from the brain to the spinal cord to control output to the periphery.
Disorders of the Basal Nuclei
Parkinson’s disease is a disorder of the basal nuclei, specifically of the substantia nigra, that demonstrates the effects of the direct and indirect pathways. Parkinson’s disease is the result of neurons in the substantia nigra pars compacta dying. These neurons release dopamine into the striatum. Without that modulatory influence, the basal nuclei are stuck in the indirect pathway, without the direct pathway being activated. The direct pathway is responsible for increasing cortical movement commands. The increased activity of the indirect pathway results in the hypokinetic disorder of Parkinson’s disease.
Parkinson’s disease is neurodegenerative, meaning that neurons die that cannot be replaced, so there is no cure for the disorder. Treatments for Parkinson’s disease are aimed at increasing dopamine levels in the striatum. Currently, the most common way of doing that is by providing the amino acid L-DOPA, which is a precursor to the neurotransmitter dopamine and can cross the blood-brain barrier. With levels of the precursor elevated, the remaining cells of the substantia nigra pars compacta can make more neurotransmitter and have a greater effect. Unfortunately, the patient will become less responsive to L-DOPA treatment as time progresses, and it can cause increased dopamine levels elsewhere in the brain, which are associated with psychosis or schizophrenia.
Section Review
The adult brain is separated into four major regions: the cerebrum, the diencephalon, the brain stem, and the cerebellum. The cerebrum is the largest portion and contains the cerebral cortex and subcortical nuclei. It is divided into two halves by the longitudinal fissure.
The cortex is separated into the frontal, parietal, temporal, and occipital lobes. The frontal lobe is responsible for motor functions, from planning movements through executing commands to be sent to the spinal cord and periphery. The most anterior portion of the frontal lobe is the prefrontal cortex, which is associated with aspects of personality through its influence on motor responses in decision-making.
The other lobes are responsible for sensory functions. The parietal lobe is where somatosensation is processed. The occipital lobe is where visual processing begins, although the other parts of the brain can contribute to visual function. The temporal lobe contains the cortical area for auditory processing, but also has regions crucial for memory formation.
Nuclei beneath the cerebral cortex, known as the subcortical nuclei, are responsible for augmenting cortical functions. The basal nuclei receive input from cortical areas and compare it with the general state of the individual through the activity of a dopamine-releasing nucleus. The output influences the activity of part of the thalamus that can then increase or decrease cortical activity that often results in changes to motor commands. The basal forebrain is responsible for modulating cortical activity in attention and memory. The limbic system includes deep cerebral nuclei that are responsible for emotion and memory.
The diencephalon includes the thalamus and the hypothalamus, along with some other structures. The thalamus is a relay between the cerebrum and the rest of the nervous system. The hypothalamus coordinates homeostatic functions through the autonomic and endocrine systems.
The brain stem is composed of the midbrain, pons, and medulla. It controls the head and neck region of the body through the cranial nerves. There are control centres in the brain stem that regulate the cardiovascular and respiratory systems.
The cerebellum is connected to the brain stem, primarily at the pons, where it receives a copy of the descending input from the cerebrum to the spinal cord. It can compare this with sensory feedback input through the medulla and send output through the midbrain that can correct motor commands for coordination.
Describe the vessels that supply the CNS with blood
Name the components of the ventricular system and the regions of the brain in which each is located
Explain the production of cerebrospinal fluid and its flow through the ventricles
Explain how a disruption in circulation would result in a stroke
The CNS is crucial to the operation of the body, and any compromise in the brain and spinal cord can lead to severe difficulties. The CNS has a privileged blood supply, as suggested by the blood-brain barrier. The function of the tissue in the CNS is crucial to the survival of the organism, so the contents of the blood cannot simply pass into the central nervous tissue. To protect this region from the toxins and pathogens that may be traveling through the blood stream, there is strict control over what can move out of the general systems and into the brain and spinal cord. Because of this privilege, the CNS needs specialised structures for the maintenance of circulation. This begins with a unique arrangement of blood vessels carrying fresh blood into the CNS. Beyond the supply of blood, the CNS filters that blood into cerebrospinal fluid (CSF), which is then circulated through the cavities of the brain and spinal cord called ventricles.
Blood Supply to the Brain
A lack of oxygen to the CNS can be devastating, and the cardiovascular system has specific regulatory reflexes to ensure that the blood supply is not interrupted. There are multiple routes for blood to get into the CNS, with specialisations to protect that blood supply and to maximise the ability of the brain to get an uninterrupted perfusion.
Arterial Supply
The major artery carrying recently oxygenated blood away from the heart is the aorta. The very first branches off the aorta supply the heart with nutrients and oxygen. The next branches give rise to the common carotid arteries, which further branch into the internal carotid arteries. The external carotid arteries supply blood to the tissues on the surface of the cranium. The bases of the common carotids contain stretch receptors that immediately respond to the drop in blood pressure upon standing. The orthostatic reflex is a reaction to this change in body position, so that blood pressure is maintained against the increasing effect of gravity (orthostatic means “standing up”). Heart rate increases—a reflex of the sympathetic division of the autonomic nervous system—and this raises blood pressure.
The internal carotid artery enters the cranium through the carotid canal in the temporal bone. A second set of vessels that supply the CNS are the vertebral arteries, which are protected as they pass through the neck region by the transverse foramina of the cervical vertebrae. The vertebral arteries enter the cranium through the foramen magnum of the occipital bone. Branches off the left and right vertebral arteries merge into the anterior spinal artery supplying the anterior aspect of the spinal cord, found along the anterior median fissure. The two vertebral arteries then merge into the basilar artery, which gives rise to branches to the brain stem and cerebellum. The left and right internal carotid arteries and branches of the basilar artery all become the circle of Willis, a confluence of arteries that can maintain perfusion of the brain even if narrowing or a blockage limits flow through one part (Figure 13.8.1).
Figure 13.8.1. Circle of Willis. The blood supply to the brain enters through the internal carotid arteries and the vertebral arteries, eventually giving rise to the circle of Willis.
Venous Return
After passing through the CNS, blood returns to the circulation through a series of dural sinuses and veins (Figure 13.8.2). The superior sagittal sinus runs in the groove of the longitudinal fissure, where it absorbs CSF from the meninges. The superior sagittal sinus drains to the confluence of sinuses, along with the occipital sinuses and straight sinus, to then drain into the transverse sinuses. The transverse sinuses connect to the sigmoid sinuses, which then connect to the jugular veins. From there, the blood continues toward the heart to be pumped to the lungs for reoxygenation.
Figure 13.8.2. Dural sinuses and veins. Blood drains from the brain through a series of sinuses that connect to the jugular veins.
Protective Coverings of the Brain and Spinal Cord
The outer surface of the CNS is covered by a series of membranes composed of connective tissue called the meninges, which protect the brain. The dura mater is a thick fibrous layer and a strong protective sheath over the entire brain and spinal cord. It is anchored to the inner surface of the cranium and vertebral cavity. The arachnoid mater is a membrane of thin fibrous tissue that forms a loose sac around the CNS. Beneath the arachnoid is a thin, filamentous mesh called the arachnoid trabeculae, which looks like a spider web, giving this layer its name. Directly adjacent to the surface of the CNS is the pia mater, a thin fibrous membrane that follows the convolutions of gyri and sulci in the cerebral cortex and fits into other grooves and indentations (Figure 13.8.3).
Figure 13.8.3. Meningeal layers of superior sagittal sinus. The layers of the meninges in the longitudinal fissure of the superior sagittal sinus are shown, with the dura mater adjacent to the inner surface of the cranium, the pia mater adjacent to the surface of the brain, and the arachnoid and subarachnoid space between them. An arachnoid villus is shown emerging into the dural sinus to allow CSF to filter back into the blood for drainage.
Dura Mater
Like a thick cap covering the brain, the dura mater is a tough outer covering. The name comes from the Latin for “tough mother” to represent its physically protective role. It encloses the entire CNS and the major blood vessels that enter the cranium and vertebral cavity. It is directly attached to the inner surface of the bones of the cranium and to the very end of the vertebral cavity.
There are infoldings of the dura that fit into large crevasses of the brain. Two infoldings go through the midline separations of the cerebrum and cerebellum; one forms a shelf-like tent between the occipital lobes of the cerebrum and the cerebellum, and the other surrounds the pituitary gland. The dura also surrounds and supports the venous sinuses.
Arachnoid Mater
The middle layer of the meninges is the arachnoid, named for the spider-web–like trabeculae between it and the pia mater. The arachnoid defines a sac-like enclosure around the CNS. The trabeculae are found in the subarachnoid space, which is filled with circulating CSF. The arachnoid emerges into the dural sinuses as the arachnoid granulations, where the CSF is filtered back into the blood for drainage from the nervous system.
The subarachnoid space is filled with circulating CSF, which also provides a liquid cushion to the brain and spinal cord. Similar to clinical blood work, a sample of CSF can be withdrawn to find chemical evidence of neuropathology or metabolic traces of the biochemical functions of nervous tissue.
Pia Mater
The outer surface of the CNS is covered in the thin fibrous membrane of the pia mater. It is thought to have a continuous layer of cells providing a fluid-impermeable membrane. The name pia mater comes from the Latin for “tender mother,” suggesting the thin membrane is a gentle covering for the brain. The pia extends into every convolution of the CNS, lining the inside of the sulci in the cerebral and cerebellar cortices. At the end of the spinal cord, a thin filament extends from the inferior end of CNS at the upper lumbar region of the vertebral column to the sacral end of the vertebral column. Because the spinal cord does not extend through the lower lumbar region of the vertebral column, a needle can be inserted through the dura and arachnoid layers to withdraw CSF. This procedure is called a lumbar puncture and avoids the risk of damaging the central tissue of the spinal cord. Blood vessels that are nourishing the central nervous tissue are between the pia mater and the nervous tissue.
Disorders of the Meninges
Meningitis is an inflammation of the meninges, the three layers of fibrous membrane that surround the CNS. Meningitis can be caused by infection by bacteria or viruses. The pathogens are not special to meningitis; it is just an inflammation of that specific set of tissues from what might be a broader infection. Bacterial meningitis can be caused by Streptococcus, Staphylococcus, or the tuberculosis pathogen, among many others. Viral meningitis is usually the result of common enteroviruses (such as those that cause intestinal disorders) but may be the result of the herpes virus or West Nile virus. Bacterial meningitis tends to be more severe.
The symptoms associated with meningitis can be fever, chills, nausea, vomiting, light sensitivity, soreness of the neck, or severe headache. More important are the neurological symptoms, such as changes in mental state (confusion, memory deficits, and other dementia-type symptoms). A serious risk of meningitis can be damage to peripheral structures because of the nerves that pass through the meninges. Hearing loss is a common result of meningitis.
The primary test for meningitis is a lumbar puncture. A needle inserted into the lumbar region of the spinal column through the dura mater and arachnoid membrane into the subarachnoid space can be used to withdraw the fluid for chemical testing. Fatality occurs in 5 to 40 percent of children and 20 to 50 percent of adults with bacterial meningitis. Treatment of bacterial meningitis is through antibiotics, but viral meningitis cannot be treated with antibiotics because viruses do not respond to that type of drug. Fortunately, the viral forms are milder.
The Ventricular System
Cerebrospinal fluid (CSF) circulates throughout and around the CNS. In other tissues, water and small molecules are filtered through capillaries as the major contributor to the interstitial fluid. In the brain, CSF is produced in special structures to perfuse through the nervous tissue of the CNS and is continuous with the interstitial fluid. Specifically, CSF circulates to remove metabolic wastes from the interstitial fluids of nervous tissues and return them to the blood stream. The ventricles are the open spaces within the brain where CSF circulates. In some of these spaces, CSF is produced by filtering of the blood that is performed by a specialised membrane known as a choroid plexus. The CSF circulates through all of the ventricles to eventually emerge into the subarachnoid space where it will be reabsorbed into the blood.
The Ventricles
There are four ventricles within the brain, all of which developed from the original hollow space within the neural tube, the central canal. The first two are named the lateral ventricles and are deep within the cerebrum. These ventricles are connected to the third ventricle by two openings called the interventricular foramina. The third ventricle is the space between the left and right sides of the diencephalon, which opens into the cerebral aqueduct that passes through the midbrain. The aqueduct opens into the fourth ventricle, which is the space between the cerebellum and the pons and upper medulla (Figure 13.8.4).
Figure 13.8.4. Cerebrospinal fluid circulation. The choroid plexus in the four ventricles produce CSF, which is circulated through the ventricular system and then enters the subarachnoid space through the median and lateral apertures. The CSF is then reabsorbed into the blood at the arachnoid granulations, where the arachnoid membrane emerges into the dural sinuses.
As the telencephalon enlarges and grows into the cranial cavity, it is limited by the space within the skull. The telencephalon is the most anterior region of what was the neural tube but cannot grow past the limit of the frontal bone of the skull. Because the cerebrum fits into this space, it takes on a C-shaped formation, through the frontal, parietal, occipital, and finally temporal regions. The space within the telencephalon is stretched into this same C-shape. The two ventricles are in the left and right sides and were at one time referred to as the first and second ventricles. The interventricular foramina connect the frontal region of the lateral ventricles with the third ventricle.
The third ventricle is the space bounded by the medial walls of the hypothalamus and thalamus. The two thalami touch in the centre in most brains as the massa intermedia, which is surrounded by the third ventricle. The cerebral aqueduct opens just inferior to the epithalamus and passes through the midbrain. The tectum and tegmentum of the midbrain are the roof and floor of the cerebral aqueduct, respectively. The aqueduct opens into the fourth ventricle. The floor of the fourth ventricle is the dorsal surface of the pons and upper medulla (that grey matter making a continuation of the tegmentum of the midbrain). The fourth ventricle then narrows into the central canal of the spinal cord.
The ventricular system opens to the subarachnoid space from the fourth ventricle. The single median aperture and the pair of lateral apertures connect to the subarachnoid space so that CSF can flow through the ventricles and around the outside of the CNS. Cerebrospinal fluid is produced within the ventricles by a type of specialised membrane called a choroid plexus. Ependymal cells (one of the types of glial cells described in the introduction to the nervous system) surround blood capillaries and filter the blood to make CSF. The fluid is a clear solution with a limited amount of the constituents of blood. It is essentially water, small molecules, and electrolytes. Oxygen and carbon dioxide are dissolved into the CSF, as they are in blood, and can diffuse between the fluid and the nervous tissue.
Cerebrospinal Fluid Circulation
The choroid plexuses are found in all four ventricles. Observed in dissection, they appear as soft, fuzzy structures that may still be pink, depending on how well the circulatory system is cleared in preparation of the tissue. The CSF is produced from components extracted from the blood, so its flow out of the ventricles is tied to the pulse of cardiovascular circulation.
From the lateral ventricles, the CSF flows into the third ventricle, where more CSF is produced, and then through the cerebral aqueduct into the fourth ventricle where even more CSF is produced. A very small amount of CSF is filtered at any one of the plexuses, for a total of about 500 millilitres daily, but it is continuously made and pulses through the ventricular system, keeping the fluid moving. From the fourth ventricle, CSF can continue down the central canal of the spinal cord, but this is essentially a cul-de-sac, so more of the fluid leaves the ventricular system and moves into the subarachnoid space through the median and lateral apertures.
Within the subarachnoid space, the CSF flows around all the CNS, providing two important functions. As with elsewhere in its circulation, the CSF picks up metabolic wastes from the nervous tissue and moves it out of the CNS. It also acts as a liquid cushion for the brain and spinal cord. By surrounding the entire system in the subarachnoid space, it provides a thin buffer around the organs within the strong, protective dura mater. The arachnoid granulations are outpocketings of the arachnoid membrane into the dural sinuses so that CSF can be reabsorbed into the blood, along with the metabolic wastes. From the dural sinuses, blood drains out of the head and neck through the jugular veins, along with the rest of the circulation for blood, to be reoxygenated by the lungs and wastes to be filtered out by the kidneys (Table 13.8.1).
Table 13.8.1. Components of CSF circulation
Lateral ventricles
Third ventricle
Cerebral aqueduct
Fourth ventricle
Central canal
Subarachnoid space
Location in CNS
Cerebrum
Diencephalon
Midbrain
Between pons/
upper medulla and cerebellum
Spinal cord
External to entire CNS
Blood vessel structure
Choroid plexus
Choroid plexus
None
Choroid Plexus
None
Arachnoid granulations
Disorders of the Central Nervous System
The supply of blood to the brain is crucial to its ability to perform many functions. Without a steady supply of oxygen, and to a lesser extent glucose, the nervous tissue in the brain cannot keep up its extensive electrical activity. These nutrients get into the brain through the blood, and if blood flow is interrupted, neurological function is compromised.
The common name for a disruption of blood supply to the brain is a stroke. It is caused by a blockage to an artery in the brain. The blockage is from some type of embolus: a blood clot, a fat embolus, or an air bubble. When the blood cannot travel through the artery, the surrounding tissue that is deprived starves and dies. Strokes will often result in the loss of very specific functions. A stroke in the lateral medulla, for example, can cause a loss in the ability to swallow. Sometimes, seemingly unrelated functions will be lost because they are dependent on structures in the same region. Along with the swallowing in the previous example, a stroke in that region could affect sensory functions from the face or extremities because important white matter pathways also pass through the lateral medulla. Loss of blood flow to specific regions of the cortex can lead to the loss of specific higher functions, from the ability to recognise faces to the ability to move a particular region of the body. Severe or limited memory loss can be the result of a temporal lobe stroke.
Related to strokes are transient ischemic attacks (TIAs), which can also be called “mini-strokes.” These are events in which a physical blockage may be temporary, cutting off the blood supply and oxygen to a region, but not to the extent that it causes cell death in that region. While the neurons in that area are recovering from the event, neurological function may be lost. Function can return if the area is able to recover from the event.
Recovery from a stroke (or TIA) is strongly dependent on the speed of treatment. Often, the person who is present and notices something is wrong must then decide. The mnemonic FAST helps people remember what to look for when someone is dealing with sudden losses of neurological function. If someone complains of feeling “funny,” check these things quickly: Look at the person’s face. Does he or she have problems moving Face muscles and making regular facial expressions? Ask the person to raise his or her Arms above the head. Can the person lift one arm but not the other? Has the person’s Speech changed? Is he or she slurring words or having trouble saying things? If any of these things have happened, then it is Time to call for help.
Sometimes, treatment with blood-thinning drugs can alleviate the problem, and recovery is possible. If the tissue is damaged, the amazing thing about the nervous system is that it is adaptable. With physical, occupational, and speech therapy, victims of strokes can recover, or more accurately relearn, functions.
Section Review
The CNS has a privileged blood supply established by the blood-brain barrier. Establishing this barrier are anatomical structures that help to protect and isolate the CNS. The arterial blood to the brain comes from the internal carotid and vertebral arteries, which both contribute to the unique circle of Willis that provides constant perfusion of the brain even if one of the blood vessels is blocked or narrowed. That blood is eventually filtered to make a separate medium, the CSF, that circulates within the spaces of the brain and then into the surrounding space defined by the meninges, the protective covering of the brain and spinal cord.
The blood that nourishes the brain and spinal cord is behind the glial-cell–enforced blood-brain barrier, which limits the exchange of material from blood vessels with the interstitial fluid of the nervous tissue. Thus, metabolic wastes are collected in cerebrospinal fluid that circulates through the CNS. This fluid is produced by filtering blood at the choroid plexuses in the four ventricles of the brain. It then circulates through the ventricles and into the subarachnoid space, between the pia mater and the arachnoid mater. From the arachnoid granulations, CSF is reabsorbed into the blood, removing the waste from the privileged central nervous tissue.
The blood, now with the reabsorbed CSF, drains out of the cranium through the dural sinuses. The dura mater is the tough outer covering of the CNS, which is anchored to the inner surface of the cranial and vertebral cavities. It surrounds the venous space known as the dural sinuses, which connect to the jugular veins, where blood drains from the head and neck.
Distinguish between somatic and autonomic structures, including the special peripheral structures of the enteric nervous system
Name the twelve cranial nerves and explain the functions associated with each
Describe the sensory and motor components of spinal nerves and the plexuses that they pass through
The PNS is not as contained as the CNS because it is defined as everything that is not the CNS. Some peripheral structures are incorporated into the other organs of the body. In describing the anatomy of the PNS, it is necessary to describe the common structures, the nerves and the ganglia, as they are found in various parts of the body. Many of the neural structures that are incorporated into other organs are features of the digestive system; these structures are known as the enteric nervous system and are a special subset of the PNS.
Ganglia
A ganglion is a group of neuron cell bodies in the periphery. Ganglia can be categorised, for the most part, as either sensory ganglia or autonomic ganglia, referring to their primary functions. The most common type of sensory ganglion is a dorsal (posterior) root ganglion. These ganglia are the cell bodies of neurons with axons that are sensory endings in the periphery, such as in the skin, and that extend into the CNS through the dorsal nerve root. The ganglion is an enlargement of the nerve root. Under microscopic inspection, it can be seen to include the cell bodies of the neurons, as well as bundles of fibres that are the posterior nerve root (Figure 13.9.1). The cells of the dorsal root ganglion are unipolar cells, classifying them by shape. Also, the small round nuclei of satellite cells can be seen surrounding—as if they were orbiting—the neuron cell bodies.
Another type of sensory ganglion is a cranial nerveganglion. This is analogous to the dorsal root ganglion, except that it is associated with a cranial nerve instead of a spinal nerve. The roots of cranial nerves are within the cranium, whereas the ganglia are outside the skull, for example, the trigeminal ganglion is superficial to the temporal bone whereas its associated nerve is attached to the mid-pons region of the brain stem. The neurons of cranial nerve ganglia are also unipolar in shape with associated satellite cells.
The other major category of ganglia are those of the autonomic nervous system, which is divided into the sympathetic and parasympathetic nervous systems. The sympathetic chain ganglia constitute a row of ganglia along the vertebral column that receive central input from the lateral horn of the thoracic and upper lumbar spinal cord. Superior to the chain ganglia are three paravertebral ganglia in the cervical region. Three other autonomic ganglia that are related to the sympathetic chain are the prevertebral ganglia, which are located outside of the chain but have similar functions. They are referred to as prevertebral because they are anterior to the vertebral column. The neurons of these autonomic ganglia are multipolar in shape, with dendrites radiating out around the cell body where synapses from the spinal cord neurons are made. The neurons of the chain, paravertebral, and prevertebral ganglia then project to organs in the head and neck, thoracic, abdominal, and pelvic cavities to regulate the sympathetic aspect of homeostatic mechanisms.
Another group of autonomic ganglia are the terminal ganglia that receive input from cranial nerves or sacral spinal nerves and are responsible for regulating the parasympathetic aspect of homeostatic mechanisms. These two sets of ganglia, sympathetic and parasympathetic, often project to the same organs—one input from the chain ganglia and one input from a terminal ganglion—to regulate the overall function of an organ. The heart, for example, receives two inputs such as these; one increases heart rate, and the other decreases it. The terminal ganglia that receive input from cranial nerves are found in the head and neck, as well as the thoracic and upper abdominal cavities, whereas the terminal ganglia that receive sacral input are in the lower abdominal and pelvic cavities.
Terminal ganglia below the head and neck are often incorporated into the wall of the target organ as a plexus. A plexus, in a general sense, is a network of fibres or vessels. This can apply to nervous tissue (as in this instance) or structures containing blood vessels (such as a choroid plexus), for example, the enteric plexus is the extensive network of axons and neurons in the wall of the small and large intestines. The enteric plexus is actually part of the enteric nervous system, along with the gastric plexuses and the oesophageal plexus. Though the enteric nervous system receives input originating from central neurons of the autonomic nervous system, it does not require CNS input to function. In fact, it operates independently to regulate the digestive system.
Nerves
Bundles of axons in the PNS are referred to as nerves. These structures in the periphery are different than the central counterpart, called a tract. Nerves are composed of more than just nervous tissue. They have connective tissues invested in their structure, as well as blood vessels supplying the tissues with nourishment. The outer surface of a nerve is a surrounding layer of fibrous connective tissue called the epineurium. Within the nerve, axons are further bundled into fascicles, which are each surrounded by their own layer of fibrous connective tissue called perineurium. Finally, individual axons are surrounded by loose connective tissue called the endoneurium (Figure 13.9.3). These three layers are similar to the connective tissue sheaths for muscles. Nerves are associated with the region of the CNS to which they are connected, either as cranial nerves connected to the brain or spinal nerves connected to the spinal cord.
The nerves attached to the brain are the cranial nerves, which are primarily responsible for the sensory and motor functions of the head and neck (one of these nerves targets organs in the thoracic and abdominal cavities as part of the parasympathetic nervous system). There are twelve cranial nerves, which are designated CNI through CNXII for “Cranial Nerve,” using Roman numerals for 1 through 12. They can be classified as sensory nerves, motor nerves, or a combination of both, meaning that the axons in these nerves originate out of sensory ganglia external to the cranium or motor nuclei within the brain stem. Sensory axons enter the brain to synapse in a nucleus. Motor axons connect to skeletal muscles of the head or neck. Three of the nerves are solely composed of sensory fibres; five are strictly motor; and the remaining four are mixed nerves.
Learning the cranial nerves is a tradition in anatomy courses, and students have always used mnemonic devices to remember the nerve names. A traditional mnemonic is the rhyming couplet, “On Old Olympus’ Towering Tops/A Finn And German Viewed Some Hops,” in which the initial letter of each word corresponds to the initial letter in the name of each nerve. The names of the nerves have changed over the years to reflect current usage and more accurate naming. An exercise to help learn this sort of information is to generate a mnemonic using words that have personal significance. The names of the cranial nerves are listed in Table 13.9.1 along with a brief description of their function, their source (sensory ganglion or motor nucleus), and their target (sensory nucleus or skeletal muscle). They are listed here with a brief explanation of each nerve (Figure 13.9.5).
The olfactory nerve and optic nerve are responsible for the sense of smell and vision, respectively. The oculomotor nerve is responsible for eye movements by controlling four of the extraocular muscles. It is also responsible for lifting the upper eyelid when the eyes point up, and for pupillary constriction. The trochlear nerve and the abducens nerve are both responsible for eye movement but do so by controlling different extraocular muscles. The trigeminal nerve is responsible for cutaneous sensations of the face and controlling the muscles of mastication. The facial nerve is responsible for the muscles involved in facial expressions, as well as part of the sense of taste and the production of saliva. The vestibulocochlear nerve is responsible for the senses of hearing and balance. The glossopharyngeal nerve is responsible for controlling muscles in the oral cavity and upper throat, as well as part of the sense of taste and the production of saliva. The vagus nerve is responsible for contributing to homeostatic control of the organs of the thoracic and upper abdominal cavities. The spinal accessory nerve is responsible for controlling the muscles of the neck, along with cervical spinal nerves. The hypoglossal nerve is responsible for controlling the muscles of the lower throat and tongue.
Figure 13.9.5. The cranial nerves. The anatomical arrangement of the roots of the cranial nerves observed from an inferior view of the brain.
Three of the cranial nerves also contain autonomic fibres, and a fourth is purely a component of the autonomic system. The oculomotor, facial, and glossopharyngeal nerves contain fibres that contact autonomic ganglia. The oculomotor fibres initiate pupillary constriction, whereas the facial and glossopharyngeal fibres both initiate salivation. The vagus nerve primarily targets autonomic ganglia in the thoracic and upper abdominal cavities.
Another important aspect of the cranial nerves that lends itself to a mnemonic is the functional role each nerve plays. The nerves fall into one of three basic groups. They are sensory, motor, or both (see Table 13.9.1). The sentence, “Some Say Marry Money But My Brother Says Brains Beauty Matter More,” corresponds to the basic function of each nerve. The first, second, and eighth nerves are purely sensory: the olfactory (CNI), optic (CNII), and vestibulocochlear (CNVIII) nerves. The three eye-movement nerves are all motor: the oculomotor (CNIII), trochlear (CNIV), and abducens (CNVI). The spinal accessory (CNXI) and hypoglossal (CNXII) nerves are also strictly motor. The remainder of the nerves contain both sensory and motor fibres. They are the trigeminal (CNV), facial (CNVII), glossopharyngeal (CNIX), and vagus (CNX) nerves. The nerves that convey both are often related to each other. The trigeminal and facial nerves both concern the face; one concerns the sensations and the other concerns the muscle movements. The facial and glossopharyngeal nerves are both responsible for conveying gustatory, or taste, sensations as well as controlling salivary glands. The vagus nerve is involved in visceral responses to taste, namely the gag reflex. This is not an exhaustive list of what these combination nerves do, but there is a thread of relation between them.
Terminal ganglia serving thoracic and upper abdominal organs (heart and small intestine)
Some
XI
Spinal accessory
Motor – head and neck (M)
Spinal accessory nucleus
Neck muscles
Hops
XII
Hypoglossal
Motor – lower throat (M)
Hypoglossal nucleus
Muscles of the larynx and lower pharynx
Spinal Nerves
The nerves connected to the spinal cord are the spinal nerves. The arrangement of these nerves is much more regular than that of the cranial nerves. All of the spinal nerves are combined sensory and motor axons that separate into two nerve roots. The sensory axons enter the spinal cord as the dorsal nerve root. The motor fibres, both somatic and autonomic, emerge as the ventral nerve root. The dorsal root ganglion for each nerve is an enlargement of the spinal nerve.
There are 31 spinal nerves, named for the level of the spinal cord at which each one emerges. There are eight pairs of cervical nerves designated C1 to C8, twelve thoracic nerves designated T1 to T12, five pairs of lumbar nerves designated L1 to L5, five pairs of sacral nerves designated S1 to S5, and one pair of coccygeal nerves. The nerves are numbered from the superior to inferior positions, and each emerges from the vertebral column through the intervertebral foramen at its level. The first nerve, C1, emerges between the first cervical vertebra and the occipital bone. The second nerve, C2, emerges between the first and second cervical vertebrae. The same occurs for C3 to C7, but C8 emerges between the seventh cervical vertebra and the first thoracic vertebra. For the thoracic and lumbar nerves, each one emerges between the vertebra that has the same designation and the next vertebra in the column. The sacral nerves emerge from the sacral foramina along the length of that unique vertebra.
Spinal nerves extend outward from the vertebral column to enervate the periphery. The nerves in the periphery are not straight continuations of the spinal nerves, but the reorganisation of the axons in those nerves to follow different courses. Axons from different spinal nerves will come together into a systemic nerve. This occurs at four places along the length of the vertebral column, each identified as a nerve plexus, whereas the other spinal nerves directly correspond to nerves at their respective levels. In this instance, the word plexus is used to describe networks of nerve fibres with no associated cell bodies.
Of the four nerve plexuses, two are found at the cervical level, one at the lumbar level, and one at the sacral level (Figure 13.9.6). The cervical plexus is composed of axons from spinal nerves C1 through C5 and branches into nerves in the posterior neck and head, as well as the phrenic nerve, which connects to the diaphragm at the base of the thoracic cavity. The other plexus from the cervical level is the brachial plexus. Spinal nerves C4 through T1 reorganise through this plexus to give rise to the nerves of the arms, as the name brachial suggests. A large nerve from this plexus is the radial nerve from which the axillary nerve branches to go to the armpit region. The radial nerve continues through the arm and is paralleled by the ulnar nerve and the median nerve. The lumbar plexus arises from all the lumbar spinal nerves and gives rise to nerves enervating the pelvic region and the anterior leg. The femoral nerve is one of the major nerves from this plexus, which gives rise to the saphenous nerve as a branch that extends through the anterior lower leg. The sacral plexus comes from the lower lumbar nerves L4 and L5 and the sacral nerves S1 to S4. The most significant systemic nerve to come from this plexus is the sciatic nerve, which is a combination of the tibial nerve and the fibular nerve. The sciatic nerve extends across the hip joint and is most associated with the condition sciatica, which is the result of compression or irritation of the nerve or any of the spinal nerves giving rise to it.
These plexuses are described as arising from spinal nerves and giving rise to certain systemic nerves, but they contain fibres that serve sensory functions or fibres that serve motor functions. This means that some fibres extend from cutaneous or other peripheral sensory surfaces and send action potentials into the CNS. Those are axons of sensory neurons in the dorsal root ganglia that enter the spinal cord through the dorsal nerve root. Other fibres are the axons of motor neurons of the anterior horn of the spinal cord, which emerge in the ventral nerve root and send action potentials to cause skeletal muscles to contract in their target regions. The radial nerve, for example, contains fibres of cutaneous sensation in the arm, as well as motor fibres that move muscles in the arm.
Spinal nerves of the thoracic region, T2 through T11, are not part of the plexuses but emerge and give rise to the intercostal nerves found between the ribs, which articulate with the vertebrae surrounding the spinal nerve.
Figure 13.9.6. Nerve plexuses of the body. There are four main nerve plexuses in the human body. The cervical plexus supplies nerves to the posterior head and neck, as well as to the diaphragm. The brachial plexus supplies nerves to the arm. The lumbar plexus supplies nerves to the anterior leg. The sacral plexus supplies nerves to the posterior leg.
Ageing and the Nervous System
Anosmia is the loss of the sense of smell. It is often the result of the olfactory nerve being severed, usually because of blunt force trauma to the head. The sensory neurons of the olfactory epithelium have a limited lifespan of approximately one to four months, and new ones are made on a regular basis. The new neurons extend their axons into the CNS by growing along the existing fibres of the olfactory nerve. The ability of these neurons to be replaced is lost with age. Age-related anosmia is not the result of impact trauma to the head, but rather a slow loss of the sensory neurons with no new neurons born to replace them.
Smell is an important sense, especially for the enjoyment of food. There are only five tastes sensed by the tongue, and two of them are generally thought of as unpleasant tastes (sour and bitter). The rich sensory experience of food is the result of odour molecules associated with the food, both as food is moved into the mouth, and therefore passes under the nose, and when it is chewed and molecules are released to move up the pharynx into the posterior nasal cavity. Anosmia results in a loss of the enjoyment of food.
As the replacement of olfactory neurons declines with age, anosmia can set in. Without the sense of smell, many sufferers complain of food tasting bland. Often, the only way to enjoy food is to add seasoning that can be sensed on the tongue, which usually means adding table salt. The problem with this solution, however, is that this increases sodium intake, which can lead to cardiovascular problems through water retention and the associated increase in blood pressure.
Section Review
The PNS is composed of the groups of neurons (ganglia) and bundles of axons (nerves) that are outside of the brain and spinal cord. Ganglia are of two types, sensory or autonomic. Sensory ganglia contain unipolar sensory neurons and are found on the dorsal root of all spinal nerves as well as associated with many of the cranial nerves. Autonomic ganglia are in the sympathetic chain, the associated paravertebral or prevertebral ganglia, or in terminal ganglia near or within the organs controlled by the autonomic nervous system.
Nerves are classified as cranial nerves or spinal nerves on the basis of their connection to the brain or spinal cord, respectively. The twelve cranial nerves can be strictly sensory in function, strictly motor in function, or a combination of the two functions. Sensory fibres are axons of sensory ganglia that carry sensory information into the brain and target sensory nuclei. Motor fibres are axons of motor neurons in motor nuclei of the brain stem and target skeletal muscles of the head and neck. Spinal nerves are all mixed nerves with both sensory and motor fibres. Spinal nerves emerge from the spinal cord and reorganise through plexuses, which then give rise to systemic nerves. Thoracic spinal nerves are not part of any plexus but give rise to the intercostal nerves directly.
Describe the structures responsible for the special senses of taste, smell, hearing, balance, and vision
Distinguish how different tastes are transduced
Describe the means of mechanoreception for hearing and balance
List the supporting structures around the eye and describe the structure of the eyeball
Describe the processes of phototransduction
A major role of sensory receptors is to help us learn about the environment around us, or about the state of our internal environment. Stimuli from varying sources, and of different types, are received and changed into the electrochemical signals of the nervous system. This occurs when a stimulus changes the cell membrane potential of a sensory neuron. The stimulus causes the sensory cell to produce an action potential that is relayed into the central nervous system (CNS), where it is integrated with other sensory information—or sometimes higher cognitive functions—to become a conscious perception of that stimulus. The central integration may then lead to a motor response.
Describing sensory function with the term sensation or perception is a deliberate distinction. Sensation is the activation of sensory receptor cells at the level of the stimulus. Perception is the central processing of sensory stimuli into a meaningful pattern. Perception is dependent on sensation, but not all sensations are perceived. Receptors are the cells or structures that detect sensations. A receptor cell is changed directly by a stimulus. A transmembrane protein receptor is a protein in the cell membrane that mediates a physiological change in a neuron, most often through the opening of ion channels or changes in the cell signalling processes. Transmembrane receptors are activated by chemicals called ligands, for example, a molecule in food can serve as a ligand for taste receptors. Other transmembrane proteins, which are not accurately called receptors, are sensitive to mechanical or thermal changes. Physical changes in these proteins increase ion flow across the membrane and can generate an action potential or a graded potential in the sensory neurons.
Sensory Receptors
Stimuli in the environment activate specialised receptor cells in the peripheral nervous system. Different types of stimuli are sensed by different types of receptor cells. Receptor cells can be classified into types based on three different criteria: cell type, position, and function. Receptors can be classified structurally based on cell type and their position in relation to stimuli they sense. They can also be classified functionally based on the transduction of stimuli, or how the mechanical stimulus, light, or chemical changed the cell membrane potential.
Structural Receptor Types
The cells that interpret information about the environment can be either (1) a neuron that has a free nerve ending, with dendrites embedded in tissue that would receive a sensation; (2) a neuron that has an encapsulated ending in which the sensory nerve endings are encapsulated in connective tissue that enhances their sensitivity; or (3) a specialised receptor cell, which has distinct structural components that interpret a specific type of stimulus (Figure 13.10.1). The pain and temperature receptors in the dermis of the skin are examples of neurons that have free nerve endings. Also located in the dermis of the skin are lamellated corpuscles, neurons with encapsulated nerve endings that respond to pressure and touch. The cells in the retina that respond to light stimuli are an example of a specialised receptor, a photoreceptor.
Figure 13.10.1. Receptor classification by cell type. Receptor cell types can be classified based on their structure. Sensory neurons can have either (a) free nerve endings or (b) encapsulated endings. Photoreceptors in the eyes, such as rod cells, are examples of (c) specialised receptor cells. These cells release neurotransmitters onto a bipolar cell, which then synapses with the optic nerve neurons.
Another way that receptors can be classified is based on their location relative to the stimuli. An exteroceptor is a receptor that is located near a stimulus in the external environment, such as the somatosensory receptors that are located in the skin. An interoceptor is one that interprets stimuli from internal organs and tissues, such as the receptors that sense the increase in blood pressure in the aorta or carotid sinus. Finally, a proprioceptor is a receptor located near a moving part of the body, such as a muscle, that interprets the positions of the tissues as they move.
Functional Receptor Types
A third classification of receptors is by how the receptor transduces stimuli into membrane potential changes. Stimuli are of three general types. Some stimuli are ions and macromolecules that affect transmembrane receptor proteins when these chemicals diffuse across the cell membrane. Some stimuli are physical variations in the environment that affect receptor cell membrane potentials. Other stimuli include the electromagnetic radiation from visible light. For humans, the only electromagnetic energy that is perceived by our eyes is visible light. Some other organisms have receptors that humans lack, such as the heat sensors of snakes, the ultraviolet light sensors of bees, or magnetic receptors in migratory birds.
Receptor cells can be further categorised based on the type of stimuli they transduce. Chemical stimuli can be interpreted by a chemoreceptor that interprets chemical stimuli, such as an object’s taste or smell. Osmoreceptors respond to solute concentrations of body fluids. Additionally, pain is primarily a chemical sense that interprets the presence of chemicals from tissue damage, or similar intense stimuli, through a nociceptor. Physical stimuli, such as pressure and vibration, as well as the sensation of sound and body position (balance), are interpreted through a mechanoreceptor. Another physical stimulus that has its own type of receptor is temperature, which is sensed through a thermoreceptor that is either sensitive to temperatures above (heat) or below (cold) normal body temperature.
Sensory Modalities
Ask anyone what the senses are, and they are likely to list the five major senses—taste, smell, touch, hearing, and sight. However, these are not all the senses. The most obvious omission from this list is balance. Also, what is referred to simply as touch can be further subdivided into pressure, vibration, stretch, and hair-follicle position, based on the type of mechanoreceptors that perceive these touch sensations. Other overlooked senses include temperature perception by thermoreceptors and pain perception by nociceptors.
Within the realm of physiology, senses can be classified as either general or specific. A general sense is one that is distributed throughout the body and has receptor cells within the structures of other organs. Mechanoreceptors in the skin, muscles, or the walls of blood vessels are examples of this type. General senses often contribute to the sense of touch, as described above, or to proprioception (body movement) and kinaesthesia (body movement), or to a visceral sense, which is most important to autonomic functions. A special sense is one that has a specific organ devoted to it, namely the eye, inner ear, tongue, or nose.
Each of the senses is referred to as a sensory modality. Modality refers to the way that information is encoded, which is similar to the idea of transduction. The main sensory modalities can be described based on how each is transduced. The chemical senses are taste and smell. The general sense that is usually referred to as touch includes chemical sensation in the form of nociception, or pain. Pressure, vibration, muscle stretch, and the movement of hair by an external stimulus, are all sensed by mechanoreceptors. Hearing and balance are also sensed by mechanoreceptors. Finally, vision involves the activation of photoreceptors.
Listing all the different sensory modalities, which can number as many as 17, involves separating the five major senses into more specific categories, or submodalities, of the larger sense. An individual sensory modality represents the sensation of a specific type of stimulus, for example, the general sense of touch, which is known as somatosensation, can be separated into light pressure, deep pressure, vibration, itch, pain, temperature, or hair movement.
Gustation (Taste)
Only a few recognised submodalities exist within the sense of taste, or gustation. Until recently, only four tastes were recognised: sweet, salty, sour, and bitter. Research at the turn of the 20th century led to recognition of the fifth taste, umami, during the mid-1980s. Umami is a Japanese word that means “delicious taste,” and is often translated to mean savory. Very recent research has suggested that there may also be a sixth taste for fats, or lipids.
Gustation is the special sense associated with the tongue. The surface of the tongue, along with the rest of the oral cavity, is lined by a stratified squamous epithelium. Raised bumps called papillae (singular = papilla) contain the structures for gustatory transduction. There are four types of papillae, based on their appearance (Figure 13.10.2): circumvallate, foliate, filiform, and fungiform. Within the structure of the papillae are taste buds that contain specialised gustatory receptor cells for the transduction of taste stimuli. These receptor cells are sensitive to the chemicals contained within foods that are ingested, and they release neurotransmitters based on the amount of the chemical in the food. Neurotransmitters from the gustatory cells can activate sensory neurons in the facial, glossopharyngeal, and vagus cranial nerves.
Salty taste is simply the perception of sodium ions (Na+) in the saliva. When you eat something salty, the salt crystals dissociate into the component ions Na+ and Cl–, which dissolve into the saliva in your mouth. The Na+ concentration becomes high outside the gustatory cells, creating a strong concentration gradient that drives the diffusion of the ion into the cells. The entry of Na+ into these cells results in the depolarisation of the cell membrane and the generation of a receptor potential.
Sour taste is the perception of H+ concentration. Just as with sodium ions in salty flavours, these hydrogen ions enter the cell and trigger depolarisation. Sour flavours are, essentially, the perception of acids in our food. Increasing hydrogen ion concentrations in the saliva (lowering saliva pH) triggers progressively stronger graded potentials in the gustatory cells. Orange juice, for example, —which contains citric acid—will taste sour because it has a pH value of approximately 3. Of course, it is often sweetened so that the sour taste is masked.
The first two tastes (salty and sour) are triggered by the cations Na+ and H+. The other tastes result from food molecules binding to a G protein–coupled receptor. A G protein signal transduction system ultimately leads to depolarisation of the gustatory cell. The sweet taste is the sensitivity of gustatory cells to the presence of glucose dissolved in the saliva. Other monosaccharides such as fructose, or artificial sweeteners such as aspartame (NutraSweet™), saccharine, or sucralose (Splenda™) also activate the sweet receptors. The affinity for each of these molecules varies, and some will taste sweeter than glucose because they bind to the G protein–coupled receptor differently.
Bitter taste is similar to sweet in that food molecules bind to G protein–coupled receptors. However, there are a number of different ways in which this can happen because there is a large diversity of bitter-tasting molecules. Some bitter molecules depolarise gustatory cells, whereas others hyperpolarise gustatory cells. Likewise, some bitter molecules increase G protein activation within the gustatory cells, whereas other bitter molecules decrease G protein activation. The specific response depends on which molecule is binding to the receptor.
One major group of bitter-tasting molecules are alkaloids. Alkaloids are nitrogen containing molecules that are commonly found in bitter-tasting plant products, such as coffee, hops (in beer), tannins (in wine), tea, and aspirin. By containing toxic alkaloids, the plant is less susceptible to microbe infection and less attractive to herbivores.
Therefore, the function of bitter taste may primarily be related to stimulating the gag reflex to avoid ingesting poisons. Because of this, many bitter foods that are normally ingested are often combined with a sweet component to make them more palatable (cream and sugar in coffee, for example). The highest concentration of bitter receptors appear to be in the posterior tongue, where a gag reflex could still spit out poisonous food.
The taste known as umami is often referred to as the savory taste. Like sweet and bitter, it is based on the activation of G protein–coupled receptors by a specific molecule. The molecule that activates this receptor is the amino acid L-glutamate. Therefore, the umami flavour is often perceived while eating protein-rich foods. Not surprisingly, dishes that contain meat are often described as savory.
Once the gustatory cells are activated by the taste molecules, they release neurotransmitters onto the dendrites of sensory neurons. These neurons are part of the facial and glossopharyngeal cranial nerves, as well as a component within the vagus nerve dedicated to the gag reflex. The facial nerve connects to taste buds in the anterior third of the tongue. The glossopharyngeal nerve connects to taste buds in the posterior two thirds of the tongue. The vagus nerve connects to taste buds in the extreme posterior of the tongue, verging on the pharynx, which are more sensitive to noxious stimuli such as bitterness.
Olfaction (Smell)
Like taste, the sense of smell, or olfaction, is also responsive to chemical stimuli. The olfactory receptor neurons are located in a small region within the superior nasal cavity (Figure 13.10.3). This region is referred to as the olfactory epithelium and contains bipolar sensory neurons. Each olfactory sensory neuron has dendrites that extend from the apical surface of the epithelium into the mucus lining the cavity. As airborne molecules are inhaled through the nose, they pass over the olfactory epithelial region and dissolve into the mucus. These odorant molecules bind to proteins that keep them dissolved in the mucus and help transport them to the olfactory dendrites. The odorant–protein complex binds to a receptor protein within the cell membrane of an olfactory dendrite. These receptors are G protein–coupled and will produce a graded membrane potential in the olfactory neurons.
The axon of an olfactory neuron extends from the basal surface of the epithelium, through an olfactory foramen in the cribriform plate of the ethmoid bone, and into the brain. The group of axons called the olfactory tract connect to the olfactory bulb on the ventral surface of the frontal lobe. From there, the axons split to travel to several brain regions. Some travel to the cerebrum, specifically to the primary olfactory cortex that is located in the inferior and medial areas of the temporal lobe. Others project to structures within the limbic system and hypothalamus, where smells become associated with long-term memory and emotional responses. This is how certain smells trigger emotional memories, such as the smell of food associated with one’s birthplace. Smell is the one sensory modality that does not synapse in the thalamus before connecting to the cerebral cortex. This intimate connection between the olfactory system and the cerebral cortex is one reason why smell can be a potent trigger of memories and emotion.
The nasal epithelium, including the olfactory cells, can be harmed by airborne toxic chemicals. Therefore, the olfactory neurons are regularly replaced within the nasal epithelium, after which the axons of the new neurons must find their appropriate connections in the olfactory bulb. These new axons grow along the axons that are already in place in the cranial nerve.
Blunt force trauma to the face, such as that common in many car accidents, can lead to the loss of the olfactory nerve, and subsequently, loss of the sense of smell. This condition is known as anosmia. When the frontal lobe of the brain moves relative to the ethmoid bone, the olfactory tract axons may be sheared apart. Professional fighters often experience anosmia because of repeated trauma to face and head. In addition, certain pharmaceuticals, such as antibiotics, can cause anosmia by killing all the olfactory neurons at once. If no axons are in place within the olfactory nerve, then the axons from newly formed olfactory neurons have no guide to lead them to their connections within the olfactory bulb. There are temporary causes of anosmia, as well, such as those caused by inflammatory responses related to respiratory infections or allergies.
Loss of the sense of smell can result in food tasting bland. A person with an impaired sense of smell may require additional spice and seasoning levels for food to be tasted. Anosmia may also be related to some presentations of mild depression because the loss of enjoyment of food may lead to a general sense of despair.
The ability of olfactory neurons to replace themselves decreases with age, leading to age-related anosmia. This explains why some elderly people salt their food more than younger people do. However, this increased sodium intake can increase blood volume and blood pressure, increasing the risk of cardiovascular diseases in the elderly.
Audition (Hearing)
Hearing, or audition, is the transduction of sound waves into a neural signal that is made possible by the structures of the ear (Figure 13.10.4). The large, fleshy structure on the lateral aspect of the head is known as the auricle. Some sources will also refer to this structure as the pinna, though that term is more appropriate for a structure that can be moved, such as the external ear of a cat. The C-shaped curves of the auricle direct sound waves toward the auditory canal. The canal enters the skull through the external auditory meatus of the temporal bone. At the end of the auditory canal is the tympanic membrane, or ear drum, which vibrates after it is struck by sound waves. The auricle, ear canal, and tympanic membrane are often referred to as the external ear. The middle ear consists of a space spanned by three small bones called the ossicles. The three ossicles are the malleus, incus, and stapes, which are Latin names that roughly translate to hammer, anvil, and stirrup. The malleus is attached to the tympanic membrane and articulates with the incus. The incus, in turn, articulates with the stapes. The stapes is then attached to the inner ear, where the sound waves will be transduced into a neural signal. The middle ear is connected to the pharynx through the Eustachian tube, which helps equilibrate air pressure across the tympanic membrane. The tube is normally closed but will pop open when the muscles of the pharynx contract during swallowing or yawning.
Figure 13.10.4. Structures of the ear. The external ear contains the auricle, ear canal, and tympanic membrane. The middle ear contains the ossicles and is connected to the pharynx by the Eustachian tube. The inner ear contains the cochlea and vestibule, which are responsible for audition and equilibrium, respectively.
The inner ear is often described as a bony labyrinth, as it is composed of a series of canals embedded within the temporal bone. It has two separate regions, the cochlea and the vestibule, which are responsible for hearing and balance, respectively. The neural signals from these two regions are relayed to the brain stem through separate fibre bundles. However, these two distinct bundles travel together from the inner ear to the brain stem as the vestibulocochlear nerve. Sound is transduced into neural signals within the cochlear region of the inner ear, which contains the sensory neurons of the spiral ganglia. These ganglia are located within the spiral-shaped cochlea of the inner ear. The cochlea is attached to the stapes through the oval window.
The oval window is located at the beginning of a fluid-filled tube within the cochlea called the scala vestibuli. The scala vestibuli extends from the oval window, travelling above the cochlear duct, which is the central cavity of the cochlea that contains the sound-transducing neurons. At the uppermost tip of the cochlea, the scala vestibuli curves over the top of the cochlear duct. The fluid-filled tube, now called the scala tympani, returns to the base of the cochlea, this time travelling under the cochlear duct. The scala tympani ends at the round window, which is covered by a membrane that contains the fluid within the scala. As vibrations of the ossicles travel through the oval window, the fluid of the scala vestibuli and scala tympani moves in a wave-like motion. The frequency of the fluid waves match the frequencies of the sound waves (Figure 13.10.5). The membrane covering the round window will bulge out or pucker in with the movement of the fluid within the scala tympani.
Figure 13.10.5. Transmission of sound waves to cochlea. A sound wave causes the tympanic membrane to vibrate. This vibration is amplified as it moves across the malleus, incus, and stapes. The amplified vibration is picked up by the oval window causing pressure waves in the fluid of the scala vestibuli and scala tympani. The complexity of the pressure waves is determined by the changes in amplitude and frequency of the sound waves entering the ear.
A cross-sectional view of the cochlea shows that the scala vestibuli and scala tympani run along both sides of the cochlear duct (Figure 13.10.6). The cochlear duct contains several organs of Corti, which transduce the wave motion of the two scala into neural signals. The organs of Corti lie on top of the basilar membrane, which is the side of the cochlear duct located between the organs of Corti and the scala tympani. As the fluid waves move through the scala vestibuli and scala tympani, the basilar membrane moves at a specific spot, depending on the frequency of the waves. Higher frequency waves move the region of the basilar membrane that is close to the base of the cochlea. Lower frequency waves move the region of the basilar membrane that is near the tip of the cochlea.
Figure 13.10.6. Cross section of the cochlea. The three major spaces within the cochlea are highlighted. The scala tympani and scala vestibuli lie on either side of the cochlear duct. The organ of Corti, containing the mechanoreceptor hair cells, is adjacent to the scala tympani, where it sits atop the basilar membrane.
The organs of Corti contain hair cells, which are named for the hair-like stereocilia extending from the cell’s apical surfaces (Figure 13.10.7). The stereocilia are an array of microvilli-like structures arranged from tallest to shortest. Protein fibres tether adjacent hairs together within each array, such that the array will bend in response to movements of the basilar membrane. The stereocilia extend up from the hair cells to the overlying tectorial membrane, which is attached medially to the organ of Corti. When the pressure waves from the scala move the basilar membrane, the tectorial membrane slides across the stereocilia. This bends the stereocilia either toward or away from the tallest member of each array. When the stereocilia bend toward the tallest member of their array, tension in the protein tethers opens ion channels in the hair cell membrane. This will depolarise the hair cell membrane, triggering nerve impulses that travel down the afferent nerve fibres attached to the hair cells. When the stereocilia bend toward the shortest member of their array, the tension on the tethers slackens and the ion channels close. When no sound is present, and the stereocilia are standing straight, a small amount of tension still exists on the tethers, keeping the membrane potential of the hair cell slightly depolarised.
As stated above, a given region of the basilar membrane will only move if the incoming sound is at a specific frequency. Because the tectorial membrane only moves where the basilar membrane moves, the hair cells in this region will also only respond to sounds of this specific frequency. Therefore, as the frequency of a sound changes, different hair cells are activated all along the basilar membrane. The cochlea encodes auditory stimuli for frequencies between 20 and 20,000 Hz, which is the range of sound that human ears can detect. The unit of Hertz measures the frequency of sound waves in terms of cycles produced per second. Frequencies as low as 20 Hz are detected by hair cells at the apex, or tip, of the cochlea. Frequencies in the higher ranges of 20 KHz are encoded by hair cells at the base of the cochlea, close to the round and oval windows. Most auditory stimuli contain a mixture of sounds at a variety of frequencies and intensities (represented by the amplitude of the sound wave). The hair cells along the length of the cochlear duct, which are each sensitive to a particular frequency, allow the cochlea to separate auditory stimuli by frequency, just as a prism separates visible light into its component colours.
Figure 13.10.9. Frequency coding in the cochlea. The standing sound wave generated in the cochlea by the movement of the oval window deflects the basilar membrane based on the frequency of sound. Therefore, hair cells at the base of the cochlea are activated only by high frequencies, whereas those at the apex of the cochlea are activated only by low frequencies.
Equilibrium (Balance)
Along with audition, the inner ear is responsible for encoding information about equilibrium, the sense of balance. A similar mechanoreceptor—a hair cell with stereocilia—senses head position, head movement, and whether our bodies are in motion. These cells are located within the vestibule of the inner ear. Head position is sensed by the utricle and saccule, whereas head movement is sensed by the semicircular canals. The neural signals generated in the vestibular ganglion are transmitted through the vestibulocochlear nerve to the brain stem and cerebellum.
The utricle and saccule are both largely composed of macula tissue (plural = maculae). The macula is composed of hair cells surrounded by support cells. The stereocilia of the hair cells extend into a viscous gel called the otolithic membrane ( Figure 13.10.10). On top of the otolithic membrane is a layer of calcium carbonate crystals, called otoliths. The otoliths make the otolithic membrane top-heavy. The otolithic membrane moves separately from the macula in response to head movements. Tilting the head causes the otolithic membrane to slide over the macula in the direction of gravity. The moving otolithic membrane, in turn, bends the sterocilia, causing some hair cells to depolarise as others hyperpolarise. The exact position of the head is interpreted by the brain based on the pattern of hair-cell depolarisation.
Figure 13.10.10. Linear acceleration coding by maculae. The maculae are specialised for sensing linear acceleration, such as when gravity acts on the tilting head, or if the head starts moving in a straight line. The difference in inertia between the hair cell stereocilia and the otolithic membrane in which they are embedded leads to a shearing force that causes the stereocilia to bend in the direction of that linear acceleration.
The semicircular canals are three ring-like extensions of the vestibule. One is oriented in the horizontal plane, whereas the other two are oriented in the vertical plane. The anterior and posterior vertical canals are oriented at approximately 45 degrees relative to the sagittal plane (Figure 13.10.11). The base of each semicircular canal, where it meets with the vestibule, connects to an enlarged region known as the ampulla. The ampulla contains the hair cells that respond to rotational movement, such as turning the head while saying “no.” The stereocilia of these hair cells extend into the cupula, a membrane that attaches to the top of the ampulla. As the head rotates in a plane parallel to the semicircular canal, the fluid lags, deflecting the cupula in the direction opposite to the head movement. The semicircular canals contain several ampullae, with some oriented horizontally and others oriented vertically. By comparing the relative movements of both the horizontal and vertical ampullae, the vestibular system can detect the direction of most head movements within three-dimensional (3-D) space.
Figure 13.10.11. Rotational coding by semicircular canals. Rotational movement of the head is encoded by the hair cells in the base of the semicircular canals. As one of the canals moves in an arc with the head, the internal fluid moves in the opposite direction, causing the cupula and stereocilia to bend. The movement of two canals within a plane results in information about the direction in which the head is moving, and activation of all six canals can give a very precise indication of head movement in three dimensions.
Somatosensation (Touch)
Somatosensation is considered a general sense, as opposed to the special senses discussed in this section. Somatosensation is the group of sensory modalities that are associated with touch, proprioception, and interoception. These modalities include pressure, vibration, light touch, tickle, itch, temperature, pain, proprioception, and kinaesthesia. This means that its receptors are not associated with a specialised organ but are instead spread throughout the body in a variety of organs. Many of the somatosensory receptors are located in the skin, but receptors are also found in muscles, tendons, joint capsules, ligaments, and in the walls of visceral organs.
Two types of somatosensory signals that are transduced by free nerve endings are pain and temperature. These two modalities use thermoreceptors and nociceptors to transduce temperature and pain stimuli, respectively. Temperature receptors are stimulated when local temperatures differ from body temperature. Some thermoreceptors are sensitive to just cold and others to just heat. Nociception is the sensation of potentially damaging stimuli. Mechanical, chemical, or thermal stimuli beyond a set threshold will elicit painful sensations. Stressed or damaged tissues release chemicals that activate receptor proteins in the nociceptors. The sensation of heat associated with spicy foods, for example, involves capsaicin, the active molecule in hot peppers. Capsaicin molecules bind to a transmembrane ion channel in nociceptors that is sensitive to temperatures above 37°C. The dynamics of capsaicin binding with this transmembrane ion channel is unusual in that the molecule remains bound for a long time. Because of this, it will decrease the ability of other stimuli to elicit pain sensations through the activated nociceptor. For this reason, capsaicin can be used as a topical analgesic, such as in products such as Icy Hot™.
If you drag your finger across a textured surface, the skin of your finger will vibrate. Such low frequency vibrations are sensed by mechanoreceptors called Merkel cells, also known as type I cutaneous mechanoreceptors. Merkel cells are located in the stratum basale of the epidermis. Deep pressure and vibration is transduced by lamellated (Pacinian) corpuscles, which are receptors with encapsulated endings found deep in the dermis, or subcutaneous tissue. Light touch is transduced by the encapsulated endings known as tactile (Meissner) corpuscles. Follicles are also wrapped in a plexus of nerve endings known as the hair follicle plexus. These nerve endings detect the movement of hair at the surface of the skin, such as when an insect may be walking along the skin. Stretching of the skin is transduced by stretch receptors known as bulbous corpuscles. Bulbous corpuscles are also known as Ruffini corpuscles, or type II cutaneous mechanoreceptors.
Other somatosensory receptors are found in the joints and muscles. Stretch receptors monitor the stretching of tendons, muscles, and the components of joints. Have you ever stretched your muscles before or after exercise and noticed that you can only stretch so far before your muscles spasm back to a less stretched state? This spasm is a reflex that is initiated by stretch receptors to avoid muscle tearing. Such stretch receptors can also prevent over-contraction of a muscle. In skeletal muscle tissue, these stretch receptors are called muscle spindles. Golgi tendon organs similarly transduce the stretch levels of tendons. Bulbous corpuscles are also present in joint capsules, where they measure stretch in the components of the skeletal system within the joint. The types of nerve endings, their locations, and the stimuli they transduce are presented in Table 13.10.1.
Table 13.10.1. Mechanoreceptors of somatosensation
Papillary dermis, especially in the fingertips and lips
Light touch, vibrations below 50 Hz
Lamellated corpuscle
Pacinian corpuscle
Deep dermis, subcutaneous tissue
Deep pressure, high frequency vibration (around 250 Hz)
Hair follicle
*
Wrapped around hair follicles in the dermis
Movement of hair
Muscle spindle
*
In line with skeletal muscle fibres
Muscle contraction and stretch
Tendon stretch organ
Golgi tendon organ
In line with tendons
Stretch of tendons
* No corresponding eponymous name
Vision
Vision is the special sense of sight that is based on the transduction of light stimuli received through the eyes. The eyes are located within either orbit in the skull. The bony orbits surround the eyeballs, protecting them and anchoring the soft tissues of the eye (Figure 13.10.12). The eyelids, with lashes at their leading edges, help to protect the eye from abrasions by blocking particles that may land on the surface of the eye. The inner surface of each lid is a thin membrane known as the palpebral conjunctiva. The conjunctiva extends over the white areas of the eye (the sclera), connecting the eyelids to the eyeball. Tears are produced by the lacrimal gland, located beneath the lateral edges of the nose. Tears produced by this gland flow through the lacrimal duct to the medial corner of the eye, where the tears flow over the conjunctiva, washing away foreign particles.
Figure 13.10.12. The eye in the orbit. The eye is located within the orbit and surrounded by soft tissues that protect and support its function. The orbit is surrounded by cranial bones of the skull.
Movement of the eye within the orbit is accomplished by the contraction of six extraocular muscles that originate from the bones of the orbit and insert into the surface of the eyeball (Figure 13.10.). Four of the muscles are arranged at the cardinal points around the eye and are named for those locations. They are the superior rectus, medial rectus, inferior rectus, and lateral rectus. When each of these muscles contract, the eye to moves toward the contracting muscle, for example, when the superior rectus contracts, the eye rotates to look up. The superior oblique originates at the posterior orbit, near the origin of the four rectus muscles. However, the tendon of the oblique muscles threads through a pulley-like piece of cartilage known as the trochlea. The tendon inserts obliquely into the superior surface of the eye. The angle of the tendon through the trochlea means that contraction of the superior oblique rotates the eye medially. The inferior oblique muscle originates from the floor of the orbit and inserts into the inferolateral surface of the eye. When it contracts, it laterally rotates the eye, in opposition to the superior oblique. Rotation of the eye by the two oblique muscles is necessary because the eye is not perfectly aligned on the sagittal plane. When the eye looks up or down, the eye must also rotate slightly to compensate for the superior rectus pulling at approximately a 20-degree angle, rather than straight up. The same is true for the inferior rectus, which is compensated by contraction of the inferior oblique. A seventh muscle in the orbit is the levator palpebrae superioris, which is responsible for elevating and retracting the upper eyelid, a movement that usually occurs in concert with elevation of the eye by the superior rectus (see Figure 13.10.13).
The extraocular muscles are innervated by three cranial nerves. The lateral rectus, which causes abduction of the eye, is innervated by the abducens nerve. The superior oblique is innervated by the trochlear nerve. All of the other muscles are innervated by the oculomotor nerve, as is the levator palpebrae superioris. The motor nuclei of these cranial nerves connect to the brain stem, which coordinates eye movements.
Figure 13.10.13. Extraocular muscles. The extraocular muscles move the eye within the orbit.
The eye itself is a hollow sphere composed of three layers of tissue. The outermost layer is the fibrous tunic, which includes the white sclera and clear cornea. The sclera accounts for five sixths of the surface of the eye, most of which is not visible, though humans are unique compared with many other species in having so much of the “white of the eye” visible (Figure 13.10.14). The transparent cornea covers the anterior tip of the eye and allows light to enter the eye. The middle layer of the eye is the vascular tunic, which is mostly composed of the choroid, ciliary body, and iris. The choroid is a layer of highly vascularised connective tissue that provides a blood supply to the eyeball. The choroid is posterior to the ciliary body, a muscular structure that is attached to the lens by zonule fibres. These two structures bend the lens, allowing it to focus light on the back of the eye. Overlaying the ciliary body, and visible in the anterior eye, is the iris—the coloured part of the eye. The iris is a smooth muscle that opens or closes the pupil, which is the hole at the centre of the eye that allows light to enter. The iris constricts the pupil in response to bright light and dilates the pupil in response to dim light. The innermost layer of the eye is the neural tunic, or retina, which contains the nervous tissue responsible for photoreception.
The eye is also divided into two cavities: the anterior cavity and the posterior cavity. The anterior cavity is the space between the cornea and lens, including the iris and ciliary body. It is filled with a watery fluid called the aqueous humour. The posterior cavity is the space behind the lens that extends to the posterior side of the interior eyeball, where the retina is located. The posterior cavity is filled with a more viscous fluid called the vitreous humour.
The retina is composed of several layers and contains specialised cells for the initial processing of visual stimuli. The photoreceptors (rods and cones) change their membrane potential when stimulated by light energy. The change in membrane potential alters the amount of neurotransmitter that the photoreceptor cells release onto bipolar cells in the outer synaptic layer. It is the bipolar cell in the retina that connects a photoreceptor to a retinal ganglion cell (RGC) in the inner synaptic layer. There, amacrine cells additionally contribute to retinal processing before an action potential is produced by the RGC. The axons of RGCs, which lie at the innermost layer of the retina, collect at the optic disc and leave the eye as the optic nerve (see Figure 13.10.14). Because these axons pass through the retina, there are no photoreceptors at the very back of the eye, where the optic nerve begins. This creates a “blind spot” in the retina, and a corresponding blind spot in our visual field.
Figure 13.10.14. Structure of the eye. The sphere of the eye can be divided into anterior and posterior chambers. The wall of the eye is composed of three layers: the fibrous tunic, vascular tunic, and neural tunic. Within the neural tunic is the retina, with three layers of cells and two synaptic layers in between. The centre of the retina has a small indentation known as the fovea.
Note that the photoreceptors in the retina (rods and cones) are located behind the axons, RGCs, bipolar cells, and retinal blood vessels. A significant amount of light is absorbed by these structures before the light reaches the photoreceptor cells. However, at the exact centre of the retina is a small area known as the fovea. At the fovea, the retina lacks the supporting cells and blood vessels, and only contains photoreceptors. Therefore, visual acuity, or the sharpness of vision, is greatest at the fovea. This is because the fovea is where the least amount of incoming light is absorbed by other retinal structures (see Figure 13.10.14). As one moves in either direction from this central point of the retina, visual acuity drops significantly. In addition, each photoreceptor cell of the fovea is connected to a single RGC. Therefore, this RGC does not have to integrate inputs from multiple photoreceptors, which reduces the accuracy of visual transduction. Toward the edges of the retina, several photoreceptors converge on RGCs (through the bipolar cells) up to a ratio of 50 to 1. The difference in visual acuity between the fovea and peripheral retina is easily evidenced by looking directly at a word in the middle of this paragraph. The visual stimulus in the middle of the field of view falls on the fovea and is in the sharpest focus. Without moving your eyes off that word, notice that words at the beginning or end of the paragraph are not in focus. The images in your peripheral vision are focused by the peripheral retina, and have vague, blurry edges and words that are not as clearly identified. As a result, a large part of the neural function of the eyes is concerned with moving the eyes and head so that important visual stimuli are centred on the fovea.
Light falling on the retina causes chemical changes to pigment molecules in the photoreceptors, ultimately leading to a change in the activity of the RGCs. Photoreceptor cells have two parts, the inner segment and the outer segment (Figure 13.10.15). The inner segment contains the nucleus and other common organelles of a cell, whereas the outer segment is a specialised region in which photoreception takes place. There are two types of photoreceptors—rods and cones—which differ in the shape of their outer segment. The rod-shaped outer segments of the rod photoreceptor contain a stack of membrane-bound discs that contain the photosensitive pigment rhodopsin. The cone-shaped outer segments of the cone photoreceptor contain their photosensitive pigments in infoldings of the cell membrane. There are three cone photopigments, called opsins, which are each sensitive to a particular wavelength of light. The wavelength of visible light determines its colour. The pigments in human eyes are specialised in perceiving three different primary colours: red, green, and blue.
At the molecular level, visual stimuli cause changes in the photopigment molecule that lead to changes in membrane potential of the photoreceptor cell. A single unit of light is called a photon, which is described in physics as a packet of energy with properties of both a particle and a wave. The energy of a photon is represented by its wavelength, with each wavelength of visible light corresponding to a particular colour. Visible light is electromagnetic radiation with a wavelength between 380 and 720 nm. Wavelengths of electromagnetic radiation longer than 720 nm fall into the infrared range, whereas wavelengths shorter than 380 nm fall into the ultraviolet range. Light with a wavelength of 380 nm is blue whereas light with a wavelength of 720 nm is dark red. All other colours fall between red and blue at various points along the wavelength scale.
Opsin pigments are actually transmembrane proteins that contain a cofactor known as retinal. Retinal is a hydrocarbon molecule related to vitamin A. When a photon hits retinal, the long hydrocarbon chain of the molecule is biochemically altered. Specifically, photons cause some of the double-bonded carbons within the chain to switch from a cis to a trans conformation. This process is called photoisomerisation. Before interacting with a photon, retinal’s flexible double-bonded carbons are in the cis conformation. This molecule is referred to as 11-cis-retinal. A photon interacting with the molecule causes the flexible double-bonded carbons to change to the trans– conformation, forming all-trans-retinal, which has a straight hydrocarbon chain (Figure 13.10.16).
The shape change of retinal in the photoreceptors initiates visual transduction in the retina. Activation of retinal and the opsin proteins result in activation of a G protein. The G protein changes the membrane potential of the photoreceptor cell, which then releases less neurotransmitter into the outer synaptic layer of the retina. Until the retinal molecule is changed back to the 11-cis-retinal shape, the opsin cannot respond to light energy, which is called bleaching. When a large group of photopigments is bleached, the retina will send information as if opposing visual information is being perceived. After a bright flash of light, afterimages are usually seen in negative. The photoisomerisation is reversed by a series of enzymatic changes so that the retinal responds to more light energy.
Figure 13.10.16. Retinal isomers. The retinal molecule has two isomers, (a) one before a photon interacts with it and (b) one that is altered through photoisomerisation.
The opsins are sensitive to limited wavelengths of light. Rhodopsin, the photopigment in rods, is most sensitive to light at a wavelength of 498 nm. The three colour opsins have peak sensitivities of 564 nm, 534 nm, and 420 nm corresponding roughly to the primary colours of red, green, and blue (Figure 13.10.17). The absorbance of rhodopsin in the rods is much more sensitive than in the cone opsins; specifically, rods are sensitive to vision in low light conditions, and cones are sensitive to brighter conditions. In normal sunlight, rhodopsin will be constantly bleached while the cones are active. In a darkened room, there is not enough light to activate cone opsins, and vision is entirely dependent on rods. Rods are so sensitive to light that a single photon can result in an action potential from a rod’s corresponding RGC.
The three types of cone opsins, being sensitive to different wavelengths of light, provide us with colour vision. By comparing the activity of the three different cones, the brain can extract colour information from visual stimuli. A bright blue light, for example, that has a wavelength of approximately 450 nm would activate the “red” cones minimally, the “green” cones marginally, and the “blue” cones predominantly. The relative activation of the three different cones is calculated by the brain, which perceives the colour as blue. However, cones cannot react to low-intensity light, and rods do not sense the colour of light. Therefore, our low-light vision is—in essence—in greyscale. In other words, in a dark room, everything appears as a shade of grey. If you think that you can see colours in the dark, it is because your brain knows what colour something is and is relying on that memory.
Figure 13.10.17. Comparison of colour sensitivity of photopigments. Comparing the peak sensitivity and absorbance spectra of the four photopigments suggests that they are most sensitive to particular wavelengths.
Sensory Nerves
Once any sensory cell transduces a stimulus into a nerve impulse, that impulse must travel along axons to reach the CNS. In many of the special senses, the axons leaving the sensory receptors have a topographical arrangement, meaning that the location of the sensory receptor relates to the location of the axon in the nerve. In the retina, for example, axons from RGCs in the fovea are located at the centre of the optic nerve, where they are surrounded by axons from the more peripheral RGCs.
Spinal Nerves
Generally, spinal nerves contain afferent axons from sensory receptors in the periphery, such as from the skin, mixed with efferent axons travelling to the muscles or other effector organs. As the spinal nerve nears the spinal cord, it splits into dorsal and ventral roots. The dorsal root contains only the axons of sensory neurons, whereas the ventral roots contain only the axons of the motor neurons. Some of the branches will synapse with local neurons in the dorsal root ganglion, posterior (dorsal) horn, or even the anterior (ventral) horn, at the level of the spinal cord where they enter. Other branches will travel a short distance up or down the spine to interact with neurons at other levels of the spinal cord. A branch may also turn into the posterior (dorsal) column of the white matter to connect with the brain. For the sake of convenience, we will use the terms ventral and dorsal in reference to structures within the spinal cord that are part of these pathways. This will help to underscore the relationships between the different components. Typically, spinal nerve systems that connect to the brain are contralateral, in that the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain.
Cranial Nerves
Cranial nerves convey specific sensory information from the head and neck directly to the brain. For sensations below the neck, the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain. Whereas spinal information is contralateral, cranial nerve systems are mostly ipsilateral, meaning that a cranial nerve on the right side of the head is connected to the right side of the brain. Some cranial nerves contain only sensory axons, such as the olfactory, optic, and vestibulocochlear nerves. Other cranial nerves contain both sensory and motor axons, including the trigeminal, facial, glossopharyngeal, and vagus nerves (however, the vagus nerve is not associated with the somatic nervous system). The general senses of somatosensation for the face travel through the trigeminal system.
Section Review
The senses are olfaction (smell), gustation (taste), somatosensation (sensations associated with the skin and body), audition (hearing), equilibrium (balance), and vision. Except for somatosensation, this list represents the special senses, or those systems of the body that are associated with specific organs such as the tongue or eye. Somatosensation belongs to the general senses, which are those sensory structures that are distributed throughout the body and in the walls of various organs. The special senses are all primarily part of the somatic nervous system in that they are consciously perceived through cerebral processes, though some special senses contribute to autonomic function. The general senses can be divided into somatosensation, which is commonly considered touch, but includes tactile, pressure, vibration, temperature, and pain perception. The general senses also include the visceral senses, which are separate from the somatic nervous system function in that they do not normally rise to the level of conscious perception.
The cells that transduce sensory stimuli into the electrochemical signals of the nervous system are classified based on structural or functional aspects of the cells. The structural classifications are either based on the anatomy of the cell that is interacting with the stimulus (free nerve endings, encapsulated endings, or specialised receptor cell), or where the cell is located relative to the stimulus (interoceptor, exteroceptor, proprioceptor). Thirdly, the functional classification is based on how the cell transduces the stimulus into a neural signal. Chemoreceptors respond to chemical stimuli and are the basis for olfaction and gustation. Related to chemoreceptors are osmoreceptors and nociceptors for fluid balance and pain reception, respectively. Mechanoreceptors respond to mechanical stimuli and are the basis for most aspects of somatosensation, as well as being the basis of audition and equilibrium in the inner ear. Thermoreceptors are sensitive to temperature changes, and photoreceptors are sensitive to light energy.
The nerves that convey sensory information from the periphery to the CNS are either spinal nerves, connected to the spinal cord, or cranial nerves, connected to the brain. Spinal nerves have mixed populations of fibres; some are motor fibres and some are sensory. The sensory fibres connect to the spinal cord through the dorsal root, which is attached to the dorsal root ganglion. Sensory information from the body that is conveyed through spinal nerves will project to the opposite side of the brain to be processed by the cerebral cortex. The cranial nerves can be strictly sensory fibres, such as the olfactory, optic, and vestibulocochlear nerves, or mixed sensory and motor nerves, such as the trigeminal, facial, glossopharyngeal, and vagus nerves. The cranial nerves are connected to the same side of the brain from which the sensory information originates.
Describe the pathways that sensory systems follow into the central nervous system
Differentiate between the two major ascending pathways in the spinal cord
Describe the pathway of somatosensory input from the face and compare it to the ascending pathways in the spinal cord
Explain topographical representations of sensory information in at least two systems
Describe two pathways of visual processing and the functions associated with each
Sensory Pathways
Specific regions of the CNS coordinate different somatic processes using sensory inputs and motor outputs of peripheral nerves. A simple case is a reflex caused by a synapse between a dorsal sensory neuron axon and a motor neuron in the ventral horn. More complex arrangements are possible to integrate peripheral sensory information with higher processes. The important regions of the CNS that play a role in somatic processes can be separated into the spinal cord brain stem, diencephalon, cerebral cortex, and subcortical structures.
Spinal Cord and Brain Stem
A sensory pathway that carries peripheral sensations to the brain is referred to as an ascending pathway, or ascending tract. The various sensory modalities each follow specific pathways through the CNS. Tactile and other somatosensory stimuli activate receptors in the skin, muscles, tendons, and joints throughout the entire body. However, the somatosensory pathways are divided into two separate systems on the basis of the location of the receptor neurons. Somatosensory stimuli from below the neck pass along the sensory pathways of the spinal cord, whereas somatosensory stimuli from the head and neck travel through the cranial nerves—specifically, the trigeminal system.
The dorsal column system (sometimes referred to as the dorsal column–medial lemniscus) and the spinothalamic tract are two major pathways that bring sensory information to the brain (Figure 13.11.1). The sensory pathways in each of these systems are composed of three successive neurons.
The dorsal column system begins with the axon of a dorsal root ganglion neuron entering the dorsal root and joining the dorsal column white matter in the spinal cord. As axons of this pathway enter the dorsal column, they take on a positional arrangement so that axons from lower levels of the body position themselves medially, whereas axons from upper levels of the body position themselves laterally. The dorsal column is separated into two component tracts, the fasciculus gracilis that contains axons from the legs and lower body, and the fasciculus cuneatus that contains axons from the upper body and arms.
The axons in the dorsal column terminate in the nuclei of the medulla, where each synapses with the second neuron in their respective pathway. The nucleus gracilis is the target of fibres in the fasciculus gracilis, whereas the nucleus cuneatus is the target of fibres in the fasciculus cuneatus. The second neuron in the system projects from one of the two nuclei and then decussates, or crosses the midline of the medulla. These axons then continue to ascend the brain stem as a bundle called the medial lemniscus. These axons terminate in the thalamus, where each synapses with the third neuron in their respective pathway. The third neuron in the system projects its axons to the postcentral gyrus of the cerebral cortex, where somatosensory stimuli are initially processed and the conscious perception of the stimulus occurs.
The spinothalamic tract also begins with neurons in a dorsal root ganglion. These neurons extend their axons to the dorsal horn, where they synapse with the second neuron in their respective pathway. The name “spinothalamic” comes from this second neuron, which has its cell body in the spinal cord grey matter and connects to the thalamus. Axons from these second neurons then decussate within the spinal cord and ascend to the brain and enter the thalamus, where each synapses with the third neuron in its respective pathway. The neurons in the thalamus then project their axons to the spinothalamic tract, which synapses in the postcentral gyrus of the cerebral cortex.
These two systems are similar in that they both begin with dorsal root ganglion cells, as with most general sensory information. The dorsal column system is primarily responsible for touch sensations and proprioception, whereas the spinothalamic tract pathway is primarily responsible for pain and temperature sensations. Another similarity is that the second neurons in both of these pathways are contralateral, because they project across the midline to the other side of the brain or spinal cord. In the dorsal column system, this decussation takes place in the brain stem; in the spinothalamic pathway, it takes place in the spinal cord at the same spinal cord level at which the information entered. The third neurons in the two pathways are the same. In both, the second neuron synapses in the thalamus, and the thalamic neuron projects to the somatosensory cortex.
Figure 13.11.1. Ascending sensory pathways of the spinal cord. The dorsal column system and spinothalamic tract are the major ascending pathways that connect the periphery with the brain.
The trigeminal pathway carries somatosensory information from the face, head, mouth, and nasal cavity. As with the previously discussed nerve tracts, the sensory pathways of the trigeminal pathway each involve three successive neurons. First, axons from the trigeminal ganglion enter the brain stem at the level of the pons. These axons project to one of three locations. The spinal trigeminal nucleus of the medulla receives information similar to that carried by spinothalamic tract, such as pain and temperature sensations. Other axons go to either the chief sensory nucleus in the pons or the mesencephalic nuclei in the midbrain. These nuclei receive information like that carried by the dorsal column system, such as touch, pressure, vibration, and proprioception. Axons from the second neuron decussate and ascend to the thalamus along the trigeminothalamic tract. In the thalamus, each axon synapses with the third neuron in its respective pathway. Axons from the third neuron then project from the thalamus to the primary somatosensory cortex of the cerebrum.
The sensory pathway for gustation travels along the facial and glossopharyngeal cranial nerves, which synapse with neurons of the solitary nucleus in the brain stem. Axons from the solitary nucleus then project to the ventral posterior nucleus of the thalamus. Finally, axons from the ventral posterior nucleus project to the gustatory cortex of the cerebral cortex, where taste is processed and consciously perceived.
The sensory pathway for audition travels along the vestibulocochlear nerve, which synapses with neurons in the cochlear nuclei of the superior medulla. Within the brain stem, input from either ear is combined to extract location information from the auditory stimuli. Whereas the initial auditory stimuli received at the cochlea strictly represent the frequency—or pitch—of the stimuli, the locations of sounds can be determined by comparing information arriving at both ears.
Sound localisation is a feature of central processing in the auditory nuclei of the brain stem. Sound localisation is achieved by the brain calculating the interaural time difference and the interaural intensity difference. A sound originating from a specific location will arrive at each ear at different times, unless the sound is directly in front of the listener. If the sound source is slightly to the left of the listener, the sound will arrive at the left ear microseconds before it arrives at the right ear (Figure 13.11.2). This time difference is an example of an interaural time difference. Also, the sound will be slightly louder in the left ear than in the right ear because some of the sound waves reaching the opposite ear are blocked by the head. This is an example of an interaural intensity difference.
Figure 13.11.2. Auditory brain stem mechanisms of sound localisation. Localising sound in the horizontal plane is achieved by processing in the medullary nuclei of the auditory system. Connections between neurons on either side are able to compare very slight differences in sound stimuli that arrive at either ear and represent interaural time and intensity differences.
Auditory processing continues on to a nucleus in the midbrain called the inferior colliculus. Axons from the inferior colliculus project to two locations, the thalamus and the superior colliculus. The medial geniculate nucleus of the thalamus receives the auditory information and then projects that information to the auditory cortex in the temporal lobe of the cerebral cortex. The superior colliculus receives input from the visual and somatosensory systems, as well as the ears, to initiate stimulation of the muscles that turn the head and neck toward the auditory stimulus.
Balance is coordinated through the vestibular system, the nerves of which are composed of axons from the vestibular ganglion that carries information from the utricle, saccule, and semicircular canals. The system contributes to controlling head and neck movements in response to vestibular signals. An important function of the vestibular system is coordinating eye and head movements to maintain visual attention. Most of the axons terminate in the vestibular nuclei of the medulla. Some axons project from the vestibular ganglion directly to the cerebellum, with no intervening synapse in the vestibular nuclei. The cerebellum is primarily responsible for initiating movements on the basis of equilibrium information.
Neurons in the vestibular nuclei project their axons to targets in the brain stem. One target is the reticular formation, which influences respiratory and cardiovascular functions in relation to body movements. A second target of the axons of neurons in the vestibular nuclei is the spinal cord, which initiates the spinal reflexes involved with posture and balance. To assist the visual system, fibres of the vestibular nuclei project to the oculomotor, trochlear, and abducens nuclei to influence signals sent along the cranial nerves. These connections constitute the pathway of the vestibulo-ocular reflex (VOR), which compensates for head and body movement by stabilising images on the retina (Figure 13.11.3). Finally, the vestibular nuclei project to the thalamus to join the proprioceptive pathway of the dorsal column system, allowing conscious perception of equilibrium.
Figure 13.11.3. Vestibulo-ocular reflex. Connections between the vestibular system and the cranial nerves controlling eye movement keep the eyes centred on a visual stimulus, even though the head is moving. During head movement, the eye muscles move the eyes in the opposite direction as the head movement, keeping the visual stimulus centred in the field of view.
The connections of the optic nerve are more complicated than those of other cranial nerves. Instead of the connections being between each eye and the brain, visual information is segregated between the left and right sides of the visual field. In addition, some of the information from one side of the visual field projects to the opposite side of the brain. Within each eye, the axons projecting from the medial side of the retina decussate at the optic chiasm, for example, the axons from the medial retina of the left eye cross over to the right side of the brain at the optic chiasm. However, within each eye, the axons projecting from the lateral side of the retina do not decussate. The axons from the lateral retina of the right eye project back to the right side of the brain. Therefore, the left field of view of each eye is processed on the right side of the brain, whereas the right field of view of each eye is processed on the left side of the brain (Figure 13.11.4).
Figure 13.11.4. Segregation of visual field information at the optic chiasm. Contralateral visual field information from the lateral retina projects to the ipsilateral brain, whereas ipsilateral visual field information must decussate at the optic chiasm to reach the opposite side of the brain.
A unique clinical presentation that relates to this anatomic arrangement is the loss of lateral peripheral vision, known as bilateral hemianopia. This is different from “tunnel vision” because the superior and inferior peripheral fields are not lost. Visual field deficits can be disturbing for a patient, but in this case, the cause is not within the visual system itself. A growth of the pituitary gland presses against the optic chiasm and interferes with signal transmission. However, the axons projecting to the same side of the brain are unaffected. Therefore, the patient loses the outermost areas of their field of vision and cannot see objects to their right and left.
Extending from the optic chiasm, the axons of the visual system are referred to as the optic tract instead of the optic nerve. The optic tract has three major targets, two in the diencephalon and one in the midbrain. The connection between the eyes and diencephalon is demonstrated during development, in which the neural tissue of the retina differentiates from that of the diencephalon by the growth of the secondary vesicles. The connections of the retina into the CNS are a holdover from this developmental association. Most of the connections of the optic tract are to the thalamus—specifically, the lateral geniculate nucleus. Axons from this nucleus then project to the visual cortex of the cerebrum, located in the occipital lobe. Another target of the optic tract is the superior colliculus.
In addition, a very small number of RGC axons project from the optic chiasm to the suprachiasmatic nucleus of the hypothalamus. These RGCs are photosensitive, in that they respond to the presence or absence of light. Unlike the photoreceptors, however, these photosensitive RGCs cannot be used to perceive images. By simply responding to the absence or presence of light, these RGCs can send information about day length. The perceived proportion of sunlight to darkness establishes the circadian rhythm of our bodies, allowing certain physiological events to occur at approximately the same time every day.
Diencephalon
The diencephalon is beneath the cerebrum and includes the thalamus and hypothalamus. In the somatic nervous system, the thalamus is an important relay for communication between the cerebrum and the rest of the nervous system. The hypothalamus has both somatic and autonomic functions. In addition, the hypothalamus communicates with the limbic system, which controls emotions and memory functions.
Sensory input to the thalamus comes from most of the special senses and ascending somatosensory tracts. Each sensory system is relayed through a particular nucleus in the thalamus. The thalamus is a required transfer point for most sensory tracts that reach the cerebral cortex, where conscious sensory perception begins. The one exception to this rule is the olfactory system. The olfactory tract axons from the olfactory bulb project directly to the cerebral cortex, along with the limbic system and hypothalamus.
The thalamus is a collection of several nuclei that can be categorised into three anatomical groups. White matter running through the thalamus defines the three major regions of the thalamus, which are an anterior nucleus, a medial nucleus, and a lateral group of nuclei. The anterior nucleus serves as a relay between the hypothalamus and the emotion and memory-producing limbic system. The medial nuclei serve as a relay for information from the limbic system and basal ganglia to the cerebral cortex. This allows memory creation during learning, but also determines alertness. The special and somatic senses connect to the lateral nuclei, where their information is relayed to the appropriate sensory cortex of the cerebrum.
Cortical Processing
As described earlier, many of the sensory axons are positioned in the same way as their corresponding receptor cells in the body. This allows identification of the position of a stimulus on the basis of which receptor cells are sending information. The cerebral cortex also maintains this sensory topography in the particular areas of the cortex that correspond to the position of the receptor cells. The somatosensory cortex provides an example in which, in essence, the locations of the somatosensory receptors in the body are mapped onto the somatosensory cortex. This mapping is often depicted using a sensory homunculus (Figure 13.11.5).
The term homunculus comes from the Latin word for “little man” and refers to a map of the human body that is laid across a portion of the cerebral cortex. In the somatosensory cortex, the external genitals, feet, and lower legs are represented on the medial face of the gyrus within the longitudinal fissure. As the gyrus curves out of the fissure and along the surface of the parietal lobe, the body map continues through the thighs, hips, trunk, shoulders, arms, and hands. The head and face are just lateral to the fingers as the gyrus approaches the lateral sulcus. The representation of the body in this topographical map is medial to lateral from the lower to upper body. It is a continuation of the topographical arrangement seen in the dorsal column system, where axons from the lower body are carried in the fasciculus gracilis, whereas axons from the upper body are carried in the fasciculus cuneatus. As the dorsal column system continues into the medial lemniscus, these relationships are maintained. Also, the head and neck axons running from the trigeminal nuclei to the thalamus run adjacent to the upper body fibres. The connections through the thalamus maintain topography such that the anatomic information is preserved. Note that this correspondence does not result in a perfectly miniature scale version of the body, but exaggerates the more sensitive areas of the body, such as the fingers and lower face. Less sensitive areas of the body, such as the shoulders and back, are mapped to smaller areas on the cortex.
Figure 13.11.5. The Sensory homunculus. A cartoon representation of the sensory homunculus arranged adjacent to the cortical region in which the processing takes place.
Likewise, the topographic relationship between the retina and the visual cortex is maintained throughout the visual pathway. The visual field is projected onto the two retinae, as described above, with sorting at the optic chiasm. The right peripheral visual field falls on the medial portion of the right retina and the lateral portion of the left retina. The right medial retina then projects across the midline through the optic chiasm. This results in the right visual field being processed in the left visual cortex. Likewise, the left visual field is processed in the right visual cortex (see Figure 13.11.4). Though the chiasm is helping to sort right and left visual information, superior and inferior visual information is maintained topographically in the visual pathway. Light from the superior visual field falls on the inferior retina, and light from the inferior visual field falls on the superior retina. This topography is maintained such that the superior region of the visual cortex processes the inferior visual field and vice versa. Therefore, the visual field information is inverted and reversed as it enters the visual cortex—up is down, and left is right. However, the cortex processes the visual information such that the final conscious perception of the visual field is correct. The topographic relationship is evident in that information from the foveal region of the retina is processed in the centre of the primary visual cortex. Information from the peripheral regions of the retina is correspondingly processed toward the edges of the visual cortex. Similar to the exaggerations in the sensory homunculus of the somatosensory cortex, the foveal-processing area of the visual cortex is disproportionately larger than the areas processing peripheral vision.
In an experiment performed in the 1960s, subjects wore prism glasses so that the visual field was inverted before reaching the eye. On the first day of the experiment, subjects would duck when walking up to a table, thinking it was suspended from the ceiling. However, after a few days of acclimation, the subjects behaved as if everything were represented correctly. Therefore, the visual cortex is somewhat flexible in adapting to the information it receives from our eyes (Figure 13.11.6).
Figure 13.11.6. Topographic mapping of the retina onto the visual cortex. The visual field projects onto the retina through the lenses and falls on the retinae as an inverted, reversed image. The topography of this image is maintained as the visual information travels through the visual pathway to the cortex.
The cortex has been described as having specific regions that are responsible for processing specific information; there is the visual cortex, somatosensory cortex, gustatory cortex, etc. However, our experience of these senses is not divided. Instead, we experience what can be referred to as a seamless percept. Our perceptions of the various sensory modalities—though distinct in their content—are integrated by the brain so that we experience the world as a continuous whole.
In the cerebral cortex, sensory processing begins at the primary sensory cortex, then proceeds to an association area, and finally, into a multimodal integration area. The visual pathway, for example, projects from the retinae through the thalamus to the primary visual cortex in the occipital lobe. This area is primarily in the medial wall within the longitudinal fissure. Here, visual stimuli begin to be recognised as basic shapes. Edges of objects are recognised and built into more complex shapes. Also, inputs from both eyes are compared to extract depth information. Because of the overlapping field of view between the two eyes, the brain can begin to estimate the distance of stimuli based on binocular depth cues.
Everyday Connections
Depth Perception, 3-D Movies, and Optical Illusions
The visual field is projected onto the retinal surface, where photoreceptors transduce light energy into neural signals for the brain to interpret. The retina is a two-dimensional surface, so it does not encode three-dimensional information. However, we can perceive depth. How is that accomplished?
Two ways in which we can extract depth information from the two-dimensional retinal signal are based on monocular cues and binocular cues, respectively. Monocular depth cues are those that are the result of information within the two-dimensional visual field. One object that overlaps another object must be in front. Relative size differences are also a cue, for example, if a basketball appears larger than the basket, then the basket must be further away. Based on experience, we can estimate how far away the basket is. Binocular depth cues compare information represented in the two retinae because they do not see the visual field exactly the same.
The centres of the two eyes are separated by a small distance, which is approximately 6 to 6.5 cm in most people. Because of this offset, visual stimuli do not fall on exactly the same spot on both retinae unless we are fixated directly on them and they fall on the fovea of each retina. All other objects in the visual field, either closer or farther away than the fixated object, will fall on different spots on the retina. When vision is fixed on an object in space, closer objects will fall on the lateral retina of each eye, and more distant objects will fall on the medial retina of either eye (Figure 13.11.7). This is easily observed by holding a finger up in front of your face as you look at a more distant object. You will see two images of your finger that represent the two disparate images that are falling on either retina.
These depth cues, both monocular and binocular, can be exploited to make the brain think there are three dimensions in two-dimensional information. This is the basis of 3-D movies. The projected image on the screen is two dimensional, but it has disparate information embedded in it. The 3-D glasses that are available at the theatre filter the information so that only one eye sees one version of what is on the screen, and the other eye sees the other version. If you take the glasses off, the image on the screen will have varying amounts of blur because both eyes are seeing both layers of information, and the third dimension will not be evident. Some optical illusions can take advantage of depth cues as well, though those are more often using monocular cues to fool the brain into seeing different parts of the scene as being at different depths.
Figure 13.11.7. Retinal disparity. Because of the interocular distance, which results in objects of different distances falling on different spots of the two retinae, the brain can extract depth perception from the two-dimensional information of the visual field.
There are two main regions that surround the primary cortex that are usually referred to as areas V2 and V3 (the primary visual cortex is area V1). These surrounding areas are the visual association cortex. The visual association regions develop more complex visual perceptions by adding colour and motion information. The information processed in these areas is then sent to regions of the temporal and parietal lobes. Visual processing has two separate streams of processing: one into the temporal lobe and one into the parietal lobe. These are the ventral and dorsal streams, respectively (Figure 13.11.8). The ventral stream identifies visual stimuli and their significance. Because the ventral stream uses temporal lobe structures, it begins to interact with the non-visual cortex and may be important in visual stimuli becoming part of memories. The dorsal stream locates objects in space and helps in guiding movements of the body in response to visual inputs. The dorsal stream enters the parietal lobe, where it interacts with somatosensory cortical areas that are important for our perception of the body and its movements. The dorsal stream can then influence frontal lobe activity where motor functions originate.
Figure 13.11.8. Ventral and dorsal visual streams. From the primary visual cortex in the occipital lobe, visual processing continues in two streams—one into the temporal lobe and one into the parietal lobe.
Disorders of the Brain: Prosopagnosia
The failures of sensory perception can be unusual and debilitating. A particular sensory deficit that inhibits an important social function of humans is prosopagnosia, or face blindness. The word comes from the Greek words prosopa, that means “faces,” and agnosia, that means “not knowing.” Some people may feel that they cannot recognise people easily by their faces. However, a person with prosopagnosia cannot recognise the most recognisable people in their respective cultures. They would not recognise the face of a celebrity, an important historical figure, or even a family member like their mother. They may not even recognise their own face.
Prosopagnosia can be caused by trauma to the brain, or it can be present from birth. The exact cause of proposagnosia and the reason that it happens to some people is unclear. A study of the brains of people born with the deficit found that a specific region of the brain, the anterior fusiform gyrus of the temporal lobe, is often underdeveloped. This region of the brain is concerned with the recognition of visual stimuli and its association with memories. Though the evidence is not yet definitive, this region is likely to be where facial recognition occurs.
Though this can be a devastating condition, people who suffer from it can get by—often by using other cues to recognise the people they see. Often, the sound of a person’s voice, or the presence of unique cues such as distinct facial features (a mole, for example) or hair colour can help the sufferer recognise a familiar person.
Section Review
Sensory input to the brain enters through pathways that travel through either the spinal cord (for somatosensory input from the body) or the brain stem (for everything else, except the visual and olfactory systems) to reach the diencephalon. In the diencephalon, sensory pathways reach the thalamus. This is necessary for all sensory systems to reach the cerebral cortex, except for the olfactory system that is directly connected to the frontal and temporal lobes.
The two major tracts in the spinal cord, originating from sensory neurons in the dorsal root ganglia, are the dorsal column system and the spinothalamic tract. The major differences between the two are in the type of information that is relayed to the brain and where the tracts decussate. The dorsal column system primarily carries information about touch and proprioception and crosses the midline in the medulla. The spinothalamic tract is primarily responsible for pain and temperature sensation and crosses the midline in the spinal cord at the level at which it enters. The trigeminal nerve adds similar sensation information from the head to these pathways.
The auditory pathway passes through multiple nuclei in the brain stem in which additional information is extracted from the basic frequency stimuli processed by the cochlea. Sound localisation is made possible through the activity of these brain stem structures. The vestibular system enters the brain stem and influences activity in the cerebellum, spinal cord, and cerebral cortex.
The visual pathway segregates information from the two eyes so that one half of the visual field projects to the other side of the brain. Within visual cortical areas, the perception of the stimuli and their location is passed along two streams, one ventral and one dorsal. The ventral visual stream connects to structures in the temporal lobe that are important for long-term memory formation. The dorsal visual stream interacts with the somatosensory cortex in the parietal lobe, and together they can influence the activity in the frontal lobe to generate movements of the body in relation to visual information.
List the components of the basic processing stream for the motor system
Describe the pathway of descending motor commands from the cortex to the skeletal muscles
Compare different descending pathways, both by structure and function
Explain the initiation of movement from the neurological connections
Describe several reflex arcs and their functional roles
The defining characteristic of the somatic nervous system is that it controls skeletal muscles. Somatic senses inform the nervous system about the external environment, but the response to that is through voluntary muscle movement. The term “voluntary” suggests that there is a conscious decision to make a movement. However, some aspects of the somatic system use voluntary muscles without conscious control. One example is the ability of our breathing to switch to unconscious control while we are focused on another task. However, the muscles that are responsible for the basic process of breathing are also utilised for speech, which is entirely voluntary.
Cortical Responses
Let’s start with sensory stimuli that have been registered through receptor cells and the information relayed to the CNS along ascending pathways. In the cerebral cortex, the initial processing of sensory perception progresses to associative processing and then integration in multimodal areas of cortex. These levels of processing can lead to the incorporation of sensory perceptions into memory, but more importantly, they lead to a response. The completion of cortical processing through the primary, associative, and integrative sensory areas initiates a similar progression of motor processing, usually in different cortical areas.
Whereas the sensory cortical areas are located in the occipital, temporal, and parietal lobes, motor functions are controlled by the frontal lobe. The most anterior regions of the frontal lobe—the prefrontal areas—are important for executive functions, which are those cognitive functions that lead to goal-directed behaviours. These higher cognitive processes include working memory, which has been called a “mental scratch pad,” that can help organise and represent information that is not in the immediate environment. The prefrontal lobe is responsible for aspects of attention, such as inhibiting distracting thoughts and actions so that a person can focus on a goal and direct behaviour toward achieving that goal.
The functions of the prefrontal cortex are integral to the personality of an individual because it is responsible for what a person intends to do and how they accomplish those plans. A famous case of damage to the prefrontal cortex is that of Phineas Gage, dating back to 1848. He was a railroad worker who had a metal spike impale his prefrontal cortex (Figure 13.12.1). He survived the accident, but according to second-hand accounts, his personality changed drastically. Friends described him as no longer acting like himself. Whereas he was a hardworking, amiable man before the accident, he turned into an irritable, temperamental, and lazy man after the accident. Many of the accounts of his change may have been inflated in the retelling, and some behaviour was attributable to alcohol used as a pain medication. However, the accounts suggest that some aspects of his personality did change. Also, there is new evidence that though his life changed dramatically, he was able to become a functioning stagecoach driver, suggesting that the brain can recover even from major trauma such as this.
Phineas Gage
Figure 13.12.1. Phineas Gage. The victim of an accident while working on a railroad in 1848, Phineas Gage had a large iron rod impaled through the prefrontal cortex of his frontal lobe. After the accident, his personality appeared to change, but he eventually learned to cope with the trauma and lived as a coach driver even after such a traumatic event. (credit b: John M. Harlow, MD).
The victim of an accident while working on a railroad in 1848, Phineas Gage had a large iron rod impaled through the prefrontal cortex of his frontal lobe. After the accident, his personality appeared to change, but he eventually learned to cope with the trauma and lived as a coach driver even after such a traumatic event.
Secondary Motor Cortices
In generating motor responses, the executive functions of the prefrontal cortex will need to initiate actual movements. One way to define the prefrontal area is any region of the frontal lobe that does not elicit movement when electrically stimulated. These are primarily in the anterior part of the frontal lobe. The regions of the frontal lobe that remain are the regions of the cortex that produce movement. The prefrontal areas project into the secondary motor cortices, which include the premotor cortex and the supplemental motor area.
Two important regions that assist in planning and coordinating movements are located adjacent to the primary motor cortex. The premotor cortex is more lateral, whereas the supplemental motor area is more medial and superior. The premotor area aids in controlling movements of the core muscles to maintain posture during movement, whereas the supplemental motor area is hypothesised to be responsible for planning and coordinating movement. The supplemental motor area also manages sequential movements that are based on prior experience (that is, learned movements). Neurons in these areas are most active leading up to the initiation of movement, for example, these areas might prepare the body for the movements necessary to drive a car in anticipation of a traffic light changing.
Adjacent to these two regions are two specialised motor planning centres. The frontal eye fields are responsible for moving the eyes in response to visual stimuli. There are direct connections between the frontal eye fields and the superior colliculus. Also, anterior to the premotor cortex and primary motor cortex is Broca’s area. This area is responsible for controlling movements of the structures of speech production. The area is named after a French surgeon and anatomist who studied patients who could not produce speech. They did not have impairments to understanding speech, only to producing speech sounds, suggesting a damaged or underdeveloped Broca’s area.
Primary Motor Cortex
The primary motor cortex is located in the precentral gyrus of the frontal lobe. A neurosurgeon, Walter Penfield, described much of the basic understanding of the primary motor cortex by electrically stimulating the surface of the cerebrum. Penfield would probe the surface of the cortex while the patient was only under local anaesthesia so that he could observe responses to the stimulation. This led to the belief that the precentral gyrus directly stimulated muscle movement. We now know that the primary motor cortex receives input from several areas that aid in planning movement, and its principal output stimulates spinal cord neurons to stimulate skeletal muscle contraction.
The primary motor cortex is arranged in a similar fashion to the primary somatosensory cortex, in that it has a topographical map of the body, creating a motor homunculus. The neurons responsible for musculature in the feet and lower legs are in the medial wall of the precentral gyrus, with the thighs, trunk, and shoulder at the crest of the longitudinal fissure. The hand and face are in the lateral face of the gyrus. Also, the relative space allotted for the different regions is exaggerated in muscles that have greater enervation. The greatest amount of cortical space is given to muscles that perform fine, agile movements, such as the muscles of the fingers and the lower face. The “power muscles” that perform coarser movements, such as the buttock and back muscles, occupy much less space on the motor cortex.
Descending Pathways
The motor output from the cortex descends into the brain stem and to the spinal cord to control the musculature through motor neurons. Neurons located in the primary motor cortex, named Betz cells, are large cortical neurons that synapse with lower motor neurons in the spinal cord or the brain stem. The two descending pathways travelled by the axons of Betz cells are the corticospinal tract and the corticobulbar tract. Both tracts are named for their origin in the cortex and their targets—either the spinal cord or the brain stem (the term “bulbar” refers to the brain stem as the bulb, or enlargement, at the top of the spinal cord).
These two descending pathways are responsible for the conscious or voluntary movements of skeletal muscles. Any motor command from the primary motor cortex is sent down the axons of the Betz cells to activate upper motor neurons in either the cranial motor nuclei or in the ventral horn of the spinal cord. The axons of the corticobulbar tract are ipsilateral, meaning they project from the cortex to the motor nucleus on the same side of the nervous system. Conversely, the axons of the corticospinal tract are largely contralateral, meaning that they cross the midline of the brain stem or spinal cord and synapse on the opposite side of the body. Therefore, the right motor cortex of the cerebrum controls muscles on the left side of the body, and vice versa.
The corticospinal tract descends from the cortex through the deep white matter of the cerebrum. It then passes between the caudate nucleus and putamen of the basal nuclei as a bundle called the internal capsule. The tract then passes through the midbrain as the cerebral peduncles, after which it burrows through the pons. Upon entering the medulla, the tracts make up the large white matter tract referred to as the pyramids (Figure 13.12.2). The defining landmark of the medullary-spinal border is the pyramidal decussation, which is where most of the fibres in the corticospinal tract cross over to the opposite side of the brain. At this point, the tract separates into two parts, which have control over different domains of the musculature.
Figure 13.12.2. Corticospinal tract. The major descending tract that controls skeletal muscle movements is the corticospinal tract. It is composed of two neurons, the upper motor neuron and the lower motor neuron. The upper motor neuron has its cell body in the primary motor cortex of the frontal lobe and synapses on the lower motor neuron, which is in the ventral horn of the spinal cord and projects to the skeletal muscle in the periphery.
Appendicular Control
The lateral corticospinal tract is composed of the fibres that cross the midline at the pyramidal decussation (see Figure 13.12.2). The axons cross over from the anterior position of the pyramids in the medulla to the lateral column of the spinal cord. These axons are responsible for controlling appendicular muscles.
This influence over the appendicular muscles means that the lateral corticospinal tract is responsible for moving the muscles of the arms and legs. The ventral horn in both the lower cervical spinal cord and the lumbar spinal cord both have wider ventral horns, representing the greater number of muscles controlled by these motor neurons. The cervical enlargement is particularly large because there is greater control over the fine musculature of the upper limbs, particularly of the fingers. The lumbar enlargement is not as significant in appearance because there is less fine motor control of the lower limbs.
Axial Control
The anterior corticospinal tract is responsible for controlling the muscles of the body trunk (see Figure 13.12.2). These axons do not decussate in the medulla. Instead, they remain in an anterior position as they descend the brain stem and enter the spinal cord. These axons then travel to the spinal cord level at which they synapse with a lower motor neuron. Upon reaching the appropriate level, the axons decussate, entering the ventral horn on the opposite side of the spinal cord from which they entered. In the ventral horn, these axons synapse with their corresponding lower motor neurons. The lower motor neurons are located in the medial regions of the ventral horn because they control the axial muscles of the trunk.
Because movements of the body trunk involve both sides of the body, the anterior corticospinal tract is not entirely contralateral. Some collateral branches of the tract will project into the ipsilateral ventral horn to control synergistic muscles on that side of the body, or to inhibit antagonistic muscles through interneurons within the ventral horn. Through the influence of both sides of the body, the anterior corticospinal tract can coordinate postural muscles in broad movements of the body. These coordinating axons in the anterior corticospinal tract are often considered bilateral, as they are both ipsilateral and contralateral.
Extrapyramidal Controls
Other descending connections between the brain and the spinal cord are called the extrapyramidal system. The name comes from the fact that this system is outside the corticospinal pathway, which includes the pyramids in the medulla. A few pathways originating from the brain stem contribute to this system.
The tectospinal tract projects from the midbrain to the spinal cord and is important for postural movements that are driven by the superior colliculus. The name of the tract comes from an alternate name for the superior colliculus, which is the tectum. The reticulospinal tract connects the reticular system, a diffuse region of grey matter in the brain stem, with the spinal cord. This tract influences trunk and proximal limb muscles related to posture and locomotion. The reticulospinal tract also contributes to muscle tone and influences autonomic functions. The vestibulospinal tract connects the brain stem nuclei of the vestibular system with the spinal cord. This allows posture, movement, and balance to be modulated based on equilibrium information provided by the vestibular system.
The pathways of the extrapyramidal system are influenced by subcortical structures, for example, connections between the secondary motor cortices and the extrapyramidal system modulate spine and cranium movements. The basal nuclei, which are important for regulating movement initiated by the CNS, influence the extrapyramidal system as well as its thalamic feedback to the motor cortex.
The conscious movement of our muscles is more complicated than simply sending a single command from the precentral gyrus down to the proper motor neurons. During the movement of any body part, our muscles relay information back to the brain, and the brain is constantly sending “revised” instructions back to the muscles. The cerebellum is important in contributing to the motor system because it compares cerebral motor commands with proprioceptive feedback. The corticospinal fibres that project to the ventral horn of the spinal cord have branches that also synapse in the pons, which project to the cerebellum. Also, the proprioceptive sensations of the dorsal column system have a collateral projection to the medulla that projects to the cerebellum. These two streams of information are compared in the cerebellar cortex. Conflicts between the motor commands sent by the cerebrum and body position information provided by the proprioceptors cause the cerebellum to stimulate the red nucleus of the midbrain. The red nucleus then sends corrective commands to the spinal cord along the rubrospinal tract. The name of this tract comes from the word for red that is seen in the English word “ruby.”
A good example of how the cerebellum corrects cerebral motor commands can be illustrated by walking in water. An original motor command from the cerebrum to walk will result in a highly coordinated set of learned movements. However, in water, the body cannot perform a typical walking movement as instructed. The cerebellum can alter the motor command, stimulating the leg muscles to take larger steps to overcome the water resistance. The cerebellum can make the necessary changes through the rubrospinal tract. Modulating the basic command to walk also relies on spinal reflexes, but the cerebellum is responsible for calculating the appropriate response. When the cerebellum does not work properly, coordination and balance are severely affected. The most dramatic example of this is during the overconsumption of alcohol. Alcohol inhibits the ability of the cerebellum to interpret proprioceptive feedback, making it more difficult to coordinate body movements, such as walking a straight line, or guide the movement of the hand to touch the tip of the nose.
Ventral Horn Output
The somatic nervous system provides output strictly to skeletal muscles. The lower motor neurons, which are responsible for the contraction of these muscles, are found in the ventral horn of the spinal cord. These large, multipolar neurons have a corona of dendrites surrounding the cell body and an axon that extends out of the ventral horn. This axon travels through the ventral nerve root to join the emerging spinal nerve. The axon is relatively long because it needs to reach muscles in the periphery of the body. The diameters of cell bodies may be on the order of hundreds of micrometres to support the long axon; some axons are a meter in length, such as the lumbar motor neurons that innervate muscles in the first digits of the feet.
The axons will also branch to innervate multiple muscle fibres. Together, the motor neuron and all the muscle fibres that it controls make up a motor unit. Motor units vary in size. Some may contain up to 1000 muscle fibres, such as in the quadriceps, or they may only have 10 fibres, such as in an extraocular muscle. The number of muscle fibres that are part of a motor unit corresponds to the precision of control of that muscle. Also, muscles that have finer motor control have more motor units connecting to them, and this requires a larger topographical field in the primary motor cortex.
Motor neuron axons connect to muscle fibres at a neuromuscular junction. This is a specialised synaptic structure at which multiple axon terminals synapse with the muscle fibre sarcolemma. The synaptic end bulbs of the motor neurons secrete acetylcholine, which binds to receptors on the sarcolemma. The binding of acetylcholine opens ligand-gated ion channels, increasing the movement of cations across the sarcolemma. This depolarises the sarcolemma, initiating muscle contraction. Whereas other synapses result in graded potentials that must reach a threshold in the postsynaptic target, activity at the neuromuscular junction reliably leads to muscle fibre contraction with every nerve impulse received from a motor neuron. However, the strength of contraction and the number of fibres that contract can be affected by the frequency of the motor neuron impulses.
Reflexes
This chapter began by introducing reflexes as an example of the basic elements of the somatic nervous system. Simple somatic reflexes do not include the higher centres discussed for conscious or voluntary aspects of movement. Reflexes can be spinal or cranial, depending on the nerves and central components that are involved. The example described at the beginning of the chapter involved heat and pain sensations from a hot stove causing withdrawal of the arm through a connection in the spinal cord that leads to contraction of the biceps brachii. The description of this withdrawal reflex was simplified, for the sake of the introduction, to emphasise the parts of the somatic nervous system. But to consider reflexes fully, more attention needs to be given to this example.
As you withdraw your hand from the stove, you do not want to slow that reflex down. As the biceps brachii contracts, the antagonistic triceps brachii needs to relax. Because the neuromuscular junction is strictly excitatory, the biceps will contract when the motor nerve is active. Skeletal muscles do not actively relax, instead the motor neuron needs to “quiet down,” or be inhibited. In the hot-stove withdrawal reflex, this occurs through an interneuron in the spinal cord. The interneuron’s cell body is located in the dorsal horn of the spinal cord. The interneuron receives a synapse from the axon of the sensory neuron that detects that the hand is being burned. In response to this stimulation from the sensory neuron, the interneuron then inhibits the motor neuron that controls the triceps brachii. This is done by releasing a neurotransmitter or other signal that hyperpolarises the motor neuron connected to the triceps brachii, making it less likely to initiate an action potential. With this motor neuron being inhibited, the triceps brachii relaxes. Without the antagonistic contraction, withdrawal from the hot stove is faster and keeps further tissue damage from occurring.
Another example of a withdrawal reflex occurs when you step on a painful stimulus, like a tack or a sharp rock. The nociceptors that are activated by the painful stimulus activate the motor neurons responsible for contraction of the tibialis anterior muscle. This causes dorsiflexion of the foot. An inhibitory interneuron, activated by a collateral branch of the nociceptor fibre, will inhibit the motor neurons of the gastrocnemius and soleus muscles to cancel plantar flexion. An important difference in this reflex is that plantar flexion is most likely in progress as the foot is pressing down onto the tack. Contraction of the tibialis anterior is not the most important aspect of the reflex, as continuation of plantar flexion will result in further damage from stepping onto the tack.
Another type of reflex is a stretch reflex. In this reflex, when a skeletal muscle is stretched, a muscle spindle receptor is activated. The axon from this receptor structure will cause direct contraction of the muscle. A collateral of the muscle spindle fibre will also inhibit the motor neuron of the antagonist muscles. The reflex helps to maintain muscles at a constant length. A common example of this reflex is the knee jerk that is elicited by a rubber hammer struck against the patellar ligament in a physical exam.
A specialised reflex to protect the surface of the eye is the corneal reflex, or the eye blink reflex. When the cornea is stimulated by a tactile stimulus, or even by bright light in a related reflex, blinking is initiated. The sensory component travels through the trigeminal nerve, which carries somatosensory information from the face, or through the optic nerve, if the stimulus is bright light. The motor response travels through the facial nerve and innervates the orbicularis oculi on the same side. This reflex is commonly tested during a physical exam using an air puff or a gentle touch of a cotton-tipped applicator.
Section Review
The motor components of the somatic nervous system begin with the frontal lobe of the brain, where the prefrontal cortex is responsible for higher functions such as working memory. The integrative and associate functions of the prefrontal lobe feed into the secondary motor areas, which help plan movements. The premotor cortex and supplemental motor area then feed into the primary motor cortex that initiates movements. Large Betz cells project through the corticobulbar and corticospinal tracts to synapse on lower motor neurons in the brain stem and ventral horn of the spinal cord, respectively. These connections are responsible for generating movements of skeletal muscles.
The extrapyramidal system includes projections from the brain stem and higher centres that influence movement, mostly to maintain balance and posture, as well as to maintain muscle tone. The superior colliculus and red nucleus in the midbrain, the vestibular nuclei in the medulla, and the reticular formation throughout the brain stem each have tracts projecting to the spinal cord in this system. Descending input from the secondary motor cortices, basal nuclei, and cerebellum connect to the origins of these tracts in the brain stem.
All these motor pathways project to the spinal cord to synapse with motor neurons in the ventral horn of the spinal cord. These lower motor neurons are the cells that connect to skeletal muscle and cause contractions. These neurons project through the spinal nerves to connect to the muscles at neuromuscular junctions. One motor neuron connects to multiple muscle fibres within a target muscle. The number of fibres that are innervated by a single motor neuron varies based on the precision necessary for that muscle and the amount of force necessary for that motor unit. The quadriceps, for example, have many fibres controlled by single motor neurons for powerful contractions that do not need to be precise. The extraocular muscles have only a small number of fibres controlled by each motor neuron because moving the eyes does not require much force but needs to be very precise.
Reflexes are the simplest circuits within the somatic nervous system. A withdrawal reflex from a painful stimulus only requires the sensory fibre that enters the spinal cord and the motor neuron that projects to a muscle. Antagonist and postural muscles can be coordinated with the withdrawal, making the connections more complex. The simple, single neuronal connection is the basis of somatic reflexes. The corneal reflex is contraction of the orbicularis oculi muscle to blink the eyelid when something touches the surface of the eye. Stretch reflexes maintain a constant length of muscles by causing a contraction of a muscle to compensate for a stretch that can be sensed by a specialised receptor called a muscle spindle.
Distinguish the types of intercellular communication, their importance, mechanisms and effects
Identify the major organs and tissues of the endocrine system and their location in the body
Communication is a process in which a sender transmits signals to one or more receivers to control and coordinate actions. In the human body, two major organ systems participate in relatively “long distance” communication: the nervous system and the endocrine system. Together, these two systems are primarily responsible for maintaining homeostasis in the body.
Neural and Endocrine Signalling
The nervous system uses two types of intercellular communication—electrical and chemical signalling—either by the direct action of an electrical potential, or in the latter case, through the action of chemical neurotransmitters such as serotonin or noradrenaline. Neurotransmitters act locally and rapidly. When an electrical signal in the form of an action potential arrives at the synaptic terminal, they diffuse across the synaptic cleft (the gap between a sending neuron and a receiving neuron or muscle cell). Once the neurotransmitters interact (bind) with receptors on the receiving (post-synaptic) cell, the receptor stimulation is transduced into a response such as continued electrical signalling or modification of cellular response. The target cell responds within milliseconds of receiving the chemical “message”; this response then ceases very quickly once the neural signalling ends. In this way, neural communication enables body functions that involve quick, brief actions, such as movement, sensation, and cognition. In contrast, the endocrine system uses just one method of communication: chemical signalling. These signals are sent by the endocrine organs, which secrete chemicals—the hormone—into the extracellular fluid. Hormones are transported primarily via the bloodstream throughout the body, where they bind to receptors on target cells, inducing a characteristic response. As a result, endocrine signalling requires more time than neural signalling to prompt a response in target cells, though the precise amount of time varies with different hormones. For example, the hormones released when you are confronted with a dangerous or frightening situation, called the fight-or-flight response, occur by the release of adrenal hormones—adrenaline and noradrenaline—within seconds. In contrast, it may take up to 48 hours for target cells to respond to certain reproductive hormones.
In addition, endocrine signalling is typically less specific than neural signalling. The same hormone may play a role in a variety of different physiological processes depending on the target cells involved, for example, the hormone oxytocin promotes uterine contractions in women in labour and it is also important in breastfeeding and may be involved in the sexual response and in feelings of emotional attachment in both males and females.
In general, the nervous system involves quick responses to rapid changes in the external environment, and the endocrine system is usually slower acting—taking care of the internal environment of the body, maintaining homeostasis, and controlling reproduction (Table 14.1.1). So how does the fight-or-flight response that was mentioned earlier happen so quickly if hormones are usually slower acting? It is because the two systems are connected. It is the fast action of the nervous system in response to the danger in the environment that stimulates the adrenal glands to secrete their hormones. As a result, the nervous system can cause rapid endocrine responses to keep up with sudden changes in both the external and internal environments when necessary.
Table 14.1.1. Endocrine and nervous systems
Endocrine System
Nervous system
Signalling mechanisms(s)
Chemical
Chemical/electrical
Primary chemical signal
Hormones
Neurotransmitters
Distance travelled
Long or short
Always short
Response time
Fast or slow
Always fast
Environment targeted
Internal
Internal and external
Structures of the Endocrine System
The endocrine system consists of cells, tissues and organs that secrete hormones as a primary or secondary function. The endocrine gland is the major player in this system. The primary function of these ductless glands is to secrete their hormones directly into the surrounding fluid. The interstitial fluid and the blood vessels then transport the hormones throughout the body. The endocrine system includes the pituitary, thyroid, parathyroid, adrenal and pineal glands (Figure 14.1.1). Some of these glands have both endocrine and non-endocrine (exocrine) functions, for example, the pancreas contains cells that function in digestion as well as cells that secrete the hormones insulin and glucagon, which regulate blood glucose levels. The hypothalamus, thymus, heart, kidneys, stomach, small intestine, liver, skin, female ovaries and male testes are other organs that contain cells with endocrine function. Additionally, adipose tissue has long been known to produce hormones and recent research has shown that even bone tissue has endocrine functions.
Figure 14.1.1. Endocrine system. Endocrine glands and cells are located throughout the body and play an important role in homeostasis.
The ductless endocrine glands are not to be confused with the body’s exocrine system, whose glands release their secretions through ducts. Examples of exocrine glands include the sebaceous and sweat glands of the skin. As just noted, the pancreas also has an exocrine function: most of its cells secrete pancreatic secretion through the pancreatic and accessory ducts to the lumen of the small intestine.
Other Types of Chemical Signalling
In endocrine signalling, hormones secreted into the extracellular fluid diffuse into the blood or lymph and can then travel great distances throughout the body. In contrast, autocrine signalling takes place within the same cell. An autocrine (auto- = “self”) is a chemical that elicits a response in the same cell that secreted it. Interleukin-1, or IL-1, is a signalling molecule that plays an important role in inflammatory response. The cells that secrete IL-1 have receptors on their cell surface that bind these molecules, resulting in autocrine signalling.
Local intercellular communication is the province of the paracrine, also called a paracrine factor, which is a chemical that induces a response in neighbouring cells. Although paracrines may enter the bloodstream, their concentration is generally too low to elicit a response from distant tissues. A familiar example to those with asthma is histamine, a paracrine that is released by immune cells in the bronchial tree. Histamine causes the smooth muscle cells of the bronchi to constrict, narrowing the airways. Another example is the neurotransmitters of the nervous system, which act only locally within the synaptic cleft.
Career Connections
Endocrinologist
Endocrinology is a specialty in the field of medicine that focuses on the treatment of endocrine system disorders. Endocrinologists—medical doctors who specialise in this field—are experts in treating diseases associated with hormonal systems, ranging from thyroid disease to diabetes mellitus. Endocrine surgeons treat endocrine disease through the removal, or resection, of the affected endocrine gland.
Patients who are referred to endocrinologists may have signs and symptoms or blood test results that suggest excessive or impaired functioning of an endocrine gland or endocrine cells. The endocrinologist may order additional blood tests to determine whether the patient’s hormonal levels are abnormal, or they may stimulate or suppress the function of the suspect endocrine gland and then have blood taken for analysis. Treatment varies according to the diagnosis. Some endocrine disorders, such as type 2 diabetes, may respond to lifestyle changes such as modest weight loss, adoption of a healthy diet, and regular physical activity. Other disorders may require medication, such as hormone replacement, and routine monitoring by the endocrinologist. These include disorders of the pituitary gland that can affect growth and disorders of the thyroid gland that can result in a variety of metabolic problems.
Some patients experience health problems as a result of the normal decline in hormones that can accompany ageing. These patients can consult with an endocrinologist to weigh the risks and benefits of hormone replacement therapy intended to boost their natural levels of reproductive hormones.
In addition to treating patients, endocrinologists may be involved in research to improve the understanding of endocrine system disorders and develop new treatments for these diseases.
Section Review
The endocrine system consists of cells, tissues and organs that secrete hormones critical to homeostasis. The body coordinates its functions through two major types of communication: neural and endocrine. Neural communication includes both electrical and chemical signalling between neurons and target cells. Endocrine communication involves chemical signalling via the release of hormones into the extracellular fluid. From there, hormones diffuse into the bloodstream and may travel to distant body regions, where they elicit a response in target cells. Endocrine glands are ductless glands that secrete hormones. Many organs of the body with other primary functions—such as the heart, stomach, and kidneys—also have hormone-secreting cells.
Identify the three major classes of hormones on the basis of chemical structure
Compare and contrast intracellular and cell membrane hormone receptors
Describe signalling pathways that involve cAMP and IP3
Identify several factors that influence a target cell’s response
Discuss the role of feedback loops and humoral, hormonal, and neural stimuli in hormone control
Although a given hormone may travel throughout the body in the bloodstream, it will affect the activity only of its target cells; that is, cells with receptors for that particular hormone. Once the hormone binds to the receptor, a chain of events is initiated that leads to the target cell’s response. Hormones play a critical role in the regulation of physiological processes because of the target cell responses they regulate. These responses contribute to human reproduction, growth and development of body tissues, metabolism, fluid and electrolyte balance, sleep and many other body functions. The major hormones of the human body and their effects are identified in Table 14.2.1.
Table 14.2.2. Endocrine glands and their major hormones
Endocrine gland
Associated hormones
Chemical effect
Effect
Pituitary
(anterior)
Growth hormone
(GH)
Protein
Promotes growth of body tissues
Prolactin
(PRL)
Peptide
Promotes milk production
Thyroid-stimulating hormone
(TSH)
Glycoprotein
Stimulates thyroid hormone release
Adrenocorticotropic hormone
(ACTH)
Peptide
Stimulates hormone release by adrenal cortex
Follicle-stimulating hormone
(FSH)
Glycoprotein
Stimulates gamete production
Luteinising hormone
(LH)
Glycoprotein
Stimulates androgen production by gonads
Pituitary
(posterior)
Antidiuretic hormone
(ADH)
Peptide
Stimulates water reabsorption by kidneys
Oxytocin
Peptide
Stimulates urine contractions during childbirth
Thyroid
Thyroxine (T4), triiodothyronine (T3)
Amine
Stimulate basal metabolic rate
Calcitonin
Peptide
Reduces blood Ca2+ levels
Parathyroid
Parathyroid hormone
(PTH)
Peptide
Increases blood Ca2+ levels
Adrenal
(cortex)
Aldosterone
Steroid
Increases blood Na+ levels
Cortisol, corticosterone, cortisone
Steroid
Increase blood glucose levels
Adrenal
(medulla)
Adrenaline, noradrenaline
Amine
Stimulate fight-or-flight response
Pineal
Melatonin
Amine
Regulates sleep cycles
Pancreas
Insulin
Protein
Reduces blood glucose levels
Glucagon
Protein
Increases blood glucose levels
Testes
Testosterone
steroid
Stimulate development of male secondary sex characteristics and sperm production
Ovaries
Oestrogens and progesterone
Steroid
Stimulate development of female secondary sex characteristics and prepare the body for childbirth
Types of Hormones
The hormones of the human body can be divided into two major groups on the basis of their chemical structure. Hormones derived from amino acids include amines, peptides, and proteins. Those derived from lipids include steroids (Figure 14.2.1). These chemical groups affect a hormone’s distribution, the type of receptors it binds to, and other aspects of its function.
Figure 14.2.1. Amine, peptide, protein and steroid hormone structure.
Amine Hormones
Hormones derived from the modification of amino acids are referred to as amine hormones. Typically, the original structure of the amino acid is modified such that a –COOH, or carboxyl, group is removed, whereas the −NH2+, −NH3+, or amine, group remains.
Amine hormones are synthesised from the amino acids tryptophan or tyrosine. An example of a hormone derived from tryptophan is melatonin, which is secreted by the pineal gland and helps regulate circadian rhythm. Tyrosine derivatives include the metabolism-regulating thyroid hormones, as well as the catecholamines, such as adrenaline, noradrenaline, and dopamine. Adrenaline and noradrenaline are secreted by the adrenal medulla and play a role in the fight-or-flight response, whereas dopamine is secreted by the hypothalamus and inhibits the release of certain anterior pituitary hormones.
Peptide and Protein Hormones
Whereas the amine hormones are derived from a single amino acid, peptide and protein hormones consist of multiple amino acids that link to form an amino acid chain. Peptide hormones consist of short chains of amino acids, whereas protein hormones are longer polypeptides. Both types are synthesised like other body proteins: DNA is transcribed into mRNA, which is translated into an amino acid chain.
Examples of peptide hormones include antidiuretic hormone (ADH), a pituitary hormone important in fluid balance, and atrial-natriuretic peptide (ANP), which is produced by the cells of the heart and helps to decrease blood pressure. Some examples of protein hormones include growth hormone, which is produced by the pituitary gland, and follicle-stimulating hormone (FSH) which has an attached carbohydrate group and is thus classified as a glycoprotein. FSH helps stimulate the maturation of eggs in the ovaries and sperm in the testes.
Steroid Hormones
The primary hormones derived from lipids are steroids. Steroid hormones are derived from the lipid cholesterol, for example the reproductive hormones testosterone and the oestrogens—which are produced by the gonads (testes and ovaries)—are steroid hormones. The adrenal glands produce the steroid hormone aldosterone, which is involved in osmoregulation and cortisol which plays a role in metabolism.
Like cholesterol, steroid hormones are not soluble in water (they are hydrophobic). Because blood is water-based, lipid-derived hormones must travel to their target cell bound to a transport protein. This more complex structure extends the half-life of steroid hormones much longer than that of hormones derived from amino acids. A hormone’s half-life is the time required for half the concentration of the hormone to be degraded, for example, the lipid-derived hormone cortisol has a half-life of approximately 60 to 90 minutes. In contrast, the amino acid–derived hormone adrenaline has a half-life of approximately one minute.
Pathways of Hormone Action
The message a hormone sends is received by a hormone receptor, a protein located either inside the cell or within the cell membrane. The receptor will process the message by initiating other signalling events or cellular mechanisms that result in the target cell’s response. Hormone receptors recognise molecules with specific shapes and side groups and respond only to those hormones that are recognised. The same type of receptor may be located on cells in different body tissues and trigger somewhat different responses. Thus, the response triggered by a hormone depends not only on the hormone, but also on the target cell.
Once the target cell receives the hormone signal, it can respond in a variety of ways. The response may include the stimulation of protein synthesis, activation or deactivation of enzymes, alteration in the permeability of the cell membrane, altered rates of mitosis and cell growth, and stimulation of the secretion of products. Moreover, a single hormone may be capable of inducing different responses in a given cell.
Intracellular hormone receptors are located inside the cell. Hormones that bind to this type of receptor must be able to cross the cell membrane. Steroid hormones are derived from cholesterol and therefore can readily diffuse through the lipid bilayer of the cell membrane to reach the intracellular receptor (Figure 14.2.2). Thyroid hormones, which contain benzene rings studded with iodine are also lipid-soluble and can enter the cell.
The location of steroid and thyroid hormone binding differs slightly: a steroid hormone may bind to its receptor within the cytosol or within the nucleus. In either case, this binding generates a hormone-receptor complex that moves toward the chromatin in the cell nucleus and binds to a particular segment of the cell’s DNA. In contrast, thyroid hormones bind to receptors already bound to DNA. For both steroid and thyroid hormones, binding of the hormone-receptor complex with DNA triggers transcription of a target gene to mRNA, which moves to the cytosol and directs protein synthesis by ribosomes.
Figure 14.2.2. Binding of lipid-soluble hormones. A steroid hormone directly initiates the production of proteins within a target cell. Steroid hormones easily diffuse through the cell membrane. The hormone binds to its receptor in the cytosol, forming a receptor–hormone complex. The receptor–hormone complex then enters the nucleus and binds to the target gene on the DNA. Transcription of the gene creates a messenger RNA that is translated into the desired protein within the cytoplasm.
Pathway Involving Cell Membrane Hormone Receptors
Hydrophilic, or water-soluble, hormones are unable to diffuse through the lipid bilayer of the cell membrane and must therefore pass on their message to a receptor located at the surface of the cell. Except for thyroid hormones, which are lipid-soluble, all amino acid–derived hormones bind to cell membrane receptors that are located, at least in part, on the extracellular surface of the cell membrane. Therefore, they do not directly affect the transcription of target genes, but instead initiate a signalling cascade that is carried out by a molecule called a second messenger. In this case, the hormone is called a first messenger.
The second messenger used by most hormones is cyclic adenosine monophosphate (cAMP). In the cAMP second messenger system, a water-soluble hormone binds to its receptor in the cell membrane (Step 1 in Figure 14.2.3). This receptor is associated with an intracellular component called a G protein and binding of the hormone activates the G-protein component (Step 2). The activated G protein in turn activates an enzyme called adenylyl cyclase, also known as adenylate cyclase (Step 3), which converts adenosine triphosphate (ATP) to cAMP (Step 4). As the second messenger, cAMP activates a type of enzyme called a protein kinase that is present in the cytosol (Step 5). Activated protein kinases initiate a phosphorylation cascade, in which multiple protein kinases phosphorylate (add a phosphate group to) numerous and various cellular proteins, including other enzymes (Step 6).
Figure 14.2.3. Binding of water-soluble hormones. Water-soluble hormones cannot diffuse through the cell membrane. These hormones must bind to a surface cell-membrane receptor. The receptor then initiates a cell-signalling pathway within the cell involving G proteins, adenylyl cyclase, the secondary messenger cyclic AMP (cAMP), and protein kinases. In the final step, these protein kinases phosphorylate proteins in the cytoplasm. This activates proteins in the cell that carry out the changes specified by the hormone.
The phosphorylation of cellular proteins can trigger a wide variety of effects, from nutrient metabolism to the synthesis of different hormones and other products. The effects vary according to the type of target cell, the G proteins and kinases involved, and the phosphorylation of proteins. Examples of hormones that use cAMP as a second messenger include calcitonin, which is important for bone construction and regulating blood calcium levels; glucagon, which plays a role in blood glucose levels; and thyroid-stimulating hormone, which causes the release of T3 and T4 from the thyroid gland.
Overall, the phosphorylation cascade significantly increases the efficiency, speed, and specificity of the hormonal response, as thousands of signalling events can be initiated simultaneously in response to a very low concentration of hormone in the bloodstream. However, the duration of the hormone signal is short, as cAMP is quickly deactivated by the enzyme phosphodiesterase (PDE), which is located in the cytosol. The action of PDE helps to ensure that a target cell’s response ceases quickly unless new hormones arrive at the cell membrane.
Importantly, there are also G proteins that decrease the levels of cAMP in the cell in response to hormone binding. For example, when growth hormone–inhibiting hormone (GHIH), also known as somatostatin, binds to its receptors in the pituitary gland, the level of cAMP decreases, thereby inhibiting the secretion of human growth hormone.
Not all water-soluble hormones initiate the cAMP second messenger system. One common alternative system uses calcium ions as a second messenger. In this system, G proteins activate the enzyme phospholipase C (PLC), which functions similarly to adenylyl cyclase. Once activated, PLC cleaves a membrane-bound phospholipid into two molecules: diacylglycerol (DAG) and inositol triphosphate (IP3). Like cAMP, DAG activates protein kinases that initiate a phosphorylation cascade. At the same time, IP3 causes calcium ions to be released from storage sites within the cytosol, such as from within the smooth endoplasmic reticulum. The calcium ions then act as second messengers in two ways: they can influence enzymatic and other cellular activities directly, or they can bind to calcium-binding proteins, the most common of which is calmodulin. Upon binding calcium, calmodulin is able to modulate protein kinase within the cell. Examples of hormones that use calcium ions as a second messenger system include angiotensin II, which helps regulate blood pressure through vasoconstriction, and growth hormone–releasing hormone (GHRH), which causes the pituitary gland to release growth hormones.
Factors Affecting Target Cell Response
You will recall that target cells must have receptors specific to a given hormone if that hormone is to trigger a response. But several other factors influence the target cell response, for example, the presence of a significant concentration of a hormone circulating in the bloodstream can cause its target cells to decrease their number of receptors for that hormone. This process is called downregulation, and it allows cells to become less reactive to the excessive hormone levels. When the concentration of a hormone is chronically reduced, target cells engage in upregulation to increase their number of receptors. This process allows cells to be more sensitive to the hormone that is present. Cells can also alter the sensitivity of the receptors themselves to various hormones.
Two or more hormones can interact to affect the response of cells in a variety of ways. The three most common types of interaction are as follows:
The permissive effect, in which the presence of one hormone enables another hormone to act, for example thyroid hormones have complex permissive relationships with certain reproductive hormones. A dietary deficiency of iodine, a component of thyroid hormones, can therefore affect reproductive system development and functioning.
The synergistic effect, in which two hormones with similar effects produce an amplified response. In some cases, two hormones are required for an adequate response. For example, two different reproductive hormones—FSH from the pituitary gland and oestrogens from the ovaries—are required for the maturation of female ova (egg cells).
The antagonistic effect, in which two hormones have opposing effects. A familiar example is the effect of two pancreatic hormones, insulin and glucagon. Insulin increases the liver’s storage of glucose as glycogen, decreasing blood glucose, whereas glucagon stimulates the breakdown of glycogen stores, increasing blood glucose.
Regulation of Hormone Secretion
To prevent abnormal hormone levels and a potential disease state, hormone concentrations must be tightly controlled. The body maintains this control by balancing hormone production and degradation. Feedback loops govern the initiation and maintenance of most hormone secretion in response to various stimuli.
Role of Feedback Loops
The contribution of feedback loops to homeostasis will only be briefly reviewed here. Positive feedback loops are characterised by the release of additional hormone in response to an original hormone release. The release of oxytocin during childbirth is a positive feedback loop. The initial release of oxytocin begins to signal the uterine muscles to contract, which pushes the foetus toward the cervix, causing it to stretch. This, in turn, signals the pituitary gland to release more oxytocin, causing labour contractions to intensify. The release of oxytocin decreases after the birth of the child.
The more common method of hormone regulation is the negative feedback loop. Negative feedback is characterised by the inhibition of further secretion of a hormone in response to adequate levels of that hormone. This allows blood concentrations of the hormone to be regulated within a narrow range. An example of a negative feedback loop is the release of glucocorticoid hormones from the adrenal glands, as directed by the hypothalamus and pituitary gland. As glucocorticoid concentrations in the blood rise, the hypothalamus and pituitary gland reduce their signalling to the adrenal glands to prevent additional glucocorticoid secretion (Figure 14.2.4).
Figure 14.2.4. Negative feedback loop. The release of adrenal glucocorticoids is stimulated by the release of hormones from the hypothalamus and pituitary gland. This signalling is inhibited when glucocorticoid levels become elevated by causing negative signals to the pituitary gland and hypothalamus.
Role of Endocrine Gland Stimuli
Reflexes triggered by both chemical and neural stimuli control endocrine activity. These reflexes may be simple, involving only one hormone response, or they may be more complex and involve many hormones, as is the case with the hypothalamic control of various anterior pituitary–controlled hormones.
Humoral stimuli are changes in blood concentrations of non-hormone chemicals, such as nutrients or ions, which cause the release or inhibition of a hormone to, in turn, maintain homeostasis. Osmoreceptors in the hypothalamus detect changes in blood osmolarity (the concentration of solutes in the blood plasma) is an example of this hormonal stimuli. If blood osmolarity is too high, meaning that the blood is not dilute enough, osmoreceptors signal the hypothalamus to release ADH. The hormone causes the kidneys to reabsorb more water and reduce the volume of urine produced. This reabsorption causes a reduction of the osmolarity of the blood, diluting the blood to the appropriate level. The regulation of blood glucose is another example. High blood glucose concentrations cause the release of insulin from the pancreas, which increases glucose uptake by cells (via special transporters) and liver storage of glucose as glycogen.
An endocrine gland may also secrete a hormone in response to the presence of another hormone produced by a different endocrine gland. Such hormonal stimuli often involve the hypothalamus, which produces releasing and inhibiting hormones that control the secretion of a variety of pituitary hormones.
In addition to these chemical signals, hormones can also be released in response to neural stimuli. A common example of neural stimuli is the activation of the fight-or-flight response by the sympathetic nervous system. When an individual perceives danger, sympathetic neurons signal the adrenal glands to secrete noradrenaline and adrenaline. The two hormones dilate blood vessels, increase the heart and respiratory rate, and suppress the digestive and immune systems. These responses boost the body’s transport of oxygen to the brain and muscles, thereby improving the body’s ability to fight or flee.
Everyday Connections
Bisphenol A and Endocrine Disruption
You may have heard news reports about the effects of a chemical called bisphenol A (BPA) in various types of food packaging. BPA is used in the manufacturing of hard plastics and epoxy resins. Common food-related items that may contain BPA include the lining of aluminium cans, plastic food-storage containers, drinking cups, as well as baby bottles and “sippy” cups. Other uses of BPA include medical equipment, thermal paper, dental fillings, and the lining of water pipes.
Research shows that BPA is an endocrine disruptor, meaning that it negatively interferes with the endocrine system, particularly during the prenatal and postnatal development period. In particular, BPA mimics the hormonal effects of oestrogens and has the opposite effect—that of androgens. Following the reports released by the World Health Organisation (WHO) and the US Food and Drug Administration (FDA), the Food Standards Australia New Zealand (FSANZ) had also released a detailed report on BPA in food packaging and consumer risk. The Australian government had announced a voluntary phase out of baby bottles containing BPA by major retailers back in 2010. Additionally, the FSANZ had conducted a study of BPA found in consumable foodstuff and determined that in Australia and New Zealand the levels of PBA consumed are considerably low and do not pose significant risk to humans. The U.S. Food and Drug Administration (FDA) notes in their statement about BPA safety that although traditional toxicology studies have supported the safety of low levels of exposure to BPA, recent studies using novel approaches to test for subtle effects have led to some concern about the potential effects of BPA on the brain, behaviour, and prostate gland in foetuses, infants, and young children. Both Canada and the European Union have completely banned the use of BPA in baby products.
The potential harmful effects of BPA have been studied in both animal models and humans and include a large variety of health effects, such as developmental delay and disease, for example, prenatal exposure to BPA during the first trimester of human pregnancy may be associated with wheezing and aggressive behaviour during childhood. Adults exposed to high levels of BPA may experience altered thyroid signalling and male sexual dysfunction. BPA exposure during the prenatal or postnatal period of development in animal models has been observed to cause neurological delays, changes in brain structure and function, sexual dysfunction, asthma, and increased risk for multiple cancers. In vitro studies have also shown that BPA exposure causes molecular changes that initiate the development of cancers of the breast, prostate, and brain. Although these studies have implicated BPA in numerous ill health effects, some experts caution that some of these studies may be flawed and that more research needs to be done. In the meantime, the FDA recommends that consumers take precautions to limit their exposure to BPA. In addition to purchasing foods in packaging free of BPA, consumers should avoid carrying or storing foods or liquids in bottles with the recycling code 3 or 7. Foods and liquids should not be microwave-heated in any form of plastic: use paper, glass, or ceramics instead.
Section Review
Hormones are derived from amino acids or lipids. Amine hormones originate from the amino acids tryptophan or tyrosine. Larger amino acid hormones include peptides and protein hormones. Steroid hormones are derived from cholesterol.
Steroid hormones and thyroid hormone are lipid soluble. All other amino acid–derived hormones are water soluble. Hydrophobic hormones are able to diffuse through the membrane and interact with an intracellular receptor. In contrast, hydrophilic hormones must interact with cell membrane receptors. These are typically associated with a G protein, which becomes activated when the hormone binds the receptor. This initiates a signalling cascade that involves a second messenger, such as cyclic adenosine monophosphate (cAMP). Second messenger systems greatly amplify the hormone signal, creating a broader, more efficient, and faster response.
Hormones are released upon stimulation that is of either chemical or neural origin. Regulation of hormone release is primarily achieved through negative feedback. Various stimuli may cause the release of hormones, but there are three major types. Humoral stimuli are changes in ion or nutrient levels in the blood. Hormonal stimuli are changes in hormone levels that initiate or inhibit the secretion of another hormone. Finally, a neural stimulus occurs when a nerve impulse prompts the secretion or inhibition of a hormone.
Explain the interrelationships of the anatomy and functions of the hypothalamus and the posterior and anterior lobes of the pituitary gland
Identify the two hormones released from the posterior pituitary, their target cells and their principal actions
Identify the six hormones produced by the anterior lobe of the pituitary gland, their target cells, their principal actions and their regulation by the hypothalamus
The hypothalamus–pituitary complex can be thought of as the “command centre” of the endocrine system. This complex secretes several hormones that directly produce responses in target tissues, as well as hormones that regulate the synthesis and secretion of hormones of other glands. In addition, the hypothalamus–pituitary complex coordinates the messages of the endocrine and nervous systems. In many cases, a stimulus received by the nervous system must pass through the hypothalamus–pituitary complex to be translated into hormones that can initiate a response.
The hypothalamus is a structure of the diencephalon of the brain located anterior and inferior to the thalamus (Figure 14.3.1). It has both neural and endocrine functions, producing and secreting many hormones. In addition, the hypothalamus is anatomically and functionally related to the pituitary gland (or hypophysis), a bean-sized organ suspended from it by a stem called the infundibulum (or pituitary stalk). The pituitary gland is cradled within the sella turcica of the sphenoid bone of the skull. It consists of two lobes that arise from distinct parts of embryonic tissue: the posterior pituitary (neurohypophysis) is neural tissue, whereas the anterior pituitary (also known as the adenohypophysis) is glandular tissue that develops from the primitive digestive tract, specifically the developing hard palate. The hormones secreted by the posterior and anterior pituitary, and the intermediate zone between the lobes are summarised in Table 14.3.1.
Figure 14.3.1. Hypothalamus–pituitary complex. The hypothalamus region lies inferior and anterior to the thalamus. It connects to the pituitary gland by the stalk-like infundibulum. The pituitary gland consists of an anterior and posterior lobe, with each lobe secreting different hormones in response to signals from the hypothalamus.
Table 14.3.3. Pituitary hormones
Pituitary lobe
Associated hormones
Chemical class
Effect
Anterior
Growth hormone
(GH)
Protein
Promotes growth of body tissues
Prolactin
(PRL)
Peptide
Promotes milk production from mammary glands
Thyroid-stimulating hormone
(TSH)
Glycoprotein
Stimulates thyroid hormone release from thyroid
Adrenocorticotropic hormone
(ACTH)
Peptide
Stimulates hormone release by adrenal cortex
Follicle-stimulating hormone
(FSH)
Glycoprotein
Stimulates gamete production in gonads
Luteinising hormone
(LH)
Glycoprotein
Stimulates androgen production by gonads
Posterior
Antidiuretic hormone
(ADH)
Peptide
Stimulates water reabsorption by kidney
Oxytocin
Peptide
Stimulates uterine contractions during childbirth
Intermediate zone
Melanocyte-stimulating hormone
Peptide
Simulates melanin formation in melanocytes
Posterior Pituitary
The posterior pituitary is actually an extension of the neurons of the paraventricular and supraoptic nuclei of the hypothalamus. The cell bodies of these regions rest in the hypothalamus, but their axons descend as the hypothalamic–hypophyseal tract within the infundibulum, and end in axon terminals that comprise the posterior pituitary (14.3.2).
Figure 14.3.2. Posterior pituitary. Neurosecretory cells in the hypothalamus release oxytocin (OT) or Antidiuretic hormone (ADH) into the posterior lobe of the pituitary gland. These hormones are stored or released into the blood via the capillary plexus.
The posterior pituitary gland does not produce hormones, but rather stores and secretes hormones produced by the hypothalamus. The paraventricular nuclei produce the hormone oxytocin, whereas the supraoptic nuclei produce ADH. These hormones travel along the axons into storage sites in the axon terminals of the posterior pituitary. In response to signals from the same hypothalamic neurons, the hormones are released from the axon terminals into the bloodstream.
Oxytocin (OT)
When foetal development is complete, the peptide-derived hormone oxytocin (tocia- = “childbirth”) stimulates uterine contractions and dilation of the cervix. Throughout most of pregnancy, oxytocin hormone receptors are not expressed at high levels in the uterus. Toward the end of pregnancy, the synthesis of oxytocin receptors in the uterus increases, and the smooth muscle cells of the uterus become more sensitive to its effects. Oxytocin is continually released throughout childbirth through a positive feedback mechanism. As noted earlier, oxytocin prompts uterine contractions that push the foetal head toward the cervix. In response, cervical stretching stimulates additional oxytocin to be synthesised by the hypothalamus and released from the pituitary. This increases the intensity and effectiveness of uterine contractions and prompts additional dilation of the cervix. The feedback loop continues until birth.
Although the mother’s high blood concentrations of oxytocin begin to decrease immediately following birth, oxytocin continues to play a role in maternal and newborn health. First, oxytocin is necessary for the milk ejection reflex (commonly referred to as “let-down”) in breastfeeding women. As the newborn begins suckling, sensory receptors in the nipples transmit signals to the hypothalamus. In response, oxytocin is secreted and released into the bloodstream. Within seconds, cells in the mother’s milk ducts contract, ejecting milk into the infant’s mouth. Secondly, in both males and females, oxytocin is thought to contribute to parent–newborn bonding, known as attachment. Oxytocin is also thought to be involved in feelings of love and closeness, as well as in the sexual response.
Antidiuretic Hormone (ADH)
The solute concentration of the blood, or blood osmolarity, may change in response to the consumption of certain foods and fluids, as well as in response to disease, injury, medications, or other factors. Blood osmolarity is constantly monitored by osmoreceptors—specialised cells within the hypothalamus that are particularly sensitive to the concentration of sodium ions and other solutes.
In response to high blood osmolarity, which can occur during dehydration or following a very salty meal, the osmoreceptors signal the posterior pituitary to release antidiuretic hormone (ADH). The target cells of ADH are located in the tubular cells of the kidneys. Its effect is to increase epithelial permeability to water, allowing increased water reabsorption. The more water reabsorbed from the filtrate, the greater the amount of water that is returned to the blood and the less that is excreted in the urine. A greater concentration of water results in a reduced concentration of solutes. ADH is also known as vasopressin because, in very high concentrations, it causes constriction of blood vessels, which increases blood pressure by increasing peripheral resistance. The release of ADH is controlled by a negative feedback loop. As blood osmolarity decreases, the hypothalamic osmoreceptors sense the change and prompt a corresponding decrease in the secretion of ADH. As a result, less water is reabsorbed from the urine filtrate.
Interestingly, drugs can affect the secretion of ADH, for example, alcohol consumption inhibits the release of ADH, resulting in increased urine production that can eventually lead to dehydration and a hangover. A disease called diabetes insipidus is characterised by chronic underproduction of ADH that causes chronic dehydration. Because little ADH is produced and secreted, not enough water is reabsorbed by the kidneys. Although patients feel thirsty, and increase their fluid consumption, this doesn’t effectively decrease the solute concentration in their blood because ADH levels are not high enough to trigger water reabsorption in the kidneys. Electrolyte imbalances can occur in severe cases of diabetes insipidus.
Anterior Pituitary
The anterior pituitary originates from the digestive tract in the embryo and migrates toward the brain during foetal development. There are three regions: the pars distalis is the most anterior, the pars intermedia is adjacent to the posterior pituitary, and the pars tuberalis is a slender “tube” that wraps the infundibulum.
Recall that the posterior pituitary does not synthesise hormones, but merely stores them. In contrast, the anterior pituitary does manufacture hormones. However, the secretion of hormones from the anterior pituitary is regulated by two classes of hormones. These hormones—secreted by the hypothalamus—are the releasing hormones that stimulate the secretion of hormones from the anterior pituitary and the inhibiting hormones that inhibit secretion.
Hypothalamic hormones are secreted by neurons but enter the anterior pituitary through blood vessels (Figur 14.3.3). Within the infundibulum is a bridge of capillaries that connects the hypothalamus to the anterior pituitary. This network, called the hypophyseal portal system, allows hypothalamic hormones to be transported to the anterior pituitary without first entering the systemic circulation. The system originates from the superior hypophyseal artery, which branches off the carotid arteries and transports blood to the hypothalamus. The branches of the superior hypophyseal artery form the hypophyseal portal system (see Figure 14.3.3). Hypothalamic releasing and inhibiting hormones travel through a primary capillary plexus to the portal veins, which carry them into the anterior pituitary. Hormones produced by the anterior pituitary (in response to releasing hormones) enter a secondary capillary plexus, and from there drain into the circulation.
Figure 14.3.3. Anterior pituitary. The anterior pituitary manufactures seven hormones. The hypothalamus produces separate hormones that stimulate or inhibit hormone production in the anterior pituitary. Hormones from the hypothalamus reach the anterior pituitary via the hypophyseal portal system.
The anterior pituitary produces seven hormones. These are the growth hormone (GH), thyroid-stimulating hormone (TSH), adrenocorticotropic hormone (ACTH), follicle-stimulating hormone (FSH), luteinising hormone (LH), beta endorphin and prolactin. Of the hormones of the anterior pituitary, TSH, ACTH, FSH, and LH are collectively referred to as tropic hormones (trope- = “turning”) because they turn on or off the function of other endocrine glands.
Growth Hormone (GH)
The endocrine system regulates the growth of the human body, protein synthesis and cellular replication. A major hormone involved in this process is growth hormone (GH), also called somatotropin—a protein hormone produced and secreted by the anterior pituitary gland. Its primary function is anabolic; it promotes protein synthesis and tissue building through direct and indirect mechanisms (14.3.4). GH levels are controlled by the release of GHRH and GHIH (also known as somatostatin) from the hypothalamus.
Figure 14.3.4. Hormonal regulation of growth. Growth hormone (GH) directly accelerates the rate of protein synthesis in skeletal muscle and bones. Insulin-like growth factor 1 (IGF-1) is activated by growth hormone and indirectly supports the formation of new proteins in muscle cells and bone.
A glucose-sparing effect occurs when GH stimulates lipolysis, or the breakdown of adipose tissue, releasing fatty acids into the blood. As a result, many tissues switch from glucose to fatty acids as their main energy source, which means that less glucose is taken up from the bloodstream.
GH also initiates the diabetogenic effect in which GH stimulates the liver to break down glycogen to glucose, which is then deposited into the blood. The name “diabetogenic” is derived from the similarity in elevated blood glucose levels observed between individuals with untreated diabetes mellitus and individuals experiencing GH excess. Blood glucose concentrations rise as the result of a combination of glucose-sparing and diabetogenic effects.
GH indirectly mediates growth and protein synthesis by triggering the liver and other tissues to produce a group of proteins called insulin-like growth factors (IGFs). These proteins enhance cellular proliferation and inhibit apoptosis, or programmed cell death. IGFs stimulate cells to increase their uptake of amino acids from the blood for protein synthesis. Skeletal muscle and cartilage cells are particularly sensitive to stimulation from IGFs.
Dysfunction of the endocrine system’s control of growth can result in several disorders, for example, gigantism is a disorder in children that is caused by the secretion of abnormally large amounts of GH, resulting in excessive growth. A similar condition in adults is acromegaly, a disorder that results in the growth of bones in the face, hands, and feet in response to excessive levels of GH in individuals who have stopped growing. Abnormally low levels of GH in children can cause growth impairment—a disorder called pituitary dwarfism (also known as growth hormone deficiency).
Thyroid-Stimulating Hormone (TSH)
The activity of the thyroid gland is regulated by thyroid-stimulating hormone (TSH), also called thyrotropin. TSH is released from the anterior pituitary in response to thyrotropin-releasing hormone (TRH) from the hypothalamus. As discussed shortly, it triggers the secretion of thyroid hormones by the thyroid gland. In a classic negative feedback loop, elevated levels of thyroid hormones in the bloodstream then trigger a drop in production of TRH and subsequently TSH.
Adrenocorticotropic Hormone (ACTH)
The adrenocorticotropic hormone (ACTH), also called corticotropin, stimulates the adrenal cortex (the more superficial “bark” of the adrenal glands) to secrete corticosteroid hormones such as cortisol. ACTH come from a precursor molecule known as pro-opiomelanotropin (POMC) which produces several biologically active molecules when cleaved, including ACTH, melanocyte-stimulating hormone, and the brain opioid peptides known as endorphins.
The release of ACTH is regulated by the corticotropin-releasing hormone (CRH) from the hypothalamus in response to normal physiologic rhythms. A variety of stressors can also influence its release, and the role of ACTH in the stress response is discussed later in this chapter.
Follicle-Stimulating Hormone (FSH) and Luteinsing Hormone (LH)
The endocrine glands secrete a variety of hormones that control the development and regulation of the reproductive system (these glands include the anterior pituitary, the adrenal cortex, and the gonads—the testes in males and the ovaries in females). Much of the development of the reproductive system occurs during puberty and is marked by the development of sex-specific characteristics in both male and female adolescents. Puberty is initiated by gonadotropin-releasing hormone (GnRH), a hormone produced and secreted by the hypothalamus. GnRH stimulates the anterior pituitary to secrete gonadotropins—hormones that regulate the function of the gonads. The levels of GnRH are regulated through a negative feedback loop; high levels of reproductive hormones inhibit the release of GnRH. Throughout life, gonadotropins regulate reproductive function and, in the case of women, the onset and cessation of reproductive capacity.
The gonadotropins include two glycoprotein hormones: follicle-stimulating hormone (FSH) stimulates the production and maturation of sex cells, or gametes, including ova in women and sperm in men. FSH also promotes follicular growth; these follicles then release oestrogens in the female ovaries. Luteinising hormone (LH) triggers ovulation in women, as well as the production of oestrogens and progesterone by the ovaries. LH stimulates production of testosterone by the male testes.
Prolactin (PRL)
As its name implies, prolactin (PRL) promotes lactation (milk production) in women. During pregnancy, it contributes to development of the mammary glands, and after birth, it stimulates the mammary glands to produce breast milk. However, the effects of prolactin depend heavily upon the permissive effects of oestrogens, progesterone, and other hormones. And as noted earlier, the let-down of milk occurs in response to stimulation from oxytocin.
In a non-pregnant woman, prolactin secretion is inhibited by prolactin-inhibiting hormone (PIH), which is actually the neurotransmitter dopamine, and is released from neurons in the hypothalamus. Only during pregnancy do prolactin levels rise in response to prolactin-releasing hormone (PRH) from the hypothalamus.
The cells in the zone between the pituitary lobes secrete a hormone known as melanocyte-stimulating hormone (MSH) that is formed by cleavage of the pro-opiomelanocortin (POMC) precursor protein. Local production of MSH in the skin is responsible for melanin production in response to UV light exposure. The role of MSH made by the pituitary is more complicated. For instance, people with lighter skin generally have the same amount of MSH as people with darker skin. Nevertheless, this hormone is capable of darkening of the skin by inducing melanin production in the skin’s melanocytes. Women also show increased MSH production during pregnancy; in combination with oestrogens, it can lead to darker skin pigmentation, especially the skin of the areolas and labia minora. Figure 14.3.5 is a summary of the pituitary hormones and their principal effects.
Figure 14.3.5. Major pituitary hormones. Major pituitary hormones and their target organs.
Section Review
The hypothalamus–pituitary complex is located in the diencephalon of the brain. The hypothalamus and the pituitary gland are connected by a structure called the infundibulum, which contains vasculature and nerve axons. The pituitary gland is divided into two distinct structures with different embryonic origins. The posterior lobe houses the axon terminals of hypothalamic neurons. It stores and releases into the bloodstream two hypothalamic hormones: oxytocin (OT) and antidiuretic hormone (ADH). The anterior lobe is connected to the hypothalamus by vasculature in the infundibulum and produces and secretes six hormones. Their secretion is regulated, however, by releasing and inhibiting hormones from the hypothalamus. The six anterior pituitary hormones are: growth hormone (GH), thyroid-stimulating hormone (TSH), adrenocorticotropic hormone (ACTH), follicle-stimulating hormone (FSH), luteinising hormone (LH) and prolactin (PRL).
Describe the location and anatomy of the thyroid gland
Discuss the synthesis of triiodothyronine and thyroxine
Explain the role of thyroid hormones in the regulation of basal metabolism
Identify the hormone produced by the parafollicular cells of the thyroid
A butterfly-shaped organ, the thyroid gland is located anterior to the trachea, just inferior to the larynx (Figure 14.4.1). The medial region, called the isthmus, is flanked by wing-shaped left and right lobes. Each of the thyroid lobes are embedded with parathyroid glands, primarily on their posterior surfaces. The tissue of the thyroid gland is composed mostly of thyroid follicles. The follicles are made up of a central cavity filled with a sticky fluid called colloid. Surrounded by a wall of epithelial follicle cells, the colloid is the centre of thyroid hormone production, and that production is dependent on the hormones’ essential and unique component: iodine.
Hormones are produced in the colloid when atoms of the mineral iodine attach to a glycoprotein, called thyroglobulin, that is secreted into the colloid by the follicle cells.
The following steps outline the hormones’ assembly:
Binding of TSH to its receptors in the follicle cells of the thyroid gland causes the cells to actively transport iodide ions (I–) across their cell membrane, from the bloodstream into the cytosol. As a result, the concentration of iodide ions “trapped” in the follicular cells is many times higher than the concentration in the bloodstream.
Iodide ions (I-) then move to the lumen of the follicle cells that border the colloid. There, the ions undergo oxidation (their negatively charged electrons are removed). The oxidation of two iodide ions (2 I–) results in iodine (I2), which passes through the follicle cell membrane into the colloid.
In the colloid, peroxidase enzymes link the iodine to the tyrosine amino acids in thyroglobulin to produce two intermediaries: a tyrosine attached to one iodine and a tyrosine attached to two iodines. When one of each of these intermediaries is linked by covalent bonds, the resulting compound is triiodothyronine (T3), a thyroid hormone with three iodines. Much more commonly, two copies of the second intermediary bond, forming tetraiodothyronine, also known as thyroxine (T4), a thyroid hormone with four iodines.
These hormones remain in the colloid centre of the thyroid follicles until TSH stimulates endocytosis of colloid back into the follicle cells. There, lysosomal enzymes break apart the thyroglobulin colloid, releasing free T3 and T4, which diffuse across the follicle cell membrane and enter the bloodstream.
In the bloodstream, less than one percent of the circulating T3 and T4 remains unbound. This free T3 and T4 can cross the lipid bilayer of cell membranes and be taken up by cells. The remaining 99 percent of circulating T3 and T4 is bound to specialised transport proteins called thyroxine-binding globulins (TBGs), to albumin, or to other plasma proteins. This “packaging” prevents their free diffusion into body cells. When blood levels of T3 and T4 begin to decline, bound T3 and T4 are released from these plasma proteins and readily cross the membrane of target cells. T3 is more potent than T4, and many cells convert T4 to T3 through the removal of an iodine atom.
Regulation of Thyroid Hormones Synthesis
The release of T3 and T4 from the thyroid gland is regulated by thyroid-stimulating hormone (TSH). As shown in Figure 14.4.2, low blood levels of T3 and T4 stimulate the release of thyrotropin-releasing hormone (TRH) from the hypothalamus, which triggers secretion of TSH from the anterior pituitary. In turn, TSH stimulates the thyroid gland to secrete T3 and T4. The levels of TRH, TSH, T3 and T4 are regulated by a negative feedback system in which increasing levels of T3 and T4 decrease the production and secretion of TSH.
Figure 14.4.2. Classic negative feedback loop. A classic negative feedback loop controls the regulation of thyroid hormone levels.
Functions of Thyroid Hormones
The thyroid hormones, T3 and T4, are often referred to as metabolic hormones because their levels influence the body’s basal metabolic rate, the amount of energy used by the body at rest. When T3 and T4 bind to intracellular receptors located on the mitochondria, they cause an increase in nutrient breakdown and the use of oxygen to produce ATP. In addition, T3 and T4 initiate the transcription of genes involved in glucose oxidation. Although these mechanisms prompt cells to produce more ATP, the process is inefficient, and an abnormally increased level of heat is released as a by-product of these reactions. This so-called calorigenic effect (calor- = “heat”) raises body temperature.
Adequate levels of thyroid hormones are also required for protein synthesis and for foetal and childhood tissue development and growth. They are especially critical for normal development of the nervous system both in utero and in early childhood, and they continue to support neurological function in adults. As noted earlier, these thyroid hormones have a complex interrelationship with reproductive hormones, and deficiencies can influence libido, fertility, and other aspects of reproductive function. Finally, thyroid hormones increase the body’s sensitivity to catecholamines (adrenaline and noradrenaline) from the adrenal medulla by upregulation of receptors in the blood vessels. When levels of T3 and T4 hormones are excessive, this effect accelerates the heart rate, strengthens the heartbeat, and increases blood pressure. Because thyroid hormones regulate metabolism, heat production, protein synthesis, and many other body functions, thyroid disorders can have severe and widespread consequences.
Disorders of the Endocrine System:
Iodine Deficiency, Hypothyroidism, and Hyperthyroidism
As discussed above, dietary iodine is required for the synthesis of T3 and T4. But for much of the world’s population, foods do not provide adequate levels of this mineral, because the amount varies according to the level in the soil in which the food was grown, as well as the irrigation and fertilisers used. Marine fish and shrimp tend to have high levels because they concentrate iodine from seawater, but many people in landlocked regions lack access to seafood. Thus, the primary source of dietary iodine in many countries is iodised salt. Fortification of salt with iodine began in the United States in 1924, and in Australia (2009) only iodised salt could be used in the manufacture of most breads. International efforts to iodise salt in the world’s poorest nations continue today.
Dietary iodine deficiency can result in the impaired ability to synthesise T3 and T4, leading to a variety of severe disorders. When T3 and T4 cannot be produced, TSH is secreted in increasing amounts. As a result of this hyperstimulation, thyroglobulin accumulates in the thyroid gland follicles, increasing their deposits of colloid. The accumulation of colloid increases the overall size of the thyroid gland, a condition called a goitre (Figure 14.4.3). A goitre is only a visible indication of the deficiency. Other iodine deficiency disorders include impaired growth and development, decreased fertility, and prenatal and infant death. Moreover, iodine deficiency is the primary cause of preventable intellectual disability worldwide. Congenital hypothyroidism is characterised by cognitive deficits, short stature, and sometimes deafness and muteness in children and adults born to mothers who were iodine-deficient during pregnancy.
In areas of the world with access to iodised salt, dietary deficiency is rare. Instead, inflammation of the thyroid gland is the more common cause of low blood levels of thyroid hormones. Called hypothyroidism, the condition is characterised by a low metabolic rate, weight gain, cold extremities, constipation, reduced libido, menstrual irregularities and reduced mental activity. In contrast, hyperthyroidism—an abnormally elevated blood level of thyroid hormones—is often caused by a pituitary or thyroid tumour. In Graves’ disease, the hyperthyroid state results from an autoimmune reaction in which antibodies overstimulate the follicle cells of the thyroid gland. Hyperthyroidism can lead to an increased metabolic rate, excessive body heat and sweating, diarrhoea, weight loss, tremors and increased heart rate. The person’s eyes may bulge (called exophthalmos) as antibodies produce inflammation in the soft tissues of the orbits. The person may also develop a goitre.
Calcitonin
The thyroid gland also secretes a hormone called calcitonin that is produced by the parafollicular cells (also called C cells) that stud the tissue between distinct follicles. Calcitonin is released in response to a rise in blood calcium levels. It appears to have a function in decreasing blood calcium concentrations by:
Inhibiting the activity of osteoclasts, bone cells that release calcium into the circulation by degrading bone matrix
Increasing osteoblastic activity
Decreasing calcium absorption in the intestines
Increasing calcium loss in the urine
However, these functions are usually not significant in maintaining calcium homeostasis, so the importance of calcitonin is not entirely understood. Pharmaceutical preparations of calcitonin are sometimes prescribed to reduce osteoclast activity in people with osteoporosis and to reduce the degradation of cartilage in people with osteoarthritis. The hormones secreted by thyroid are summarised in Table 14.4.1.
Table 14.4.1. Thyroid hormones
Associated hormones
Chemical class
Effect
Thyroxine (T4),
Triiodothyronine (T3)
Amine
Stimulate basal metabolic rate
Calcitonin
Peptide
Reduces blood Ca2+ levels
Of course, calcium is critical for many other biological processes. It is a second messenger in many signalling pathways, and is essential for muscle contraction, nerve impulse transmission, and blood clotting. Given these roles, it is not surprising that blood calcium levels are tightly regulated by the endocrine system. The organs involved in the regulation are the parathyroid glands.
Section Review
The thyroid gland is a butterfly-shaped organ located in the neck anterior to the trachea. Its hormones regulate basal metabolism, oxygen use, nutrient metabolism, the production of ATP, and calcium homeostasis. They also contribute to protein synthesis and the normal growth and development of body tissues, including maturation of the nervous system, and they increase the body’s sensitivity to catecholamines. The thyroid hormones triiodothyronine (T3) and thyroxine (T4) are produced and secreted by the thyroid gland in response to thyroid-stimulating hormone (TSH) from the anterior pituitary. Synthesis of the amino acid–derived T3 and T4 hormones requires iodine. Insufficient amounts of iodine in the diet can lead to goitre, congenital hypothyroidism and many other disorders.
Describe the location and structure of the parathyroid glands
Describe the hormonal control of blood calcium levels
Discuss the physiological response of parathyroid dysfunction
The parathyroid glands are tiny, round structures usually found embedded in the posterior surface of the thyroid gland (Figure 14.5.1). A thick connective tissue capsule separates the glands from the thyroid tissue. Most people have four parathyroid glands, but occasionally there are more in tissues of the neck or chest. The function of one type of parathyroid cells, the oxyphil cells, is not clear. The primary functional cells of the parathyroid glands are the chief cells. These epithelial cells produce and secrete the parathyroid hormone (PTH), the major hormone involved in the regulation of blood calcium levels.
The parathyroid glands produce and secrete PTH, a peptide hormone, in response to low blood calcium concentrations (Figure 14.5.2). PTH secretion causes the release of calcium from the bones by stimulating osteoclasts, which secrete enzymes that degrade bone and release calcium into the interstitial fluid. PTH also inhibits osteoblasts, the cells involved in bone deposition, thereby sparing blood calcium. PTH causes increased reabsorption of calcium (and magnesium) in the kidney tubules from the urine filtrate. In addition, PTH initiates the production of the steroid hormone calcitriol (also known as 1,25-dihydroxyvitamin D), which is the active form of vitamin D3, in the kidneys. Calcitriol then stimulates increased absorption of dietary calcium by the intestines. A negative feedback loop regulates the levels of PTH, with rising blood calcium levels inhibiting further release of PTH.
Figure 14.5.2. Parathyroid hormone in maintaining blood calcium homeostasis. Parathyroid hormone increases blood calcium levels when they drop too low. Conversely, calcitonin, which is released from the thyroid gland, decreases blood calcium levels when they become too high. These two mechanisms constantly maintain blood calcium concentration at homeostasis.
Abnormally high activity of the parathyroid gland can cause hyperparathyroidism, a disorder caused by an overproduction of PTH that results in excessive calcium reabsorption from bone. Hyperparathyroidism can significantly decrease bone density, leading to spontaneous fractures or deformities. As blood calcium levels rise, cell membrane permeability to sodium is decreased, and the responsiveness of the nervous system is reduced. At the same time, calcium deposits may collect in the body’s tissues and organs, impairing their functioning.
In contrast, abnormally low blood calcium levels may be caused by parathyroid hormone deficiency, called hypoparathyroidism, which may develop following injury or surgery involving the thyroid gland. Low blood calcium increases membrane permeability to sodium, resulting in muscle twitching, cramping, spasms, or convulsions. Severe deficits can paralyse muscles, including those involved in breathing, and can be fatal.
When blood calcium levels are elevated, calcitonin is produced and secreted by the parafollicular cells of the thyroid gland. As discussed earlier, calcitonin inhibits the activity of osteoclasts, reduces the absorption of dietary calcium in the intestine, and signals the kidneys to reabsorb less calcium, resulting in larger amounts of calcium excreted in the urine.
Section Review
Calcium is required for a variety of important physiologic processes, including neuromuscular functioning; thus, blood calcium levels are closely regulated. The parathyroid glands are small structures located on the posterior thyroid gland that produce parathyroid hormone (PTH), which regulates blood calcium levels. Low blood calcium levels cause the production and secretion of PTH. In contrast, elevated blood calcium levels inhibit secretion of PTH and trigger secretion of the thyroid hormone calcitonin. Underproduction of PTH can result in hypoparathyroidism. In contrast, overproduction of PTH can result in hyperparathyroidism.
Describe the location and structure of the adrenal glands
Identify the hormones produced by the adrenal cortex and adrenal medulla and summarise their target cells and effects
The adrenal glands are wedges of glandular and neuroendocrine tissue adhering to the top of the kidneys by a fibrous capsule (Figure 14.6.1). The adrenal glands have a rich blood supply and experience one of the highest rates of blood flow in the body. They are served by several arteries branching off the aorta, including the suprarenal and renal arteries. Blood flows to each adrenal gland at the adrenal cortex and then drains into the adrenal medulla. Adrenal hormones are released into the circulation via the left and right suprarenal veins.
The adrenal gland consists of an outer cortex of glandular tissue and an inner medulla of nervous tissue. The cortex itself is divided into three zones: the zona glomerulosa, the zona fasciculata, and the zona reticularis. Each region secretes its own set of hormones.
The adrenal cortex, as a component of the hypothalamic-pituitary-adrenal (HPA) axis, secretes steroid hormones important for the regulation of the long-term stress response, blood pressure and blood volume, nutrient uptake and storage, fluid and electrolyte balance, and inflammation. The HPA axis involves the stimulation of hormone release of adrenocorticotropic hormone (ACTH) from the pituitary by the hypothalamus. ACTH then stimulates the adrenal cortex to produce the hormone cortisol. This pathway will be discussed in more detail below.
The adrenal medulla is neuroendocrine tissue composed of postganglionic sympathetic nervous system (SNS) neurons. It is really an extension of the autonomic nervous system, which regulates homeostasis in the body. The sympathomedullary (SAM) pathway involves the stimulation of the medulla by impulses from the hypothalamus via neurons from the thoracic spinal cord. The medulla is stimulated to secrete the amine hormones adrenaline and noradrenaline.
One of the major functions of the adrenal gland is to respond to stress. Stress can be either physical or psychological or both. Physical stresses include exposing the body to injury, walking outside in cold and wet conditions without a coat on, or malnutrition. Psychological stresses include the perception of a physical threat, a fight with a loved one, or just a bad day at school.
The body responds in different ways to short-term stress and long-term stress following a pattern known as the general adaptation syndrome (GAS). Stage one of GAS is called the alarm reaction. This is short-term stress, the fight-or-flight response, mediated by the hormones adrenaline and noradrenaline from the adrenal medulla via the SAM pathway. Their function is to prepare the body for extreme physical exertion. Once this stress is relieved, the body quickly returns to normal. The section on the adrenal medulla covers this response in more detail.
If the stress is not soon relieved, the body adapts to the stress in the second stage called the stage of resistance. If a person is starving for example, the body may send signals to the gastrointestinal tract to maximise the absorption of nutrients from food.
If the stress continues for a longer term however, the body responds with symptoms quite different than the fight-or-flight response. During the stage of exhaustion, individuals may begin to suffer depression, the suppression of their immune response, severe fatigue, or even a fatal heart attack. These symptoms are mediated by the hormones of the adrenal cortex, especially cortisol, released as a result of signals from the HPA axis.
Adrenal hormones also have several non–stress-related functions, including the increase of blood sodium and glucose levels, which will be described in detail below.
Adrenal Cortex
The adrenal cortex consists of multiple layers of lipid-storing cells that occur in three structurally distinct regions. Each of these regions produces different hormones.
Hormones of the Zona Glomerulosa
The most superficial region of the adrenal cortex is the zona glomerulosa, which produces a group of hormones collectively referred to as mineralocorticoids because of their effect on body minerals, especially sodium and potassium. These hormones are essential for fluid and electrolyte balance.
Aldosterone is the major mineralocorticoid. It is important in the regulation of the concentration of sodium and potassium ions in urine, sweat, and saliva. For example, it is released in response to elevated blood K+, low blood Na+, low blood pressure, or low blood volume. In response, aldosterone increases the excretion of K+ and the retention of Na+, which in turn increases blood volume and blood pressure. Its secretion is prompted when CRH from the hypothalamus triggers ACTH release from the anterior pituitary.
Aldosterone is also a key component of the renin-angiotensin-aldosterone system (RAAS) in which specialised cells of the kidneys secrete the enzyme renin in response to low blood volume or low blood pressure. Renin then catalyses the conversion of the blood protein angiotensinogen, produced by the liver, to the hormone angiotensin I. Angiotensin I is converted in the lungs to angiotensin II by angiotensin-converting enzyme (ACE). Angiotensin II has three major functions:
Initiating vasoconstriction of the arterioles, decreasing blood flow
Stimulating kidney tubules to reabsorb NaCl and water, increasing blood volume
Signalling the adrenal cortex to secrete aldosterone, the effects of which further contribute to fluid retention, restoring blood pressure and blood volume
For individuals with hypertension, or high blood pressure, drugs are available that block the production of angiotensin II. These drugs, known as ACE inhibitors, block the ACE enzyme from converting angiotensin I to angiotensin II, thus mitigating the latter’s ability to increase blood pressure.
Hormones of the Zona Fasciculata
The intermediate region of the adrenal cortex is the zona fasciculata, named as such because the cells form small fascicles (bundles) separated by tiny blood vessels. The cells of the zona fasciculata produce hormones called glucocorticoids because of their role in glucose metabolism. The most important of these is cortisol, some of which the liver converts to cortisone. A glucocorticoid produced in much smaller amounts is corticosterone. In response to long-term stressors, the hypothalamus secretes CRH, which in turn triggers the release of ACTH by the anterior pituitary. ACTH triggers the release of the glucocorticoids. Their overall effect is to inhibit tissue building while stimulating the breakdown of stored nutrients to maintain adequate fuel supplies. In conditions of long-term stress, for example, cortisol promotes the catabolism of glycogen to glucose, the catabolism of stored triglycerides into fatty acids and glycerol, and the catabolism of muscle proteins into amino acids. These raw materials can then be used to synthesise additional glucose and ketones for use as body fuels. The hippocampus, which is part of the temporal lobe of the cerebral cortices and important in memory formation, is highly sensitive to stress levels because of its many glucocorticoid receptors.
You are probably familiar with prescription and over-the-counter medications containing glucocorticoids, such as cortisone injections into inflamed joints, prednisone tablets and steroid-based inhalers used to manage severe asthma, and hydrocortisone creams applied to relieve itchy skin rashes. These drugs reflect another role of cortisol—the downregulation of the immune system, which inhibits the inflammatory response.
Hormones of the Zona Reticularis
The deepest region of the adrenal cortex is the zona reticularis, which produces small amounts of a class of steroid sex hormones called androgens. During puberty and most of adulthood, androgens are produced in the gonads. The androgens produced in the zona reticularis supplement the gonadal androgens. They are produced in response to ACTH from the anterior pituitary and are converted in the tissues to testosterone or oestrogens. In adult women, they may contribute to the sex drive, but their function in adult men is not well understood. In post-menopausal women, as the functions of the ovaries decline, the main source of oestrogens becomes the androgens produced by the zona reticularis.
Adrenal Medulla
As noted earlier, the adrenal cortex releases glucocorticoids in response to long-term stress such as severe illness. In contrast, the adrenal medulla releases its hormones in response to acute, short-term stress mediated by the sympathetic nervous system (SNS).
The medullary tissue is composed of unique postganglionic SNS neurons called chromaffin cells, which are large and irregularly shaped, and produce the neurotransmitters adrenaline and noradrenaline. Adrenaline is produced in greater quantities—approximately a 4 to 1 ratio with noradrenaline—and is the more powerful hormone. Because the chromaffin cells release adrenaline and noradrenaline into the systemic circulation, where they travel widely and exert effects on distant cells, they are considered hormones. Derived from the amino acid tyrosine, they are chemically classified as catecholamines.
The secretion of medullary adrenaline and noradrenaline is controlled by a neural pathway that originates from the hypothalamus in response to danger or stress (the SAM pathway). Both adrenaline and noradrenaline signal the liver and skeletal muscle cells to convert glycogen into glucose, resulting in increased blood glucose levels. These hormones increase the heart rate, pulse, and blood pressure to prepare the body to fight the perceived threat or flee from it. In addition, the pathway dilates the airways, raising blood oxygen levels. It also prompts vasodilation, further increasing the oxygenation of important organs such as the lungs, brain, heart, and skeletal muscle. At the same time, it triggers vasoconstriction to blood vessels serving less essential organs such as the gastrointestinal tract, kidneys, and skin, and downregulates some components of the immune system. Other effects include a dry mouth, loss of appetite, pupil dilation, and a loss of peripheral vision. The major hormones of the adrenal glands are summarised in Table 14.6.1.
Table 14.6.1. Hormones of the adrenal glands
Adrenal gland
Associated hormones
Chemical class
Effect
Adrenal cortex
Aldosterone
Steroid
Increases blood Na+ levels
Cortisol, corticosterone, cortisone
Steroid
Increase blood glucose levels
Adrenal medulla
Adrenaline, noradrenaline
Amine
Stimulate fight-or-flight response
Disorders Involving the Adrenal Glands
Several disorders are caused by the dysregulation of the hormones produced by the adrenal glands. For example, Cushing’s disease is a disorder characterised by high blood glucose levels and the accumulation of lipid deposits on the face and neck. It is caused by hypersecretion of cortisol. The most common source of Cushing’s disease is a pituitary tumour that secretes cortisol or ACTH in abnormally high amounts. Other common signs of Cushing’s disease include the development of a moon-shaped face, a buffalo hump on the back of the neck, rapid weight gain, and hair loss. Chronically elevated glucose levels are also associated with an elevated risk of developing type 2 diabetes. In addition to hyperglycaemia, chronically elevated glucocorticoids compromise immunity, resistance to infection, and memory, and can result in rapid weight gain and hair loss.
In contrast, the hyposecretion of corticosteroids can result in Addison’s disease, a rare disorder that causes low blood glucose levels and low blood sodium levels. The signs and symptoms of Addison’s disease are vague and are typical of other disorders as well, making diagnosis difficult. They may include general weakness, abdominal pain, weight loss, nausea, vomiting, sweating and cravings for salty food.
Section Review
The adrenal glands, located superior to each kidney, consist of two regions: the adrenal cortex and adrenal medulla. The adrenal cortex—the outer layer of the gland—produces mineralocorticoids, glucocorticoids, and androgens. The adrenal medulla at the core of the gland produces adrenaline and noradrenaline.
The adrenal glands mediate a short-term stress response and a long-term stress response. A perceived threat results in the secretion of adrenaline and noradrenaline from the adrenal medulla, which mediate the fight-or-flight response. The long-term stress response is mediated by the secretion of CRH from the hypothalamus, which triggers ACTH, which in turn stimulates the secretion of corticosteroids from the adrenal cortex. The mineralocorticoids, chiefly aldosterone, cause sodium and fluid retention, which increases blood volume and blood pressure.
Describe the location and structure of the pineal gland
Discuss the function of melatonin
Recall that the hypothalamus, part of the diencephalon of the brain, sits inferior and somewhat anterior to the thalamus. Inferior but somewhat posterior to the thalamus is the pineal gland, a tiny endocrine gland whose functions are not entirely clear. The pinealocyte cells that make up the pineal gland are known to produce and secrete the amine hormone melatonin, which is derived from serotonin.
The secretion of melatonin varies according to the level of light received from the environment. When photons of light stimulate the retinas of the eyes, a nerve impulse is sent to a region of the hypothalamus called the suprachiasmatic nucleus (SCN), which is important in regulating biological rhythms. From the SCN, the nerve signal is carried to the spinal cord and eventually to the pineal gland, where the production of melatonin is inhibited. As a result, blood levels of melatonin fall, promoting wakefulness. In contrast, as light levels decline—such as during the evening—melatonin production increases, boosting blood levels and causing drowsiness.
The secretion of melatonin may influence the body’s circadian rhythms, the dark-light fluctuations that affect not only sleepiness and wakefulness, but also appetite and body temperature. Interestingly, children have higher melatonin levels than adults, which may prevent the release of gonadotropins from the anterior pituitary, thereby inhibiting the onset of puberty. Finally, an antioxidant role of melatonin is the subject of current research.
Jet lag occurs when a person travels across several time zones and feels sleepy during the day or wakeful at night. Traveling across multiple time zones significantly disturbs the light-dark cycle regulated by melatonin. It can take up to several days for melatonin synthesis to adjust to the light-dark patterns in the new environment, resulting in jet lag. Some air travellers take melatonin supplements to induce sleep.
Section Review
The pineal gland is an endocrine structure of the diencephalon of the brain and is located inferior and posterior to the thalamus. It is made up of pinealocytes. These cells produce and secrete the hormone melatonin in response to low light levels. High blood levels of melatonin induce drowsiness. Jet lag, caused by travelling across several time zones, occurs because melatonin synthesis takes several days to readjust to the light-dark patterns in the new environment.
Identify the most important hormones produced by the testes and ovaries
Name the hormones produced by the placenta and state their functions
This section briefly discusses the hormonal role of the gonads—the male testes and female ovaries—which produce the sex cells (sperm and ova) and secrete the gonadal hormones. The roles of the gonadotropins released from the anterior pituitary (FSH and LH) were discussed earlier.
The primary hormone produced by the male testes is testosterone, a steroid hormone important in the development of the male reproductive system, the maturation of sperm cells, and the development of male secondary sex characteristics such as a deepened voice, body hair, and increased muscle mass. Interestingly, testosterone is also produced in the female ovaries, but at a much reduced level. In addition, the testes produce the peptide hormone inhibin, which inhibits the secretion of FSH from the anterior pituitary gland. FSH stimulates spermatogenesis.
The primary hormones produced by the ovaries are oestrogens, which include oestradiol, oestriol, and oestrone. Oestrogens play an important role in a larger number of physiological processes, including the development of the female reproductive system, regulation of the menstrual cycle, the development of female secondary sex characteristics such as increased adipose tissue and the development of breast tissue, and the maintenance of pregnancy. Another significant ovarian hormone is progesterone, which contributes to regulation of the menstrual cycle and is important in preparing the body for pregnancy as well as maintaining pregnancy. In addition, the granulosa cells of the ovarian follicles produce inhibin, which—as in males—inhibits the secretion of FSH. During the initial stages of pregnancy, an organ called the placenta develops within the uterus. The placenta supplies oxygen and nutrients to the foetus, excretes waste products, and produces and secretes oestrogens and progesterone. The placenta produces human chorionic gonadotropin (hCG) as well. The hCG hormone promotes progesterone synthesis and reduces the mother’s immune function to protect the foetus from immune rejection. It also secretes human placental lactogen (hPL), which plays a role in preparing the breasts for lactation, and relaxin, which is thought to help soften and widen the pubic symphysis in preparation for childbirth. The hormones controlling reproduction are summarised in Table 14.8.1.
Table 14.8.1. Reproductive hormones
Gonad
Associated hormones
Chemical class
Effect
Testes
Testosterone
Steroid
Stimulates development of male secondary sex characteristics and sperm production
Inhibin
Protein
Inhibits FSH release from pituitary
Ovaries
Oestrogens and progesterone
Steroid
Stimulates development of female secondary sex characteristics and prepare the body for childbirth
Placenta
Human chorionic gonadotropin
Protein
Promotes progesterone synthesis during pregnancy and inhibits immune response against foetus
Everyday Connections
Anabolic Steroids – The endocrine system can be exploited for illegal or unethical purposes. A prominent example of this is the use of steroid drugs by professional athletes.
Commonly used for performance enhancement, anabolic steroids are synthetic versions of the male sex hormone, testosterone. By boosting natural levels of this hormone, athletes experience increased muscle mass. Synthetic versions of human growth hormone are also used to build muscle mass.
The use of performance-enhancing drugs is banned by all major collegiate and professional sports organisations in the world because they impart an unfair advantage to athletes who take them. This is regulated by the World Anti-Doping Agency (WADA) which is an international independent agency formed in 1999. Their website provides a list of all substances and methods that are prohibited at all times. In addition, the drugs can cause significant and dangerous side effects. Anabolic steroid use can increase cholesterol levels, raise blood pressure, and damage the liver. Altered testosterone levels (both too low or too high) have been implicated in causing structural damage to the heart and increasing the risk for cardiac arrhythmias, heart attacks, congestive heart failure and sudden death. Paradoxically, steroids can have a feminising effect in males, including shrivelled testicles and enlarged breast tissue. In females, their use can cause masculinising effects such as an enlarged clitoris and growth of facial hair. In both sexes, their use can promote increased aggression (commonly known as “roid-rage”), depression, sleep disturbances, severe acne and infertility.
Section Review
The male and female reproductive system is regulated by follicle-stimulating hormone (FSH) and luteinising hormone (LH) produced by the anterior lobe of the pituitary gland in response to gonadotropin-releasing hormone (GnRH) from the hypothalamus. In males, FSH stimulates sperm maturation, which is inhibited by the hormone inhibin. The steroid hormone testosterone, a type of androgen, is released in response to LH and is responsible for the maturation and maintenance of the male reproductive system, as well as the development of male secondary sex characteristics. In females, FSH promotes egg maturation and LH signals the secretion of the female sex hormones, the oestrogens and progesterone. Both of these hormones are important in the development and maintenance of the female reproductive system, as well as maintaining pregnancy. The placenta develops during early pregnancy and secretes several hormones important for maintaining the pregnancy.
Describe the location and structure of the pancreas, and the morphology and function of the pancreatic islets
Compare and contrast the functions of insulin and glucagon
The pancreas is a long, slender organ, most of which is located posterior to the bottom half of the stomach (Figure 14.9.1). Although it is primarily an exocrine gland, secreting a variety of digestive enzymes, the pancreas has an endocrine function. Its pancreatic islets—clusters of cells formerly known as the islets of Langerhans—secrete the hormones glucagon, insulin, somatostatin and pancreatic polypeptide (PP) among others.
The pancreatic islets each contain five main varieties of cells:
The alpha cell produces the hormone glucagon and makes up approximately 20 percent of each islet. Glucagon plays an important role in blood glucose regulation; low blood glucose levels stimulate its release.
The beta cell produces the hormones insulin and amylin and makes up approximately 75 percent of each islet. Elevated blood glucose levels stimulate the release of insulin.
The delta cell accounts for about four percent of the islet cells and secretes the peptide hormone somatostatin. Recall that somatostatin is also released by the hypothalamus (as GHIH), and the stomach and intestines also secrete it. An inhibiting hormone, pancreatic somatostatin inhibits the release of both glucagon and insulin.
The gamma cell (PP cell) accounts for about one percent of islet cells and secretes the pancreatic polypeptide hormone. It is thought to play a role in appetite, as well as in the regulation of pancreatic exocrine and endocrine secretions. Pancreatic polypeptide released following a meal may reduce further food consumption; however, it is also released in response to fasting.
The epsilon cell produces the hormone ghrelin and accounts for less than 1 percent of islet cells.
Regulation of Blood Glucose Levels by Insulin and Glucagon
Glucose is required for cellular respiration and is the preferred fuel for all body cells. The body derives glucose from the breakdown of the carbohydrate-containing foods and drinks we consume. Glucose not immediately taken up by cells for fuel can be stored by the liver and muscles as glycogen or converted to triglycerides and stored in the adipose tissue. Hormones regulate both the storage and the utilisation of glucose as required. Receptors located in the pancreas sense blood glucose levels, and subsequently the pancreatic cells secrete glucagon or insulin to maintain normal levels.
Glucagon
Receptors in the pancreas can sense the decline in blood glucose levels, such as during periods of fasting or during prolonged labour or exercise (Figure 14.9.2). In response, the alpha cells of the pancreas secrete the hormone glucagon, which has several effects:
It stimulates the liver to convert its stores of glycogen back into glucose. This response is known as glycogenolysis. The glucose is then released into the circulation for use by body cells.
It stimulates the liver to take up amino acids from the blood and convert them into glucose. This response is known as gluconeogenesis.
It stimulates lipolysis, the breakdown of stored triglycerides into free fatty acids and glycerol. Some of the free glycerol released into the bloodstream travels to the liver, which converts it into glucose. This is also a form of gluconeogenesis.
Taken together, these actions increase blood glucose levels. The activity of glucagon is regulated through a negative feedback mechanism; rising blood glucose levels inhibit further glucagon production and secretion.
Figure 14.9.2. Homeostatic regulation of blood glucose levels. Blood glucose concentration is tightly maintained between 70 mg/dL and 110 mg/dL (or between 4.0 mmol/L and 7.8 mmol/L). If blood glucose concentration rises above this range, insulin is released, which stimulates body cells to remove glucose from the blood. If blood glucose concentration drops below this range, glucagon is released, which stimulates body cells to release glucose into the blood.
Insulin
The primary function of insulin is to facilitate the uptake of glucose into body cells. Red blood cells, as well as cells of the brain, liver, kidneys, and the lining of the small intestine, do not have insulin receptors on their cell membranes and do not require insulin for glucose uptake. Although all other body cells do require insulin if they are to take glucose from the bloodstream, skeletal muscle cells and adipose cells are the primary targets of insulin.
The presence of food in the intestine triggers the release of gastrointestinal tract hormones such as glucose-dependent insulinotropic peptide (previously known as gastric inhibitory peptide). This is in turn the initial trigger for insulin production and secretion by the beta cells of the pancreas. Once nutrient absorption occurs, the resulting surge in blood glucose levels further stimulates insulin secretion.
Precisely how insulin facilitates glucose uptake is not entirely clear. However, insulin appears to activate a tyrosine kinase receptor, triggering the phosphorylation of many substrates within the cell. These multiple biochemical reactions converge to support the movement of intracellular vesicles containing facilitative glucose transporters to the cell membrane. In the absence of insulin, these transport proteins are normally recycled slowly between the cell membrane and cell interior. Insulin triggers the rapid movement of a pool of glucose transporter vesicles to the cell membrane, where they fuse and expose the glucose transporters to the extracellular fluid. The transporters then move glucose by facilitated diffusion into the cell interior.
Insulin also reduces blood glucose levels by stimulating glycolysis, the metabolism of glucose for generation of ATP. Moreover, it stimulates the liver to convert excess glucose into glycogen for storage, and it inhibits enzymes involved in glycogenolysis and gluconeogenesis. Finally, insulin promotes triglyceride and protein synthesis. The secretion of insulin is regulated through a negative feedback mechanism. As blood glucose levels decrease, further insulin release is inhibited. The pancreatic hormones are summarised in Table 14.9.1.
Table 14.9.1. Hormones of the pancreas
Associated hormones
Chemical class
Effect
Glucagon (alpha cells)
Protein
Increases blood glucose levels
Insulin (beta cells)
Protein
Reduces blood glucose levels
Somatostatin (delta cells)
Protein
Inhibits insulin and glucagon release
Pancreatic polypeptide (gamma or PP cells)
Protein
Role in appetite
Ghrelin (epsilon cells)
Protein
Stimulates hunger
Disorders of the Endocrine System: Diabetes Mellitus
Dysfunction of insulin production and secretion, as well as the target cells’ responsiveness to insulin, can lead to a condition called diabetes mellitus. An increasingly common disease, diabetes mellitus has been diagnosed in approximately 463 million adults and 1.1 million children and adolescents under 19 years of age worldwide. In Australia, almost 1.2 million people (~5% of the total population) had been diagnosed with diabetes mellitus. In the United States, 18 million adults and more than 200,000 children were diagnosed with this disease. It is estimated that up to 7 million more adults have the condition but have not been diagnosed. In addition, approximately 79 million people in the US are estimated to have pre-diabetes, a condition in which blood glucose levels are abnormally high, but not yet high enough to be classified as diabetes.There are two main forms of diabetes mellitus. Type 1 diabetes is an autoimmune disease affecting the beta cells of the pancreas. Certain genes are recognised to increase susceptibility. The beta cells of people with type 1 diabetes do not produce insulin; thus, synthetic insulin must be administered by injection or infusion. This form of diabetes accounts for less than five percent of all diabetes cases.Type 2 diabetes accounts for approximately 95 percent of all cases. It is acquired, and lifestyle factors such as poor diet, inactivity, and the presence of pre-diabetes greatly increase a person’s risk. About 80 to 90 percent of people with type 2 diabetes are overweight or obese. In type 2 diabetes, cells become resistant to the effects of insulin. In response, the pancreas increases its insulin secretion, but over time, the beta cells become exhausted. In many cases, type 2 diabetes can be reversed by moderate weight loss, regular physical activity, and consumption of a healthy diet; however, if blood glucose levels cannot be controlled, the diabetic will eventually require insulin.Two of the early manifestations of diabetes are excessive urination and excessive thirst. They demonstrate how the out-of-control levels of glucose in the blood affect kidney function. The kidneys are responsible for filtering glucose from the blood. Excessive blood glucose draws water into the urine, and as a result the person eliminates an abnormally large quantity of sweet urine. The use of body water to dilute the urine leaves the body dehydrated, and so the person is unusually and continually thirsty. The person may also experience persistent hunger because the body cells are unable to access the glucose in the bloodstream.Over time, persistently high levels of glucose in the blood injure tissues throughout the body, especially those of the blood vessels and nerves. Inflammation and injury of the lining of arteries lead to atherosclerosis and an increased risk of heart attack and stroke. Damage to the microscopic blood vessels of the kidney impairs kidney function and can lead to kidney failure. Damage to blood vessels that serve the eyes can lead to blindness. Blood vessel damage also reduces circulation to the limbs, whereas nerve damage leads to a loss of sensation, called neuropathy, particularly in the hands and feet. Together, these changes increase the risk of injury, infection, and tissue death (necrosis), contributing to a high rate of toe, foot, and lower leg amputations in people with diabetes. Uncontrolled diabetes can also lead to a dangerous form of metabolic acidosis called ketoacidosis. Deprived of glucose, cells increasingly rely on fat stores for fuel. However, in a glucose-deficient state, the liver is forced to use an alternative lipid metabolism pathway that results in the increased production of ketone bodies (or ketones), which are acidic. The build-up of ketones in the blood causes ketoacidosis, which—if left untreated—may lead to a life-threatening “diabetic coma.” Together, these complications make diabetes the seventh leading cause of death in the United States.Diabetes is diagnosed when lab tests reveal that blood glucose concentrations are higher than normal, a condition called hyperglycaemia. The treatment of diabetes depends on the type, the severity of the condition, and the ability of the patient to make lifestyle changes. As noted earlier, moderate weight loss, regular physical activity, and consumption of a healthful diet can reduce blood glucose levels. Some patients with type 2 diabetes may be unable to control their disease with these lifestyle changes and will require medication. Historically, the first-line treatment of type 2 diabetes was insulin. Research advances have resulted in alternative options, including medications that enhance pancreatic function.
Section Review
The pancreas has both exocrine and endocrine functions. The pancreatic islet cell types include alpha cells, which produce glucagon; beta cells, which produce insulin; delta cells, which produce somatostatin; gamma cell which produce pancreatic polypeptide and epsilon cells which produce ghrelin. Insulin and glucagon are involved in the regulation of glucose metabolism. Insulin is produced by the beta cells in response to high blood glucose levels. It enhances glucose uptake and utilisation by target cells, as well as the storage of excess glucose for later use. Dysfunction of the production of insulin or target cell resistance to the effects of insulin causes diabetes mellitus, a disorder characterised by high blood glucose levels. The hormone glucagon is produced and secreted by the alpha cells of the pancreas in response to low blood glucose levels. Glucagon stimulates mechanisms that increase blood glucose levels, such as the catabolism of glycogen into glucose.
Identify the organs with a secondary endocrine function, the hormone they produce and its effects
In your study of anatomy and physiology, you have already encountered a few of the many organs of the body that have secondary endocrine functions. Here, you will learn about the hormone-producing activities of the heart, gastrointestinal tract, kidneys, skeleton, adipose tissue, skin and thymus.
Heart
When the body experiences an increase in blood volume or pressure, the cells of the heart’s atrial wall stretch. In response, specialised cells in the wall of the atria produce and secrete the peptide hormone atrial natriuretic peptide (ANP). ANP signals the kidneys to reduce sodium reabsorption, thereby decreasing the amount of water reabsorbed from the urine filtrate and reducing blood volume. Other actions of ANP include the inhibition of renin secretion and the initiation of the renin-angiotensin-aldosterone system (RAAS) and vasodilation. Therefore, ANP aids in decreasing blood pressure, blood volume and blood sodium levels.
Gastrointestinal Tract
The endocrine cells of the GI tract are located in the mucosa of the stomach and small intestine. Some of these hormones are secreted in response to eating a meal and aid in digestion. An example of a hormone secreted by the stomach cells is gastrin, a peptide hormone secreted in response to stomach distention that stimulates the release of hydrochloric acid. Secretin is a peptide hormone secreted by the small intestine as acidic chyme (partially digested food and fluid) moves from the stomach into the duodenum. It stimulates the release of bicarbonate from the pancreas, which buffers the acidic chyme, and inhibits the further secretion of hydrochloric acid by the stomach. Cholecystokinin (CCK) is another peptide hormone released from the small intestine. It promotes the secretion of pancreatic enzymes and the release of bile from the gallbladder, both of which facilitate digestion. Other hormones produced by the intestinal cells aid in glucose metabolism, such as by stimulating the pancreatic beta cells to secrete insulin, reducing glucagon secretion from the alpha cells, or enhancing cellular sensitivity to insulin.
Kidneys
The kidneys participate in several complex endocrine pathways and produce several hormones. A decline in blood flow to the kidneys stimulates them to release the enzyme renin, triggering the renin-angiotensin-aldosterone (RAAS) system, and stimulating the reabsorption of sodium and water. The reabsorption increases blood flow and blood pressure. The kidneys also play a role in regulating blood calcium levels through the production of calcitriol from vitamin D3, which is released in response to the secretion of parathyroid hormone (PTH). In addition, the kidneys produce the hormone erythropoietin (EPO) in response to low oxygen levels. EPO stimulates the production of red blood cells (erythrocytes) in the bone marrow, thereby increasing oxygen delivery to tissues. You may have heard of EPO as a performance-enhancing drug (in a synthetic form).
Skeleton
Although bone has long been recognised as a target for hormones, only recently have researchers recognised that the skeleton itself produces at least two hormones. Fibroblast growth factor 23 (FGF23) is produced by bone cells in response to increased blood levels of vitamin D3 or phosphate. It triggers the kidneys to inhibit the formation of calcitriol from vitamin D3 and to increase phosphorus excretion. Osteocalcin, produced by osteoblasts, stimulates the pancreatic beta cells to increase insulin production. It also acts on peripheral tissues to increase their sensitivity to insulin and their utilisation of glucose.
Adipose Tissue
Adipose tissue produces and secretes several hormones involved in lipid metabolism and storage. One important example is leptin, a protein manufactured by adipose cells that circulates in amounts directly proportional to levels of body fat. Leptin is released in response to food consumption and acts by binding to brain neurons involved in energy intake and expenditure. Binding of leptin produces a feeling of satiety after a meal, thereby reducing appetite. It also appears that the binding of leptin to brain receptors triggers the sympathetic nervous system to regulate bone metabolism, increasing deposition of cortical bone. Adiponectin—another hormone synthesised by adipose cells—appears to reduce cellular insulin resistance and to protect blood vessels from inflammation and atherosclerosis. Its levels are lower in people who are obese and rise following weight loss.
Skin
The skin functions as an endocrine organ in the production of the inactive form of vitamin D3, cholecalciferol. When cholesterol present in the epidermis is exposed to ultraviolet radiation, it is converted to cholecalciferol, which then enters the blood. In the liver, cholecalciferol is converted to an intermediate that travels to the kidneys and is further converted to calcitriol, the active form of vitamin D3. Vitamin D is important in a variety of physiological processes, including intestinal calcium absorption and immune system function. In some studies, low levels of vitamin D have been associated with increased risks of cancer, severe asthma, and multiple sclerosis. Vitamin D deficiency in children causes rickets, and in adults, osteomalacia—both of which are characterised by bone deterioration.
Thymus
The thymus is an organ of the immune system that is larger and more active during infancy and early childhood and begins to atrophy as we age. Its endocrine function is the production of a group of hormones called thymosins that contribute to the development and differentiation of T lymphocytes, which are immune cells. Although the role of thymosins is not yet well understood, it is clear that they contribute to the immune response. Thymosins have been found in tissues other than the thymus and have a wide variety of functions, so the thymosins cannot be strictly categorised as thymic hormones.
Liver
The liver is responsible for secreting at least four important hormones or hormone precursors: insulin-like growth factor (somatomedin), angiotensinogen, thrombopoietin, and hepcidin. Insulin-like growth factor-1 is the immediate stimulus for growth in the body, especially of the bones. Angiotensinogen is the precursor to angiotensin, mentioned earlier, which increases blood pressure. Thrombopoietin stimulates the production of the blood’s platelets. Hepcidins block the release of iron from cells in the body, helping to regulate iron homeostasis in our body fluids. The major hormones of these other organs are summarised in Table 14.10.1.
Table 14.10.1. Organs with secondary endocrine functions and their major hormones
Organ
Major hormones
Effects
Heart
Atrial natriuretic peptide (ANP)
Reduces blood volume, blood pressure, and Na+ concentration
Gastrointestinal tract
Gastrin, secretin and cholecystokinin,
Aid in digestion of food and buffering of stomach acids
glucose-dependent insulinotropic peptide (GIP) and glucagon-like peptide 1 (GLP-1)
Stimulate beta cells of the pancreas to release insulin
Kidneys
Renin
Enzyme that begins the RAAS and allows stimulation for release of aldosterone
Calcitriol
Aids in the reabsorption of Ca 2+
Erythropoietin
Triggers formation of red blood cells in the bone marrow
Skeleton
FGF23
Inhibits production of calcitriol and increases phosphate excretion
osteocalcin
Increases insulin production
Adipose tissue
Leptin
Promotes satiety signals in the brain
Adiponectin
Reduces insulin resistance
Skin
Cholecalciferol
Modifies to form vitamin D
Thymus (and other organs)
Thymosin
Among other things, aids in the development of T lymphocytes of the immune system
Liver
Insulin-like growth factor-1
Stimulates bodily growth
Angiotensinogen
Raises blood pressure
Thrombopoietin
Causes increase in platelets
Hepcidin
Blocks release of iron into the body fluids
Section Review
Some organs have a secondary endocrine function. For example, the walls of the atria of the heart produce the hormone atrial natriuretic peptide (ANP), the gastrointestinal tract produces the hormones gastrin, secretin, and cholecystokinin, which aid in digestion and the kidneys produce erythropoietin (EPO), which stimulates the formation of red blood cells. Even bone, adipose tissue and the skin have secondary endocrine functions.
14.11 Development and Ageing of the Endocrine System
Learning Objectives
By the end of this section, you will be able to:
Describe the embryonic origins of the endocrine system
Discuss the effects of ageing on the endocrine system
The endocrine system arises from all three embryonic germ layers. The endocrine glands that produce the steroid hormones, such as the gonads and adrenal cortex, arise from the mesoderm. In contrast, endocrine glands that arise from the endoderm and ectoderm produce the amine, peptide and protein hormones. The pituitary gland arises from two distinct areas of the ectoderm: the anterior pituitary gland arises from the oral ectoderm, whereas the posterior pituitary gland arises from the neural ectoderm at the base of the hypothalamus. The pineal gland also arises from the ectoderm. The two structures of the adrenal glands arise from two different germ layers: the adrenal cortex from the mesoderm and the adrenal medulla from ectoderm neural cells. The endoderm gives rise to the thyroid and parathyroid glands, as well as the pancreas and the thymus.
As the body ages, changes occur that affect the endocrine system, sometimes altering the production, secretion, and catabolism of hormones. For example, the structure of the anterior pituitary gland changes as vascularisation decreases and the connective tissue content increases with increasing age. This restructuring affects the gland’s hormone production. For example, the amount of human growth hormone that is produced declines with age, resulting in the reduced muscle mass commonly observed in the elderly.
The adrenal glands also undergo changes as the body ages; as fibrous tissue increases, the production of cortisol and aldosterone decreases. Interestingly, the production and secretion of adrenaline and noradrenaline remain normal throughout the ageing process.
A well-known example of the ageing process affecting an endocrine gland is menopause and the decline of ovarian function. With increasing age, the ovaries decrease in both size and weight and become progressively less sensitive to gonadotropins. This gradually causes a decrease in oestrogen and progesterone levels, leading to menopause and the inability to reproduce. Low levels of oestrogens and progesterone are also associated with some disease states, such as osteoporosis, atherosclerosis and hyperlipidaemia, or abnormal blood lipid levels.
Testosterone levels also decline with age, a condition called andropause (or viropause); however, this decline is much less dramatic than the decline of oestrogens in women, and much more gradual, rarely affecting sperm production until very old age. Although this means that males maintain their ability to father children for decades longer than females, the quantity, quality and motility of their sperm is often reduced.
As the body ages, the thyroid gland produces less of the thyroid hormones, causing a gradual decrease in the basal metabolic rate. The lower metabolic rate reduces the production of body heat and increases levels of body fat. Parathyroid hormones, on the other hand, increase with age. This may be because of reduced dietary calcium levels, causing a compensatory increase in parathyroid hormone. However, increased parathyroid hormone levels combined with decreased levels of calcitonin (and oestrogens in women) can lead to osteoporosis as PTH stimulates demineralisation of bones to increase blood calcium levels. Notice that osteoporosis is common in both elderly males and females.
Increasing age also affects glucose metabolism, as blood glucose levels spike more rapidly and take longer to return to normal in the elderly. In addition, increasing glucose intolerance may occur because of a gradual decline in cellular insulin sensitivity. Almost 27 percent of Americans aged 65 and older have diabetes and fewer than 5% of Australians (of all ages) self-reported as having diabetes in 2017-2018 (AIHW, 2020).
Section Review
The endocrine system originates from all three germ layers of the embryo, including the endoderm, ectoderm, and mesoderm. In general, different hormone classes arise from distinct germ layers. Ageing affects the endocrine glands, potentially affecting hormone production and secretion, and can cause disease. The production of hormones, such as human growth hormone, cortisol, aldosterone, sex hormones and the thyroid hormones, decreases with age.
15.1 Anatomy and Physiology of the Male Reproductive System
Learning Objectives
By the end of this section, you will be able to:
Describe the structure and function of the organs of the male reproductive system
Describe the structure and function of the sperm cell
Explain the events during spermatogenesis that produce haploid sperm from diploid cells
Identify the importance of testosterone in male reproductive function
Unique for its role in human reproduction, a gamete is a specialised sex cell carrying 23 chromosomes—one half the number in body cells. At fertilisation, the chromosomes in one male gamete, called a sperm (or spermatozoon), combine with the chromosomes in one female gamete, called an oocyte. The function of the male reproductive system (Figure 15.1.1) is to produce sperm and transfer them to the female reproductive tract. The paired testes are a crucial component in this process, as they produce both sperm and androgens, the hormones that support male reproductive physiology. In humans, the most important male androgen is testosterone. Several accessory organs and ducts aid the process of sperm maturation and transport the sperm and other seminal components to the penis, which delivers sperm to the female reproductive tract. In this section, we examine each of these different structures and discuss the process of sperm production and transport.
Figure 15.1.1. Male reproductive system. The structures of the male reproductive system include the testes, the epididymides, the penis, and the ducts and glands that produce and carry semen. Sperm exit the scrotum through the ductus deferens, which is bundled in the spermatic cord. The seminal vesicles and prostate gland add fluids to the sperm to create semen.
Scrotum
The testes are located in a skin-covered, highly pigmented, muscular sack called the scrotum that extends from the body behind the penis (see Figure 15.1.1). This location is important in sperm production, which occurs within the testes, and proceeds more efficiently when the testes are kept 2 to 4°C below core body temperature.
The dartos muscle makes up the subcutaneous muscle layer of the scrotum (Figure 15.1.2). It continues internally to make up the scrotal septum, a wall that divides the scrotum into two compartments, each housing one testis. Descending from the internal oblique muscle of the abdominal wall are the two cremaster muscles, which cover each testis like a muscular net. By contracting simultaneously, the dartos and cremaster muscles can elevate the testes in cold weather (or water), moving the testes closer to the body and decreasing the surface area of the scrotum to retain heat. Alternatively, as the environmental temperature increases, the scrotum relaxes, moving the testes farther from the body core and increasing scrotal surface area, which promotes heat loss. Externally, the scrotum has a raised medial thickening on the surface called the raphae.
Figure 15.1.2. The scrotum and testes. This anterior view shows the structures of the scrotum and testes.
Testes
The testes (singular = testis) are the male gonads—that is, the male reproductive organs. They produce both sperm and androgens, such as testosterone, and are active throughout the reproductive lifespan of the male.
Paired ovals, the testes are each approximately 4 to 5 cm in length and are housed within the scrotum (see Figure 15.1.2). They are surrounded by two distinct layers of protective connective tissue (Figure 15.1.3). The outer tunica vaginalis is a serous membrane that has both a parietal and a thin visceral layer. Beneath the tunica vaginalis is the tunica albuginea, a tough, white, dense connective tissue layer covering the testis itself. Not only does the tunica albuginea cover the outside of the testis, it also invaginates to form septa that divide the testis into 300 to 400 structures called lobules. Within the lobules, sperm develop in structures called seminiferous tubules. During the seventh month of the developmental period of a male foetus, each testis moves through the abdominal musculature to descend (via the inguinal canal) into the scrotal cavity. This is called the “descent of the testis.” Cryptorchidism is the clinical term used when one or both testes fail to descend into the scrotum prior to birth.
Figure 15.1.3. Anatomy of the testis. This sagittal view shows the seminiferous tubules, the site of sperm production. Formed sperm are transferred to the epididymis, where they mature. They leave the epididymis during an ejaculation via the ductus deferens.
The tightly coiled seminiferous tubules form the bulk of each testis. They are composed of developing sperm cells surrounding a lumen, the hollow centre of the tubule, where formed sperm are released into the duct system of the testis. Specifically, from the lumens of the seminiferous tubules, sperm move into the straight tubules (or tubuli recti), and from there into a fine meshwork of tubules called the rete testes. Sperm leave the rete testes, and the testis itself, through the 15 to 20 efferent ductules that cross the tunica albuginea.
Inside the seminiferous tubules are six different cell types. These include supporting cells called sustentacular (Sertoli) cells, as well as five types of developing sperm cells called germ cells. Germ cell development progresses from the basement membrane—at the perimeter of the tubule—toward the lumen. Let’s look more closely at these cell types.
Sertoli Cells
Surrounding all stages of the developing sperm cells are elongate, branching Sertoli cells. Sertoli cells are a type of supporting cell called a sustentacular cell, or sustenocyte, that are typically found in epithelial tissue. Sertoli cells secrete signalling molecules that promote sperm production and can control whether germ cells live or die. They extend physically around the germ cells from the peripheral basement membrane of the seminiferous tubules to the lumen. Tight junctions between these sustentacular cells create the blood–testis barrier, which keeps bloodborne substances from reaching the germ cells and, at the same time, keeps surface antigens on developing germ cells from escaping into the bloodstream and prompting an autoimmune response.
Germ Cells
The least mature cells, the spermatogonia (singular = spermatogonium), line the basement membrane inside the tubule. Spermatogonia are the stem cells of the testis, which means that they are still able to differentiate into a variety of different cell types throughout adulthood. Spermatogonia divide to produce primary and secondary spermatocytes, then spermatids, which finally produce formed sperm. The process that begins with spermatogonia and concludes with the production of sperm is called spermatogenesis.
Spermatogenesis
As just noted, spermatogenesis occurs in the seminiferous tubules that form the bulk of each testis (see Figure 15.1.3). The process begins at puberty, after which time sperm are produced constantly throughout a man’s life. One production cycle, from spermatogonia through formed sperm, takes approximately 64 days. A new cycle starts approximately every 16 days, although this timing is not synchronous across the seminiferous tubules. Sperm counts—the total number of sperm a man produces—slowly decline after age 35 and some studies suggest that smoking can lower sperm counts irrespective of age.
The process of spermatogenesis begins with mitosis of the diploid spermatogonia (Figure 15.1.4). Because these cells are diploid (2n), they each have a complete copy of the father’s genetic material, or 46 chromosomes. However, mature gametes are haploid (1n), containing 23 chromosomes—meaning that daughter cells of spermatogonia must undergo a second cellular division through the process of meiosis.
Two identical diploid cells result from spermatogonia mitosis. One of these cells remains a spermatogonium, and the other becomes a primary spermatocyte, the next stage in the process of spermatogenesis. As in mitosis, DNA is replicated in a primary spermatocyte, and the cell undergoes cell division to produce two cells with identical chromosomes. Each of these is a secondary spermatocyte. Now a second round of cell division occurs in both secondary spermatocytes, separating the chromosome pairs. This second meiotic division results in a total of four cells with only half of the number of chromosomes. Each of these new cells is a spermatid. Although haploid, early spermatids look remarkably similar to cells in the earlier stages of spermatogenesis, with a round shape, central nucleus, and large amount of cytoplasm. A process called spermiogenesis transforms these early spermatids, reducing the cytoplasm, and beginning the formation of the parts of a true sperm. The fifth stage of germ cell formation—spermatozoa, or formed sperm—is the end result of this process, which occurs in the portion of the tubule nearest the lumen. Eventually, the sperm are released into the lumen and are moved along a series of ducts in the testis toward a structure called the epididymis for the next step of sperm maturation.
Structure of Formed Sperm
Sperm are smaller than most cells in the body; in fact, the volume of a sperm cell is 85,000 times less than that of the female gamete. Approximately 100 to 300 million sperm are produced each day, whereas women typically ovulate only one oocyte per month as is true for most cells in the body, the structure of sperm cells speaks to their function. Sperm have a distinctive head, mid-piece, and tail region (Figure 15.1.5). The head of the sperm contains the extremely compact haploid nucleus with very little cytoplasm. These qualities contribute to the overall small size of the sperm (the head is only 5 μm long). A structure called the acrosome covers most of the head of the sperm cell as a “cap” that is filled with lysosomal enzymes important for preparing sperm to participate in fertilisation. Tightly packed mitochondria fill the mid-piece of the sperm. ATP produced by these mitochondria will power the flagellum, which extends from the neck and the mid-piece through the tail of the sperm, enabling it to move the entire sperm cell. The central strand of the flagellum, the axial filament, is formed from one centriole inside the maturing sperm cell during the final stages of spermatogenesis.
Figure 15.1.5. Structure of sperm. Sperm cells are divided into a head, containing DNA; a mid-piece, containing mitochondria; and a tail, providing motility. The acrosome is oval and somewhat flattened.
Sperm Transport
To fertilise an egg, sperm must be moved from the seminiferous tubules in the testes, through the epididymis and—later during ejaculation—along the length of the penis and out into the female reproductive tract.
Role of the Epididymis
From the lumen of the seminiferous tubules, the immotile sperm are surrounded by testicular fluid and moved to the epididymis (plural = epididymides), a coiled tube attached to the testis where newly formed sperm continue to mature (see Figure 15.1.3). Though the epididymis does not take up much room in its tightly coiled state, it would be approximately 6 m long if straightened. It takes an average of 12 days for sperm to move through the coils of the epididymis, with the shortest recorded transit time in humans being one day. Sperm enter the head of the epididymis and are moved along predominantly by the contraction of smooth muscles lining the epididymal tubes. As they are moved along the length of the epididymis, the sperm further mature and acquire the ability to move under their own power. Once inside the female reproductive tract, they will use this ability to move independently toward the unfertilised egg. The more mature sperm are then stored in the tail of the epididymis (the final section) until ejaculation occurs.
Ductal System
During ejaculation, sperm exit the tail of the epididymis and are pushed by smooth muscle contraction to the ductus deferens (also called the vas deferens). The ductus deferens is a thick, muscular tube that is bundled together inside the scrotum with connective tissue, blood vessels, and nerves into a structure called the spermatic cord (see Figure 15.1.1 and Figure 15.1.2). Because the ductus deferens is physically accessible within the scrotum, surgical sterilisation to interrupt sperm delivery can be performed by cutting and sealing a small section of the ductus (vas) deferens. This procedure is called a vasectomy and it is an effective form of male birth control. Although it may be possible to reverse a vasectomy, clinicians consider the procedure permanent and advise men to undergo it only if they are certain they no longer wish to father children.
From each epididymis, each ductus deferens extends superiorly into the abdominal cavity through the inguinal canal in the abdominal wall. From here, the ductus deferens continues posteriorly to the pelvic cavity, ending posterior to the bladder where it dilates in a region called the ampulla (meaning “flask”).
Sperm make up only five percent of the final volume of semen, the thick, milky fluid that the male ejaculates. The bulk of semen is produced by three critical accessory glands of the male reproductive system: the seminal vesicles, the prostate and the bulbourethral glands.
Seminal Vesicles
As sperm pass through the ampulla of the ductus deferens at ejaculation, they mix with fluid from the associated seminal vesicle (see Figure 15.1.1). The paired seminal vesicles are glands that contribute approximately 60 percent of the semen volume. Seminal vesicle fluid contains substantial amounts of fructose, which is used by the sperm mitochondria to generate ATP to allow movement through the female reproductive tract.
The fluid, now containing both sperm and seminal vesicle secretions, next moves into the associated ejaculatory duct, a short structure formed from the ampulla of the ductus deferens and the duct of the seminal vesicle. The paired ejaculatory ducts transport the seminal fluid into the next structure, the prostate gland.
Prostate Gland
As shown in Figure 15.1.1, the centrally located prostate gland sits anterior to the rectum at the base of the bladder surrounding the prostatic urethra (the portion of the urethra that runs within the prostate). About the size of a walnut, the prostate is formed of both muscular and glandular tissues. It excretes an alkaline, milky fluid to the passing seminal fluid—now called semen—that is critical to first coagulate and then de-coagulate the semen following ejaculation. The temporary thickening of semen helps retain it within the female reproductive tract, providing time for sperm to utilise the fructose provided by seminal vesicle secretions. When the semen regains its fluid state, sperm can then pass farther into the female reproductive tract.
The prostate normally doubles in size during puberty. At approximately age 25, it gradually begins to enlarge again. This enlargement does not usually cause problems; however, abnormal growth of the prostate, or benign prostatic hyperplasia (BPH), can cause constriction of the urethra as it passes through the middle of the prostate gland, leading to a number of lower urinary tract symptoms, such as a frequent and intense urge to urinate, a weak stream and a sensation that the bladder has not emptied completely. By age 60, approximately 40 percent of men have some degree of BPH. By age 80, the number of affected individuals can increase to as many as 80 percent. Treatments for BPH attempt to relieve the pressure on the urethra so that urine can flow more normally. Mild to moderate symptoms are treated with medication, whereas severe enlargement of the prostate is treated by surgery in which a portion of the prostate tissue is removed.
Another common disorder involving the prostate is prostate cancer. According to the Centres for Disease Control and Prevention (CDC), prostate cancer is the second most common cancer in men in the US and is the most commonly diagnosed cancer in males in Australia (AIHW, 2019). However, some forms of prostate cancer grow very slowly and thus may not ever require treatment. Aggressive forms of prostate cancer, in contrast, involve metastasis to vulnerable organs like the lungs and brain. There is no link between BPH and prostate cancer, but the symptoms are similar. Prostate cancer is detected by a medical history, a blood test, and a rectal exam that allows physicians to palpate the prostate and check for unusual masses. If a mass is detected, the cancer diagnosis is confirmed by biopsy of the cells.
Bulbourethral Glands
The final addition to semen is made by two bulbourethral glands (or Cowper’s glands) that release a thick, salty, alkaline fluid that lubricates the end of the urethra and the vagina and helps to clean and neutralise urine residues from the penile urethra. The fluid from these accessory glands is released after the male becomes sexually aroused, and shortly before the release of the semen. It is therefore sometimes called pre-ejaculate. It is important to note that, in addition to the lubricating proteins, it is possible for bulbourethral fluid to pick up sperm already present in the urethra and therefore it may be able to cause pregnancy.
The Penis
The penis is the male organ of copulation (sexual intercourse). It is flaccid for non-sexual actions, such as urination, and turgid and rod-like with sexual arousal. When erect, the stiffness of the organ allows it to penetrate the vagina and deposit semen into the female reproductive tract.
Figure 15.1.6. Cross-sectional anatomy of the penis. Three columns of erectile tissue make up most of the volume of the penis.
The shaft of the penis surrounds the urethra (Figure 15.1.6). The shaft is composed of three column-like chambers of erectile tissue that span the length of the shaft. Each of the two larger lateral chambers is called a corpus cavernosum (plural = corpora cavernosa). Together, these make up the bulk of the penis. The corpus spongiosum, which can be felt as a raised ridge on the erect penis, is a smaller chamber that surrounds the spongy, or penile, urethra. The end of the penis, called the glans penis, has a high concentration of nerve endings, resulting in extremely sensitive skin that influences the likelihood of ejaculation (see Figure 15.1.1). The skin from the shaft extends down over the glans and forms a collar called the prepuce (or foreskin). The foreskin also contains a dense concentration of nerve endings, and both lubricate and protect the sensitive skin of the glans penis. A surgical procedure called circumcision, often performed for religious or social reasons, removes the prepuce, typically within days of birth.
Both sexual arousal and REM sleep (during which dreaming occurs) can induce an erection. Penile erections are the result of vasocongestion, or engorgement of the tissues because of more arterial blood flowing into the penis than is leaving in the veins. During sexual arousal, nitric oxide (NO) is released from nerve endings near blood vessels within the corpora cavernosa and spongiosum. Release of NO activates a signalling pathway that results in relaxation of the smooth muscles that surround the penile arteries, causing them to dilate. This dilation increases the amount of blood that can enter the penis and induces the endothelial cells in the penile arterial walls to also secrete NO and perpetuate the vasodilation. The rapid increase in blood volume fills the erectile chambers, and the increased pressure of the filled chambers compresses the thin-walled penile venules, preventing venous drainage of the penis. The result of this increased blood flow to the penis and reduced blood return from the penis is erection. Depending on the flaccid dimensions of a penis, it can increase in size slightly or greatly during erection, with the average length of an erect penis measuring approximately 15 cm.
Disorders of the Male Reproductive System
Erectile dysfunction (ED) is a condition in which a man has difficulty either initiating or maintaining an erection. The combined prevalence of minimal, moderate, and complete ED is approximately 40 percent in men at age 40 and reaches nearly 70 percent by 70 years of age. In addition to ageing, ED is associated with diabetes, vascular disease, psychiatric disorders, prostate disorders, the use of some drugs such as certain antidepressants, and problems with the testes resulting in low testosterone concentrations. These physical and emotional conditions can lead to interruptions in the vasodilation pathway and result in an inability to achieve an erection.
Recall that the release of NO induces relaxation of the smooth muscles that surround the penile arteries, leading to the vasodilation necessary to achieve an erection. To reverse the process of vasodilation, an enzyme called phosphodiesterase (PDE) degrades a key component of the NO signalling pathway called cGMP. There are several different forms of this enzyme, and PDE type 5 is the type of PDE found in the tissues of the penis. Scientists discovered that inhibiting PDE5 increases blood flow and allows vasodilation of the penis to occur.
PDEs and the vasodilation signalling pathway are found in the vasculature in other parts of the body. In the 1990s, clinical trials of a PDE5 inhibitor called sildenafil were initiated to treat hypertension and angina pectoris (chest pain caused by poor blood flow through the heart). The trial showed that the drug was not effective at treating heart conditions, but many men experienced erection and priapism (erection lasting longer than 4 hours). Because of this, a clinical trial was started to investigate the ability of sildenafil to promote erections in men suffering from ED. In 1998, the FDA approved the drug, marketed as Viagra®. Since approval of the drug, sildenafil and similar PDE inhibitors now generate over a billion dollars a year in sales and are reported to be effective in treating approximately 70 to 85 percent of cases of ED. Importantly, men with health problems—especially those with cardiac disease taking nitrates—should avoid Viagra or talk to their physician to find out if they are a candidate for the use of this drug, as deaths have been reported for at-risk users.
Testosterone
Testosterone, an androgen, is a steroid hormone produced by Leydig cells. The alternate term for Leydig cells, interstitial cells, reflects their location between the seminiferous tubules in the testes. In male embryos, testosterone is secreted by Leydig cells by the seventh week of development, with peak concentrations reached in the second trimester. This early release of testosterone results in the anatomical differentiation of the male sexual organs. In childhood, testosterone concentrations are low. They increase during puberty, activating characteristic physical changes and initiating spermatogenesis.
Functions of Testosterone
The continued presence of testosterone is necessary to keep the male reproductive system working properly, and Leydig cells produce approximately 6 to 7 mg of testosterone per day. Testicular steroidogenesis (the manufacture of androgens, including testosterone) results in testosterone concentrations that are 100 times higher in the testes than in the circulation. Maintaining these normal concentrations of testosterone promotes spermatogenesis, whereas low levels of testosterone can lead to infertility. In addition to intratesticular secretion, testosterone is also released into the systemic circulation and plays a key role in muscle development, bone growth, the development of secondary sex characteristics and maintaining libido (sex drive) in both males and females. In females, the ovaries secrete small amounts of testosterone, although most is converted to oestradiol. A small amount of testosterone is also secreted by the adrenal glands in both sexes.
Control of Testosterone
The regulation of testosterone concentrations throughout the body is critical for male reproductive function. The intricate interplay between the endocrine system and the reproductive system is shown in Figure 15.1.7.
Figure 15.1.7. Regulation of testosterone production. The hypothalamus and pituitary gland regulate the production of testosterone and the cells that assist in spermatogenesis. GnRH activates the anterior pituitary to produce LH and FSH, which in turn stimulate Leydig cells and Sertoli cells, respectively. The system is a negative feedback loop because the end products of the pathway, testosterone, and inhibin, interact with the activity of GnRH to inhibit their own production.
The regulation of Leydig cell production of testosterone begins outside of the testes. The hypothalamus and the pituitary gland in the brain integrate external and internal signals to control testosterone synthesis and secretion. The regulation begins in the hypothalamus. Pulsatile release of a hormone called gonadotropin-releasing hormone (GnRH) from the hypothalamus stimulates the endocrine release of hormones from the pituitary gland. Binding of GnRH to its receptors on the anterior pituitary gland stimulates release of the two gonadotropins: luteinising hormone (LH) and follicle-stimulating hormone (FSH). These two hormones are critical for reproductive function in both men and women. In men, FSH binds to the Sertoli cells within the seminiferous tubules to promote spermatogenesis. FSH also stimulates the Sertoli cells to produce hormones called inhibins, which function to inhibit FSH release from the pituitary, thus reducing testosterone secretion. These polypeptide hormones correlate directly with Sertoli cell function and sperm number; inhibin B can be used as a marker of spermatogenic activity. In men, LH binds to receptors on Leydig cells in the testes and upregulates the production of testosterone.
A negative feedback loop controls the synthesis and secretion of both FSH and LH. Low blood concentrations of testosterone stimulate the hypothalamic release of GnRH. GnRH then stimulates the anterior pituitary to secrete LH into the bloodstream. In the testis, LH binds to LH receptors on Leydig cells and stimulates the release of testosterone. When concentrations of testosterone in the blood reach a critical threshold, testosterone itself will bind to androgen receptors on both the hypothalamus and the anterior pituitary, inhibiting the synthesis and secretion of GnRH and LH, respectively. When the blood concentrations of testosterone once again decline, testosterone no longer interacts with the receptors to the same degree and GnRH and LH are once again secreted, stimulating more testosterone production. This same process occurs with FSH and inhibin to control spermatogenesis.
Ageing and the Male Reproductive System
Declines in Leydig cell activity can occur in men beginning at 40 to 50 years of age. The resulting reduction in circulating testosterone concentrations can lead to symptoms of andropause, also known as male menopause. While the reduction in sex steroids in men is akin to female menopause, there is no clear sign—such as a lack of a menstrual period—to denote the initiation of andropause. Instead, men report feelings of fatigue, reduced muscle mass, depression, anxiety, irritability, loss of libido and insomnia. A reduction in spermatogenesis resulting in lowered fertility is also reported, and sexual dysfunction can also be associated with andropausal symptoms.
Whereas some researchers believe that certain aspects of andropause are difficult to tease apart from ageing in general, testosterone replacement is sometimes prescribed to alleviate some symptoms. Recent studies have shown a benefit from androgen replacement therapy on the new onset of depression in elderly men; however, other studies caution against testosterone replacement for long-term treatment of andropause symptoms, showing that high doses can sharply increase the risk of both heart disease and prostate cancer.
Section Review
Gametes are the reproductive cells that combine to form offspring. Organs called gonads produce the gametes, along with the hormones that regulate human reproduction. The male gametes are called sperm. Spermatogenesis, the production of sperm, occurs within the seminiferous tubules that make up most of the testis. The scrotum is the muscular sac that holds the testes outside of the body cavity.
Spermatogenesis begins with mitotic division of spermatogonia (stem cells) to produce primary spermatocytes that undergo the two divisions of meiosis to become secondary spermatocytes, then the haploid spermatids. During spermiogenesis, spermatids are transformed into spermatozoa (formed sperm). Upon release from the seminiferous tubules, sperm are moved to the epididymis where they continue to mature. During ejaculation, sperm exit the epididymis through the ductus deferens, a duct in the spermatic cord that leaves the scrotum. The ampulla of the ductus deferens meets the seminal vesicle, a gland that contributes fructose and proteins, at the ejaculatory duct. The fluid continues through the prostatic urethra, where secretions from the prostate are added to form semen. These secretions help the sperm to travel through the urethra and into the female reproductive tract. Secretions from the bulbourethral glands protect sperm and cleanse and lubricate the penile (spongy) urethra.
The penis is the male organ of copulation. Columns of erectile tissue called the corpora cavernosa and corpus spongiosum fill with blood when sexual arousal activates vasodilatation in the blood vessels of the penis. Testosterone regulates and maintains the sex organs and sex drive and induces the physical changes of puberty. Interplay between the testes and the endocrine system precisely control the production of testosterone with a negative feedback loop.
15.2 Anatomy and Physiology of the Female Reproductive System
Learning Objectives
By the end of this section, you will be able to:
Describe the structure and function of the organs of the female reproductive system
List the steps of oogenesis
Describe the hormonal changes that occur during the ovarian and menstrual cycles
Trace the path of an oocyte from ovary to fertilisation
The female reproductive system functions to produce gametes and reproductive hormones, just like the male reproductive system; however, it also has the additional task of supporting the developing foetus and delivering it to the outside world. Unlike its male counterpart, the female reproductive system is located primarily inside the pelvic cavity (Figure 15.2.1). Recall that the ovaries are the female gonads. The gamete they produce is called an oocyte. We will discuss the production of oocytes in detail shortly. First, let us look at some of the structures of the female reproductive system.
Figure 15.2.1. Female reproductive system. The major organs of the female reproductive system are located inside the pelvic cavity.
External Female Genitals
The external female reproductive structures are referred to collectively as the vulva (Figure 15.2.2). The mons pubis is a pad of fat that is located at the anterior, over the pubic bone. After puberty, it becomes covered in pubic hair. The labia majora (labia = “lips”; majora = “larger”) are folds of hair-covered skin that begin just posterior to the mons pubis. The thinner and more pigmented labia minora (labia = “lips”; minora = “smaller”) extend medial to the labia majora. Although they naturally vary in shape and size from woman to woman, the labia minora serve to protect the female urethra and the entrance to the female reproductive tract.
The superior, anterior portions of the labia minora come together to encircle the clitoris (or glans clitoris), an organ that originates from the same cells as the glans penis and has abundant nerves that make it important in sexual sensation and orgasm. The hymen is a thin membrane that sometimes partially covers the entrance to the vagina. An intact hymen cannot be used as an indication of “virginity”; even at birth, this is only a partial membrane, as menstrual fluid and other secretions must be able to exit the body, regardless of penile–vaginal intercourse. The vaginal opening is located between the opening of the urethra and the anus. It is flanked by outlets to the Bartholin’s glands (or greater vestibular glands).
Figure 15.2.2. The Vulva. The external female genitalia are referred to collectively as the vulva.
Vagina
The vagina, shown at the bottom of Figure 15.2.1 and Figure 15.2.2, is a muscular canal (approximately 10 cm long) that serves as the entrance to the reproductive tract. It also serves as the exit from the uterus during menses and childbirth. The outer walls of the anterior and posterior vagina are formed into longitudinal columns, or ridges, and the superior portion of the vagina—called the fornix—meets the protruding uterine cervix. The walls of the vagina are lined with an outer, fibrous adventitia; a middle layer of smooth muscle; and an inner mucous membrane with transverse folds called rugae. Together, the middle and inner layers allow the expansion of the vagina to accommodate intercourse and childbirth. The thin, perforated hymen can partially surround the opening to the vaginal orifice. The hymen can be ruptured with strenuous physical exercise, penile–vaginal intercourse, and childbirth. The Bartholin’s glands and the lesser vestibular glands (located near the clitoris) secrete mucus, which keeps the vestibular area moist.
The vagina is home to a normal population of microorganisms that help to protect against infection by pathogenic bacteria, yeast, or other organisms that can enter the vagina. In a healthy woman, the most predominant type of vaginal bacteria is from the genus Lactobacillus. This family of beneficial bacterial flora secretes lactic acid, and thus protects the vagina by maintaining an acidic pH (below 4.5). Potential pathogens are less likely to survive in these acidic conditions. Lactic acid, in combination with other vaginal secretions, makes the vagina a self-cleansing organ. However, douching—or washing out the vagina with fluid—can disrupt the normal balance of healthy microorganisms and increase a woman’s risk for infections and irritation. Indeed, many doctors recommend that women do not douche and that they allow the vagina to maintain its normal healthy population of protective microbiota.
Ovaries
The ovaries are the female gonads (see Figure 15.2.1). Paired ovals, they are each about 2 to 3 cm in length, about the size of an almond. The ovaries are located within the pelvic cavity and are supported by the mesovarium, an extension of the peritoneum that connects the ovaries to the broad ligament. Extending from the mesovarium itself is the suspensory ligament that contains the ovarian blood and lymph vessels. Finally, the ovary itself is attached to the uterus via the ovarian ligament.
The ovary comprises an outer covering of cuboidal epithelium called the ovarian surface epithelium that is superficial to a dense connective tissue covering called the tunica albuginea. Beneath the tunica albuginea is the cortex, or outer portion, of the organ. The cortex is composed of a tissue framework called the ovarian stroma that forms the bulk of the adult ovary. Oocytes develop within the outer layer of this stroma, each surrounded by supporting cells. This grouping of an oocyte and its supporting cells is called a follicle. The growth and development of ovarian follicles will be described shortly. Beneath the cortex lies the inner ovarian medulla, the site of blood vessels, lymph vessels and the nerves of the ovary. You will learn more about the overall anatomy of the female reproductive system at the end of this section.
The Ovarian Cycle
The ovarian cycle is a set of predictable changes in a female’s oocytes and ovarian follicles. During a woman’s reproductive years, it is a roughly 28-day cycle that can be correlated with, but is different from, the menstrual cycle (discussed shortly). The cycle includes two interrelated processes: oogenesis (the production of female gametes) and folliculogenesis (the growth and development of ovarian follicles).
Oogenesis
Gametogenesis in females is called oogenesis. The process begins with the ovarian stem cells, or oogonia (Figure 15.2.3). Oogonia are formed during foetal development and divide via mitosis, much like spermatogonia in the testis. Unlike spermatogonia, however, oogonia form primary oocytes in the foetal ovary prior to birth. These primary oocytes are then arrested in this stage of meiosis I, only to resume it years later, beginning at puberty and continuing until the woman is near menopause (the cessation of a woman’s reproductive functions). The number of primary oocytes present in the ovaries declines from one to two million in an infant, to approximately 400,000 at puberty, to zero by the end of menopause.
The initiation of ovulation—the release of an oocyte from the ovary—marks the transition from puberty into reproductive maturity for women. From then on, throughout a woman’s reproductive years, ovulation occurs approximately once every 28 days. Just prior to ovulation, a surge of luteinising hormone triggers the resumption of meiosis in a primary oocyte. This initiates the transition from primary to secondary oocyte. However, as you can see in Figure 15.2.3, this cell division does not result in two identical cells. Instead, the cytoplasm is divided unequally, and one daughter cell is much larger than the other. This larger cell, the secondary oocyte, eventually leaves the ovary during ovulation. The smaller cell, called the first polar body, may or may not complete meiosis and produce second polar bodies; in either case, it eventually disintegrates. Therefore, even though oogenesis produces up to four cells, only one survives.
Figure 15.2.3. Oogenesis. The unequal cell division of oogenesis produces one to three polar bodies that later degrade, as well as a single haploid ovum, which is produced only if there is penetration of the secondary oocyte by a sperm cell.
How does the diploid secondary oocyte become an ovum—the haploid female gamete? Meiosis of a secondary oocyte is completed only if a sperm succeeds in penetrating its barriers. Meiosis II then resumes, producing one haploid ovum that, at the instant of fertilisation by a (haploid) sperm, becomes the first diploid cell of the new offspring (a zygote). Thus, the ovum can be thought of as a brief, transitional, haploid stage between the diploid oocyte and diploid zygote.
The larger amount of cytoplasm contained in the female gamete is used to supply the developing zygote with nutrients during the period between fertilisation and implantation into the uterus. Interestingly, sperm contribute only DNA at fertilisation —not cytoplasm. Therefore, the cytoplasm and all of the cytoplasmic organelles in the developing embryo are of maternal origin. This includes mitochondria, which contain their own DNA. Scientific research in the 1980s determined that mitochondrial DNA was maternally inherited, meaning that you can trace your mitochondrial DNA directly to your mother, her mother and so on, back through your female ancestors.
Everyday Connections
Mapping Human History with Mitochondrial DNA
When we talk about human DNA, we’re usually referring to nuclear DNA; that is, the DNA coiled into chromosomal bundles in the nucleus of our cells. We inherit half of our nuclear DNA from our father, and half from our mother. However, mitochondrial DNA (mtDNA) comes only from the mitochondria in the cytoplasm of the fat ovum we inherit from our mother. She received her mtDNA from her mother, who got it from her mother, and so on. Each of our cells contains approximately 1700 mitochondria, with each mitochondrion packed with mtDNA containing approximately 37 genes.
Mutations (changes) in mtDNA occur spontaneously in a somewhat organised pattern at regular intervals in human history. By analysing these mutational relationships, researchers have been able to determine that we can all trace our ancestry back to one woman who lived in Africa about 200,000 years ago. Scientists have given this woman the biblical name Eve, although she is not, of course, the first Homo sapien female. More precisely, she is our most recent common ancestor through matrilineal descent.
This does not mean that everyone’s mtDNA today looks exactly like that of our ancestral Eve. Because of the spontaneous mutations in mtDNA that have occurred over the centuries, researchers can map different “branches” off of the “main trunk” of our mtDNA family tree. Your mtDNA might have a pattern of mutations that aligns more closely with one branch, and your neighbour’s may align with another branch. Still, all branches eventually lead back to Eve.
But what happened to the mtDNA of all of the other Homo sapiens females who were living at the time of Eve? Researchers explain that, over the centuries, their female descendants died childless or with only male children and thus, their maternal line—and its mtDNA—ended.
Folliculogenesis
Again, ovarian follicles are oocytes and their supporting cells. They grow and develop in a process called folliculogenesis, which typically leads to ovulation of one follicle approximately every 28 days, along with death to multiple other follicles. The death of ovarian follicles is called atresia and can occur at any point during follicular development. Recall that, a female infant at birth will have one to two million oocytes within her ovarian follicles, and that this number declines throughout life until menopause, when no follicles remain. As you will see next, follicles progress from primordial, to primary, to secondary and tertiary stages prior to ovulation—with the oocyte inside the follicle remaining as a primary oocyte until right before ovulation.
Folliculogenesis begins with follicles in a resting state. These small primordial follicles are present in newborn females and are the prevailing follicle type in the adult ovary (Figure 15.2.4). Primordial follicles have only a single flat layer of support cells, called granulosa cells, that surround the oocyte and they can stay in this resting state for years—some until right before menopause.
After puberty, a few primordial follicles will respond to a recruitment signal each day and will join a pool of immature growing follicles called primary follicles. Primary follicles start with a single layer of granulosa cells, but the granulosa cells then become active and transition from a flat or squamous shape to a rounded, cuboidal shape as they increase in size and proliferate. As the granulosa cells divide, the follicles—now called secondary follicles (see Figure 15.2.4)—increase in diameter, adding a new outer layer of connective tissue, blood vessels and theca cells—cells that work with the granulosa cells to produce oestrogens.
Within the growing secondary follicle, the primary oocyte now secretes a thin acellular membrane called the zona pellucida that will play a critical role in fertilisation. A thick fluid, called follicular fluid, that has formed between the granulosa cells also begins to collect into one large pool, or antrum. Follicles in which the antrum has become large and fully formed are considered tertiary follicles (or antral follicles). Several follicles reach the tertiary stage at the same time and most of these will undergo atresia. The one that does not die will continue to grow and develop until ovulation, when it will expel its secondary oocyte surrounded by several layers of granulosa cells from the ovary. Keep in mind that most follicles do not make it to this point. In fact, roughly 99 percent of the follicles in the ovary will undergo atresia, which can occur at any stage of folliculogenesis.
The process of development that we have just described, from primordial follicle to early tertiary follicle, takes approximately two months in humans. The final stages of development of a small cohort of tertiary follicles, ending with ovulation of a secondary oocyte, occur over a course of approximately 28 days. These changes are regulated by many of the same hormones that regulate the male reproductive system, including GnRH, LH and FSH.
As in men, the hypothalamus produces GnRH, a hormone that signals the anterior pituitary gland to produce the gonadotropins FSH and LH (Figure 15.2.5). These gonadotropins leave the pituitary and travel through the bloodstream to the ovaries, where they bind to receptors on the granulosa and theca cells of the follicles. FSH stimulates the follicles to grow (hence its name of follicle-stimulating hormone), and the five or six tertiary follicles expand in diameter. The release of LH also stimulates the granulosa and theca cells of the follicles to produce the sex steroid hormone oestradiol, a type of oestrogen. This phase of the ovarian cycle, when the tertiary follicles are growing and secreting oestrogen, is known as the follicular phase.
The more granulosa and theca cells a follicle has (that is, the larger and more developed it is), the more oestrogen it will produce in response to LH stimulation. As a result of these large follicles producing substantial amounts of oestrogen, systemic plasma oestrogen concentrations increase. Following a classic negative feedback loop, the high concentrations of oestrogen will stimulate the hypothalamus and pituitary to reduce the production of GnRH, LH, and FSH. Because the large tertiary follicles require FSH to grow and survive at this point, this decline in FSH caused by negative feedback leads most of them to die (atresia). Typically, only one follicle, now called the dominant follicle, will survive this reduction in FSH, and this follicle will be the one that releases an oocyte. Scientists have studied many factors that lead to a particular follicle becoming dominant: size, the number of granulosa cells, and the number of FSH receptors on those granulosa cells all contribute to a follicle becoming the one surviving dominant follicle.
Figure 15.2.5. Hormonal regulation of ovulation. The hypothalamus and pituitary gland regulate the ovarian cycle and ovulation. GnRH activates the anterior pituitary to produce LH and FSH, which stimulate the production of oestrogen and progesterone by the ovaries.
When only the one dominant follicle remains in the ovary, it again begins to secrete oestrogen. It produces more oestrogen than all the developing follicles did together before the negative feedback occurred. It produces so much oestrogen that the normal negative feedback does not occur. Instead, these extremely high concentrations of systemic plasma oestrogen trigger a regulatory switch in the anterior pituitary that responds by secreting substantial amounts of LH and FSH into the bloodstream (see Figure 15.2.5). The positive feedback loop by which more oestrogen triggers release of more LH and FSH only occurs at this point in the cycle.
It is this large burst of LH (called the LH surge) that leads to ovulation of the dominant follicle. The LH surge induces many changes in the dominant follicle, including stimulating the resumption of meiosis of the primary oocyte to a secondary oocyte. As noted earlier, the polar body that results from unequal cell division simply degrades. The LH surge also triggers proteases (enzymes that cleave proteins) to break down structural proteins in the ovary wall on the surface of the bulging dominant follicle. This degradation of the wall, combined with pressure from the large, fluid-filled antrum, results in the expulsion of the oocyte surrounded by granulosa cells into the peritoneal cavity. This release is ovulation.
In the next section, you will follow the ovulated oocyte as it travels toward the uterus, but there is one more important event that occurs in the ovarian cycle. The surge of LH also stimulates a change in the granulosa and theca cells that remain in the follicle after the oocyte has been ovulated. This change is called luteinisation (recall that the full name of LH is luteinising hormone), and it transforms the collapsed follicle into a new endocrine structure called the corpus luteum, a term meaning “yellowish body” (see Figure 15.2.4). Instead of oestrogen, the luteinised granulosa and theca cells of the corpus luteum begin to produce substantial amounts of the sex steroid hormone progesterone, a hormone that is critical for the establishment and maintenance of pregnancy. Progesterone triggers negative feedback at the hypothalamus and pituitary, which keeps GnRH, LH and FSH secretions low, so no new dominant follicles develop at this time.
The post-ovulatory phase of progesterone secretion is known as the luteal phase of the ovarian cycle. If pregnancy does not occur within 10 to 12 days, the corpus luteum will stop secreting progesterone and degrade into the corpus albicans, a non-functional “whitish body” that will disintegrate in the ovary over a period of several months. During this time of reduced progesterone secretion, FSH and LH are once again stimulated and the follicular phase begins again with a new cohort of early tertiary follicles beginning to grow and secrete oestrogen.
The Uterine Tubes
The uterine tubes (also called fallopian tubes or oviducts) serve as the conduit of the oocyte from the ovary to the uterus (Figure 15.2.6). Each of the two uterine tubes is close to, but not directly connected to, the ovary and is divided into sections. The isthmus is the narrow medial end of each uterine tube that is connected to the uterus. The wide distal infundibulum flares out with slender, finger-like projections called fimbriae. The middle region of the tube, called the ampulla, is where fertilisation often occurs. The uterine tubes also have three layers: an outer serosa, a middle smooth muscle layer and an inner mucosal layer. In addition to its mucus-secreting cells, the inner mucosa contains ciliated cells that beat in the direction of the uterus, producing a current that will be critical to move the oocyte.
Following ovulation, the secondary oocyte surrounded by a few granulosa cells is released into the peritoneal cavity. The nearby uterine tube, either left or right, receives the oocyte. Unlike sperm, oocytes lack flagella, and therefore cannot move on their own. So how do they travel into the uterine tube and toward the uterus? High concentrations of oestrogen that occur around the time of ovulation induce contractions of the smooth muscle along the length of the uterine tube. These contractions occur every 4 to 8 seconds, and the result is a coordinated movement that sweeps the surface of the ovary and the pelvic cavity. Current flowing toward the uterus is generated by coordinated beating of the cilia that line the outside and lumen of the length of the uterine tube. These cilia beat more strongly in response to the high oestrogen concentrations that occur around the time of ovulation. As a result of these mechanisms, the oocyte–granulosa cell complex is pulled into the interior of the tube. Once inside, the muscular contractions and beating cilia move the oocyte slowly toward the uterus. When fertilisation does occur, sperm typically meet the egg while it is still moving through the ampulla.
If the oocyte is successfully fertilised, the resulting zygote will begin to divide into two cells, then four, and so on, as it makes its way through the uterine tube and into the uterus. There, it will implant and continue to grow. If the egg is not fertilised, it will simply degrade—either in the uterine tube or in the uterus, where it may be shed with the next menstrual period.
The open-ended structure of the uterine tubes can have significant health consequences if bacteria or other infectious agents enter through the vagina and move through the uterus, into the tubes, and then into the pelvic cavity. If this is left unchecked, an infection (typically bacterial) could quickly become life-threatening due to sepsis. The spread of an infection in this manner is of special concern when unskilled practitioners perform abortions in non-sterile conditions. Sepsis is also associated with sexually transmitted bacterial infections, especially gonorrhoea and chlamydia. These increase a woman’s risk for pelvic inflammatory disease (PID), infection of the uterine tubes or other reproductive organs. Even when resolved, PID can leave scar tissue in the tubes, leading to infertility.
The Uterus and Cervix
The uterus is the muscular organ that nourishes and supports the growing embryo (see Figure 15.2.6). Its average size is approximately 5 cm wide by 7 cm long (approximately 2 in by 3 in) when a female is not pregnant. It has three sections. The portion of the uterus superior to the opening of the uterine tubes is called the fundus. The middle section of the uterus is called the body (or corpus) of uterus . The cervix is the narrow inferior portion of the uterus that projects into the vagina. The cervix produces mucus secretions that become thin and stringy under the influence of high systemic plasma oestrogen concentrations and these secretions can facilitate sperm movement through the reproductive tract.
Several ligaments maintain the position of the uterus within the abdominopelvic cavity. The broad ligament is a fold of peritoneum that serves as a primary support for the uterus, extending laterally from both sides of the uterus and attaching it to the pelvic wall. The round ligament attaches to the uterus near the uterine tubes and extends to the labia majora. Finally, the uterosacral ligament stabilises the uterus posteriorly by its connection from the cervix to the pelvic wall.
The wall of the uterus is made up of three layers. The most superficial layer is the serous membrane, or perimetrium, which consists of epithelial tissue that covers the exterior portion of the uterus. The middle layer, or myometrium, is a thick layer of smooth muscle responsible for uterine contractions. Most of the uterus is myometrial tissue and the muscle fibres run horizontally, vertically and diagonally, allowing the powerful contractions that occur during labour and the less powerful contractions (or cramps) that help to expel menstrual blood during a woman’s period. Anteriorly directed myometrial contractions also occur near the time of ovulation and are thought to facilitate the transport of sperm through the female reproductive tract.
The innermost layer of the uterus is called the endometrium. The endometrium contains a connective tissue lining, the lamina propria, which is covered by epithelial tissue that lines the lumen. Structurally, the endometrium consists of two layers: the stratum basalis and the stratum functionalis (the basal and functional layers). The stratum basalis layer is part of the lamina propria and is adjacent to the myometrium; this layer does not shed during menses. In contrast, the thicker stratum functionalis layer contains the glandular portion of the lamina propria and the endothelial tissue that lines the uterine lumen. It is the stratum functionalis that grows and thickens in response to increased levels of oestrogen and progesterone. In the luteal phase of the menstrual cycle, special branches off the uterine artery called spiral arteries supply the thickened stratum functionalis. This inner functional layer provides the proper site of implantation for the fertilised egg, and—should fertilisation not occur—it is only the stratum functionalis layer of the endometrium that sheds during menstruation.
Recall that during the follicular phase of the ovarian cycle, the tertiary follicles are growing and secreting oestrogen. At the same time, the stratum functionalis of the endometrium is thickening to prepare for a potential implantation. The post-ovulatory increase in progesterone, which characterises the luteal phase, is key for maintaining a thick stratum functionalis. If a functional corpus luteum is present in the ovary, the endometrial lining is prepared for implantation. Indeed, if an embryo implants, signals are sent to the corpus luteum to continue secreting progesterone to maintain the endometrium, and thus maintain the pregnancy. If an embryo does not implant, no signal is sent to the corpus luteum and it degrades, ceasing progesterone production and ending the luteal phase. Without progesterone, the endometrium thins and, under the influence of prostaglandins, the spiral arteries of the endometrium constrict and rupture, preventing oxygenated blood from reaching the endometrial tissue. As a result, endometrial tissue dies and blood, pieces of the endometrial tissue, and white blood cells are shed through the vagina during menstruation, or the menses. The first menses after puberty, called menarche, can occur either before or after the first ovulation.
The Menstrual Cycle
Now that we have discussed the maturation of the cohort of tertiary follicles in the ovary, the build-up and then shedding of the endometrial lining in the uterus and the function of the uterine tubes and vagina, we can put everything together to talk about the three phases of the menstrual cycle—the series of changes in which the uterine lining is shed, rebuilds and prepares for implantation.
The timing of the menstrual cycle starts with the first day of menses, referred to as day one of a woman’s period. Cycle length is determined by counting the days between the onset of bleeding in two subsequent cycles. Because the average length of a woman’s menstrual cycle is 28 days, this is the time period used to identify the timing of events in the cycle. However, the length of the menstrual cycle varies among women and even in the same woman from one cycle to the next, typically from 21 to 32 days.
Just as the hormones produced by the granulosa and theca cells of the ovary “drive” the follicular and luteal phases of the ovarian cycle, they also control the three distinct phases of the menstrual cycle. These are the menses phase, the proliferative phase and the secretory phase.
Menses Phase
The menses phase of the menstrual cycle is the phase during which the lining is shed; that is, the days that the woman menstruates. Although it averages approximately five days, the menses phase can last from 2 to 7 days, or longer. As shown in Figure 15.2.7, the menses phase occurs during the early days of the follicular phase of the ovarian cycle, when progesterone, FSH and LH levels are low. Recall that progesterone concentrations decline because of the degradation of the corpus luteum, marking the end of the luteal phase. This decline in progesterone triggers the shedding of the stratum functionalis of the endometrium.
Figure 15.2.7. Hormone levels in ovarian and menstrual cycles. The correlation of the hormone levels and their effects on the female reproductive system is shown in this timeline of the ovarian and menstrual cycles. The menstrual cycle begins at day one with the start of menses. Ovulation occurs around day 14 of a 28-day cycle, triggered by the LH surge.
Proliferative Phase
Once menstrual flow ceases, the endometrium begins to proliferate again, marking the beginning of the proliferative phase of the menstrual cycle (see Figure 15.2.7). It occurs when the granulosa and theca cells of the tertiary follicles begin to produce increased amounts of oestrogen. These rising oestrogen concentrations stimulate the endometrial lining to rebuild.
Recall that the high oestrogen concentrations will eventually lead to a decrease in FSH because of negative feedback, resulting in atresia of all but one of the developing tertiary follicles. The switch to positive feedback—which occurs with the elevated oestrogen production from the dominant follicle—then stimulates the LH surge that will trigger ovulation. In a typical 28-day menstrual cycle, ovulation occurs on day 14. Ovulation marks the end of the proliferative phase as well as the end of the follicular phase.
Secretory Phase
In addition to prompting the LH surge, high oestrogen levels increase the uterine tube contractions that facilitate the pick-up and transfer of the ovulated oocyte. High oestrogen levels also slightly decrease the acidity of the vagina, making it more hospitable to sperm. In the ovary, the luteinisation of the granulosa cells of the collapsed follicle forms the progesterone-producing corpus luteum, marking the beginning of the luteal phase of the ovarian cycle. In the uterus, progesterone from the corpus luteum begins the secretory phase of the menstrual cycle, in which the endometrial lining prepares for implantation (see Figure 15.2.7). Over the next 10 to 12 days, the endometrial glands secrete a fluid rich in glycogen. If fertilisation has occurred, this fluid will nourish the ball of cells now developing from the zygote. At the same time, the spiral arteries develop to provide blood to the thickened stratum functionalis.
If no pregnancy occurs within approximately 10 to 12 days, the corpus luteum will degrade into the corpus albicans. Levels of both oestrogen and progesterone will fall, and the endometrium will grow thinner. Prostaglandins will be secreted that cause constriction of the spiral arteries, reducing oxygen supply. The endometrial tissue will die, resulting in menses—or the first day of the next cycle.
Disorders of the Female Reproductive System
Research over many years has confirmed that cervical cancer is most often caused by a sexually transmitted infection with human papillomavirus (HPV). There are over 100 related viruses in the HPV family, and the characteristics of each strain determine the outcome of the infection. In all cases, the virus enters body cells and uses its own genetic material to take over the host cell’s metabolic machinery and produce more virus particles.
HPV infections are common in both men and women. Indeed, a recent study determined that 42.5 percent of females had HPV at the time of testing. These women ranged in age from 14 to 59 years and differed in race, ethnicity and number of sexual partners. Of note, the prevalence of HPV infection was 53.8 percent among women aged 20 to 24 years, the age group with the highest infection rate.
HPV strains are classified as high or low risk according to their potential to cause cancer. Though most HPV infections do not cause disease, the disruption of normal cellular functions in the low-risk forms of HPV can cause the male or female human host to develop genital warts. Often, the body can clear an HPV infection by normal immune responses within 2 years. However, the more serious, high-risk infection by certain types of HPV can result in cancer of the cervix (Figure 15.2.8). Infection with either of the cancer-causing variants HPV 16 or HPV 18 has been linked to more than 70 percent of all cervical cancer diagnoses. Although even these high-risk HPV strains can be cleared from the body over time, infections persist in some individuals. If this happens, the HPV infection can influence the cells of the cervix to develop precancerous changes.
Risk factors for cervical cancer include having unprotected sex; having multiple sexual partners; a first sexual experience at a younger age, when the cells of the cervix are not fully mature; failure to receive the HPV vaccine; a compromised immune system; and smoking. The risk of developing cervical cancer is doubled with cigarette smoking.
Figure 15.2.8. Development of cervical cancer. In most cases, cells infected with the HPV virus heal on their own. In some cases, however, the virus continues to spread and becomes an invasive cancer.
When the high-risk types of HPV enter a cell, two viral proteins are used to neutralise proteins that the host cells use as checkpoints in the cell cycle. The best studied of these proteins is p53. In a normal cell, p53 detects DNA damage in the cell’s genome and either halts the progression of the cell cycle—allowing time for DNA repair to occur—or initiates apoptosis. Both processes prevent the accumulation of mutations in a cell’s genome. High-risk HPV can neutralise p53, keeping the cell in a state in which fast growth is possible and impairing apoptosis, allowing mutations to accumulate in the cellular DNA.
The prevalence of cervical cancer in Australia has dropped dramatically from approximately 1200 cases in 1994, down to approximately 700 cases in 2002. However, cervical cancer cases are steadily rising in Australia with 950 new cases in 2019.
Until 2017, the method of screening for cervical cancer in Australia was through a Pap test. Pap smears sample cells of the cervix, allowing the detection of abnormal cells. If pre-cancerous cells are detected, there are several highly effective techniques that are currently in use to remove them before they pose a danger. However, women in developing countries often do not have access to regular pap smears. As a result, these women account for as many as 80 percent of the cases of cervical cancer worldwide. In 2017, the Cervical Screening Test replaced the Pap test in Australia, which is estimated to protect an additional 30% of women. The cervical screening test is designed to find evidence of the human papillomavirus (HPV), rather than looking for abnormal cells as was done in the pap smear test.
In 2006, the first vaccine against the high-risk types of HPV was approved. There are now two vaccines available in Australia. Gardasil9 is designed to protect against nine of the HPV types that are responsible for 90% of cervical cancers in women and 95% of HPV-related cancers in men, and also against 90% of genital warts in both sexes. This vaccine is administered to females aged from nine to 45, and males aged 9 to 27. Cervarix® is given to females from as early as ten years of age, up to 45 years of age and is designed to prevent early-stage cervical cancers known as pre-cancerous lesions. A recent study suggests that the HPV vaccine has cut the rates of HPV infection by the four targeted strains at least in half. Unfortunately, the high cost of manufacturing the vaccine is currently limiting access to many people worldwide.
The Breasts
Whereas the breasts are located far from the other female reproductive organs, they are considered accessory organs of the female reproductive system. The function of the breasts is to supply milk to an infant in a process called lactation. The external features of the breast include a nipple surrounded by a pigmented areola (Figure 15.2.9), whose colouration may deepen during pregnancy. The areola is typically circular and can vary in size from 25 to 100 mm in diameter. The areolar region is characterised by small, raised areolar glands that secrete lubricating fluid during lactation to protect the nipple from chafing. When a baby nurses, or draws milk from the breast, the entire areolar region is taken into the mouth.
Breast milk is produced by the mammary glands, which are modified sweat glands. The milk itself exits the breast through the nipple via 15 to 20 lactiferous ducts that open on the surface of the nipple. These lactiferous ducts each extend to a lactiferous sinus that connects to a glandular lobe within the breast itself that contains groups of milk-secreting cells in clusters called alveoli (see Figure 15.2.9). The clusters can change in size depending on the amount of milk in the alveolar lumen. Once milk is made in the alveoli, stimulated myoepithelial cells that surround the alveoli contract to push the milk to the lactiferous sinuses. From here, the baby can draw milk through the lactiferous ducts by suckling. The lobes themselves are surrounded by fat tissue, which determines the size of the breast; breast size differs between individuals and does not affect the amount of milk produced. Supporting the breasts are multiple bands of connective tissue called suspensory ligaments that connect the breast tissue to the dermis of the overlying skin.
Figure 15.2.9. Anatomy of the breast. During lactation, milk moves from the alveoli through the lactiferous ducts to the nipple.
During the normal hormonal fluctuations in the menstrual cycle, breast tissue responds to changing levels of oestrogen and progesterone, which can lead to swelling and breast tenderness in some individuals, especially during the secretory phase. If pregnancy occurs, the increase in hormones leads to further development of the mammary tissue and enlargement of the breasts.
Hormonal Birth Control
Birth control pills take advantage of the negative feedback system that regulates the ovarian and menstrual cycles to stop ovulation and prevent pregnancy. Typically, they work by providing a constant level of both oestrogen and progesterone, which negatively feeds back onto the hypothalamus and pituitary, thus preventing the release of FSH and LH. Without FSH, the follicles do not mature, and without the LH surge, ovulation does not occur. Although the oestrogen in birth control pills does stimulate some thickening of the endometrial wall, it is reduced compared with a normal cycle and is less likely to support implantation.
Some birth control pills contain 21 active pills containing hormones, and 7 inactive pills (placebos). The decline in hormones during the week that the woman takes the placebo pills triggers menses, although it is typically lighter than a normal menstrual flow because of the reduced endometrial thickening. Newer types of birth control pills have been developed that deliver low-dose oestrogens and progesterone for the entire cycle (these are meant to be taken 365 days a year), and menses never occurs. While some women prefer to have the proof of a lack of pregnancy that a monthly period provides, menstruation every 28 days is not required for health reasons and there are no reported adverse effects of not having a menstrual period in an otherwise healthy individual.
Because birth control pills function by providing constant oestrogen and progesterone levels and disrupting negative feedback, skipping even just one or two pills at certain points of the cycle (or even being several hours late taking the pill) can lead to an increase in FSH and LH and result in ovulation. It is important, therefore, that the woman follow the directions on the birth control pill package to successfully prevent pregnancy.
Ageing and the Female Reproductive System
Female fertility (the ability to conceive) peaks when women are in their twenties and is slowly reduced until a woman reaches 35 years of age. After that time, fertility declines more rapidly, until it ends completely at the end of menopause. Menopause is the cessation of the menstrual cycle that occurs because of the loss of ovarian follicles and the hormones that they produce. A woman is considered to have completed menopause if she has not menstruated in a full year. After that point, she is considered postmenopausal. The average age for this change is consistent worldwide at between 50 and 52 years of age, but it can normally occur in a woman’s forties, or later in her fifties. Poor health, including smoking, can lead to earlier loss of fertility and earlier menopause.
As a woman reaches the age of menopause, depletion of the number of viable follicles in the ovaries due to atresia affects the hormonal regulation of the menstrual cycle. During the years leading up to menopause, there is a decrease in the levels of the hormone inhibin, which normally participates in a negative feedback loop to the pituitary to control the production of FSH. The menopausal decrease in inhibin leads to an increase in FSH. The presence of FSH stimulates more follicles to grow and secrete oestrogen. Because small, secondary follicles also respond to increases in FSH levels, larger numbers of follicles are stimulated to grow; however, most undergo atresia and die. Eventually, this process leads to the depletion of all follicles in the ovaries, and the production of oestrogen falls off dramatically. It is primarily the lack of oestrogens that leads to the symptoms of menopause.
The earliest changes occur during the menopausal transition, often referred to as peri-menopause, when a women’s cycle becomes irregular but does not stop entirely. Although the levels of oestrogen are still the same as before the transition, the level of progesterone produced by the corpus luteum is reduced. This decline in progesterone can lead to abnormal growth, or hyperplasia, of the endometrium. This condition is a concern because it increases the risk of developing endometrial cancer. Two harmless conditions that can develop during the transition are uterine fibroids, which are benign masses of cells, and irregular bleeding. As oestrogen levels change, other symptoms that occur are hot flashes and night sweats, trouble sleeping, vaginal dryness, mood swings, difficulty focussing, and thinning of hair on the head along with the growth of more hair on the face. Depending on the individual, these symptoms can be entirely absent, moderate, or severe.
After menopause, lower amounts of oestrogens can lead to other changes. Cardiovascular disease becomes as prevalent in women as in men, possibly because oestrogens reduce the amount of cholesterol in the blood vessels. When oestrogen is lacking, many women find that they suddenly have problems with high cholesterol and the cardiovascular issues that accompany it. Osteoporosis is another problem because bone density decreases rapidly in the first years after menopause. The reduction in bone density leads to a higher incidence of fractures.
Menopausal Hormone Therapy (MHT)/Hormone replacement therapy (HRT), which employs medication (synthetic oestrogens and progestins) to increase oestrogen and progestin levels, can alleviate some of the symptoms of menopause. In 2002, the Women’s Health Initiative began a study to observe women for the long-term outcomes of hormone replacement therapy over 8.5 years. However, the study was prematurely terminated after 5.2 years because of evidence of a higher than normal risk of breast cancer in patients taking oestrogen-only HRT. The potential positive effects on cardiovascular disease were also not realised in the oestrogen-only patients. The results of other hormone replacement studies over the last 50 years, including a 2019 study of preclinical and clinical evidence, which found that use of oestrogen support the cardiovascular protective benefits of timely MHT. Additionally, the study showed that although MHT is correlated with a reduced risk of breast cancer development, its correlation with ovarian cancer is still unknown. Some researchers believe that the age group tested in the 2002 trial may have been too old to benefit from the therapy, thus skewing the results. In the meantime, intense debate and study of the benefits and risks of replacement therapy is ongoing. Current guidelines approve MHT for the reduction of hot flashes or flushes, but The Royal Australian and New Zealand College of Obstetricians and Gynaecologists (RANZCOG) recommends that each patient requires an individual assessment of risks and benefits prior to commencement of MHT.
Section Review
The external female genitalia are collectively called the vulva. The vagina is the pathway into and out of the uterus. The man’s penis is inserted into the vagina to deliver sperm and the baby exits the uterus through the vagina during childbirth.
The ovaries produce oocytes, the female gametes, in a process called oogenesis. As with spermatogenesis, meiosis produces the haploid gamete (in this case, an ovum); however, it is completed only in an oocyte that has been penetrated by a sperm. In the ovary, an oocyte surrounded by supporting cells is called a follicle. In folliculogenesis, primordial follicles develop into primary, secondary and tertiary follicles. Early tertiary follicles with their fluid-filled antrum will be stimulated by an increase in FSH, a gonadotropin produced by the anterior pituitary, to grow in the 28-day ovarian cycle. Supporting granulosa and theca cells in the growing follicles produce oestrogens, until the concentration of oestrogen in the bloodstream is high enough that it triggers negative feedback at the hypothalamus and pituitary. This results in a reduction of FSH and LH, and most tertiary follicles in the ovary undergo atresia (they die). One follicle, usually the one with the most FSH receptors, survives this period and is now called the dominant follicle. The dominant follicle produces more oestrogen, triggering positive feedback and the LH surge that will induce ovulation. Following ovulation, the granulosa cells of the empty follicle luteinise and transform into the progesterone-producing corpus luteum. The ovulated oocyte with its surrounding granulosa cells is picked up by the infundibulum of the uterine tube and beating cilia help to transport it through the tube toward the uterus. Fertilisation occurs within the uterine tube and the final stage of meiosis is completed.
The uterus has three regions: the fundus, the body and the cervix. It has three layers: the outer perimetrium, the muscular myometrium and the inner endometrium. The endometrium responds to oestrogen released by the follicles during the menstrual cycle and grows thicker with an increase in blood vessels in preparation for pregnancy. If the egg is not fertilised, no signal is sent to extend the life of the corpus luteum, and it degrades, stopping progesterone production. This decline in progesterone results in the sloughing of the inner portion of the endometrium in a process called menses, or menstruation.
The breasts are accessory sexual organs that are utilised after the birth of a child to produce milk in a process called lactation. Birth control pills provide constant levels of oestrogen and progesterone to negatively feedback on the hypothalamus and pituitary, and suppress the release of FSH and LH, which inhibits ovulation and prevents pregnancy.
15.3 Development of the Male and Female Reproductive Systems
Learning Objectives
By the end of this section, you will be able to:
Explain how bipotential tissues are directed to develop into male or female sex organs
Name the rudimentary duct systems in the embryo that are precursors to male or female internal sex organs
Describe the hormonal changes that bring about puberty and the secondary sex characteristics of men and women
The development of the reproductive systems begins soon after fertilisation of the egg, with primordial gonads beginning to develop approximately one month after conception. Reproductive development continues in utero, but there is little change in the reproductive system between infancy and puberty.
Development of the Sexual Organs in the Embryo and Foetus
Females are considered the “fundamental” sex—that is, without much chemical prompting, all fertilised eggs would develop into females. To become a male, an individual must be exposed to the cascade of factors initiated by a single gene on the male Y chromosome. This is called the SRY (Sex-determining Region of the Y chromosome). Because females do not have a Y chromosome, they do not have the SRY gene. Without a functional SRY gene, an individual will be female.
In both male and female embryos, the same group of cells has the potential to develop into either the male or female gonads; this tissue is considered bipotential. The SRY gene actively recruits other genes that begin to develop the testes and suppresses genes that are important in female development. As part of this SRY-prompted cascade, germ cells in the bipotential gonads differentiate into spermatogonia. Without SRY, different genes are expressed, oogonia form, and primordial follicles develop in the primitive ovary.
Soon after the formation of the testis, the Leydig cells begin to secrete testosterone. Testosterone can influence tissues that are bipotential to become male reproductive structures. With exposure to testosterone, cells that could become either the glans penis or the glans clitoris form the glans penis. Without testosterone, these same cells differentiate into the clitoris.
Not all tissues in the reproductive tract are bipotential. The internal reproductive structures (for example the uterus, uterine tubes, and part of the vagina in females; and the epididymis, ductus deferens, and seminal vesicles in males) form from one of two rudimentary duct systems in the embryo. For proper reproductive function in the adult, one set of these ducts must develop properly, and the other must degrade. In males, secretions from sustentacular cells trigger a degradation of the female duct, called the Müllerian duct. At the same time, testosterone secretion stimulates growth of the male tract, the Wolffian duct. Without such sustentacular cell secretion, the Müllerian duct will develop; without testosterone, the Wolffian duct will degrade. Thus, the developing offspring will be female. For more information and a figure of differentiation of the gonads, seek additional content on foetal development.
Further Sexual Development Occurs at Puberty
Puberty is the stage of development at which individuals become sexually mature. Though the outcomes of puberty for boys and girls are very different, the hormonal control of the process is very similar. In addition, though the timing of these events varies between individuals, the sequence of changes that occur is predictable for male and female adolescents. As shown in Figure 15.3.1, a concerted release of hormones from the hypothalamus (GnRH), the anterior pituitary (LH and FSH) and the gonads (either testosterone or oestrogen) is responsible for the maturation of the reproductive systems and the development of secondary sex characteristics, which are physical changes that serve auxiliary roles in reproduction.
The first changes begin around the age of eight or nine when the production of LH becomes detectable. The release of LH occurs primarily at night during sleep and precedes the physical changes of puberty by several years. In pre-pubertal children, the sensitivity of the negative feedback system in the hypothalamus and pituitary is remarkably high. This means that very low concentrations of androgens or oestrogens will negatively feed back onto the hypothalamus and pituitary, keeping the production of GnRH, LH, and FSH low.
As an individual approaches puberty, two changes in sensitivity occur. The first is a decrease of sensitivity in the hypothalamus and pituitary to negative feedback, meaning that it takes increasingly larger concentrations of sex steroid hormones to stop the production of LH and FSH. The second change in sensitivity is an increase in sensitivity of the gonads to the FSH and LH signals, meaning the gonads of adults are more responsive to gonadotropins than are the gonads of children. As a result of these two changes, the levels of LH and FSH slowly increase and lead to the enlargement and maturation of the gonads, which in turn leads to secretion of higher levels of sex hormones and the initiation of spermatogenesis and folliculogenesis.
In addition to age, multiple factors can affect the age of onset of puberty, including genetics, environment, and psychological stress. One of the more important influences may be nutrition; historical data demonstrate the effect of better and more consistent nutrition on the age of menarche in girls in the United States, which decreased from an average age of approximately 17 years of age in 1860 to the current age of approximately 12.75 years in 1960, as it remains today. Some studies indicate a link between puberty onset and the amount of stored fat in an individual. This effect is more pronounced in girls but has been documented in both sexes. Body fat, corresponding with secretion of the hormone leptin by adipose cells, appears to have a strong role in determining menarche. This may reflect the high metabolic costs of gestation and lactation. In girls who are lean and highly active, such as gymnasts, there is often a delay in the onset of puberty.
Figure 15.3.1. Hormones of puberty. During puberty, the release of LH and FSH from the anterior pituitary stimulates the gonads to produce sex hormones in both male and female adolescents.
Signs of Puberty
Different sex steroid hormone concentrations between the sexes also contribute to the development and function of secondary sexual characteristics and examples of secondary sexual characteristics are listed in Table 15.3.1.
Table 15.3.1. Development of secondary sexual characteristics
Male
Female
Increased larynx size and deepening of the voice
Deposition of fat, predominantly in breast and hips
Increased muscular development
Breast development
Growth of facial, axillary and pubic hair and increased growth of body hair
Broadening of the pelvis and growth of axillary and pubic hair
As a girl reaches puberty, typically the first change that is visible is the development of the breast tissue. This is followed by the growth of axillary and pubic hair. A growth spurt normally starts at approximately age 9 to 11 and may last two years or more. During this time, a girl’s height can increase approximately 7.6 cm (~3 inches) a year. The next step in puberty is menarche, the start of menstruation.
In boys, the growth of the testes is typically the first physical sign of the beginning of puberty, which is followed by growth and pigmentation of the scrotum and growth of the penis. The next step is the growth of hair, including armpit, pubic, chest and facial hair. Testosterone stimulates the growth of the larynx and thickening and lengthening of the vocal folds, which causes the voice to drop in pitch. The first fertile ejaculations typically appear at approximately 15 years of age, but this age can vary widely across individual boys. Unlike the early growth spurt observed in females, the male growth spurt occurs toward the end of puberty, at approximately age 11 to 13 and a boy’s height can increase as much as 10 cm (4 inches) a year. In some males, pubertal development can continue through the early 20s.
Section Review
The reproductive systems of males and females begin to develop soon after conception. A gene on the male’s Y chromosome called SRY is critical in stimulating a cascade of events that simultaneously stimulate testis development and repress the development of female structures. Testosterone produced by Leydig cells in the embryonic testis stimulates the development of male sexual organs. If testosterone is not present, female sexual organs will develop.
Whereas the gonads and some other reproductive tissues are considered bipotential, the tissue that forms the internal reproductive structures stems from ducts that will develop into only male (Wolffian) or female (Müllerian) structures. To be able to reproduce as an adult, one of these systems must develop properly and the other must degrade.
Further development of the reproductive systems occurs at puberty. The initiation of the changes that occur in puberty is the result of a decrease in sensitivity to negative feedback in the hypothalamus and pituitary gland, and an increase in sensitivity of the gonads to FSH and LH stimulation. These changes lead to increases in either oestrogen or testosterone, in female and male adolescents, respectively. The increase in sex steroid hormones leads to maturation of the gonads and other reproductive organs. The initiation of spermatogenesis begins in boys and girls begin ovulating and menstruating. Increases in sex steroid hormones also lead to the development of secondary sex characteristics such as breast development in girls and facial hair and larynx growth in boys.
Describe the obstacles that sperm must overcome to reach an oocyte to enable fertilisation
Explain capacitation and its importance in fertilisation
Summarise the events that occur as a sperm fertilises an oocyte in order to form a zygote
Explain the artificial reproductive technologies (ART) that are used for assisted reproduction in humans
Fertilisation occurs when a sperm and an oocyte (egg) combine and their nuclei fuse. Because each of these reproductive cells is a haploid cell containing half of the genetic material needed to form a human being, their combination forms a diploid cell. This new single cell, called a zygote, contains all the genetic material needed to form a human – half from the mother and half from the father.
Transit of Sperm
Fertilisation is a numbers game. During ejaculation, hundreds of millions of sperm (spermatozoa) are released into the vagina. Almost immediately, millions of these sperm are overcome by the acidity of the vagina (approximately pH 3.8) and millions more may be blocked from entering the uterus by thick cervical mucus. Of those that do enter, thousands are destroyed by phagocytic uterine leukocytes. Thus, the race into the uterine tubes, which is the most typical site for sperm to encounter the oocyte, is reduced to a few thousand contenders. Their journey—thought to be facilitated by uterine contractions—usually takes from 30 minutes to two hours. If the sperm do not encounter an oocyte immediately, they can survive in the uterine tubes for another 3–5 days. Thus, fertilisation can still occur if intercourse takes place a few days before ovulation. In comparison, an oocyte can survive independently for only approximately 24 hours following ovulation. Intercourse more than a day after ovulation will therefore usually not result in fertilisation.
During the journey, fluids in the female reproductive tract prepare the sperm for fertilisation through a process called capacitation or priming. The fluids improve the motility of the spermatozoa. They also deplete cholesterol molecules embedded in the membrane of the head of the sperm, thinning the membrane in such a way that will help facilitate the release of the lysosomal (digestive) enzymes needed for the sperm to penetrate the oocyte’s exterior once contact is made. Sperm must undergo the process of capacitation in order to have the “capacity” to fertilise an oocyte. If they reach the oocyte before capacitation is complete, they will be unable to penetrate the oocyte’s thick outer layer of cells.
Contact Between Sperm and Oocyte
Upon ovulation, the oocyte released by the ovary is swept into—and along—the uterine tube. Fertilisation must occur in the distal uterine tube because an unfertilised oocyte cannot survive the 72-hour journey to the uterus. As you will recall from your study of the oogenesis, this oocyte (specifically a secondary oocyte) is surrounded by two protective layers. The corona radiata is an outer layer of follicular (granulosa) cells that form around a developing oocyte in the ovary and remain with it upon ovulation. The underlying zona pellucida (pellucid = “transparent”) is a transparent, but thick, glycoprotein membrane that surrounds the cell’s plasma membrane.
As it is swept along the distal uterine tube, the oocyte encounters the surviving capacitated sperm, which stream toward it in response to chemical attractants released by the cells of the corona radiata. To reach the oocyte itself, the sperm must penetrate the two protective layers. The sperm first burrow through the cells of the corona radiata. Then, upon contact with the zona pellucida, the sperm bind to receptors in the zona pellucida. This initiates a process called the acrosomal reaction in which the enzyme-filled “cap” of the sperm, called the acrosome, releases its stored digestive enzymes. These enzymes clear a path through the zona pellucida that allows sperm to reach the oocyte. Finally, molecules on the sperm head of a single sperm contact the sperm-binding receptors on the oocyte’s plasma membrane (Figure 16.1.1). The plasma membrane of that sperm then fuses with the oocyte’s plasma membrane, and the head and mid-piece (not the tail) of the “winning” sperm enter the cytoplasm (interior) of the oocyte.
How do sperm penetrate the corona radiata? Some sperm undergo a spontaneous acrosomal reaction, which is an acrosomal reaction not triggered by contact with the zona pellucida. The digestive enzymes released by this reaction digest the extracellular matrix of the corona radiata. As you can see, the first sperm to reach the oocyte is never the one to fertilise it. Rather, hundreds of sperm cells must undergo the acrosomal reaction, each helping to degrade the corona radiata and zona pellucida until a path is created to allow one sperm to contact and fuse with the plasma membrane of the oocyte. If you consider the loss of millions of sperm between entry into the vagina and degradation of the zona pellucida, you can understand why a low sperm count can cause male infertility.
Figure 16.1.1. Sperm and the process of fertilisation. Before fertilisation, hundreds of capacitated sperm must break through the surrounding corona radiata and zona pellucida so that one can contact and fuse with the oocyte plasma membrane.
When the first sperm fuses with the oocyte, the oocyte deploys two mechanisms to prevent polyspermy, which is penetration by more than one sperm. This is critical because if more than one sperm were to fertilise the oocyte, the resulting zygote would be a triploid organism with three sets of chromosomes. This is incompatible with life.
The first mechanism is the fast block, which involves a near instantaneous change in sodium ion permeability upon binding of the first sperm, depolarising the oocyte plasma membrane and preventing the fusion of additional sperm cells. The fast block sets in almost immediately and lasts for about a minute, during which time an influx of calcium ions following sperm penetration triggers the second mechanism, the slow block. In this process, referred to as the cortical reaction, cortical granules sitting immediately below the oocyte plasma membrane fuse with the membrane and release zonal inhibiting proteins and mucopolysaccharides into the space between the plasma membrane and the zona pellucida. Zonal inhibiting proteins cause the release of any other attached sperm and destroy the oocyte’s sperm receptors, thus preventing any more sperm from binding. The mucopolysaccharides then coat the nascent zygote in an impenetrable barrier that, together with hardened zona pellucida, is called a fertilisation membrane.
The Zygote
Recall that at the point of fertilisation, the oocyte has not yet completed meiosis; all secondary oocytes remain arrested in metaphase of meiosis II until fertilisation. Only upon fertilisation does the oocyte complete meiosis. The unneeded complement of genetic material that results is stored in a second polar body that is eventually ejected. At this moment, the oocyte has become an ovum, the female haploid gamete. The two haploid nuclei derived from the sperm and oocyte and contained within the egg are referred to as pronuclei. They decondense, expand, and replicate their DNA in preparation for mitosis. The pronuclei then migrate toward each other, their nuclear envelopes disintegrate, and the male- and female-derived genetic material intermingles. This step completes the process of fertilisation and results in a single-celled diploid zygote with all the genetic instructions it needs to develop into a human.
Most of the time, a woman releases a single egg during an ovulation cycle. However, in approximately 1 percent of ovulation cycles, two eggs are released, and both are fertilised. Two zygotes form, implant, and develop, resulting in the birth of dizygotic (or fraternal) twins. Because dizygotic twins develop from two eggs fertilised by two sperm, they are no more identical than siblings born at different times.
Much less commonly, a zygote can divide into two separate offspring during early development. This results in the birth of monozygotic (or identical) twins. Although the zygote can split as early as the two-cell stage, splitting occurs most commonly during the early blastocyst stage, with roughly 70–100 cells present. These two scenarios are distinct from each other, in that the twin embryos that separated at the two-cell stage will have individual placentas, whereas twin embryos that form from separation at the blastocyst stage will share a placenta and a chorionic cavity.
Everyday Connections
In Vitro Fertilisation (IVF)
IVF, which stands for in vitro fertilisation, is an assisted reproductive technology. In vitro, which in Latin translates to “in glass,” refers to a procedure that takes place outside of the body. There are many different indications for IVF, for example, a woman may produce normal eggs, but the eggs cannot reach the uterus because the uterine tubes are blocked or otherwise compromised. A man may have a low sperm count, low sperm motility, sperm with an unusually high percentage of morphological abnormalities, or sperm that are incapable of penetrating the zona pellucida of an egg.
A typical IVF procedure begins with egg collection. A normal ovulation cycle produces only one oocyte, but the number can be boosted significantly (to 10–20 oocytes) by administering a short course of gonadotropins. The course begins with follicle-stimulating hormone (FSH) analogs, which support the development of multiple follicles, and ends with a luteinising hormone (LH) analog that triggers ovulation. Right before the ova would be released from the ovary, they are harvested using ultrasound-guided oocyte retrieval. In this procedure, ultrasound allows a physician to visualise mature follicles. The ova are aspirated (sucked out) using a syringe.
In parallel, sperm are obtained from the male partner or from a sperm bank. The sperm are prepared by washing to remove seminal fluid because seminal fluid contains a peptide, FPP (or, fertilisation promoting peptide), that—in high concentrations—prevents capacitation of the sperm. The sperm sample is also concentrated, to increase the sperm count per millilitre.
Next, the eggs and sperm are mixed in a petri dish. The ideal ratio is 75,000 sperm to one egg. If there are severe problems with the sperm—for example, the count is exceedingly low, or the sperm are completely nonmotile, or incapable of binding to or penetrating the zona pellucida—a sperm can be injected into an egg. This is called intracytoplasmic sperm injection (ICSI).
The embryos are then incubated until they either reach the eight-cell stage or the blastocyst stage. In most countries, for example Australia, UK, US and Canada, fertilised eggs are typically cultured to the blastocyst stage because this results in a higher pregnancy rate. Finally, the embryos are transferred to a woman’s uterus using a plastic catheter (tube). Figure 16.1.2 illustrates the steps involved in IVF.
Figure 16.1.2. IVF.In vitro fertilisation involves egg collection from the ovaries, fertilisation in a petri dish, and the transfer of embryos into the uterus.
The first successful IVF procedure was conducted in 1973. Seven years later, in 1980, the first IVF birth in Australia occurred. In 1983 the world saw the first birth from a frozen embryo prior to transfer. Despite the advances and milestones reached over the last 50 years, IVF technology is a continuously evolving technique.
The rate of success for IVF is correlated with a woman’s age. In Australia, the success rate for live births from fresh embryo transfers is approximately 40% in patients 30 years and younger, however drops to only approximately 9% in women over the age of 40. Similarly, the rate of live births from frozen embryo transfer is approximately 35% in women 30 years and younger and approximately 27% in women 40 years and older.
Career Connection
IVF Scientist/Embryologist
The IVF or Fertility Clinics industry in Australia and world-wide is growing as more couples decide to delay pregnancy. Medical facilities in this industry offer services such as assisted reproductive technologies (ART) including IVF, frozen embryo transfer, fertility treatments, intrauterine insemination and donor insemination as well as diagnostic and genetic testing. An IVF Scientist or Embryologist requires a minimum of a bachelor’s degree in biomedical science or biological science focussing on human biology.
Section Review
Hundreds of millions of sperm deposited in the vagina travel toward the oocyte, but only a few hundred reach it. The number of sperm that reach the oocyte is greatly reduced because of conditions within the female reproductive tract. Many sperm are overcome by the acidity of the vagina, others are blocked by mucus in the cervix, whereas others are attacked by phagocytic leukocytes in the uterus. Those sperm that do survive undergo a change in response to those conditions. They go through the process of capacitation, which improves their motility and alters the membrane surrounding the acrosome, the cap-like structure in the head of a sperm that contains the digestive enzymes needed for it to attach to and penetrate the oocyte.
The oocyte that is released by ovulation is protected by a thick outer layer of granulosa cells known as the corona radiata and by the zona pellucida, a thick glycoprotein membrane that lies just outside the oocyte’s plasma membrane. When capacitated sperm make contact with the oocyte, they release the digestive enzymes in the acrosome (the acrosomal reaction) and are thus able to attach to the oocyte and burrow through to the oocyte’s zona pellucida. One of the sperm will then break through to the oocyte’s plasma membrane and release its haploid nucleus into the oocyte. The oocyte’s membrane structure changes in response (cortical reaction), preventing any further penetration by another sperm and forming a fertilisation membrane. Fertilisation is complete upon unification of the haploid nuclei of the two gametes, producing a diploid zygote.
Distinguish the stages of embryonic development that occur before implantation
Describe the process of implantation
List and describe four embryonic membranes
Explain gastrulation
Describe how the placenta is formed and identify its functions
Explain how an embryo transforms from a flat disc of cells into a three-dimensional shape resembling a human
Summarise the process of organogenesis
Throughout this chapter, we will express embryonic and foetal ages in terms of weeks from fertilisation, commonly called conception. The period of time required for full development of a foetusin utero is referred to as gestation (gestare = “to carry” or “to bear”). It can be subdivided into distinct gestational periods. The first 2 weeks of prenatal development are referred to as the pre-embryonic stage. A developing human is referred to as an embryo during weeks 3–8 and a foetus from the ninth week of gestation until birth. In this section, we’ll cover the pre-embryonic and embryonic stages of development, which are characterised by cell division, migration, and differentiation. By the end of the embryonic period, all the organ systems are structured in rudimentary form, although the organs themselves are either non-functional or only semi-functional.
Pre-Implantation Embryonic Development
Following fertilisation, the zygote and its associated membranes, together referred to as the conceptus, continue to be projected toward the uterus by peristalsis and beating cilia. During its journey to the uterus, the zygote undergoes five or six rapid mitotic cell divisions. Although each cleavage results in more cells, it does not increase the total volume of the conceptus (Figure 16.2.1). Each daughter cell produced by cleavage is called a blastomere (blastos = “germ,” in the sense of a seed or sprout).
Approximately three days after fertilisation, a 16-cell conceptus reaches the uterus. The cells that had been loosely grouped are now compacted and look more like a solid mass. The name given to this structure is the morula (morula = “little mulberry”). Once inside the uterus, the conceptus floats freely for several more days. It continues to divide, creating a ball of approximately 100 cells, and consuming nutritive endometrial secretions called uterine milk while the uterine lining thickens. The ball of now tightly bound cells starts to secrete fluid and organise themselves around a fluid-filled cavity, the blastocoel. At this developmental stage, the conceptus is referred to as a blastocyst. Within this structure, a group of cells forms into an inner cell mass, which is fated to become the embryo. The cells that form the outer shell are called trophoblasts (trophe = “to feed” or “to nourish”). These cells will develop into the chorionic sac and the foetal portion of the placenta (the organ of nutrient, waste and gas exchange between mother and the developing offspring).
The inner mass of embryonic cells is totipotent during this stage, meaning that each cell has the potential to differentiate into any cell type in the human body. Totipotency lasts for only a few days before the cells’ fates are set as being the precursors to a specific lineage of cells.
Figure 16.2.1. Pre-embryonic cleavages. Pre-embryonic cleavages make use of the abundant cytoplasm of the conceptus as the cells rapidly divide without changing the total volume.
As the blastocyst forms, the trophoblast excretes enzymes that begin to degrade the zona pellucida. In a process called “hatching,” the conceptus breaks free of the zona pellucida in preparation for implantation.
Implantation
At the end of the first week, the blastocyst comes in contact with the uterine wall and adheres to it, embedding itself in the uterine lining via the trophoblast cells. Thus, begins the process of implantation, which signals the end of the pre-embryonic stage of development (Figure 16.2.2). Implantation can be accompanied by minor bleeding. The blastocyst typically implants in the fundus of the uterus or on the posterior wall. However, if the endometrium is not fully developed and ready to receive the blastocyst, the blastocyst will detach and find a better spot. A significant percentage (50–75 percent) of blastocysts fail to implant; when this occurs, the blastocyst is shed with the endometrium during menses. The high rate of implantation failure is one reason why pregnancy typically requires several ovulation cycles to achieve.
Figure 16.2.2. Pre-embryonic development. Ovulation, fertilisation, pre-embryonic development, and implantation occur at specific locations within the female reproductive system in a time span of approximately one week.
When implantation succeeds and the blastocyst adheres to the endometrium, the superficial cells of the trophoblast fuse with each other, forming the syncytiotrophoblast, a multinucleated body that digests endometrial cells to firmly secure the blastocyst to the uterine wall. In response, the uterine mucosa rebuilds itself and envelops the blastocyst (Figure 16.2.3). The trophoblast secretes human chorionic gonadotropin (hCG), a hormone that directs the corpus luteum to survive, enlarge, and continue producing progesterone and oestrogen to suppress menses. These functions of hCG are necessary for creating an environment suitable for the developing embryo. As a result of this increased production, hCG accumulates in the maternal bloodstream and is excreted in the urine. Implantation is complete by the middle of the second week. Just a few days after implantation, the trophoblast has secreted enough hCG for an at-home urine pregnancy test to give a positive result.
Figure 16.2.3. Implantation. During implantation, the trophoblast cells of the blastocyst adhere to the endometrium and digest endometrial cells until it is attached securely.
Most of the time an embryo implants within the body of the uterus in a location that can support growth and development. However, in one to two percent of cases, the embryo implants either outside the uterus (an ectopic pregnancy) or in a region of uterus that can create complications for the pregnancy. If the embryo implants in the inferior portion of the uterus, the placenta can potentially grow over the opening of the cervix, a condition call placenta previa.
Disorders of the Development of the Embryo
In the vast majority of ectopic pregnancies, the embryo does not complete its journey to the uterus and implants in the uterine tube, referred to as a tubal pregnancy. However, there are also ovarian ectopic pregnancies (in which the egg never left the ovary) and abdominal ectopic pregnancies (in which an egg was “lost” to the abdominal cavity during the transfer from ovary to uterine tube, or in which an embryo from a tubal pregnancy re-implanted in the abdomen). Once in the abdominal cavity, an embryo can implant into any well-vascularised structure—the rectouterine cavity (Douglas’ pouch), the mesentery of the intestines, and the greater omentum are some common sites.
Tubal pregnancies can be caused by scar tissue within the tube following a sexually transmitted bacterial infection. The scar tissue impedes the progress of the embryo into the uterus—in some cases “snagging” the embryo and, in other cases, blocking the tube completely. Approximately one half of tubal pregnancies resolve spontaneously. Implantation in a uterine tube causes bleeding, which appears to stimulate smooth muscle contractions and expulsion of the embryo. In the remaining cases, medical or surgical intervention is necessary. If an ectopic pregnancy is detected early, the embryo’s development can be arrested by the administration of the cytotoxic drug methotrexate, which inhibits the metabolism of folic acid. If diagnosis is late and the uterine tube is already ruptured, surgical repair is essential.
Even if the embryo has successfully found its way to the uterus, it does not always implant in an optimal location (the fundus or the posterior wall of the uterus). Placenta previa can result if an embryo implants close to the internal os of the uterus (the internal opening of the cervix). As the foetus grows, the placenta can partially or completely cover the opening of the cervix (Figure 16.2.4). Although it occurs in only 0.5 percent of pregnancies, placenta previa is the leading cause of antepartum haemorrhage (profuse vaginal bleeding after week 24 of pregnancy but prior to childbirth).
Figure 16.2.4. Placenta previa. An embryo that implants too close to the opening of the cervix can lead to placenta previa, a condition in which the placenta partially or completely covers the cervix.
Embryonic Membranes
During the second week of development, with the embryo implanted in the uterus, cells within the blastocyst start to organise into layers. Some grow to form the extra-embryonic membranes needed to support and protect the growing embryo: the amnion, the yolk sac, the allantois, and the chorion.
At the beginning of the second week, the cells of the inner cell mass form into a two-layered disc of embryonic cells, and a space—the amniotic cavity—opens between it and the trophoblast (Figure 16.2.5). Cells from the upper layer of the disc (the epiblast) extend around the amniotic cavity, creating a membranous sac that forms into the amnion by the end of the second week. The amnion fills with amniotic fluid and eventually grows to surround the embryo. Early in development, amniotic fluid consists almost entirely of a filtrate of maternal plasma, but as the kidneys of the foetus begin to function at approximately the eighth week, they add urine to the volume of amniotic fluid. Floating within the amniotic fluid, the embryo—and later, the foetus—is protected from trauma and rapid temperature changes. It can move freely within the fluid and can prepare for swallowing and breathing out of the uterus.
Figure 16.2.5. Development of the embryonic disc. Formation of the embryonic disc leaves spaces on either side that develop into the amniotic cavity and the yolk sac.
On the ventral side of the embryonic disc, opposite the amnion, cells in the lower layer of the embryonic disk (the hypoblast) extend into the blastocyst cavity and form a yolk sac. The yolk sac supplies some nutrients absorbed from the trophoblast and provides primitive blood circulation to the developing embryo for the second and third week of development. When the placenta takes over nourishing the embryo at approximately week 4, the yolk sac has been greatly reduced in size and its main function is to serve as the source of blood cells and germ cells (cells that will give rise to gametes). During week 3, a finger-like out-pocketing of the yolk sac develops into the allantois, a primitive excretory duct of the embryo that will become part of the urinary bladder. Together, the stalks of the yolk sac and allantois establish the outer structure of the umbilical cord.
The last of the extra-embryonic membranes is the chorion, which is the one membrane that surrounds all others. The development of the chorion will be discussed in more detail shortly, as it relates to the growth and development of the placenta.
Embryogenesis
As the third week of development begins, the two-layered disc of cells becomes a three-layered disc through the process of gastrulation, during which the cells transition from totipotency to multipotency. The embryo, which takes the shape of an oval-shaped disc, forms an indentation called the primitive streak along the dorsal surface of the epiblast. A node at the caudal or “tail” end of the primitive streak emits growth factors that direct cells to multiply and migrate. Cells migrate toward and through the primitive streak and then move laterally to create two new layers of cells. The first layer is the endoderm, a sheet of cells that displaces the hypoblast and lies adjacent to the yolk sac. The second layer of cells fills in as the middle layer, or mesoderm. The cells of the epiblast that remain (not having migrated through the primitive streak) become the ectoderm (Figure 16.2.6).
Figure 16.2.6. Germ layers. Formation of the three primary germ layers occurs during the first 2 weeks of development. The embryo at this stage is only a few millimetres in length.
Each of these germ layers will develop into specific structures in the embryo. Whereas the ectoderm and endoderm form tightly connected epithelial sheets, the mesodermal cells are less organised and exist as a loosely connected cell community. The ectoderm gives rise to cell lineages that differentiate to become the central and peripheral nervous systems, sensory organs, epidermis, hair and nails. Mesodermal cells ultimately become the skeleton, muscles, connective tissue, heart, blood vessels and kidneys. The endoderm goes on to form the epithelial lining of the gastrointestinal tract, liver and pancreas, as well as the lungs (Figure 16.2.7).
Figure 16.2.7. Fates of germ layers in embryo. Following gastrulation of the embryo in the third week, embryonic cells of the ectoderm, mesoderm, and endoderm begin to migrate and differentiate into the cell lineages that will give rise to mature organs and organ systems in the infant.
Development of the Placenta
During the first several weeks of development, the cells of the endometrium—referred to as decidual cells—nourish the nascent embryo. During prenatal weeks 4–12, the developing placenta gradually takes over the role of feeding the embryo and the decidual cells are no longer needed. The mature placenta is composed of tissues derived from the embryo, as well as maternal tissues of the endometrium. The placenta connects to the conceptus via the umbilical cord, which carries deoxygenated blood and wastes from the foetus through two umbilical arteries; nutrients and oxygen are carried from the mother to the foetus through the single umbilical vein. The umbilical cord is surrounded by the amnion and the spaces within the cord around the blood vessels are filled with Wharton’s jelly, a mucous connective tissue.
The maternal portion of the placenta develops from the deepest layer of the endometrium, the decidua basalis. To form the embryonic portion of the placenta, the syncytiotrophoblast and the underlying cells of the trophoblast (cytotrophoblast cells) begin to proliferate along with a layer of extraembryonic mesoderm cells. These form the chorionic membrane, which envelops the entire conceptus as the chorion. The chorionic membrane forms finger-like structures called chorionic villi that burrow into the endometrium like tree roots, making up the foetal portion of the placenta. The cytotrophoblast cells perforate the chorionic villi, burrow farther into the endometrium, and remodel maternal blood vessels to augment maternal blood flow surrounding the villi. Meanwhile, foetal mesenchymal cells derived from the mesoderm fill the villi and differentiate into blood vessels, including the three umbilical blood vessels that connect the embryo to the developing placenta (Figure 16.2.8).
Figure 16.2.8. Cross-section of the placenta. In the placenta, maternal and foetal blood components are conducted through the surface of the chorionic villi, but maternal and foetal bloodstreams never mix directly.
The placenta develops throughout the embryonic period and during the first several weeks of the foetal period; placentation is complete by weeks 14–16. As a fully developed organ, the placenta provides nutrition and excretion, respiration and endocrine function (Table 16.2.1 and Figure 16.2.9). It receives blood from the foetus through the umbilical arteries. Capillaries in the chorionic villi filter foetal wastes out of the blood and return clean, oxygenated blood to the foetus through the umbilical vein. Nutrients and oxygen are transferred from maternal blood surrounding the villi through the capillaries and into the foetal bloodstream. Some substances move across the placenta by simple diffusion. Oxygen, carbon dioxide, and any other lipid-soluble substances take this route. Other substances move across by facilitated diffusion. This includes water-soluble glucose. The foetus has a high demand for amino acids and iron, and those substances are moved across the placenta by active transport.
Figure 16.2.9. Placenta. This post-expulsion placenta and umbilical cord (white) are viewed from the foetal side.
Maternal and foetal blood does not commingle because blood cells cannot move across the placenta. This separation prevents the mother’s cytotoxic T cells from reaching and subsequently destroying the foetus, which bears “non-self” antigens. Further, it ensures the foetal red blood cells do not enter the mother’s circulation and trigger antibody development (if they carry “non-self” antigens)—at least until the final stages of pregnancy or birth. This is the reason that, even in the absence of preventive treatment, an Rh− mother doesn’t develop antibodies that could cause haemolytic disease in her first Rh+ foetus.
Although blood cells are not exchanged, the chorionic villi provide ample surface area for the two-way exchange of substances between maternal and foetal blood. The rate of exchange increases throughout gestation as the villi become thinner and increasingly branched. The placenta is permeable to lipid-soluble fetotoxic substances: alcohol, nicotine, barbiturates, antibiotics, certain pathogens, and many other substances that can be dangerous or fatal to the developing embryo or foetus. For these reasons, pregnant women should avoid foetotoxic substances. Alcohol consumption by pregnant women, for example, can result in a range of abnormalities referred to as foetal alcohol spectrum disorders (FASD). These include organ and facial malformations, as well as cognitive and behavioural disorders.
Table 16.2.1. Functions of the placenta
Nutrition and digestion
Respiration
Endocrine function
Mediates diffusion of maternal glucose, amino acids, fatty acids, vitamins and minerals
Mediates maternal-to-foetal oxygen transport and foetal-to-maternal carbon dioxide transport
Secretes several hormones, including hCG, oestrogens and progesterone to maintain the pregnancy and stimulate maternal and foetal development
Stores nutrients during early to accommodate increased foetal demand later in pregnancy
Mediates the transmission of maternal hormones into foetal blood and vice versa
Excretes and filters foetal nitrogenous wastes into maternal blood
Organogenesis
Following gastrulation, rudiments of the central nervous system develop from the ectoderm in the process of neurulation (Figure 16.2.10). Specialised neuroectodermal tissues along the length of the embryo thicken into the neural plate. During the fourth week, tissues on either side of the plate fold upward into a neural fold. The two folds converge to form the neural tube. The tube lies atop a rod-shaped, mesoderm-derived notochord, which eventually becomes the nucleus pulposus of intervertebral discs. Block-like structures called somites form on either side of the tube, eventually differentiating into the axial skeleton, skeletal muscle and dermis. During the fourth and fifth weeks, the anterior neural tube dilates and subdivides to form vesicles that will become the brain structures.
Folate, one of the B vitamins, is important to the healthy development of the neural tube. A deficiency of maternal folate in the first weeks of pregnancy can result in neural tube defects, including spina bifida—a birth defect in which spinal tissue protrudes through the newborn’s vertebral column which has failed to completely close. A more severe neural tube defect is anencephaly, a partial or complete absence of brain tissue.
Figure 16.2.10. Neurulation. The embryonic process of neurulation establishes the rudiments of the future central nervous system and skeleton.
The embryo, which begins as a flat sheet of cells, begins to acquire a cylindrical shape through the process of embryonic folding (Figure 16.2.11). The embryo folds laterally and again at either end, forming a C-shape with distinct head and tail ends. The embryo envelops a portion of the yolk sac, which protrudes with the umbilical cord from what will become the abdomen. The folding essentially creates a tube, called the primitive gut, that is lined by the endoderm. The amniotic sac, which was sitting on top of the flat embryo, envelops the embryo as it folds.
Figure 16.2.11. Embryonic folding. Embryonic folding converts a flat sheet of cells into a hollow, tube-like structure.
Within the first eight weeks of gestation, a developing embryo establishes the rudimentary structures of all its organs and tissues from the ectoderm, mesoderm and endoderm. This process is called organogenesis.
Figure 16.2.12. Embryo at seven weeks. An embryo at the end of seven weeks of development is only 10 mm in length, but its developing eyes, limb buds, and tail are already visible. (This embryo was derived from an ectopic pregnancy.) (credit: Ed Uthman).
Like the central nervous system, the heart also begins its development in the embryo as a tube-like structure, connected via capillaries to the chorionic villi. Cells of the primitive tube-shaped heart are capable of electrical conduction and contraction. The heart begins beating in the beginning of the fourth week, although it does not actually pump embryonic blood until a week later, when the oversized liver has begun producing red blood cells. (This is a temporary responsibility of the embryonic liver that the bone marrow will assume during foetal development.) During weeks four–five, the eye pits form, limb buds become apparent and the rudiments of the pulmonary system are formed.
During the sixth week, uncontrolled foetal limb movements begin to occur. The gastrointestinal system develops too rapidly for the embryonic abdomen to accommodate it and the intestines temporarily loop into the umbilical cord. Paddle-shaped hands and feet develop fingers and toes by the process of apoptosis (programmed cell death), which causes the tissues between the fingers to disintegrate. By week seven, the facial structure is more complex and includes nostrils, outer ears and lenses (Figure 16.2.12). By the eighth week, the head is nearly as large as the rest of the embryo’s body and all major brain structures are in place. The external genitalia are apparent, but at this point, male and female embryos are indistinguishable. Bone begins to replace cartilage in the embryonic skeleton through the process of ossification. By the end of the embryonic period, the embryo is approximately three cm from crown to rump and weighs approximately eight grams.
Section Review
As the zygote travels toward the uterus, it undergoes numerous cleavages in which the number of cells doubles (blastomeres). Upon reaching the uterus, the conceptus has become a tightly packed sphere of cells called the morula, which then forms into a blastocyst consisting of an inner cell mass within a fluid-filled cavity surrounded by trophoblasts. The blastocyst implants in the uterine wall, the trophoblasts fuse to form a syncytiotrophoblast, and the conceptus is enveloped by the endometrium. Four embryonic membranes form to support the growing embryo: the amnion, the yolk sac, the allantois and the chorion. The chorionic villi of the chorion extend into the endometrium to form the foetal portion of the placenta. The placenta supplies the growing embryo with oxygen and nutrients; it also removes carbon dioxide and other metabolic wastes.
Following implantation, embryonic cells undergo gastrulation, in which they differentiate and separate into an embryonic disc and establish three primary germ layers (the endoderm, mesoderm and ectoderm). Through the process of embryonic folding, the foetus begins to take shape. Neurulation starts the process of the development of structures of the central nervous system and organogenesis establishes the basic plan for all organ systems.
Differentiate between the embryonic period and the foetal period
Briefly describe the process of sexual differentiation
Describe the foetal circulatory system and explain the role of the shunts
Trace the development of a foetus from the end of the embryonic period to birth
As you will recall, a developing human is called a foetus from the ninth week of gestation until birth. This 30-week period of development is marked by continued cell growth and differentiation, which fully develop the structures and functions of the immature organ systems formed during the embryonic period. The completion of foetal development results in a newborn who, although still immature in many ways, is capable of survival outside the womb.
Sexual Differentiation
Sexual differentiation does not begin until the foetal period, during weeks 9–12. Embryonic males (with SRY gene) and females, though genetically distinguishable, are morphologically identical (Figure 16.3.1). Bipotential gonads, or gonads that can develop into male or female sexual organs, are connected to a central cavity called the cloaca via Müllerian ducts and Wolffian ducts. (The cloaca is an extension of the primitive gut.) Several events lead to sexual differentiation during this period.
During male foetal development, the bipotential gonads become the testes and associated epididymis. The Müllerian ducts degenerate. The Wolffian ducts become the vas deferens and the cloaca becomes the urethra and rectum.
During female foetal development, the bipotential gonads develop into ovaries. The Wolffian ducts degenerate. The Müllerian ducts become the uterine tubes and uterus and the cloaca divides and develops into a vagina, a urethra and a rectum.
Figure 16.3.1. Sexual differentiation. Differentiation of the male and female reproductive systems does not occur until the foetal period of development.
The Foetal Circulatory System
During prenatal development, the foetal circulatory system is integrated with the placenta via the umbilical cord so that the foetus receives both oxygen and nutrients from the placenta. However, after childbirth, the umbilical cord is severed, and the newborn’s circulatory system must be reconfigured. When the heart first forms in the embryo, it exists as two parallel tubes derived from mesoderm and lined with endothelium, which then fuse together. As the embryo develops into a foetus, the tube-shaped heart folds and further differentiates into the four chambers present in a mature heart. Unlike a mature cardiovascular system, however, the foetal cardiovascular system also includes circulatory shortcuts, or shunts. A shunt is an anatomical (or sometimes surgical) diversion that allows blood flow to bypass immature organs such as the lungs and liver until childbirth.
The placenta provides the foetus with necessary oxygen and nutrients via the umbilical vein. (Remember that veins carry blood toward the heart. In this case however, the blood flowing to the foetal heart is oxygenated because it comes from the placenta. The respiratory system is immature and cannot yet oxygenate blood on its own.) From the umbilical vein, the oxygenated blood flows toward the inferior vena cava, all but bypassing the immature liver, via the ductus venosus shunt (Figure 16.3.2). The liver receives just a trickle of blood, which is all that it needs in its immature, semifunctional state. Blood flows from the inferior vena cava to the right atrium, mixing with foetal venous blood along the way.
Although the foetal liver is semifunctional, the foetal lungs are non-functional. The foetal circulation therefore bypasses the lungs by shifting some of the blood through the foramen ovale, a shunt that directly connects the right and left atria and avoids the pulmonary trunk altogether. Most of the rest of the blood is pumped to the right ventricle, and from there, into the pulmonary trunk, which splits into pulmonary arteries. However, a shunt within the pulmonary artery, the ductus arteriosus, diverts a portion of this blood into the aorta. This ensures that only a small volume of oxygenated blood passes through the immature pulmonary circuit, which has only minor metabolic requirements. Blood vessels of uninflated lungs have high resistance to flow, a condition that encourages blood to flow to the aorta, which presents much lower resistance. The oxygenated blood moves through the foramen ovale into the left atrium, where it mixes with the now deoxygenated blood returning from the pulmonary circuit. This blood then moves into the left ventricle, where it is pumped into the aorta. Some of this blood moves through the coronary arteries into the myocardium and some moves through the carotid arteries to the brain.
The descending aorta carries partially oxygenated and partially deoxygenated blood into the lower regions of the body. It eventually passes into the umbilical arteries through branches of the internal iliac arteries. The deoxygenated blood collects waste as it circulates through the foetal body and returns to the umbilical cord. Thus, the two umbilical arteries carry blood low in oxygen and high in carbon dioxide and foetal wastes. This blood is filtered through the placenta, where wastes diffuse into the maternal circulation. Oxygen and nutrients from the mother diffuse into the placenta and from there into the foetal blood, and the process repeats.
Figure 16.3.2. Foetal circulatory system. The foetal circulatory system includes three shunts to divert blood from undeveloped and partially functioning organs, as well as blood supply to and from the placenta.
Other Organ Systems
During weeks 9–12 of foetal development, the brain continues to expand, the body elongates, and ossification continues. Foetal movements are frequent during this period but are jerky and not well-controlled. The bone marrow begins to take over the process of erythrocyte production—a task that the liver performed during the embryonic period. The liver now secretes bile. The foetus circulates amniotic fluid by swallowing it and producing urine. The eyes are well-developed by this stage, but the eyelids are fused shut. The fingers and toes begin to develop nails. By the end of week 12, the foetus measures approximately 9 cm from crown to rump.
Weeks 13–16 are marked by sensory organ development. The eyes move closer together; blinking motions begin, although the eyes remain sealed shut. The lips exhibit sucking motions. The ears move upward and lie flatter against the head. The scalp begins to grow hair. The excretory system is also developing: the kidneys are well-formed, and meconium, or foetal faeces, begins to accumulate in the intestines. Meconium consists of ingested amniotic fluid, cellular debris, mucus and bile.
During approximately weeks 16–20, as the foetus grows and limb movements become more powerful, the mother may begin to feel quickening, or foetal movements. However, space restrictions limit these movements and typically force the growing foetus into the “foetal position,” with the arms crossed and the legs bent at the knees. Sebaceous glands coat the skin with a waxy, protective substance called vernix caseosa that protects and moisturises the skin and may provide lubrication during childbirth. A silky hair called lanugo also covers the skin during weeks 17–20, but it is shed as the foetus continues to grow. Extremely premature infants sometimes exhibit residual lanugo.
Developmental weeks 21–30 are characterised by rapid weight gain, which is important for maintaining a stable body temperature after birth. The bone marrow completely takes over erythrocyte synthesis, and the axons of the spinal cord begin to be myelinated or coated in the electrically insulating glial cell sheaths that are necessary for efficient nervous system functioning. (The process of myelination is not completed until adolescence.) During this period, the foetus grows eyelashes. The eyelids are no longer fused and can be opened and closed. The lungs begin producing surfactant, a substance that reduces surface tension in the lungs and assists proper lung expansion after birth. Inadequate surfactant production in premature newborns may result in respiratory distress syndrome, and as a result, the newborn may require surfactant replacement therapy, supplemental oxygen, or maintenance in a continuous positive airway pressure (CPAP) chamber during their first days or weeks of life. In male foetuses, the testes descend into the scrotum near the end of this period. The foetus at 30 weeks measures 28 cm from crown to rump and exhibits the approximate body proportions of a full-term newborn, but still is much leaner.
The foetus continues to lay down subcutaneous fat from week 31 until birth. The added fat fills out the hypodermis and the skin transitions from red and wrinkled to soft and pink. Lanugo is shed, and the nails grow to the tips of the fingers and toes. Immediately before birth, the average crown-to-rump length is 35.5–40.5 cm, and the foetus weighs approximately 2.5–4 kg. Once born, the newborn is no longer confined to the foetal position, so subsequent measurements are made from head-to-toe instead of from crown-to-rump. At birth, the average length is approximately 51 cm.
Disorders of the Developing Foetus
Throughout the second half of gestation, the foetal intestines accumulate a tarry, greenish black meconium. The newborn’s first stools consist entirely of meconium; they later transition to seedy yellow stools or slightly formed tan stools as meconium is cleared and replaced with digested breast milk or formula, respectively. Unlike these later stools, meconium is sterile; it is devoid of bacteria because the foetus is in a sterile environment and has not consumed any breast milk or formula. Typically, an infant does not pass meconium until after birth. However, in 5–20 percent of births, the foetus has a bowel movement in utero, which can cause major complications in the newborn.
The passage of meconium in the uterus signals foetal distress, particularly foetal hypoxia (i.e., oxygen deprivation). This may be caused by maternal drug abuse (especially tobacco or cocaine), maternal hypertension, depletion of amniotic fluid, long labour or difficult birth, or a defect in the placenta that prevents it from delivering adequate oxygen to the foetus. Meconium passage is typically a complication of full-term or post-term newborns because it is rarely passed before 34 weeks of gestation, when the gastrointestinal system has matured and is appropriately controlled by nervous system stimuli. Foetal distress can stimulate the vagus nerve to trigger gastrointestinal peristalsis and relaxation of the anal sphincter. Notably, foetal hypoxic stress also induces a gasping reflex, increasing the likelihood that meconium will be inhaled into the foetal lungs.
Although meconium may be considered a sterile substance, it interferes with the antibiotic properties of the amniotic fluid and makes the newborn and mother more vulnerable to bacterial infections at birth and during the perinatal period. Specifically, inflammation of the foetal membranes, inflammation of the uterine lining, or neonatal sepsis (infection in the newborn) may occur. Meconium also irritates delicate foetal skin and can cause a rash.
The first sign that a foetus has passed meconium usually does not come until childbirth, when the amniotic sac ruptures. Normal amniotic fluid is clear and watery, but amniotic fluid in which meconium has been passed is stained greenish or yellowish. Antibiotics given to the mother may reduce the incidence of maternal bacterial infections, but it is critical that meconium is aspirated from the newborn before the first breath. Under these conditions, an obstetrician will extensively aspirate the infant’s airways as soon as the head is delivered, while the rest of the infant’s body is still inside the birth canal.
Aspiration of meconium with the first breath can result in laboured breathing, a barrel-shaped chest, or a low Apgar score. An obstetrician can identify meconium aspiration by listening to the lungs with a stethoscope for a coarse rattling sound. Blood gas tests and chest X-rays of the infant can confirm meconium aspiration. Inhaled meconium after birth could obstruct a newborn’s airways leading to alveolar collapse, interfere with surfactant function by stripping it from the lungs or cause pulmonary inflammation or hypertension. Any of these complications will make the newborn much more vulnerable to pulmonary infection, including pneumonia.
Section Review
The foetal period lasts from the ninth week of development until birth. During this period, male and female gonads differentiate. The foetal circulatory system becomes much more specialised and efficient than its embryonic counterpart. It includes three shunts—the ductus venosus, the foramen ovale and the ductus arteriosus—that enable it to bypass the semifunctional liver and pulmonary circuit until after childbirth. The brain continues to grow and its structures differentiate. Facial features develop, the body elongates and the skeleton ossifies. In the womb, the developing foetus moves, blinks, practices sucking and circulates amniotic fluid. The foetus grows from an embryo measuring approximately 3.3 cm and weighing 7 g to an infant measuring approximately 51 cm and weighing an average of approximately 3.4 kg. Embryonic organ structures that were primitive and non-functional develop to the point that the newborn can survive in the outside world.
Compare and contrast blood plasma, glomerular filtrate, and urine characteristics
Describe the characteristics of a normal urine sample, including normal range of pH, osmolarity, and volume
The urinary system’s ability to filter the blood resides in about 2 to 3 million tufts of specialised capillaries—the glomeruli—distributed more or less equally between the two kidneys. Because the glomeruli filter the blood based mostly on particle size, large elements like blood cells, platelets, antibodies, and albumen are excluded. The glomerulus is the first part of the nephron, which then continues as a highly specialised tubular structure responsible for creating the final urine composition. All other solutes, such as ions, amino acids, vitamins, and wastes, are filtered to create a filtrate composition very similar to plasma. The glomeruli create about 200 litres of this filtrate every day, yet you excrete less than two litres of waste per day of urine.
Characteristics of the urine vary, depending on influences such as water intake, exercise, environmental temperature, nutrient intake, and other factors (Table 17.1.1.). Some of the characteristics such as colour and odour are rough descriptors of your state of hydration, for example, if you exercise or work outside, and sweat a great deal, your urine will turn darker and produce a slight odour, even if you drink plenty of water. Athletes are often advised to consume water until their urine is clear. This is (mostly) good advice; however, it takes time for the kidneys to process body fluids and store it in the bladder. Another way of looking at this is that the quality of the urine produced is an average over the time it takes to make that urine. Producing clear urine may take only a few minutes if you are drinking a lot of water or several hours if you are working outside and not drinking much.
Table 17.1.1. Normal urine characteristics
Characteristic
Normal values
Colour
Pale yellow to deep amber
Odour
Odourless
Volume
750-2000mL/24h
pH
4.5-8.0
Specific gravity
1.003-1.0032
Osmolarity
40-1350 mOsmol/kg
Urobilinogen
0.2-1.0 mg/100mL
White blood cells
0-2 HPF (per high-power field of microscope)
Leukocyte esterase
None
Protein
None or trace
Bilirubin
<0.3 mg/100mL
Ketones
None
Nitrites
None
Blood
None
Glucose
None
Urinalysis (urine analysis) often provides clues to renal disease. Normally, only traces of protein are found in urine, and when higher amounts are found, damage to the glomeruli is the likely basis. Unusually large quantities of urine may point to diseases like diabetes mellitus or hypothalamic tumours that cause diabetes insipidus. The colour of urine is determined mostly by the breakdown products of red blood cell destruction (Figure 17.1.1). The “haem” of haemoglobin is converted by the liver into water-soluble forms that can be excreted into the bile and indirectly into the urine. This yellow pigment is urochrome. Urine colour may also be affected by certain foods like beets, berries and fava beans. A kidney stone or a cancer of the urinary system may produce sufficient bleeding to manifest as pink or even bright red urine. Diseases of the liver or obstructions of bile drainage from the liver impart a dark “tea” or “cola” hue to the urine. Dehydration produces darker, concentrated urine that may also possess the slight odour of ammonia. Most of the ammonia produced from protein breakdown is converted into urea by the liver, so ammonia is rarely detected in fresh urine. The strong ammonia odour you may detect in bathrooms or alleys is due to the breakdown of urea into ammonia by bacteria in the environment. About one in five people detect a distinctive odour in their urine after consuming asparagus; other foods such as onions, garlic and fish can impart their own aromas! These food-caused odours are harmless.
Figure 17.1.1. Urine Colour.
Urine volume varies considerably. The normal range is one to two litres per day (Table 17.1.2). The kidneys must produce a minimum urine volume of about 500 mL/day to rid the body of wastes. Output below this level may be caused by severe dehydration or renal disease and is termed oliguria. The virtual absence of urine production is termed anuria. Excessive urine production is polyuria, which may be due to diabetes mellitus or diabetes insipidus. In diabetes mellitus, blood glucose levels exceed the number of available sodium-glucose transporters in the kidney and glucose appears in the urine. The osmotic nature of glucose attracts water, leading to its loss in the urine. In the case of diabetes insipidus, insufficient pituitary antidiuretic hormone (ADH) release or insufficient numbers of ADH receptors in the collecting ducts means that too few water channels are inserted into the cell membranes that line the collecting ducts of the kidney. Insufficient numbers of water channels (aquaporins) reduce water absorption, resulting in high volumes of very dilute urine.
Table 17.1.2. Urine volumes
Volume condition
Volume
Causes
Normal
1-2 L/day
Polyuria
>2.5 L/day
Diabetes mellitus; diabetes insipidus; excess caffeine or alcohol; kidney disease; certain drugs, such as diuretics; sickle cell anaemia; excessive water intake
Kidney failure; obstruction, such as kidney stone or tumour; enlarged prostate
The pH (hydrogen ion concentration) of the urine can vary more than 1000-fold, from a normal low of 4.5 to a maximum of 8.0. Diet can influence pH; meats lower the pH, whereas citrus fruits, vegetables and dairy products raise the pH. Chronically high or low pH can lead to disorders, such as the development of kidney stones or osteomalacia.
Specific gravity is a measure of the quantity of solutes per unit volume of a solution and is traditionally easier to measure than osmolarity. Urine will always have a specific gravity greater than pure water (water = 1.0) due to the presence of solutes. Laboratories can now measure urine osmolarity directly, which is a more accurate indicator of urinary solutes than specific gravity. Remember that osmolarity is the number of osmoles or milliosmoles per litre of fluid (mOsmol/L). Urine osmolarity ranges from a low of 50–100 mOsmol/L to as high as 1200 mOsmol/L H2O.
Cells are not normally found in the urine. The presence of leukocytes may indicate a urinary tract infection. Leukocyte esterase is released by leukocytes; if detected in the urine, it can be taken as indirect evidence of a urinary tract infection (UTI).
Protein does not normally leave the glomerular capillaries, so only trace amounts of protein should be found in the urine, approximately 10 mg/100 mL in a random sample. If excessive protein is detected in the urine, it usually means that the glomerulus is damaged and is allowing protein to “leak” into the filtrate.
Ketones are by-products of fat metabolism. Finding ketones in the urine suggests that the body is using fat as an energy source in preference to glucose. In diabetes mellitus when there is not enough insulin (Type I diabetes mellitus) or because of insulin resistance (Type 2 diabetes mellitus), there is plenty of glucose, but without the action of insulin, the cells cannot take it up, so it remains in the bloodstream. Instead, the cells are forced to use fat as their energy source, and fat consumed at such a level produces excessive ketones as by-products. These excess ketones will appear in the urine. Ketones may also appear if there is a severe deficiency of proteins or carbohydrates in the diet.
Nitrates (NO3–) occur normally in the urine. Gram-negative bacteria metabolise nitrate into nitrite (NO2–), and its presence in the urine is indirect evidence of infection.
There should be no blood found in the urine. It may sometimes appear in urine samples as a result of menstrual contamination, but this is not an abnormal condition. Now that you understand what the normal characteristics of urine are, the next section will introduce you to how you store and dispose of this waste product and how you make it.
Section Review
The kidney glomerulus filters blood mainly based on particle size to produce a filtrate lacking cells or large proteins. Most of the ions and molecules in the filtrate are needed by the body and must be reabsorbed farther down the nephron tubules, resulting in the formation of urine. Urine characteristics change depending on water intake, exercise, environmental temperature, and nutrient intake. Urinalysis analyses characteristics of the urine and is used to diagnose diseases. A minimum of 400 to 500 mL urine must be produced daily to rid the body of wastes. Excessive quantities of urine may indicate diabetes insipidus or diabetes mellitus. The pH range of urine is 4.5 to 8.0 and is affected by diet. Osmolarity ranges from 50 to 1200 milliosmoles and reflects the amount of water being recovered or lost by renal nephrons.
Identify the ureters, urinary bladder, and urethra, as well as their location, structure, histology and function
Compare and contrast male and female urethras
Describe the micturition reflex
Describe voluntary and involuntary neural control of micturition
Rather than start with urine formation, this section will start with urine excretion. Urine is a fluid of variable composition that requires specialised structures to remove it from the body safely and efficiently. Blood is filtered and the filtrate is transformed into urine at a relatively constant rate throughout the day. This processed liquid is stored until a convenient time for excretion. All structures involved in the transport and storage of the urine are large enough to be visible to the naked eye. This transport and storage system not only stores the waste, but it protects the tissues from damage due to the wide range of pH and osmolarity of the urine, prevents infection by foreign organisms and for the male, provides reproductive functions.
Urethra
The urethra transports urine from the bladder to the outside of the body for disposal. The urethra is the only urologic organ that shows any significant anatomic difference between males and females; all other urine transport structures are identical (Figure 17.2.1).
Figure 17.2.1. Female and male urethras. The urethra transports urine from the bladder to the outside of the body. This image shows (a) a female urethra and (b) a male urethra.
The urethra in both males and females begins inferior and central to the two ureteral openings forming the three points of a triangular-shaped area at the base of the bladder called the trigone (Greek tri- = “triangle” and the root of the word “trigonometry”). The urethra tracks posterior and inferior to the pubic symphysis (see Figure 17.2.1a). In both males and females, the proximal urethra is lined by transitional epithelium, whereas the terminal portion is a nonkeratinised, stratified squamous epithelium. In the male, pseudostratified columnar epithelium lines the urethra between these two cell types. Voiding is regulated by an involuntary autonomic nervous system-controlled internal urinary sphincter, consisting of smooth muscle and voluntary skeletal muscle that forms the external urinary sphincter below it.
Female Urethra
The external urethral orifice is embedded in the anterior vaginal wall inferior to the clitoris, superior to the vaginal opening (introitus), and medial to the labia minora. Its short length, about 4 cm, is less of a barrier to faecal bacteria than the longer male urethra and the best explanation for the greater incidence of UTI in women. Voluntary control of the external urethral sphincter is a function of the pudendal nerve. It arises in the sacral region of the spinal cord, travelling via the S2–S4 nerves of the sacral plexus.
Male Urethra
The male urethra passes through the prostate gland immediately inferior to the bladder before passing below the pubic symphysis (see Figure 17.2.1b). The length of the male urethra varies between men but averages 20 cm in length. It is divided into four regions: the preprostatic urethra, the prostatic urethra, the membranous urethra and the spongy or penile urethra. The preprostatic urethra is very short and incorporated into the bladder wall. The prostatic urethra passes through the prostate gland. During sexual intercourse, it receives sperm via the ejaculatory ducts and secretions from the seminal vesicles. Paired Cowper’s glands (bulbourethral glands) produce and secrete mucus into the urethra to buffer urethral pH during sexual stimulation. The mucus neutralises the usually acidic environment and lubricates the urethra, decreasing the resistance to ejaculation. The membranous urethra passes through the deep muscles of the perineum, where it is invested by the overlying urethral sphincters. The spongy urethra exits at the tip (external urethral orifice) of the penis after passing through the corpus spongiosum. Mucous glands are found along much of the length of the urethra and protect the urethra from extremes of urine pH. Innervation is the same in both males and females.
Bladder
The urinary bladder collects urine from both ureters (Figure 17.2.2). The bladder lies anterior to the uterus in females, posterior to the pubic bone and anterior to the rectum. During late pregnancy, its capacity is reduced due to compression by the enlarging uterus, resulting in increased frequency of urination. In males, the anatomy is similar, minus the uterus and with the addition of the prostate inferior to the bladder. The bladder is partially retroperitoneal (outside the peritoneal cavity) with its peritoneal covered “dome” projecting into the abdomen when the bladder is distended with urine.
The bladder is a highly distensible organ comprised of irregular crisscrossing bands of smooth muscle collectively called the detrusor muscle. The interior surface is made of transitional cellular epithelium that is structurally suited for the large volume fluctuations of the bladder. When empty, it resembles columnar epithelia, but when stretched, it “transitions” (hence the name) to a squamous appearance (see Figure 17.2.2). Capacity of the bladder varies with age and volumes in adults can range from nearly zero to 500–600 mL.
The detrusor muscle contracts with significant force in the young. The bladder’s strength diminishes with age, but voluntary contractions of abdominal skeletal muscles can increase intra-abdominal pressure to promote more forceful bladder emptying. Such voluntary contraction is also used in forceful defaecation and childbirth.
Micturition Reflex
Micturition is a less-often used, but proper term for urination or voiding. It results from an interplay of involuntary and voluntary actions by the internal and external urethral sphincters. When bladder volume reaches about 150 mL, an urge to void is sensed but is easily overridden. Voluntary control of urination relies on consciously preventing relaxation of the external urethral sphincter to maintain urinary continence. As the bladder fills, subsequent urges become harder to ignore. Ultimately, voluntary constraint fails and may result in incontinence, which will occur as bladder volume approaches 300 to 400 mL.
Normal micturition is a result of stretch receptors in the bladder wall that transmit nerve impulses to the sacral region of the spinal cord to generate a spinal reflex. The resulting parasympathetic neural outflow causes contraction of the detrusor muscle and relaxation of the involuntary internal urethral sphincter. At the same time, the spinal cord inhibits somatic motor neurons, resulting in the relaxation of the skeletal muscle of the external urethral sphincter. The micturition reflex is active in infants but with maturity, children learn to override the reflex by asserting external sphincter control, thereby delaying voiding (potty training). This reflex may be preserved even in the face of spinal cord injury that results in paraplegia or quadriplegia. However, relaxation of the external sphincter may not be possible in all cases, and therefore, periodic catheterisation may be necessary for bladder emptying.
Nerves involved in the control of urination include the hypogastric, pelvic and pudendal (Figure 17.2.3). Voluntary micturition requires an intact spinal cord and functional pudendal nerve arising from the sacral micturition centre. Since the external urinary sphincter is voluntary skeletal muscle, actions by cholinergic neurons maintain contraction (and thereby continence) during filling of the bladder. At the same time, sympathetic nervous activity via the hypogastric nerves suppresses contraction of the detrusor muscle. With further bladder stretch, afferent signals travelling over sacral pelvic nerves activate parasympathetic neurons. This activates efferent neurons to release acetylcholine at the neuromuscular junctions, producing detrusor contraction and bladder emptying.
Figure 17.2.3. Nerves Innervating the urinary system.
Ureters
The kidneys and ureters are completely retroperitoneal, and the bladder has a peritoneal covering only over the dome. As urine is formed, it drains into the calyces of the kidney, which merge to form the funnel-shaped renal pelvis in the hilum of each kidney. The hilum narrows to become the ureter of each kidney. As urine passes through the ureter, it does not passively drain into the bladder but rather is propelled by waves of peristalsis. As the ureters enter the pelvis, they sweep laterally, hugging the pelvic walls. As they approach the bladder, they turn medially and pierce the bladder wall obliquely. This is important because it creates a one-way valve (a physiological sphincter rather than an anatomical sphincter) that allows urine into the bladder but prevents reflux of urine from the bladder back into the ureter. Children born lacking this oblique course of the ureter through the bladder wall are susceptible to “vesicoureteral reflux,” which dramatically increases their risk of serious UTI. Pregnancy also increases the likelihood of reflux and UTI.
The ureters are approximately 30 cm long. The inner mucosa is lined with transitional epithelium (Figure 17.2.4) and scattered goblet cells that secrete protective mucus. The muscular layer of the ureter consists of longitudinal and circular smooth muscles that create the peristaltic contractions to move the urine into the bladder without the aid of gravity. Finally, a loose adventitial layer composed of collagen and fat anchors the ureters between the parietal peritoneum and the posterior abdominal wall.
The urethra is the only urinary structure that differs significantly between males and females. This is due to the dual role of the male urethra in transporting both urine and semen. The urethra arises from the trigone area at the base of the bladder. Urination is controlled by an involuntary internal sphincter of smooth muscle and a voluntary external sphincter of skeletal muscle. The shorter female urethra contributes to the higher incidence of bladder infections in females. The male urethra receives secretions from the prostate gland, Cowper’s gland, and seminal vesicles as well as sperm. The bladder is largely retroperitoneal and can hold up to approximately 500–600 mL urine. Micturition is the process of voiding the urine and involves both involuntary and voluntary actions. Voluntary control of micturition requires a mature and intact sacral micturition centre. It also requires an intact spinal cord. Loss of control of micturition is called incontinence. The ureters are retroperitoneal and lead from the renal pelvis of the kidney to the trigone area at the base of the bladder. A thick muscular wall consisting of longitudinal and circular smooth muscle helps move urine toward the bladder by way of peristaltic contractions.
Describe the external structure of the kidney, including its location, support structures and covering
Identify the major internal divisions and structures of the kidney
Identify the major blood vessels associated with the kidney and trace the path of blood through the kidney
Compare and contrast the cortical and juxtamedullary nephrons
Name structures found in the cortex and medulla
Describe the physiological characteristics of the cortex and medulla
The kidneys lie on either side of the spine in the retroperitoneal space between the parietal peritoneum and the posterior abdominal wall, well protected by muscle, fat, and ribs. They are roughly the size of your fist, and the male kidney is typically a bit larger than the female kidney. The kidneys are well vascularised, receiving about 25 percent of the cardiac output at rest.
External Anatomy
In humans, the left kidney is located at about the T12 to L3 vertebrae, whereas the right is lower due to slight displacement by the liver. Upper portions of the kidneys are somewhat protected by the eleventh and twelfth ribs (Figure 17.3.1). Each kidney weighs about 125–175 g in males and 115–155 g in females. They are about 11–14 cm in length, 6 cm wide, and 4 cm thick and are directly covered by a fibrous capsule composed of dense, irregular connective tissue that helps to hold their shape and protect them. This capsule is covered by a shock-absorbing layer of adipose tissue called the renal fat pad, which in turn is encompassed by a tough renal fascia. The fascia and, to a lesser extent, the overlying peritoneum serve to firmly anchor the kidneys to the posterior abdominal wall in a retroperitoneal position.
Figure 17.3.1. Kidneys. The kidneys are slightly protected by the ribs and are surrounded by fat for protection (not shown).
On the superior aspect of each kidney is the adrenal gland. The adrenal cortex directly influences renal function through the production of the hormone aldosterone to stimulate sodium reabsorption.
Internal Anatomy
A frontal section through the kidney reveals an outer region called the renal cortex and an inner region called the medulla (Figure 17.3.2). The renal columns are connective tissue extensions that radiate downward from the cortex through the medulla to separate the most characteristic features of the medulla, the renal pyramids and renal papillae. The papillae are bundles of collecting ducts that transport urine made by nephrons to the calyces of the kidney for excretion. The renal columns also serve to divide the kidney into six-eight lobes and provide a supportive framework for vessels that enter and exit the cortex. The pyramids and renal columns taken together constitute the kidney lobes.
Figure 17.3.2. Left Kidney.
Renal Hilum
The renal hilum is the entry and exit site for structures servicing the kidneys: vessels, nerves, lymphatics and ureters. The medial-facing hila are tucked into the sweeping convex outline of the cortex. Emerging from the hilum is the renal pelvis, which is formed from the major and minor calyxes in the kidney. The smooth muscle in the renal pelvis funnels urine, via peristalsis, into the ureter. The renal arteries form directly from the descending aorta, whereas the renal veins return ‘cleansed’ blood directly to the inferior vena cava. The artery, vein and renal pelvis are arranged in an anterior-to-posterior order.
Nephrons and Vessels
The renal artery first divides into segmental arteries, followed by further branching to form interlobar arteries that pass through the renal columns to reach the cortex (Figure 17.3.3). The interlobar arteries, in turn, branch into arcuate arteries, cortical radiate arteries and then into afferent arterioles. The afferent arterioles service about 1.3 million nephrons in each kidney.
Figure 17.3.3. Blood Flow in the Kidney.
Nephrons are the “functional units” of the kidney; they cleanse the blood and balance the constituents of the circulation. The afferent arterioles form a tuft of high-pressure capillaries about 200 µm in diameter, the glomerulus. The rest of the nephron consists of a continuous sophisticated tubule whose proximal end surrounds the glomerulus in an intimate embrace—this is Bowman’s (or glomerular) capsule. The glomerulus and Bowman’s capsule together form the renal corpuscle. As mentioned earlier, these glomerular capillaries filter the blood based on particle size. After passing through the renal corpuscle, the capillaries form a second arteriole, the efferent arteriole (Figure 17.3.4). These will next form a capillary network around the more distal portions of the nephron tubule, the peritubular capillaries and vasa recta, before returning to the venous system. As the glomerular filtrate progresses through the nephron, these capillary networks recover most of the solutes and water and return them to the circulation. Since a capillary bed (the glomerulus) drains into a vessel that in turn forms a second capillary bed, the definition of a portal system is met. This is the only portal system in which an arteriole is found between the first and second capillary beds. (Portal systems also link the hypothalamus to the anterior pituitary, and the blood vessels of the digestive viscera to the liver.)
Figure 17.3.4. Blood flow in the nephron. The two capillary beds are clearly shown in this figure. The efferent arteriole is the connecting vessel between the glomerulus and the peritubular capillaries and vasa recta.
Cortex
In a dissected kidney, it is easy to identify the cortex; it appears lighter in colour compared to the rest of the kidney. All of the renal corpuscles as well as both the proximal convoluted tubules (PCTs) and distal convoluted tubules are found here. Some nephrons have a short loop of Henle that does not dip beyond the cortex. These nephrons are called cortical nephrons. About 15 percent of nephrons have long loops of Henle that extend deep into the medulla and are called juxtamedullary nephrons.
Section Review
As noted previously, the structure of the kidney is divided into two principal regions—the peripheral rim of cortex and the central medulla. The two kidneys receive about 25 percent of cardiac output. They are protected in the retroperitoneal space by the renal fat pad and overlying ribs and muscle. Ureters, blood vessels, lymph vessels and nerves enter and leave at the renal hilum. The renal arteries arise directly from the aorta, and the renal veins drain directly into the inferior vena cava. Kidney function is derived from the actions of about 1.3 million nephrons per kidney; these are the “functional units.” A capillary bed, the glomerulus, filters blood and the filtrate is captured by Bowman’s capsule. A portal system is formed when the blood flows through a second capillary bed surrounding the proximal and distal convoluted tubules and the loop of Henle. Most water and solutes are recovered by this second capillary bed. This filtrate is processed and finally gathered by collecting ducts that drain into the minor calyces, which merge to form major calyces; the filtrate then proceeds to the renal pelvis and finally the ureters.
Distinguish the histological differences between the renal cortex and medulla
Describe the structure of the filtration membrane
Identify the major structures and subdivisions of the renal corpuscles, renal tubules and renal capillaries
Discuss the function of the peritubular capillaries and vasa recta
Identify the location of the juxtaglomerular apparatus and describe the cells that line it
Describe the histology of the proximal convoluted tubule, loop of Henle, distal convoluted tubule and collecting ducts
The renal structures that conduct the essential work of the kidney cannot be seen by the naked eye. Only a light or electron microscope can reveal these structures. Even then, serial sections and computer reconstruction are necessary to give us a comprehensive view of the functional anatomy of the nephron and its associated blood vessels.
Nephrons: the Functional Unit
Nephrons take a simple filtrate of the blood and modify it into urine. Many changes take place in the different parts of the nephron before urine is created for disposal. The term ‘forming urine’ will be used hereafter to describe the filtrate as it is modified into true urine. The main task of the nephron population is to balance the plasma to homeostatic set points and excrete potential toxins in the urine. They do this by accomplishing three principal functions—filtration, reabsorption and secretion. They also have additional secondary functions that exert control in three areas: blood pressure (via production of renin), red blood cell production (via the hormone EPO) and calcium absorption (via conversion of calcidiol into calcitriol, the active form of vitamin D).
Renal Corpuscle
As discussed earlier, the renal corpuscle consists of a tuft of capillaries called the glomerulus that is largely surrounded by Bowman’s (glomerular) capsule. The glomerulus is a high-pressure capillary bed between afferent and efferent arterioles. Bowman’s capsule surrounds the glomerulus to form a lumen and captures and directs this filtrate to the PCT. The outermost part of Bowman’s capsule, the parietal layer, is a simple squamous epithelium. It transitions onto the glomerular capillaries in an intimate embrace to form the visceral layer of the capsule. Here, the cells are not squamous, but uniquely shaped cells (podocytes) extending finger-like arms (pedicels) to cover the glomerular capillaries (Figure 17.4.1). These projections interdigitate to form filtration slits, leaving small gaps between the digits to form a sieve. As blood passes through the glomerulus, 10 to 20 percent of the plasma filters between these sieve-like fingers to be captured by Bowman’s capsule and funnelled to the PCT. Where the fenestrae (windows) in the glomerular capillaries match the spaces between the podocyte “fingers,” the only thing separating the capillary lumen and the lumen of Bowman’s capsule is their shared basement membrane (Figure 17.4.2). These three features comprise what is known as the filtration membrane. This membrane permits very rapid movement of filtrate from capillary to capsule though pores that are only 70 nm in diameter.
Figure 17.4.1. Podocytes. Podocytes interdigitate with structures called pedicels and filter substances in a way similar to fenestrations. In (a), the large cell body can be seen at the top right corner, with branches extending from the cell body. The smallest finger-like extensions are the pedicels. Pedicels on one podocyte always interdigitate with the pedicels of another podocyte. (b) This capillary has three podocytes wrapped around it.Figure 17.4.2. Fenestrated capillary. Fenestrations allow many substances to diffuse from the blood based primarily on size.
The fenestrations prevent filtration of blood cells or large proteins but allow most other constituents through. These substances cross readily if they are less than 4 nm in size and most pass freely up to 8 nm in size. An additional factor affecting the ability of substances to cross this barrier is their electric charge. The proteins associated with these pores are negatively charged, so they tend to repel negatively charged substances and allow positively charged substances to pass more readily. The basement membrane prevents filtration of medium-to-large proteins such as globulins. There are also mesangial cells in the filtration membrane that can contract to help regulate the rate of filtration of the glomerulus. Overall, filtration is regulated by fenestrations in capillary endothelial cells, podocytes with filtration slits, membrane charge, and the basement membrane between capillary cells. The result is the creation of a filtrate that does not contain cells or large proteins and has a slight predominance of positively charged substances.
Lying just outside Bowman’s capsule and the glomerulus is the juxtaglomerular apparatus (JGA) (Figure 17.4.3). At the juncture where the afferent and efferent arterioles enter and leave Bowman’s capsule, the initial part of the distal convoluted tubule (DCT) comes into direct contact with the arterioles. The wall of the DCT at that point forms a part of the JGA known as the macula densa. This cluster of cuboidal epithelial cells monitors the fluid composition of fluid flowing through the DCT. In response to the concentration of Na+ in the fluid flowing past them, these cells release paracrine signals. They also have a single, nonmotile cilium that responds to the rate of fluid movement in the tubule. The paracrine signals released in response to changes in flow rate and Na+ concentration are adenosine triphosphate (ATP) and adenosine.
A second cell type in this apparatus is the juxtaglomerular cell. This is a modified, smooth muscle cell lining the afferent arteriole that can contract or relax in response to ATP or adenosine released by the macula densa. Such contraction and relaxation regulate blood flow to the glomerulus. If the osmolarity of the filtrate is too high (hyperosmotic), the juxtaglomerular cells will contract, decreasing the glomerular filtration rate (GFR) so less plasma is filtered, leading to less urine formation and greater retention of fluid. This will ultimately decrease blood osmolarity toward the physiologic norm. If the osmolarity of the filtrate is too low, the juxtaglomerular cells will relax, increasing the GFR and enhancing the loss of water to the urine, causing blood osmolarity to rise. In other words, when osmolarity goes up, filtration and urine formation decrease and water is retained. When osmolarity goes down, filtration and urine formation increase and water is lost by way of the urine. The net result of these opposing actions is to keep the rate of filtration relatively constant. A second function of the macula densa cells is to regulate renin release from the juxtaglomerular cells of the afferent arteriole (Figure 17.4.4). Active renin is a protein comprised of 304 amino acids that cleaves several amino acids from angiotensinogen to produce angiotensin I. Angiotensin I is not biologically active until converted to angiotensin II by angiotensin-converting enzyme (ACE) from the lungs. Angiotensin II is a systemic vasoconstrictor that helps to regulate blood pressure by increasing it. Angiotensin II also stimulates the release of the steroid hormone aldosterone from the adrenal cortex. Aldosterone stimulates Na+ reabsorption by the kidney, which also results in water retention and increased blood pressure.
Figure 17.4.4. Conversion of angiotensin I to angiotensin II. The enzyme renin catalyses the conversion of angiotensinogen to angiotensin I; the lung-derived enzyme ACE converts angiotensin I into active angiotensin II.
Proximal Convoluted Tubule (PCT)
Filtered fluid collected by Bowman’s capsule enters the PCT. It is called convoluted due to its tortuous path. Simple cuboidal cells form this tubule with prominent microvilli on the luminal surface, forming a brush border. These microvilli create a large surface area to maximise the absorption and secretion of solutes (Na+, Cl–, glucose, etc.), the most essential function of this portion of the nephron. These cells actively transport ions across their membranes, so they possess a high concentration of mitochondria to produce sufficient ATP.
Loop of Henle
The descending and ascending portions of the loop of Henle (sometimes referred to as the nephron loop) are, of course, just continuations of the same tubule. They run adjacent and parallel to each other after having made a hairpin turn at the deepest point of their descent. The descending loop of Henle consists of an initial short, thick portion and long, thin portion, whereas the ascending loop consists of an initial short, thin portion followed by a long, thick portion. The descending thick portion consists of simple cuboidal epithelium similar to that of the PCT. The descending and ascending thin portions consist of simple squamous epithelium. As you will see later, these are important differences, since different portions of the loop have different permeabilities for solutes and water. The ascending thick portion consists of simple cuboidal epithelium similar to the DCT.
Distal Convoluted Tubule (DCT)
The DCT, like the PCT, is very tortuous and formed by simple cuboidal epithelium, but it is shorter than the PCT. These cells are not as active as those in the PCT; thus, there are fewer microvilli on the apical surface. However, these cells must also pump ions against their concentration gradient, so there are a large number of mitochondria, although fewer than in the PCT.
Collecting Ducts
The collecting ducts are continuous with the nephron but not technically part of it. In fact, each duct collects filtrate from several nephrons for final modification. Collecting ducts merge as they descend deeper in the medulla to form about 30 terminal ducts, which empty at a papilla. They are lined with simple squamous epithelium with receptors for ADH. When stimulated by ADH, these cells will insert aquaporin channel proteins into their membranes, which as their name suggests, allow water to pass from the duct lumen through the cells and into the interstitial spaces to be recovered by the vasa recta. This process allows for the recovery of large amounts of water from the filtrate back into the blood. In the absence of ADH, these channels are not inserted, resulting in the excretion of water in the form of dilute urine. Most, if not all, cells of the body contain aquaporin molecules, whose channels are so small that only water can pass. At least 10 types of aquaporins are known in humans, and six of those are found in the kidney. The function of all aquaporins is to allow the movement of water across the lipid-rich, hydrophobic cell membrane (Figure 17.4.5).
Figure 17.4.5. Aquaporin water channel. Positive charges inside the channel prevent the leakage of electrolytes across the cell membrane, while allowing water to move due to osmosis.
Section Review
The functional unit of the kidney, the nephron, consists of the renal corpuscle, PCT, loop of Henle and DCT. Cortical nephrons have short loops of Henle, whereas juxtamedullary nephrons have long loops of Henle extending into the medulla. About 15 percent of nephrons are juxtamedullary. The glomerulus is a capillary bed that filters blood principally based on particle size. The filtrate is captured by Bowman’s capsule and directed to the PCT. A filtration membrane is formed by the fused basement membranes of the podocytes and the capillary endothelial cells that they embrace. Contractile mesangial cells further perform a role in regulating the rate at which the blood is filtered. Specialised cells in the JGA produce paracrine signals to regulate blood flow and filtration rates of the glomerulus. Other JGA cells produce the enzyme renin, which plays a leading role in blood pressure regulation. The filtrate enters the PCT where absorption and secretion of several substances occur. The descending and ascending limbs of the loop of Henle consist of thick and thin segments. Absorption and secretion continue in the DCT but to a lesser extent than in the PCT. Each collecting duct collects forming urine from several nephrons and responds to the posterior pituitary hormone ADH by inserting aquaporin water channels into the cell membrane to fine tune water recovery.
Describe the hydrostatic and colloid osmotic forces that favour and oppose filtration
Describe glomerular filtration rate (GFR), state the average value of GFR and explain how clearance rate can be used to measure GFR
Predict specific factors that will increase or decrease GFR
State the percent of the filtrate that is normally reabsorbed and explain why the process of reabsorption is so important
Calculate daily urine production
List common symptoms of kidney failure
Having reviewed the anatomy and microanatomy of the urinary system, now is the time to focus on the physiology. You will discover that different parts of the nephron utilise specific processes to produce urine: filtration, reabsorption and secretion. You will learn how each of these processes works and where they occur along the nephron and collecting ducts. The physiologic goal is to modify the composition of the plasma and, in doing so, produce the waste product urine.
Failure of the renal anatomy and/or physiology can lead suddenly or gradually to renal failure. In this event, a number of symptoms, signs, or laboratory findings point to the diagnosis (Table 17.5.1).
Table 17.5.1. Symptoms of Kidney Failure
Symptoms
Weakness
Lethargy
Shortness of breath
Widespread oedema
Anaemia
Metabolic acidosis
Metabolic alkalosis
Heart arrhythmias
Uraemia (high urea level in the blood)
Loss of appetite
Fatigue
Excessive urination
Oliguria (too little urine output)
Glomerular Filtration Rate (GFR)
The volume of filtrate formed by both kidneys per minute is termed the glomerular filtration rate (GFR). The heart pumps about 5 L blood per min under resting conditions. Approximately 20-25 percent or one to 1.5 litres enters the kidneys to be filtered. On average, this results in the production of about 125 mL/min filtrate produced in men (range of 90 to 140 mL/min) and 105 mL/min filtrate produced in women (range of 80 to 125 mL/min). This then equates to a volume of about 180 L/day in men and 150 L/day in women. Ninety-nine percent of this filtrate is returned to the circulation by reabsorption so that only about 1–2 litres of urine are produced per day (Table 17.5.2).
Table 17.5.2. Calculating urine formation per day
Flow
Flow per minute (mL)
Calculation
Renal blood flow
1050
Cardiac output is about 5000 mL/minute, of which 21% flows through the kidney
5000*0.21 = 1050 mL blood/min
Renal plasma flow
578
Renal plasma flow equals to the blood flow per minute times the haematocrit. If a person has a haematocrit of 45, then the renal plasma flow is 55%.
1050*0.55 = 578 mL plasma/min
Glomerular filtration rate
110
The GFR is the amount of plasma entering Bowman’s capsule per minute. It is the renal plasma flow times the filtration that enters the renal capsule (19%).
587*0.55 = 110 mL filtrate/min
Urine
1296
The filtrate not recovered by the kidney is the urine that will be eliminated. It is the GFR times the fraction of the filtrate that is not reabsorbed (0.8%).
110*0.008 = 0.9 mL urine/min
Multiply urine/min times 60 minutes times 24 hours to get daily urine production.
0.9*60*24 = 1296 mL/day urine
GFR is influenced by the hydrostatic pressure and colloid osmotic pressure on either side of the capillary membrane of the glomerulus. Recall that filtration occurs as pressure forces fluid and solutes through a semipermeable barrier with the solute movement constrained by particle size. Hydrostatic pressure is the pressure produced by a fluid against a surface. If you have a fluid on both sides of a barrier, both fluids exert a pressure in opposing directions. Net fluid movement will be in the direction of the lower pressure. Osmosis is the movement of solvent (water) across a membrane that is impermeable to a solute in the solution. This creates a pressure, osmotic pressure, which will exist until the solute concentration is the same on both sides of a semipermeable membrane. If the concentration differs, water will move. Glomerular filtration occurs when glomerular hydrostatic pressure exceeds the luminal hydrostatic pressure of Bowman’s capsule. There is also an opposing force, the osmotic pressure, which is typically higher in the glomerular capillary.
To understand why this is so, look more closely at the microenvironment on either side of the filtration membrane. You will find osmotic pressure exerted by the solutes inside the lumen of the capillary as well as inside of Bowman’s capsule. Since the filtration membrane limits the size of particles crossing the membrane, the osmotic pressure inside the glomerular capillary is higher than the osmotic pressure in Bowman’s capsule. Recall that cells and the medium-to-large proteins cannot pass between the podocyte processes or through the fenestrations of the capillary endothelial cells. This means that red and white blood cells, platelets, albumins, and other proteins too large to pass through the filter remain in the capillary, creating an average colloid osmotic pressure of 30 mm Hg within the capillary. The absence of proteins in Bowman’s space (the lumen within Bowman’s capsule) results in an osmotic pressure near zero. Thus, the only pressure moving fluid across the capillary wall into the lumen of Bowman’s space is hydrostatic pressure. Hydrostatic (fluid) pressure is sufficient to push water through the membrane despite the osmotic pressure working against it. The sum of all the influences, both osmotic and hydrostatic, results in a net filtration pressure (NFP) of about 10 mm Hg (Figure 17.5.1).
Figure 17.5.1. Net Filtration pressure. The NFP is the sum of osmotic and hydrostatic pressures.
A proper concentration of solutes in the blood is important in maintaining osmotic pressure both in the glomerulus and systemically. There are disorders in which too much protein passes through the filtration slits into the kidney filtrate. This excess protein in the filtrate leads to a deficiency of circulating plasma proteins. In turn, the presence of protein in the urine increases its osmolarity; this holds more water in the filtrate and results in an increase in urine volume. Because there is less circulating protein, principally albumin, the osmotic pressure of the blood falls. Less osmotic pressure pulling water into the capillaries tips the balance towards hydrostatic pressure, which tends to push it out of the capillaries. The net effect is that water is lost from the circulation to interstitial tissues and cells. This “plumps up” the tissues and cells, a condition termed systemic oedema.
Net Filtration Pressure (NFP)
NFP determines filtration rates through the kidney. It is determined as follows:
As you can see, there is a low net pressure across the filtration membrane. Intuitively, you should realise that minor changes in osmolarity of the blood or changes in capillary blood pressure result in major changes in the amount of filtrate formed at any given point in time. The kidney can cope with a wide range of blood pressures. In large part, this is due to the autoregulatory nature of smooth muscle. When you stretch it, it contracts. Thus, when blood pressure goes up, smooth muscle in the afferent capillaries contracts to limit any increase in blood flow and filtration rate. When blood pressure drops, the same capillaries relax to maintain blood flow and filtration rate. The net result is a steady flow of blood into the glomerulus and a steady filtration rate despite significant systemic blood pressure changes. Mean arterial blood pressure is calculated by adding 1/3 of the difference between the systolic and diastolic pressures to the diastolic pressure. Therefore, if the blood pressure is 110/80, the difference between systolic and diastolic pressure is 30. One third of this is 10, and when you add this to the diastolic pressure of 80, you arrive at a calculated mean arterial pressure of 90 mm Hg. Therefore, if you use mean arterial pressure for the GBHP in the formula for calculating NFP, you can determine that if mean arterial pressure is above approximately 60 mm Hg, the pressure will be adequate to maintain glomerular filtration. Blood pressures below this level will impair renal function and cause systemic disorders that are severe enough to threaten survival. This condition is called shock.
Determination of the GFR is one of the tools used to assess the kidney’s excretory function. This is more than just an academic exercise. Since many drugs are excreted in the urine, a decline in renal function can lead to toxic accumulations. Additionally, administration of appropriate drug dosages for those drugs primarily excreted by the kidney requires an accurate assessment of GFR. GFR can be estimated closely by intravenous administration of inulin. Inulin is a plant polysaccharide that is neither reabsorbed nor secreted by the kidney. Its appearance in the urine is directly proportional to the rate at which it is filtered by the renal corpuscle. However, since measuring inulin clearance is cumbersome in the clinical setting, most often, the GFR is estimated by measuring naturally occurring creatinine, a protein-derived molecule produced by muscle metabolism that is not reabsorbed and only slightly secreted by the nephron.
Section Review
The entire volume of the blood is filtered through the kidneys about 300 times per day and 99 percent of the water filtered is recovered. The GFR is influenced by hydrostatic pressure and colloid osmotic pressure. Under normal circumstances, hydrostatic pressure is significantly greater and filtration occurs. The hydrostatic pressure of the glomerulus depends on systemic blood pressure, autoregulatory mechanisms, sympathetic nervous activity and paracrine hormones. The kidney can function normally under a wide range of blood pressures due to the autoregulatory nature of smooth muscle.
List specific transport mechanisms occurring in different parts of the nephron, including active transport, osmosis, facilitated diffusion and passive electrochemical gradients
List the different membrane proteins of the nephron, including channels, transporters and ATPase pumps
Compare and contrast passive and active tubular reabsorption
Explain why the differential permeability or impermeability of specific sections of the nephron tubules is necessary for urine formation
Describe how and where water, organic compounds, and ions are reabsorbed in the nephron
Explain the role of the loop of Henle, the vasa recta and the counter-current multiplication mechanisms in the concentration of urine
List the locations in the nephron where tubular secretion occurs
With up to 180 litres per day passing through the nephrons of the kidney, it is quite obvious that most of that fluid and its contents must be reabsorbed. That recovery occurs in the PCT, loop of Henle, DCT and the collecting ducts (Table 17.6.1 and Figure 17.6.1). Various portions of the nephron differ in their capacity to reabsorb water and specific solutes. While much of the reabsorption and secretion occur passively based on concentration gradients, the amount of water that is reabsorbed or lost is tightly regulated. This control is exerted directly by ADH and aldosterone and indirectly by renin. Most water is recovered in the PCT, loop of Henle and DCT. About 10 percent (about 18 L) reaches the collecting ducts. The collecting ducts, under the influence of ADH, can recover almost all of the water passing through them, in cases of dehydration, or almost none of the water, in cases of over-hydration.
Figure 17.6.1. Locations of secretion and reabsorption in the nephron.
Table 17.6.1. Substances secreted or reabsorbed in the nephron and their locations.
Substance
PCT
Loop of Henle
DCT
Collecting ducts
Glucose
Almost 100% reabsorbed; secondary active transport with Na+
Oligopeptides, proteins, amino acids
Almost 100% reabsorbed; symport with Na+
Vitamins
Reabsorbed
Lactate
Reabsorbed
Creatinine
Secreted
Urea
50% reabsorbed by diffusion; also secreted
Secretion, diffusion in descending limb
Reabsorption in medullary collecting ducts; diffusion
Sodium
65% actively reabsorbed
25% reabsorbed in thick ascending limb; active transport
5% reabsorbed; active
5% reabsorbed, stimulated by aldosterone; active
Chloride
Reabsorbed, symport with Na+, diffusion
Reabsorbed in thin and thick ascending limb; diffusion in ascending limb
Reabsorbed; diffusion
Reabsorbed; symport
Water
67% reabsorbed osmotically with solutes
15% reabsorbed in descending limb; osmosis
8% reabsorbed if ADH; osmosis
Variable amounts reabsorbed, controlled by ADH, osmosis
Bicarbonate
80-90% symport reabsorption with Na+
Reabsorbed, symport with Na+ and antiport with Cl-; in ascending limb
Reabsorbed antiport with Cl-
H+
Secreted; diffusion
Secreted; active
Secreted; active
NH4+
Secreted; diffusion
Secreted; diffusion
Secreted; diffusion
HCO3-
Reabsorbed; diffusion
Reabsorbed; diffusion in ascending limb
Reabsorbed; diffusion
Reabsorbed; antiport with Na+
Some drugs
Secreted
Secreted; active
Secreted; active
Potassium
65% reabsorbed; diffusion
20% reabsorbed in thick ascending limb; symport
Secreted; active
Secretion controlled by aldosterone; active
Calcium
Reabsorbed; diffusion
Reabsorbed in thick ascending limbs; diffusion
Reabsorbed if parathyroid hormone present; active
Magnesium
Reabsorbed; diffusion
Reabsorbed in thick ascending limb; diffusion
Reabsorbed
Phosphate
85% reabsorbed, inhibited by parathyroid hormone, diffusion
Reabsorbed; diffusion
Mechanisms of Recovery
Mechanisms by which substances move across membranes for reabsorption or secretion include active transport, diffusion, facilitated diffusion, secondary active transport and osmosis.
Active transport utilises energy, usually the energy found in a phosphate bond of ATP, to move a substance across a membrane from a low to a high concentration. It is very specific and must have an appropriately shaped receptor for the substance to be transported. An example would be the active transport of Na+ out of a cell and K+ into a cell by the Na+/K+ pump. Both ions are moved in opposite directions from a lower to a higher concentration.
Simple diffusion moves a substance from a higher to a lower concentration down its concentration gradient. It requires no energy and only needs to be soluble.
Facilitated diffusion is similar to diffusion in that it moves a substance down its concentration gradient. The difference is that it requires specific membrane receptors or channel proteins for movement. The movement of glucose and, in certain situations, Na+ ions, is an example of facilitated diffusion. In some cases of facilitated diffusion, two different substances share the same channel protein port; these mechanisms are described by the terms symport and antiport.
Symport mechanisms move two or more substances in the same direction at the same time, whereas antiport mechanisms move two or more substances in opposite directions across the cell membrane. Both mechanisms may utilise concentration gradients maintained by ATP pumps. This is a mechanism described by the term “secondary active transport.” For example, a Na+ ATPase pump on the basilar membrane of a cell may constantly pump Na+ out of a cell, maintaining a strong electrochemical gradient. On the opposite (apical) surface, a Na+/glucose symport protein channel assists both Na+ and glucose into the cell as Na+ moves down the concentration gradient created by the basilar Na+ ATPase pumps. The glucose molecule then diffuses across the basal membrane by facilitated diffusion into the interstitial space and from there into peritubular capillaries.
Most of the Ca2+, Na+, glucose, and amino acids must be reabsorbed by the nephron to maintain homeostatic plasma concentrations. Other substances, such as urea, K+, ammonia (NH3), creatinine, and some drugs are secreted into the filtrate as waste products. Acid–base balance is maintained through actions of the lungs and kidneys: The lungs rid the body of H+, whereas the kidneys secrete or reabsorb H+ and HCO3– (Table 17.6.2). In the case of urea, about 50 percent is passively reabsorbed by the PCT. More is recovered by in the collecting ducts as needed. ADH induces the insertion of urea transporters and aquaporin channel proteins.
Table 17.6.2. Substances filtered and reabsorbed by the kidney per 24 hours
Substance
Amount filtered (grams)
Amount reabsorbed (grams)
Amount in urine (grams)
Water
180 L
179 L
1 L
Proteins
10-20
10-20
0
Chlorine
630
625
5
Sodium
540
537
3
Bicarbonate
300
299.7
0.3
Glucose
180
180
0
Urea
53
28
25
Potassium
28
24
4
Uric acid
8.5
7.7
0.8
Creatinine
1.4
0
1.5
Reabsorption and Secretion in the PCT
The renal corpuscle filters the blood to create a filtrate that differs from blood mainly in the absence of cells and large proteins. From this point to the ends of the collecting ducts, the filtrate or forming urine is undergoing modification through secretion and reabsorption before true urine is produced. The first point at which the forming urine is modified is in the PCT. Here, some substances are reabsorbed, whereas others are secreted. Note the use of the term “reabsorbed.” All these substances were “absorbed” in the digestive tract—99 percent of the water and most of the solutes filtered by the nephron must be reabsorbed. Water and substances that are reabsorbed are returned to the circulation by the peritubular and vasa recta capillaries. It is important to understand the difference between the glomerulus and the peritubular and vasa recta capillaries. The glomerulus has a high pressure inside its capillaries and can sustain this by dilating the afferent arteriole while constricting the efferent arteriole. This assures adequate filtration pressure even as the systemic blood pressure varies. Movement of water into the peritubular capillaries and vasa recta will be influenced primarily by osmolarity and concentration gradients. Sodium is actively pumped out of the PCT into the interstitial spaces between cells and diffuses down its concentration gradient into the peritubular capillary. As it does so, water will follow passively to maintain an isotonic fluid environment inside the capillary. This is called obligatory water reabsorption because water is “obliged” to follow the Na+ (Figure 17.6.2).
Figure 17.6.2. Substances reabsorbed and secreted by the PCT.
More substances move across the membranes of the PCT than any other portion of the nephron. Many of these substances (amino acids and glucose) use symport mechanisms for transport along with Na+. Antiport, active transport, diffusion and facilitated diffusion are additional mechanisms by which substances are moved from one side of a membrane to the other. Recall that cells have two surfaces: apical and basal. The apical surface is the one facing the lumen or open space of a cavity or tube, in this case, the inside of the PCT. The basal surface of the cell faces the connective tissue base to which the cell attaches (basement membrane) or the cell membrane closer to the basement membrane if there is a stratified layer of cells. In the PCT, there is a single layer of simple cuboidal endothelial cells against the basement membrane. The numbers and particular types of pumps and channels vary between the apical and basilar surfaces (Table 17.6.3). A few of the substances that are transported with Na+ (symport mechanism) on the apical membrane include Cl–, Ca2+, amino acids, glucose and PO43−. Sodium is actively exchanged for K+ using ATP on the basal membrane. Most of the substances transported by a symport mechanism on the apical membrane are transported by facilitated diffusion on the basal membrane. At least three ions, K+, Ca2+ and Mg2+, diffuse laterally between adjacent cell membranes (transcellular movement).
Table 17.6.3. Reabsorption of major solutes by the PCT
Basal membrane
Apical membrane
Active transport
Symport with Na+
Na+ (exchange for K+)
K+
Facilitated diffusion
Cl–
K+
Ca2+
Cl–
Mg2+
Ca2+
HCO3–
HCO3–
PO43-
PO43-
Amino acids
Amino acids
Glucose
Glucose
Fructose
Fructose
Galactose
Lactate
Succinate
Succinate
Citrate
Citrate
Diffusion between nephron cells:
· K+
· Ca2+
· Mg2+
About 67 percent of the water, Na+ and K+ entering the nephron is reabsorbed in the PCT and returned to the circulation. Almost 100 percent of glucose, amino acids and other organic substances such as vitamins are normally recovered here. Some glucose may appear in the urine if circulating glucose levels are high enough that all the glucose transporters in the PCT are saturated, so that their capacity to move glucose is exceeded (transport maximum, or Tm). In men, the maximum amount of glucose that can be recovered is about 375 mg/min, whereas in women, it is about 300 mg/min. This recovery rate translates to an arterial concentration of about 200 mg/dL. Though an exceptionally high sugar intake might cause glucose to appear briefly in the urine, the appearance of glycosuria usually points to type I or II diabetes mellitus. The transport of glucose from the lumen of the PCT to the interstitial space is similar to the way it is absorbed by the small intestine. Both glucose and Na+ bind simultaneously to the same symport proteins on the apical surface of the cell to be transported in the same direction, toward the interstitial space. Sodium moves down its electrochemical and concentration gradient into the cell and takes glucose with it. Na+ is then actively pumped out of the cell at the basal surface of the cell into the interstitial space. Glucose leaves the cell to enter the interstitial space by facilitated diffusion. The energy to move glucose comes from the Na+/K+ ATPase that pumps Na+ out of the cell on the basal surface. Fifty percent of Cl– and variable quantities of Ca2+, Mg2+ and HPO42− are also recovered in the PCT.
Recovery of bicarbonate (HCO3–) is vital to the maintenance of acid–base balance, since it is a very powerful and fast-acting buffer. An important enzyme is used to catalyse this mechanism: carbonic anhydrase (CA). This same enzyme and reaction occurs in red blood cells in the transportation of CO2 in the stomach to produce hydrochloric acid and in the pancreas to produce HCO3– to buffer acidic chyme from the stomach. In the kidney, most of the CA is located within the cell, but a small amount is bound to the brush border of the membrane on the apical surface of the cell. In the lumen of the PCT, HCO3– combines with hydrogen ions to form carbonic acid (H2CO3). This is enzymatically catalysed into CO2 and water, which diffuse across the apical membrane into the cell. Water can move osmotically across the lipid bilayer membrane due to the presence of aquaporin water channels. Inside the cell, the reverse reaction occurs to produce bicarbonate ions (HCO3–). These bicarbonate ions are co-transported with Na+ across the basal membrane to the interstitial space around the PCT (Figure 16.7.3). At the same time this is occurring, a Na+/H+ antiporter excretes H+ into the lumen, while it recovers Na+. Note how the hydrogen ion is recycled so that bicarbonate can be recovered. Also, note that a Na+ gradient is created by the Na+/K+ pump.
HCO3−+ H+ ↔ H2CO3 ↔ CO2 + H2O
The significant recovery of solutes from the PCT lumen to the interstitial space creates an osmotic gradient that promotes water recovery. As noted before, water moves through channels created by the aquaporin proteins. These proteins are found in all cells in varying amounts and help regulate water movement across membranes and through cells by creating a passageway across the hydrophobic lipid bilayer membrane. Changing the number of aquaporin proteins in membranes of the collecting ducts also helps to regulate the osmolarity of the blood. The movement of many positively charged ions also creates an electrochemical gradient. This charge promotes the movement of negative ions toward the interstitial spaces and the movement of positive ions toward the lumen.
Figure 16.7.3. Reabsorption of bicarbonate (HCO3–) from the PCT.
Reabsorption and Secretion in the Loop of Henle
The loop of Henle consists of two sections: thick and thin descending and thin and thick ascending sections. The loops of cortical nephrons do not extend into the renal medulla very far, if at all. Juxtamedullary nephrons have loops that extend variable distances, some very deep into the medulla. The descending and ascending portions of the loop are highly specialised to enable recovery of much of the Na+ and water that were filtered by the glomerulus. As the forming urine moves through the loop, the osmolarity will change from isosmotic with blood (about 278–300 mOsmol/kg) to both a very hypertonic solution of about 1200 mOsmol/kg and a very hypotonic solution of about 100 mOsmol/kg. These changes are accomplished by osmosis in the descending limb and active transport in the ascending limb. Solutes and water recovered from these loops are returned to the circulation by way of the vasa recta.
Descending Loop
Most of the descending loop is comprised of simple squamous epithelial cells; to simplify the function of the loop, this discussion focuses on these cells. These membranes have permanent aquaporin channel proteins that allow unrestricted movement of water from the descending loop into the surrounding interstitium as osmolarity increases from about 300 mOsmol/kg to about 1200 mOsmol/kg. This increase results in reabsorption of up to 15 percent of the water entering the nephron. Modest amounts of urea, Na+ and other ions are also recovered here.
Most of the solutes that were filtered in the glomerulus have now been recovered along with the majority of water (about 82 percent). As the forming urine enters the ascending loop, major adjustments will be made to the concentration of solutes to create what is known as urine.
Ascending Loop
The ascending loop is made of very short thin and longer thick portions. Once again, to simplify the function, this section only considers the thick portion. The thick portion is lined with simple cuboidal epithelium without a brush border. It is completely impermeable to water due to the absence of aquaporin proteins, but ions, mainly Na+, are actively pumped out of the loop by large quantities of the Na+/K+ ATPase pump. This has two significant effects: Removal of Na+ while retaining water leads to a hypotonic filtrate by the time it reaches the DCT; pumping Na+ into the interstitial space contributes to the hyperosmotic environment in the kidney medulla.
The Na+/K+ ATPase pumps in the basal membrane create an electrochemical gradient, allowing reabsorption of Cl– by Na+/Cl– symporters in the apical membrane. At the same time that Na+ is actively pumped from the basal side of the cell into the interstitial fluid, Cl– follows the Na+ from the lumen into the interstitial fluid by a paracellular route between cells through leaky tight junctions. These are found between cells of the ascending loop, where they allow certain solutes to move according to their concentration gradient. Most of the K+ that enters the cell via symporters returns to the lumen (down its concentration gradient) through leaky channels in the apical membrane. Note the environment now created in the interstitial space: with the “back door exiting” K+, there is one Na+ and two Cl– ions left in the interstitium surrounding the ascending loop. Therefore, in comparison to the lumen of the loop, the interstitial space is now a negatively charged environment. This negative charge attracts cations (Na+, K+, Ca2+ and Mg 2+) from the lumen via a paracellular route to the interstitial space and vasa recta.
Counter-current Multiplier System
The structure of the loop of Henle and associated vasa recta create a counter current multiplier system (Figure 16.7.4). The counter current term comes from the fact that the descending and ascending loops are next to each other and their fluid flows in opposite directions (counter current). The multiplier term is due to the action of solute pumps that increase (multiply) the concentrations of urea and Na+ deep in the medulla.
Figure 16.7.4. Counter current multiplier system.
As discussed above, the ascending loop has many Na+ pumps that actively pump Na+ out of the forming urine into the interstitial spaces. In addition, collecting ducts have urea pumps that actively pump urea into the interstitial spaces. This results in the recovery of Na+ to the circulation via the vasa recta and creates a high osmolar environment in the depths of the medulla.
Ammonia (NH3) is a toxic by-product of protein metabolism. It is formed as amino acids are deaminated by liver hepatocytes. That means that the amine group, NH2, is removed from amino acids as they are broken down. Most of the resulting ammonia is converted into urea by liver hepatocytes. Urea is not only less toxic but is utilised to aid in the recovery of water by the loop of Henle and collecting ducts. While water is freely diffusing out of the descending loop through aquaporin channels into the interstitial spaces of the medulla, urea freely diffuses into the lumen of the descending loop as it descends deeper into the medulla, much of it to be reabsorbed from the forming urine when it reaches the collecting duct. Thus, the movement of Na+ and urea into the interstitial spaces by these mechanisms creates the hyperosmotic environment of the medulla. The net result of this counter current multiplier system is to recover both water and Na+ in the circulation.
The amino acid glutamine can be deaminated by the kidney. As NH2 from the amino acid is converted into NH3 and pumped into the lumen of the PCT, Na+ and HCO3– are excreted into the interstitial fluid of the renal pyramid via a symport mechanism. When this process occurs in the cells of the PCT, the added benefit is a net loss of a hydrogen ion (complexed to ammonia to form the weak acid NH4+) in the urine and a gain of a bicarbonate ion (HCO3–) in the blood. Ammonia and bicarbonate are exchanged in a one-to-one ratio. This exchange is yet another means by which the body can buffer and excrete acid. The presence of aquaporin channels in the descending loop allows prodigious quantities of water to leave the loop and enter the hyperosmolar interstitium of the pyramid, where it is returned to the circulation by the vasa recta. As the loop turns to become the ascending loop, there is an absence of aquaporin channels, so water cannot leave the loop. However, in the basal membrane of cells of the thick ascending loop, ATPase pumps actively remove Na+ from the cell. A Na+/K+/2Cl– symporter in the apical membrane passively allows these ions to enter the cell cytoplasm from the lumen of the loop down a concentration gradient created by the pump. This mechanism works to dilute the fluid of the ascending loop ultimately to approximately 50–100 mOsmol/L.
At the transition from the DCT to the collecting duct, about 20 percent of the original water is still present and about 10 percent of the sodium. If no other mechanism for water reabsorption existed, about 20–25 litres of urine would be produced. Now consider what is happening in the adjacent capillaries, the vasa recta. They are recovering both solutes and water at a rate that preserves the counter current multiplier system. In general, blood flows slowly in capillaries to allow time for exchange of nutrients and wastes. In the vasa recta particularly, this rate of flow is important for two additional reasons. The flow must be slow to allow blood cells to lose and regain water without either crenating or bursting. Second, a rapid flow would remove too much Na+ and urea, destroying the osmolar gradient that is necessary for the recovery of solutes and water. Thus, by flowing slowly to preserve the counter current mechanism, as the vasa recta descend, Na+ and urea are freely able to enter the capillary, while water freely leaves; as they ascend, Na+ and urea are secreted into the surrounding medulla, while water re-enters the vasa recta and re-enters the circulation.
Reabsorption and Secretion in the Distal Convoluted Tubule
Approximately 80 percent of filtered water has been recovered by the time the dilute forming urine enters the DCT. The DCT will recover another 10–15 percent before the forming urine enters the collecting ducts. Aldosterone increases the amount of Na+/K+ ATPase in the basal membrane of the DCT and collecting duct. The movement of Na+ out of the lumen of the collecting duct creates a negative charge that promotes the movement of Cl– out of the lumen into the interstitial space by a paracellular route across tight junctions. Peritubular capillaries receive the solutes and water, returning them to the circulation.
Cells of the DCT also recover Ca2+ from the filtrate. Receptors for parathyroid hormone (PTH) are found in DCT cells and when bound to PTH, induce the insertion of calcium channels on their luminal surface. The channels enhance Ca2+ recovery from the forming urine. In addition, as Na+ is pumped out of the cell, the resulting electrochemical gradient attracts Ca2+ into the cell. Finally, calcitriol (1,25 dihydroxyvitamin D, the active form of vitamin D) is especially important for calcium recovery. It induces the production of calcium-binding proteins that transport Ca2+ into the cell. These binding proteins are also important for the movement of calcium inside the cell and aid in exocytosis of calcium across the basolateral membrane. Any Ca2+ not reabsorbed at this point is lost in the urine.
Collecting Ducts and Recovery of Water
Solutes move across the membranes of the collecting ducts, which contain two distinct cell types, principal cells and intercalated cells. A principal cell possesses channels for the recovery or loss of sodium and potassium. An intercalated cell secretes or absorbs acid or bicarbonate. As in other portions of the nephron, there is an array of micromachines (pumps and channels) on display in the membranes of these cells.
Regulation of urine volume and osmolarity are major functions of the collecting ducts. By varying the amount of water that is recovered, the collecting ducts play a significant role in maintaining the body’s normal osmolarity. If the blood becomes hyperosmotic, the collecting ducts recover more water to dilute the blood; if the blood becomes hyposmotic, the collecting ducts recover less of the water, leading to concentration of the blood. Another way of saying this is: If plasma osmolarity rises, more water is recovered and urine volume decreases; if plasma osmolarity decreases, less water is recovered and urine volume increases. This function is regulated by the posterior pituitary hormone ADH (vasopressin). With mild dehydration, plasma osmolarity rises slightly. This increase is detected by osmoreceptors in the hypothalamus, which stimulates the release of ADH from the posterior pituitary. If plasma osmolarity decreases slightly, the opposite occurs.
When stimulated by ADH, aquaporin channels are inserted into the apical membrane of principal cells, which line the collecting ducts. As the ducts descend through the medulla, the osmolarity surrounding them increases (due to the counter current mechanisms described above). If aquaporin water channels are present, water will be osmotically pulled from the collecting duct into the surrounding interstitial space and into the peritubular capillaries. Therefore, the final urine will be more concentrated. If less ADH is secreted, fewer aquaporin channels are inserted and less water is recovered, resulting in dilute urine. By altering the number of aquaporin channels, the volume of water recovered or lost is altered. This, in turn, regulates the blood osmolarity, blood pressure, and osmolarity of the urine.
As Na+ is pumped from the forming urine, water is passively recaptured for the circulation; this preservation of vascular volume is critically important for the maintenance of a normal blood pressure. Aldosterone is secreted by the adrenal cortex in response to angiotensin II stimulation. As an extremely potent vasoconstrictor, angiotensin II functions immediately to increase blood pressure. By also stimulating aldosterone production, it provides a longer-lasting mechanism to support blood pressure by maintaining vascular volume (water recovery).
In addition to receptors for ADH, principal cells have receptors for the steroid hormone aldosterone. While ADH is primarily involved in the regulation of water recovery, aldosterone regulates Na+ recovery. Aldosterone stimulates principal cells to manufacture luminal Na+ and K+ channels as well as Na+/K+ ATPase pumps on the basal membrane of the cells. When aldosterone output increases, more Na+ is recovered from the forming urine and water follows the Na+ passively. As the pump recovers Na+ for the body, it is also pumping K+ into the forming urine since the pump moves K+ in the opposite direction. When aldosterone decreases, more Na+ remains in the forming urine and more K+ is recovered in the circulation. Symport channels move Na+ and Cl– together. Still other channels in the principal cells secrete K+ into the collecting duct in direct proportion to the recovery of Na+.
Intercalated cells play significant roles in regulating blood pH. Intercalated cells reabsorb K+ and HCO3– while secreting H+. This function lowers the acidity of the plasma while increasing the acidity of the urine.
Section Review
The kidney regulates water recovery and blood pressure by producing the enzyme renin. It is renin that starts a series of reactions, leading to the production of the vasoconstrictor angiotensin II and the salt-retaining steroid aldosterone. Water recovery is also powerfully and directly influenced by the hormone ADH. Even so, it only influences the last 10 percent of water available for recovery after filtration at the glomerulus, because 90 percent of water is recovered before reaching the collecting ducts. Depending on the body’s fluid status at any given time, the collecting ducts can recover none or almost all of the water reaching them.
Mechanisms of solute recovery include active transport, simple diffusion, and facilitated diffusion. Most filtered substances are reabsorbed. Urea, NH3, creatinine, and some drugs are filtered or secreted as wastes. H+ and HCO3– are secreted or reabsorbed as needed to maintain acid–base balance. Movement of water from the glomerulus is primarily due to pressure, whereas that of peritubular capillaries and vasa recta is due to osmolarity and concentration gradients. The PCT is the most metabolically active part of the nephron and uses a wide array of protein micromachines to maintain homeostasis—symporters, antiporters and ATPase active transporters—in conjunction with diffusion, both simple and facilitated. Almost 100 percent of glucose, amino acids and vitamins are recovered in the PCT. Bicarbonate (HCO3–) is recovered using the same enzyme, carbonic anhydrase (CA), found in erythrocytes. The recovery of solutes creates an osmotic gradient to promote the recovery of water. The descending loop of the juxtaglomerular nephrons reaches an osmolarity of up to 1200 mOsmol/kg, promoting the recovery of water. The ascending loop is impervious to water but actively recovers Na+, reducing filtrate osmolarity to 50–100 mOsmol/kg. The descending and ascending loop and vasa recta form a counter current multiplier system to increase Na+ concentration in the kidney medulla. The collecting ducts actively pump urea into the medulla, further contributing to the high osmotic environment. The vasa recta recover the solute and water in the medulla, returning them to the circulation. Nearly 90 percent of water is recovered before the forming urine reaches the DCT, which will recover another 10 percent. Calcium recovery in the DCT is influenced by PTH and active vitamin D. In the collecting ducts, ADH stimulates aquaporin channel insertion to increase water recovery and thereby regulate osmolarity of the blood. Aldosterone stimulates Na+ recovery by the collecting duct.
Describe the myogenic and tubuloglomerular feedback mechanisms and explain how they affect urine volume and composition
Describe the function of the juxtaglomerular apparatus (JGA)
It is vital that the flow of blood through the kidney be at a suitable rate to allow for filtration. This rate determines how much solute is retained or discarded, how much water is retained or discarded and the osmolarity of blood and the blood pressure of the body.
Sympathetic Nerves
The kidneys are innervated by the sympathetic neurons of the autonomic nervous system via the celiac plexus and splanchnic nerves. Reduction of sympathetic stimulation results in vasodilation and increased blood flow through the kidneys during resting conditions. When the frequency of action potentials increases, the arteriolar smooth muscle constricts (vasoconstriction), resulting in diminished glomerular flow, so less filtration occurs. Under conditions of stress, sympathetic nervous activity increases, resulting in the direct vasoconstriction of afferent arterioles (noradrenaline effect) as well as stimulation of the adrenal medulla. The adrenal medulla, in turn, produces a generalised vasoconstriction through the release of adrenaline. This includes vasoconstriction of the afferent arterioles, further reducing the volume of blood flowing through the kidneys. This process redirects blood to other organs with more immediate needs. If blood pressure falls, the sympathetic nerves will also stimulate the release of renin. Additional renin increases production of the powerful vasoconstrictor angiotensin II. Angiotensin II, as discussed above, will also stimulate aldosterone production to augment blood volume through retention of more Na+ and water. Only a 10 mm Hg pressure differential across the glomerulus is required for normal GFR, so very small changes in afferent arterial pressure significantly increase or decrease GFR.
Autoregulation
The kidneys are very effective at regulating the rate of blood flow over a wide range of blood pressures. Your blood pressure will decrease when you are relaxed or sleeping. It will increase when exercising. Yet, despite these changes, the filtration rate through the kidney will change very little. This is due to two internal autoregulatory mechanisms that operate without outside influence: the myogenic mechanism and the tubuloglomerular feedback mechanism.
Arteriole Myogenic Mechanism
The myogenic mechanism regulating blood flow within the kidney depends upon a characteristic shared by most smooth muscle cells of the body. When you stretch a smooth muscle cell, it contracts; when you stop, it relaxes, restoring its resting length. This mechanism works in the afferent arteriole that supplies the glomerulus. When blood pressure increases, smooth muscle cells in the wall of the arteriole are stretched and respond by contracting to resist the pressure, resulting in little change in flow. When blood pressure drops, the same smooth muscle cells relax to lower resistance, allowing a continued even flow of blood.
Tubuloglomerular Feedback
The tubuloglomerular feedback mechanism involves the JGA and a paracrine signalling mechanism utilising ATP, adenosine and nitric oxide (NO). This mechanism stimulates either contraction or relaxation of afferent arteriolar smooth muscle cells (Table 17.7.1). Recall that the DCT is in intimate contact with the afferent and efferent arterioles of the glomerulus. Specialised macula densa cells in this segment of the tubule respond to changes in the fluid flow rate and Na+ concentration. As GFR increases, there is less time for NaCl to be reabsorbed in the PCT, resulting in higher osmolarity in the filtrate. The increased fluid movement more strongly deflects single nonmotile cilia on macula densa cells. This increased osmolarity of the forming urine and the greater flow rate within the DCT, activates macula densa cells to respond by releasing ATP and adenosine (a metabolite of ATP). ATP and adenosine act locally as paracrine factors to stimulate the myogenic juxtaglomerular cells of the afferent arteriole to constrict, slowing blood flow and reducing GFR. Conversely, when GFR decreases, less Na+ is in the forming urine, and most will be reabsorbed before reaching the macula densa, which will result in decreased ATP and adenosine, allowing the afferent arteriole to dilate and increase GFR. NO has the opposite effect, relaxing the afferent arteriole at the same time ATP and adenosine are stimulating it to contract. Thus, NO fine-tunes the effects of adenosine and ATP on GFR.
ATP and adenosine increase, causing vasoconstriction
Vasoconstriction slows GFR
Decreased GFR
Tubular NaCl decreases
ATP and adenosine decrease, causing vasodilation
Vasodilation increases GFR
Increased GFR
Tubular NaCl increases
NO increases, causing vasodilation
Vasodilation increases GFR
Decreased GFR
Tubular NaCl decreases
NO decreases, causing vasoconstriction
Vasoconstriction decreases GFR
Section Review
The kidneys are innervated by sympathetic nerves of the autonomic nervous system. Sympathetic nervous activity decreases blood flow to the kidney, making more blood available to other areas of the body during times of stress. The arteriolar myogenic mechanism maintains a steady blood flow by causing arteriolar smooth muscle to contract when blood pressure increases and causing it to relax when blood pressure decreases. Tubuloglomerular feedback involves paracrine signalling at the JGA to cause vasoconstriction or vasodilation to maintain a steady rate of blood flow.
Describe how each of the following functions in the extrinsic control of GFR: renin–angiotensin mechanism, natriuretic peptides and sympathetic adrenergic activity
Describe how each of the following works to regulate reabsorption and secretion, to affect urine volume and composition: renin–angiotensin system, aldosterone, antidiuretic hormone and natriuretic peptides
Name and define the roles of other hormones that regulate kidney control
Several hormones have specific, important roles in regulating kidney function. They act to stimulate or inhibit blood flow. Some of these are endocrine, acting from a distance, whereas others are paracrine, acting locally.
Renin-Angiotensin-Aldosterone
Renin is an enzyme that is produced by the granular cells of the afferent arteriole at the JGA. It enzymatically converts angiotensinogen (made by the liver, freely circulating) into angiotensin I. Its release is stimulated by prostaglandins and NO from the JGA in response to decreased extracellular fluid volume.
Angiotensin Converting Enzyme (ACE) is not a hormone (it is an enzyme) but it is functionally important in regulating systemic blood pressure and kidney function. It is produced (mainly) in the lungs but binds to the surfaces of endothelial cells in the afferent arterioles and glomerulus. It enzymatically converts inactive angiotensin I into active angiotensin II. ACE is important in raising blood pressure. People with high blood pressure are sometimes prescribed ACE inhibitors to lower their blood pressure.
Angiotensin II is a potent vasoconstrictor that plays an immediate role in the regulation of blood pressure. It acts systemically to cause vasoconstriction as well as constriction of both the afferent and efferent arterioles of the glomerulus. In instances of blood loss or dehydration, it reduces both GFR and renal blood flow, thereby limiting fluid loss and preserving blood volume. Its release is usually stimulated by decreases in blood pressure and so the preservation of adequate blood pressure is its primary role.
Aldosterone, often called the “salt-retaining hormone,” is released from the adrenal cortex in response to angiotensin II or directly in response to increased plasma K+. It promotes Na+ reabsorption by the nephron, promoting the retention of water. It is also important in regulating K+, promoting its excretion. This dual effect on two minerals and its origin in the adrenal cortex explains its designation as a mineralocorticoid. As a result, renin has an immediate effect on blood pressure due to angiotensin II–stimulated vasoconstriction and a prolonged effect through Na+ recovery due to aldosterone. While aldosterone causes increased recovery of Na+, it also causes greater loss of K+. Progesterone is a steroid that is structurally like aldosterone. It binds to the aldosterone receptor and weakly stimulates Na+ reabsorption and increased water recovery. This process is unimportant in men due to low levels of circulating progesterone. It may cause increased retention of water during some periods of the menstrual cycle in women when progesterone concentrations increase.
Antidiuretic Hormone (ADH)
Diuretics are drugs that can increase water loss by interfering with the recapture of solutes and water from the forming urine. They are often prescribed to lower blood pressure. Coffee, tea and alcoholic beverages are familiar diuretics. ADH, a 9-amino acid peptide released by the posterior pituitary, works to do the exact opposite. It promotes the recovery of water, decreases urine volume, and maintains plasma osmolarity and blood pressure. It does so by stimulating the movement of aquaporin proteins into the apical cell membrane of principal cells of the collecting ducts to form water channels, allowing the transcellular movement of water from the lumen of the collecting duct into the interstitial space in the medulla of the kidney by osmosis. From there, it enters the vasa recta capillaries to return to the circulation. Water is attracted by the high osmotic environment of the deep kidney medulla.
Endothelin
Endothelins, 21-amino acid peptides, are extremely powerful vasoconstrictors. They are produced by endothelial cells of the renal blood vessels, mesangial cells and cells of the DCT. Hormones stimulating endothelin release include angiotensin II, bradykinin and adrenaline. They do not typically influence blood pressure in healthy people. However, in people with diabetic kidney disease, endothelin is chronically elevated, resulting in sodium retention. They also diminish GFR by damaging the podocytes and by potently vasoconstricting both the afferent and efferent arterioles.
Natriuretic Hormones
Natriuretic hormones are peptides that stimulate the kidneys to excrete sodium—an effect opposite that of aldosterone. Natriuretic hormones act by inhibiting aldosterone release and therefore inhibiting Na+ recovery in the collecting ducts. If Na+ remains in the forming urine, its osmotic force will cause a concurrent loss of water. Natriuretic hormones also inhibit ADH release, which of course will result in less water recovery. Therefore, natriuretic peptides inhibit both Na+ and water recovery. One example from this family of hormones is atrial natriuretic peptide (ANP), a 28-amino acid peptide produced by heart atria in response to over-stretching of the atrial wall. The over-stretching occurs in persons with elevated blood pressure or heart failure. It increases GFR through concurrent vasodilation of the afferent arteriole and vasoconstriction of the efferent arteriole. These events lead to an increased loss of water and sodium in the forming urine. It also decreases sodium reabsorption in the DCT. There is also B-type natriuretic peptide (BNP) of 32 amino acids produced in the ventricles of the heart. It has a 10-fold lower affinity for its receptor, so its effects are less than those of ANP. Its role may be to provide “fine tuning” for the regulation of blood pressure. BNP’s longer biologic half-life makes it a good diagnostic marker of congestive heart failure (Table 17.8.1).
Parathyroid Hormone
Parathyroid hormone (PTH) is an 84-amino acid peptide produced by the parathyroid glands in response to decreased circulating Ca2+ levels. Among its targets is the PCT, where it stimulates the hydroxylation of calcidiol to calcitriol (1,25-hydroxycholecalciferol, the active form of vitamin D). It also blocks reabsorption of phosphate (PO3–), causing its loss in the urine. The retention of phosphate would result in the formation of calcium phosphate in the plasma, reducing circulating Ca2+ levels. By ridding the blood of phosphate, higher circulating Ca2+ levels are permitted.
Table 17.8.1. Major hormones that influence Glomerular Filtration Rate (GFR) and Renal Blood Flow (RBF)
Stimulus
Effect on GFR
Effect on RBF
Vasoconstrictors
Sympathetic nerves (adrenaline and noradrenaline)
ECFV decreases
Decreases
Decreases
Angiotensin II
ECFV decreases
Decreases
Decreases
Endothelin
Starch, bradykinin, angiotensin II, and adrenaline increase; ECFV decrease
Decreases
Decreases
Vasodilators
Prostaglandins (PGE1, PGE2, and PGI2)
ECFV decreases; shear stress, and angiotensin II increases
No change/Increases
Increases
Nitric Oxide (NO)
Sheer stress, acetylcholine, histamine, bradykinin, ATP, and adenosine increases
Endocrine hormones act from a distance and paracrine hormones act locally. The renal enzyme renin converts angiotensinogen into angiotensin I. The lung enzyme, ACE, converts angiotensin I into active angiotensin II. Angiotensin II is an active vasoconstrictor that increases blood pressure. Angiotensin II also stimulates aldosterone release from the adrenal cortex, causing the collecting duct to retain Na+, which promotes water retention and a longer-term rise in blood pressure. ADH promotes water recovery by the collecting ducts by stimulating the insertion of aquaporin water channels into cell membranes. Endothelins are elevated in cases of diabetic kidney disease, increasing Na+ retention and decreasing GFR. Natriuretic hormones, released primarily from the atria of the heart in response to stretching of the atrial walls, stimulate Na+ excretion and thereby decrease blood pressure. PTH stimulates the last step in the formation of active vitamin D3 and reduces phosphate reabsorption, resulting in higher circulating Ca2+ levels.
Explain why the differential permeability or impermeability of specific sections of the nephron tubules is necessary for urine formation
The major hormones influencing total body water are ADH, aldosterone and ANP. Circumstances that lead to fluid depletion in the body include blood loss and dehydration. Homeostasis requires that volume and osmolarity be preserved. Blood volume is important in maintaining sufficient blood pressure, and there are non-renal mechanisms involved in its preservation, including vasoconstriction, which can act within seconds of a decrease in pressure. Thirst mechanisms are also activated to promote the consumption of water lost through respiration, evaporation or urination. Hormonal mechanisms are activated to recover volume while maintaining a normal osmotic environment. These mechanisms act principally on the kidney.
Volume-Sensing Mechanisms
The body cannot directly measure blood volume, but blood pressure can be measured. Blood pressure often reflects blood volume and is measured by baroreceptors in the aorta and carotid sinuses. When blood pressure increases, baroreceptors send more frequent action potentials to the central nervous system, leading to widespread vasodilation. Included in this vasodilation are the afferent arterioles supplying the glomerulus, resulting in increased GFR and water loss by the kidneys. If pressure decreases, fewer action potentials travel to the central nervous system, resulting in more sympathetic stimulation-producing vasoconstriction, which will result in decreased filtration and GFR and water loss.
Decreased blood pressure is also sensed by the granular cells in the afferent arteriole of the JGA. In response, the enzyme renin is released. You saw earlier in the chapter that renin activity leads to an almost immediate rise in blood pressure as activated angiotensin II produces vasoconstriction. The rise in pressure is sustained by the aldosterone effects initiated by angiotensin II; this includes an increase in Na+ retention and water volume. As an aside, late in the menstrual cycle, progesterone has a modest influence on water retention. Due to its structural similarity to aldosterone, progesterone binds to the aldosterone receptor in the collecting duct of the kidney, causing the same, albeit weaker, effect on Na+ and water retention.
Cardiomyocytes of the atria also respond to greater stretch (as blood pressure rises) by secreting ANP. ANP opposes the action of aldosterone by inhibiting the recovery of Na+ by the DCT and collecting ducts. More Na+ is lost, and as water follows salt, total blood volume and pressure decline. In low-pressure states, ANP does not seem to have much effect.
ADH is also called vasopressin. Early researchers found that in cases of unusually high secretion of ADH, the hormone caused vasoconstriction (vasopressor activity, hence the name). Only later were its antidiuretic properties identified. Synthetic ADH is still used occasionally to stem life-threatening oesophagus bleeding in alcoholics.
When blood volume drops 5–10 percent, causing a decrease in blood pressure, there is a rapid and significant increase in ADH release from the posterior pituitary. Immediate vasoconstriction to increase blood pressure is the result. ADH also causes activation of aquaporin channels in the collecting ducts to affect the recovery of water to help restore vascular volume.
Diuretics and Fluid Volume
A diuretic is a compound that increases urine volume. Three familiar drinks contain diuretic compounds: coffee, tea, and alcohol. The caffeine in coffee and tea works by promoting vasodilation in the nephron, which increases GFR. Alcohol increases GFR by inhibiting ADH release from the posterior pituitary, resulting in less water recovery by the collecting duct. In cases of high blood pressure, diuretics may be prescribed to reduce blood volume and, thereby, reduce blood pressure. The most frequently prescribed anti-hypertensive diuretic is hydrochlorothiazide. It inhibits the Na+/ Cl– symporter in the DCT and collecting duct. The result is a loss of Na+ with water following passively by osmosis.
Osmotic diuretics promote water loss by osmosis. An example is the indigestible sugar mannitol, which is most often administered to reduce brain swelling after head injury. However, it is not the only sugar that can produce a diuretic effect. In cases of poorly controlled diabetes mellitus, glucose levels exceed the capacity of the tubular glucose symporters, resulting in glucose in the urine. The unrecovered glucose becomes a powerful osmotic diuretic. Classically, in the days before glucose could be detected in the blood and urine, clinicians identified diabetes mellitus by the three Ps: polyuria (diuresis), polydipsia (increased thirst) and polyphagia (increased hunger).
Regulation of Extracellular Sodium Ions (Na+)
Sodium has a strong osmotic effect and attracts water. It plays a larger role in the osmolarity of the plasma than any other circulating component of the blood. If there is too much Na+ present, either due to poor control or excess dietary consumption, a series of metabolic problems ensue. There is an increase in total volume of water, which leads to hypertension (high blood pressure). Over a long period, this increases the risk of serious complications such as heart attacks, strokes and aneurysms. It can also contribute to system-wide oedema (swelling).
Mechanisms for regulating Na+ concentration include the renin–angiotensin–aldosterone system and ADH. Aldosterone stimulates the uptake of Na+ on the apical cell membrane of cells in the DCT and collecting ducts, whereas ADH helps to regulate Na+ concentration indirectly by regulating the reabsorption of water.
Regulation of Extracellular Potassium Ions (K+)
Potassium is present in a 30-fold greater concentration inside the cell than outside the cell. A generalisation can be made that K+ and Na+ concentrations will move in opposite directions. When more Na+ is reabsorbed, more K+ is secreted; when less Na+ is reabsorbed (leading to excretion by the kidney), more K+ is retained. When aldosterone causes a recovery of Na+ in the nephron, a negative electrical gradient is created that promotes the secretion of K+ and Cl– into the lumen.
Regulation of Chloride Ions (Cl–)
Chloride is important in acid–base balance in the extracellular space and has other functions, such as in the stomach, where it combines with hydrogen ions in the stomach lumen to form hydrochloric acid, aiding digestion. Its close association with Na+ in the extracellular environment makes it the dominant anion of this compartment and its regulation closely mirrors that of Na+.
Regulation of Ca2+ and Phosphate
The parathyroid glands monitor and respond to circulating levels of Ca2+ in the blood. When levels drop too low, PTH is released to stimulate the DCT to reabsorb Ca2+ from the forming urine. When levels are adequate or high, less PTH is released and more Ca2+ remains in the forming urine to be lost. Phosphate levels move in the opposite direction. When Ca2+ levels are low, PTH inhibits reabsorption of HPO42− so that its blood level drops, allowing Ca2+ levels to rise. PTH also stimulates the renal conversion of calcidiol into calcitriol, the active form of vitamin D. Calcitriol then stimulates the intestines to absorb more Ca2+ from the diet.
Regulation of H+, Bicarbonate and pH
The acid–base homeostasis of the body is a function of chemical buffers and physiologic buffering provided by the lungs and kidneys. Buffers, especially proteins, HCO32−, and ammonia have a very large capacity to absorb or release H+ as needed to resist a change in pH. They can act within fractions of a second. The lungs can rid the body of excess acid very rapidly (seconds to minutes) through the conversion of HCO3– into CO2, which is then exhaled. It is rapid but has limited capacity in the face of a significant acid challenge. The kidneys can rid the body of both acid and base. The renal capacity is large but slow (minutes to hours). The cells of the PCT actively secrete H+ into the forming urine as Na+ is reabsorbed. The body rids itself of excess H+ and raises blood pH. In the collecting ducts, the apical surfaces of intercalated cells have proton pumps that actively secrete H+ into the luminal, forming urine to remove it from the body.
As hydrogen ions are pumped into the forming urine, it is buffered by bicarbonate (HCO3–), H2PO4– (dihydrogen phosphate ion) or ammonia (forming NH4+, ammonium ion). Urine pH typically varies in a normal range from 4.5 to 8.0.
Regulation of Nitrogen Wastes
Nitrogen wastes are produced by the breakdown of proteins during normal metabolism. Proteins are broken down into amino acids, which in turn are deaminated by having their nitrogen groups removed. Deamination converts the amino (NH2) groups into ammonia (NH3), ammonium ion (NH4+), urea or uric acid (Figure 17.9.1). Ammonia is extremely toxic, so most of it is very rapidly converted into urea in the liver. Human urinary wastes typically contain primarily urea with small amounts of ammonium and very little uric acid.
Figure 17.9.1. Nitrogen wastes.
Elimination of Drugs and Hormones
Water-soluble drugs may be excreted in the urine and are influenced by one or all of the following processes: glomerular filtration, tubular secretion or tubular reabsorption. Drugs that are structurally small can be filtered by the glomerulus with the filtrate. Large drug molecules such as heparin or those that are bound to plasma proteins cannot be filtered and are not readily eliminated. Some drugs can be eliminated by carrier proteins that enable secretion of the drug into the tubule lumen. There are specific carriers that eliminate basic (such as dopamine or histamine) or acidic drugs (such as penicillin or indomethacin). As is the case with other substances, drugs may be both filtered and reabsorbed passively along a concentration gradient.
Section Review
The major hormones regulating body fluids are ADH, aldosterone and ANP. Progesterone is similar in structure to aldosterone and can bind to and weakly stimulate aldosterone receptors, providing a similar but diminished response. Blood pressure reflects blood volume and is monitored by baroreceptors in the aortic arch and carotid sinuses. When blood pressure increases, more action potentials are sent to the central nervous system, resulting in greater vasodilation, greater GFR, and more water lost in the urine. ANP is released by the cardiomyocytes when blood pressure increases, causing Na+ and water loss. ADH at high levels causes vasoconstriction in addition to its action on the collecting ducts to recover more water. Diuretics increase urine volume. Mechanisms for controlling Na+ concentration in the blood include the renin–angiotensin–aldosterone system and ADH. When Na+ is retained, K+ is excreted; when Na+ is lost, K+ is retained. When circulating Ca2+ decreases, PTH stimulates the reabsorption of Ca2+ and inhibits reabsorption of HPO42−. pH is regulated through buffers, expiration of CO2 and excretion of acid or base by the kidneys. The breakdown of amino acids produces ammonia. Most ammonia is converted into less-toxic urea in the liver and excreted in the urine. Regulation of drugs is by glomerular filtration, tubular secretion and tubular reabsorption.
Describe the role of the kidneys in vitamin D activation
Describe the role of the kidneys in regulating erythropoiesis
Provide specific examples to demonstrate how the urinary system responds to maintain homeostasis in the body
Explain how the urinary system relates to other body systems in maintaining homeostasis
Predict factors or situations affecting the urinary system that could disrupt homeostasis
Predict the types of problems that would occur in the body if the urinary system could not maintain homeostasis
All systems of the body are interrelated. A change in one system may affect all other systems in the body, with mild to devastating effects. A failure of urinary continence can be embarrassing and inconvenient but is not life threatening. The loss of other urinary functions may prove fatal. A failure to synthesise vitamin D is one such example.
Vitamin D Synthesis
In order for vitamin D to become active, it must undergo a hydroxylation reaction in the kidney, that is, an –OH group must be added to calcidiol to make calcitriol (1,25-dihydroxycholecalciferol). Activated vitamin D is important for absorption of Ca2+ in the digestive tract, its reabsorption in the kidney and the maintenance of normal serum concentrations of Ca2+ and phosphate. Calcium is vitally important in bone health, muscle contraction, hormone secretion, and neurotransmitter release. Inadequate Ca2+ leads to disorders like osteoporosis and osteomalacia in adults and rickets in children. Deficits may also result in problems with cell proliferation, neuromuscular function, blood clotting and the inflammatory response. Recent research has confirmed that vitamin D receptors are present in most, if not all, cells of the body, reflecting the systemic importance of vitamin D. Many scientists have suggested it be referred to as a hormone rather than a vitamin.
Erythropoiesis
EPO is a 193-amino acid protein that stimulates the formation of red blood cells in the bone marrow. The kidney produces 85 percent of circulating EPO; the liver, the remainder. If you move to a higher altitude, the partial pressure of oxygen is lower, meaning there is less pressure to push oxygen across the alveolar membrane and into the red blood cell. One way the body compensates is to manufacture more red blood cells by increasing EPO production. If you start an aerobic exercise program, your tissues will need more oxygen to cope and the kidney will respond with more EPO. If erythrocytes are lost due to severe or prolonged bleeding, or under produced due to disease or severe malnutrition, the kidneys come to the rescue by producing more EPO. Renal failure (loss of EPO production) is associated with anaemia, which makes it difficult for the body to cope with increased oxygen demands or to supply oxygen adequately even under normal conditions. Anaemia diminishes performance and can be life threatening.
Blood Pressure Regulation
Due to osmosis, water follows where Na+ leads. Much of the water the kidneys recover from the forming urine follows the reabsorption of Na+. ADH stimulation of aquaporin channels allows for regulation of water recovery in the collecting ducts. Normally, all the glucose is recovered, but loss of glucose control (diabetes mellitus) may result in an osmotic diuresis severe enough to produce severe dehydration and death. A loss of renal function means a loss of effective vascular volume control, leading to hypotension (low blood pressure) or hypertension (high blood pressure), which can lead to stroke, heart attack and aneurysm formation.
The kidneys cooperate with the lungs, liver and adrenal cortex through the renin–angiotensin–aldosterone system. The liver synthesises and secretes the inactive precursor angiotensinogen. When the blood pressure is low, the kidney synthesises and releases renin. Renin converts angiotensinogen into angiotensin I and ACE produced in the lung converts angiotensin I into biologically active angiotensin II (Figure 17.10.1). The immediate and short-term effect of angiotensin II is to raise blood pressure by causing widespread vasoconstriction. angiotensin II also stimulates the adrenal cortex to release the steroid hormone aldosterone, which results in renal reabsorption of Na+ and its associated osmotic recovery of water. The reabsorption of Na+ helps to raise and maintain blood pressure over a longer term.
Figure 17.10.1. The enzyme renin converts the pro-enzyme angiotensin.
Regulation of Osmolarity
Blood pressure and osmolarity are regulated in a similar fashion. Severe hypo-osmolarity can cause problems like lysis (rupture) of blood cells or widespread oedema, which is due to a solute imbalance. Inadequate solute concentration (such as protein) in the plasma results in water moving toward an area of greater solute concentration, in this case, the interstitial space and cell cytoplasm. If the kidney glomeruli are damaged by an autoimmune illness, large quantities of protein may be lost in the urine. The resultant drop in serum osmolarity leads to widespread oedema that, if severe, may lead to damaging or fatal brain swelling. Severe hypertonic conditions may arise with severe dehydration from lack of water intake, severe vomiting or uncontrolled diarrhoea. When the kidney is unable to recover sufficient water from the forming urine, the consequences may be severe (lethargy, confusion, muscle cramps and finally, death).
Recovery of Electrolytes
Sodium, calcium and potassium must be closely regulated. The role of Na+ and Ca2+ homeostasis has been discussed at length. Failure of K+ regulation can have serious consequences on nerve conduction, skeletal muscle function, and most significantly, on cardiac muscle contraction and rhythm.
pH Regulation
Recall that enzymes lose their three-dimensional conformation and therefore their function, if the pH is too acidic or basic. This loss of conformation may be a consequence of the breaking of hydrogen bonds. Move the pH away from the optimum for a specific enzyme and you may severely hamper its function throughout the body, including hormone binding, central nervous system signalling or myocardial contraction. Proper kidney function is essential for pH homeostasis.
Everyday Connection
Stem Cells and Repair of Kidney Damage
Stem cells are unspecialised cells that can reproduce themselves via cell division, sometimes after years of inactivity. Under certain conditions, they may differentiate into tissue-specific or organ-specific cells with special functions. In some cases, stem cells may continually divide to produce a mature cell and to replace themselves. Stem cell therapy has an enormous potential to improve the quality of life or save the lives of people suffering from debilitating or life-threatening diseases. There have been several studies in animals, but since stem cell therapy is still in its infancy, there have been limited experiments in humans.
Acute kidney injury can be caused by a number of factors, including transplants and other surgeries. It affects 7–10 percent of all hospitalised patients, resulting in the deaths of 35–40 percent of inpatients. In limited studies using mesenchymal stem cells, there have been fewer instances of kidney damage after surgery, the length of hospital stays has been reduced, and there have been fewer readmissions after release.
How do these stem cells work to protect or repair the kidney? Scientists are unsure at this point, but some evidence has shown that these stem cells release several growth factors in endocrine and paracrine ways. As further studies are conducted to assess the safety and effectiveness of stem cell therapy, we will move closer to a day when kidney injury is rare and curative treatments are routine.
Section Review
The effects of failure of parts of the urinary system may range from inconvenient (incontinence) to fatal (loss of filtration and many others). The kidneys catalyse the final reaction in the synthesis of active vitamin D that in turn helps regulate Ca2+. The kidney hormone EPO stimulates erythrocyte development and promotes adequate O2 transport. The kidneys help regulate blood pressure through Na+ and water retention and loss. The kidneys work with the adrenal cortex, lungs and liver in the renin–angiotensin–aldosterone system to regulate blood pressure. They regulate osmolarity of the blood by regulating both solutes and water. Three electrolytes are more closely regulated than others: Na+, Ca2+ and K+. The kidneys share pH regulation with the lungs and plasma buffers, so that proteins can preserve their three-dimensional conformation and thus their function.
Contrast the composition of the intracellular fluid with that of the extracellular fluid
Explain the importance of protein channels in the movement of solutes
Identify the causes and symptoms of oedema
The chemical reactions of life take place in aqueous solutions. The dissolved substances in a solution are called solutes. In the human body, solutes vary in different parts of the body, but may include proteins—including those that transport lipids, carbohydrates and, very importantly, electrolytes. Often in medicine, a mineral dissociated from a salt that carries an electrical charge (an ion) is called an electrolyte. For instance, sodium ions (Na+) and chloride ions (Cl–) are often referred to as electrolytes.
In the body, water moves through semi-permeable membranes of cells and from one compartment of the body to another by a process called osmosis. Osmosis is basically the diffusion of water from regions of higher concentration to regions of lower concentration, along an osmotic gradient across a semi-permeable membrane. As a result, water will move into and out of cells and tissues, depending on the relative concentrations of the water and solutes found there. An appropriate balance of solutes inside and outside of cells must be maintained to ensure normal function.
Body Water Content
Human beings are mostly water, ranging from about 75 percent of body mass in infants to about 50–60 percent in adult men and women, to as low as 45 percent in old age. The percent of body water changes with development, because the proportions of the body given over to each organ and to muscles, fat, bone and other tissues change from infancy to adulthood (Figure 17.11.1). The brain and kidneys have the highest proportions of water, which composes 80–85 percent of their masses. In contrast, teeth have the lowest proportion of water, at 8–10 percent.
Figure 17.11.1. Water content of the body’s organs and tissues. Water content varies in different body organs and tissues, from as little as 8 percent in the teeth to as much as 85 percent in the brain.
Fluid Compartments
Body fluids can be discussed in terms of their specific fluid compartment, a location that is largely separate from another compartment by some form of a physical barrier. The intracellular fluid (ICF) compartment is the system that includes all fluid enclosed in cells by their plasma membranes. Extracellular fluid (ECF) surrounds all cells in the body. Extracellular fluid has two primary constituents: the fluid component of the blood (called plasma) and the interstitial fluid (IF) that surrounds all cells not in the blood (Figure 17.11.2).
Figure 17.11.2. Fluid compartments in the human body. The intracellular fluid (ICF) is the fluid within cells. The interstitial fluid (IF) is part of the extracellular fluid (ECF) between the cells. Blood plasma is the second part of the ECF. Materials travel between cells and the plasma in capillaries through the IF.
Intracellular Fluid (ICF)
The ICF lies within cells and is the principal component of the cytosol/cytoplasm. The ICF makes up about 60 percent of the total water in the human body, and in an average-size adult male, the ICF accounts for about 25 litres of fluid (Figure 17.11.3). This fluid volume tends to be very stable, because the amount of water in living cells is closely regulated. If the volume of water inside a cell falls to a value that is too low, the cytosol becomes too concentrated with solutes to carry on normal cellular activities; if too much water enters a cell, the cell may burst (lyse) and be destroyed.
Figure 17.11.3. A pie graph showing the proportion of total body fluid in each of the body’s fluid compartments. Most of the water in the body is intracellular fluid. The second largest volume is the interstitial fluid, which surrounds cells that are not blood cells.
Extracellular Fluid (ECF)
The ECF accounts for the other one-third of the body’s water content. Approximately 20 percent of the ECF is found in plasma. Plasma travels through the body in blood vessels and transports a range of materials, including blood cells, proteins (including clotting factors and antibodies), electrolytes, nutrients, gases and wastes. Gases, nutrients, and waste materials travel between capillaries and cells through the IF. Cells are separated from the IF by a selectively permeable cell membrane that helps regulate the passage of materials between the IF and the interior of the cell.
The body has other water-based ECF. These include the cerebrospinal fluid that bathes the brain and spinal cord, lymph, the synovial fluid in joints, the pleural fluid in the pleural cavities, the pericardial fluid in the cardiac sac, the peritoneal fluid in the peritoneal cavity, and the aqueous humour of the eye. Because these fluids are outside of cells, these fluids are also considered components of the ECF compartment.
Composition of Body Fluids
The compositions of the two components of the ECF—plasma and IF—are more similar to each other than either is to the ICF (Figure 17.11.4). Blood plasma has high concentrations of sodium, chloride, bicarbonate, and protein. The IF has high concentrations of sodium, chloride, and bicarbonate, but a relatively lower concentration of protein. In contrast, the ICF has elevated amounts of potassium, phosphate, magnesium, and protein. Overall, the ICF contains high concentrations of potassium and phosphate (HPO42−), whereas both plasma and the ECF contain high concentrations of sodium and chloride.
Figure 17.11.4. The concentrations of different elements in key bodily fluids. The graph shows the composition of the ICF, IF, and plasma. The compositions of plasma and IF are similar to one another but are quite different from the composition of the ICF.
Most body fluids are neutral in charge. Thus, cations, or positively charged ions, and anions, or negatively charged ions, are balanced in fluids. As seen in the previous graph, sodium (Na+) ions and chloride (Cl–) ions are concentrated in the ECF of the body, whereas potassium (K+) ions are concentrated inside cells. Although sodium and potassium can “leak” through “pores” into and out of cells, respectively, the high levels of potassium and low levels of sodium in the ICF are maintained by sodium-potassium pumps in the cell membranes. These pumps use the energy supplied by ATP to pump sodium out of the cell and potassium into the cell (Figure 17.11.5).
Figure 17.11.5. The sodium-potassium pump. The sodium-potassium pump is powered by ATP to transfer sodium out of the cytoplasm and into the ECF. The pump also transfers potassium out of the ECF and into the cytoplasm. (credit: modification of work by Mariana Ruiz Villarreal).
Fluid Movement Between Compartments
Hydrostatic pressure, the force exerted by a fluid against a wall, causes movement of fluid between compartments. The hydrostatic pressure of blood is the pressure exerted by blood against the walls of the blood vessels by the pumping action of the heart. In capillaries, hydrostatic pressure (also known as capillary blood pressure) is higher than the opposing “colloid osmotic pressure” in blood—a “constant” pressure primarily produced by circulating albumin—at the arteriolar end of the capillary (Figure 17.11.6). This pressure forces plasma and nutrients out of the capillaries and into surrounding tissues. Fluid and the cellular wastes in the tissues enter the capillaries at the venule end, where the hydrostatic pressure is less than the osmotic pressure in the vessel. Filtration pressure squeezes fluid from the plasma in the blood to the IF surrounding the tissue cells. The surplus fluid in the interstitial space that is not returned directly back to the capillaries is drained from tissues by the lymphatic system, and then re-enters the vascular system at the subclavian veins.
Figure 17.11.6. Capillary exchange. Net filtration occurs near the arterial end of the capillary since capillary hydrostatic pressure (CHP) is greater than blood colloidal osmotic pressure (BCOP). There is no net movement of fluid near the midpoint of the capillary since CHP = BCOP. Net reabsorption occurs near the venous end of the capillary since BCOP is greater than CHP.
Hydrostatic pressure is especially important in governing the movement of water in the nephrons of the kidneys to ensure proper filtering of the blood to form urine. As hydrostatic pressure in the kidneys increases, the amount of water leaving the capillaries also increases, and more urine filtrate is formed. If hydrostatic pressure in the kidneys drops too low, as can happen in dehydration, the functions of the kidneys will be impaired, and less nitrogenous wastes will be removed from the bloodstream. Extreme dehydration can result in kidney failure.
Fluid also moves between compartments along an osmotic gradient. Recall that an osmotic gradient is produced by the difference in concentration of all solutes on either side of a semi-permeable membrane. The magnitude of the osmotic gradient is proportional to the difference in the concentration of solutes on one side of the cell membrane to that on the other side. Water will move by osmosis from the side where its concentration is high (and the concentration of solute is low) to the side of the membrane where its concentration is low (and the concentration of solute is high). In the body, water moves by osmosis from plasma to the IF (and the reverse) and from the IF to the ICF (and the reverse). In the body, water moves constantly into and out of fluid compartments as conditions change in different parts of the body.
If you are sweating, for example, you will lose water through your skin. Sweating depletes your tissues of water and increases the solute concentration in those tissues. As this happens, water diffuses from your blood into sweat glands and surrounding skin tissues that have become dehydrated because of the osmotic gradient. Additionally, as water leaves the blood, it is replaced by the water in other tissues throughout your body that are not dehydrated. If this continues, dehydration spreads throughout the body. When a dehydrated person drinks water and rehydrates, the water is redistributed by the same gradient, but in the opposite direction, replenishing water in all of the tissues.
Solute Movement Between Compartments
The movement of some solutes between compartments is active, which consumes energy and is an active transport process, whereas the movement of other solutes is passive, which does not require energy. Active transport allows cells to move a specific substance against its concentration gradient through a membrane protein, requiring energy in the form of ATP, for example, the sodium-potassium pump employs active transport to pump sodium out of cells and potassium into cells, with both substances moving against their concentration gradients.
Passive transport of a molecule or ion depends on its ability to pass through the membrane, as well as the existence of a concentration gradient that allows the molecules to diffuse from an area of higher concentration to an area of lower concentration. Some molecules, like gases, lipids, and water itself (which also utilises water channels in the membrane called aquaporins), slip fairly easily through the cell membrane; others, including polar molecules like glucose, amino acids, and ions do not. Some of these molecules enter and leave cells using facilitated transport, whereby the molecules move down a concentration gradient through specific protein channels in the membrane. This process does not require energy, for example, glucose is transferred into cells by glucose transporters that use facilitated transport (Figure 17.11.7).
Figure 17.11.7. Facilitated diffusion. Glucose molecules use facilitated diffusion to move down a concentration gradient through the carrier protein channels in the membrane. (credit: modification of work by Mariana Ruiz Villarreal).
Disorders of the Fluid Balance: Oedema
Figure 17.11.8. Oedema. An allergic reaction can cause capillaries in the hand to leak excess fluid that accumulates in the tissues. (credit: Jane Whitney).
Oedema is the accumulation of excess water in the tissues. It is most common in the soft tissues of the extremities. The physiological causes of oedema include water leakage from blood capillaries. Oedema is almost always caused by an underlying medical condition, by the use of certain therapeutic drugs, by pregnancy, by localised injury, or by an allergic reaction. In the limbs, the symptoms of oedema include swelling of the subcutaneous tissues, an increase in the normal size of the limb and stretched, tight skin. One quick way to check for subcutaneous oedema localised in a limb is to press a finger into the suspected area. Oedema is likely if the depression persists for several seconds after the finger is removed (which is called “pitting”).
Pulmonary oedema is excess fluid in the air sacs of the lungs, a common symptom of heart and/or kidney failure. People with pulmonary oedema likely will experience difficulty breathing and they may experience chest pain. Pulmonary oedema can be life threatening, because it compromises gas exchange in the lungs and anyone having symptoms should immediately seek medical care.
In pulmonary oedema resulting from heart failure, excessive leakage of water occurs because fluids get “backed up” in the pulmonary capillaries of the lungs when the left ventricle of the heart is unable to pump sufficient blood into the systemic circulation. Because the left side of the heart is unable to pump out its normal volume of blood, the blood in the pulmonary circulation gets “backed up,” starting with the left atrium, then into the pulmonary veins, and then into pulmonary capillaries. The resulting increased hydrostatic pressure within pulmonary capillaries, as blood is still coming in from the pulmonary arteries, causes fluid to be pushed out of them and into lung tissues.
Other causes of oedema include damage to blood vessels and/or lymphatic vessels or a decrease in osmotic pressure in chronic and severe liver disease, where the liver is unable to manufacture plasma proteins (Figure 17.11.8). A decrease in the normal levels of plasma proteins results in a decrease of colloid osmotic pressure (which counterbalances the hydrostatic pressure) in the capillaries. This process causes loss of water from the blood to the surrounding tissues, resulting in oedema.
Mild, transient oedema of the feet and legs may be caused by sitting or standing in the same position for long periods of time, as in the work of a supermarket cashier. This is because deep veins in the lower limbs rely on skeletal muscle contractions to push on the veins and thus “pump” blood back to the heart. Otherwise, the venous blood pools in the lower limbs and can leak into surrounding tissues.
Medications that can result in oedema include vasodilators, calcium channel blockers used to treat hypertension, non-steroidal anti-inflammatory drugs, oestrogen therapies and some diabetes medications. Underlying medical conditions that can contribute to oedema include congestive heart failure, kidney damage and kidney disease, disorders that affect the veins of the legs, and cirrhosis and other liver disorders.
Therapy for oedema usually focuses on elimination of the cause. Activities that can reduce the effects of the condition include appropriate exercises to keep the blood and lymph flowing through the affected areas. Other therapies include elevation of the affected part to assist drainage, massage and compression of the areas to move the fluid out of the tissues and decreased salt intake to decrease sodium and water retention.
Section Review
Your body is mostly water. Body fluids are aqueous solutions with differing concentrations of materials, called solutes. An appropriate balance of water and solute concentrations must be maintained to ensure cellular functions. If the cytosol becomes too concentrated due to water loss, cell functions deteriorate. If the cytosol becomes too dilute due to water intake by cells, cell membranes can be damaged and the cell can burst. Hydrostatic pressure is the force exerted by a fluid against a wall and causes movement of fluid between compartments. Fluid can also move between compartments along an osmotic gradient. Active transport processes require ATP to move some solutes against their concentration gradients between compartments. Passive transport of a molecule or ion depends on its ability to pass easily through the membrane, as well as the existence of a high to low concentration gradient.
Explain how water levels in the body influence the thirst cycle
Identify the main route by which water leaves the body
Describe the role of ADH and its effect on body water levels
Define dehydration and identify common causes of dehydration
On a typical day, the average adult will take in about 2500 mL of aqueous fluids. Although most of the intake comes through the digestive tract, about 230 mL per day is generated metabolically, in the last steps of aerobic respiration. Additionally, each day about the same volume (2500 mL) of water leaves the body by different routes; most of this lost water is removed as urine. The kidneys also can adjust blood volume though mechanisms that draw water out of the filtrate and urine. The kidneys can regulate water levels in the body; they conserve water if you are dehydrated and they can make urine more dilute to expel excess water if necessary. Water is lost through the skin through evaporation from the skin surface without overt sweating and from air expelled from the lungs. This type of water loss is called insensible water loss because a person is usually unaware of it.
Regulation of Water Intake
Osmolality is the ratio of solutes in a solution to a volume of solvent in a solution. Plasma osmolality is thus the ratio of solutes to water in blood plasma. A person’s plasma osmolality value reflects their state of hydration. A healthy body maintains plasma osmolality within a narrow range, by employing several mechanisms that regulate both water intake and output.
Drinking water is considered voluntary. So how is water intake regulated by the body? Consider someone who is experiencing dehydration, a net loss of water that results in insufficient water in blood and other tissues. The water that leaves the body, as exhaled air, sweat, or urine, is ultimately extracted from blood plasma. As the blood becomes more concentrated, the thirst response—a sequence of physiological processes—is triggered (Figure 17.12.1). Osmoreceptors are sensory receptors in the thirst centre in the hypothalamus that monitor the concentration of solutes (osmolality) of the blood. If blood osmolality increases above its ideal value, the hypothalamus transmits signals that result in a conscious awareness of thirst. The person should (and normally does) respond by drinking water. The hypothalamus of a dehydrated person also releases antidiuretic hormone (ADH) through the posterior pituitary gland. ADH signals the kidneys to recover water from urine, effectively diluting the blood plasma. To conserve water, the hypothalamus of a dehydrated person also sends signals via the sympathetic nervous system to the salivary glands in the mouth. The signals result in a decrease in watery, serous output (and an increase in stickier, thicker mucus output). These changes in secretions result in a “dry mouth” and the sensation of thirst.
Figure 17.12.1. A flowchart showing the thirst response. The thirst response begins when osmoreceptors detect a decrease in water levels in the blood.
Decreased blood volume resulting from water loss has two additional effects. First, baroreceptors, blood-pressure receptors in the arch of the aorta and the carotid arteries in the neck, detect a decrease in blood pressure that results from decreased blood volume. The heart is ultimately signalled to increase its rate and/or strength of contractions to compensate for the lowered blood pressure.
Second, the kidneys have a renin-angiotensin-aldosterone hormonal system that increases the production of the active form of the hormone angiotensin II, which helps stimulate thirst, but also stimulates the release of the hormone aldosterone from the adrenal glands. Aldosterone increases the reabsorption of sodium in the distal tubules of the nephrons in the kidneys and water follows this reabsorbed sodium back into the blood.
If adequate fluids are not consumed, dehydration results and a person’s body contains too little water to function correctly. A person who repeatedly vomits or who has diarrhoea may become dehydrated and infants, because their body mass is so low, can become dangerously dehydrated very quickly. Endurance athletes such as distance runners often become dehydrated during long races. Dehydration can be a medical emergency and a dehydrated person may lose consciousness, become comatose, or die, if their body is not rehydrated quickly.
Regulation of Water Output
Water loss from the body occurs predominantly through the renal system. A person produces an average of 1.5 litres of urine per day. Although the volume of urine varies in response to hydration levels, there is a minimum volume of urine production required for proper bodily functions. The kidney excretes 100 to 1200 milliosmoles of solutes per day to rid the body of a variety of excess salts and other water-soluble chemical wastes, most notably creatinine, urea and uric acid. Failure to produce the minimum volume of urine means that metabolic wastes cannot be effectively removed from the body, a situation that can impair organ function. The minimum level of urine production necessary to maintain normal function is about 0.47 litres per day.
The kidneys also must make adjustments in the event of ingestion of too much fluid. Diuresis, which is the production of urine in excess of normal levels, begins about 30 minutes after drinking a large quantity of fluid. Diuresis reaches a peak after about 1 hour and normal urine production is re-established after about 3 hours.
Role of Antidiuretic Hormone (ADH)
Antidiuretic hormone (ADH), also known as vasopressin, controls the volume of water reabsorbed from the collecting ducts and tubules in the kidney. This hormone is produced in the hypothalamus and is delivered to the posterior pituitary for storage and release (Figure 17.12.2). When the osmoreceptors in the hypothalamus detect an increase in the concentration of blood plasma, the hypothalamus signals the release of ADH from the posterior pituitary into the blood.
Figure 17.12.2. Antidiuretic hormone (ADH). ADH is produced in the hypothalamus and released by the posterior pituitary gland. It causes the kidneys to retain water, constricts arterioles in the peripheral circulation, and affects some social behaviours in mammals.
ADH has two major effects. It constricts the arterioles in the peripheral circulation, which reduces the flow of blood to the extremities and thereby increases the blood supply to the core of the body. ADH also causes the epithelial cells that line the renal collecting tubules to move water channel proteins, called aquaporins, from the interior of the cells to the apical surface, where these proteins are inserted into the cell membrane (Figure 17.12.3). The result is an increase in the water permeability of these cells and thus, a large increase in water passage from the urine through the walls of the collecting tubules, leading to more reabsorption of water into the bloodstream. When the blood plasma becomes less concentrated and the level of ADH decreases, aquaporins are removed from collecting tubule cell membranes, and the passage of water out of urine and into the blood decreases.
Figure 17.12.3. Aquaporins. The binding of ADH to receptors on the cells of the collecting tubule results in aquaporins being inserted into the plasma membrane, shown in the lower cell. This dramatically increases the flow of water out of the tubule and into the bloodstream.
A diuretic is a compound that increases urine output and therefore decreases water conservation by the body. Diuretics are used to treat hypertension, congestive heart failure, and fluid retention associated with menstruation. Alcohol acts as a diuretic by inhibiting the release of ADH. Additionally, caffeine, when consumed in high concentrations, also acts as a diuretic.
Section Review
Homeostasis requires that water intake and output be balanced. Most water intake comes through the digestive tract via liquids and food, but roughly 10 percent of water available to the body is generated at the end of aerobic respiration during cellular metabolism. Urine produced by the kidneys accounts for the largest volume of water leaving the body. The kidneys can adjust the concentration of the urine to reflect the body’s water needs, conserving water if the body is dehydrated or making urine more dilute to expel excess water when necessary. ADH is a hormone that helps the body to retain water by increasing water reabsorption by the kidneys.
List the role of the six most important electrolytes in the body
Name the disorders associated with abnormally high and low levels of the six electrolytes
Identify the predominant extracellular anion
Describe the role of aldosterone on the level of water in the body
The body contains a large variety of ions, or electrolytes, which perform a variety of functions. Some ions assist in the transmission of electrical impulses along cell membranes in neurons and muscles. Other ions help to stabilise protein structures in enzymes. Still others aid in releasing hormones from endocrine glands. All of the ions in plasma contribute to the osmotic balance that controls the movement of water between cells and their environment.
Electrolytes in living systems include sodium, potassium, chloride, bicarbonate, calcium, phosphate, magnesium, copper, zinc, iron, manganese, molybdenum, copper and chromium. In terms of body functioning, six electrolytes are most important: sodium, potassium, chloride, bicarbonate, calcium and phosphate.
Roles of Electrolytes (Ions)
These six ions aid in nerve excitability, endocrine secretion, membrane permeability, buffering body fluids and controlling the movement of fluids between compartments. These ions enter the body through the digestive tract. More than 90 percent of the calcium and phosphate that enters the body is incorporated into bones and teeth, with bone serving as a mineral reserve for these ions. In the event that calcium and phosphate are needed for other functions, bone tissue can be broken down to supply the blood and other tissues with these minerals. Phosphate is a normal constituent of nucleic acids; hence, blood levels of phosphate will increase whenever nucleic acids are broken down.
Excretion of ions occurs mainly through the kidneys, with lesser amounts lost in sweat and in faeces. Excessive sweating may cause a significant loss, especially of sodium and chloride. Severe vomiting or diarrhoea will cause a loss of chloride and bicarbonate ions. Adjustments in respiratory and renal functions allow the body to regulate the levels of these ions in the ECF.
Table 17.13.1 lists the reference values for blood plasma, cerebrospinal fluid (CSF) and urine for the six ions addressed in this section. In a clinical setting, sodium, potassium, and chloride are typically analysed in a routine urine sample. In contrast, calcium and phosphate analysis requires a collection of urine across a 24-hour period, because the output of these ions can vary over the course of a day. Urine values reflect the rates of excretion of these ions. Bicarbonate is the one ion that is not normally excreted in urine; instead, it is conserved by the kidneys for use in the body’s buffering systems.
Table 17.13.1. Electrolyte and iron reference values
Name
Chemical symbol
Plasma
CSF
Urine
Sodium
Na+
136.00-146.00 (mM)
138.00-150.00 (mM)
40.00-220.00 (mM)
Potassium
K+
3.50-5.00 (mM)
0.35-3.5 (mM)
25.00-125.00 (mM)
Chloride
Cl–
98.00-107.00 (mM)
118.00-132.00 (mM)
110.00-250.00 (mM)
Bicarbonate
HCO3–
22.00-29.00 (mM)
Calcium
Ca2+
2.15-2.55 (mmol/day)
Up to 7.49 (mmol/day)
Phosphate
HPO42-
0.81-1.45 (mmol/day)
12.90-42.00 (mmol/day)
Sodium (Na+)
Sodium is the major cation of the extracellular fluid. It is responsible for one-half of the osmotic pressure gradient that exists between the interior of cells and their surrounding environment. People eating a typical Western diet, which is very high in NaCl, routinely take in 130 to 160 mmol/day of sodium, however according to WHO, the daily recommendation of NaCl intake is less than 5 g (approximately 87 mmol/day). This excess sodium is a major factor in hypertension (high blood pressure) in some people. Excretion of sodium is accomplished primarily by the kidneys. Sodium is freely filtered through the glomerular capillaries of the kidneys and, although much of the filtered sodium is reabsorbed in the proximal convoluted tubule, some remains in the filtrate and urine and is normally excreted.
Hyponatraemia is a lower-than-normal concentration of sodium, usually associated with excess water accumulation in the body, which dilutes the sodium. An absolute loss of sodium may be due to a decreased intake of the ion coupled with its continual excretion in the urine. An abnormal loss of sodium from the body can result from several conditions, including excessive sweating, vomiting, or diarrhoea; the use of diuretics; excessive production of urine, which can occur in diabetes; and acidosis, either metabolic acidosis or diabetic ketoacidosis.
A relative decrease in blood sodium can occur because of an imbalance of sodium in one of the body’s other fluid compartments, like IF, or from a dilution of sodium due to water retention related to oedema or congestive heart failure. At the cellular level, hyponatraemia results in increased entry of water into cells by osmosis, because the concentration of solutes within the cell exceeds the concentration of solutes in the now-diluted ECF. The excess water causes swelling of the cells; the swelling of red blood cells—decreasing their oxygen-carrying efficiency and making them potentially too large to fit through capillaries—along with the swelling of neurons in the brain can result in brain damage or even death.
Hypernatremia is an abnormal increase of blood sodium. It can result from water loss from the blood, resulting in the haemoconcentration of all blood constituents. Hormonal imbalances involving ADH and aldosterone may also result in higher-than-normal sodium values.
Potassium (K+)
Potassium is the major intracellular cation. It helps establish the resting membrane potential in neurons and muscle fibres after membrane depolarisation and action potentials. In contrast to sodium, potassium has little effect on osmotic pressure. The low levels of potassium in blood and CSF are due to the sodium-potassium pumps in cell membranes, which maintain the normal potassium concentration gradients between the ICF and ECF. The recommendation for daily intake/consumption of potassium is at least 90 mmol/day for adults, according to WHO. Potassium is excreted, both actively and passively, through the renal tubules, especially the distal convoluted tubule and collecting ducts. Potassium participates in the exchange with sodium in the renal tubules under the influence of aldosterone, which also relies on basolateral sodium-potassium pumps.
Hypokalaemia is an abnormally low potassium blood level. Similar to the situation with hyponatraemia, Hypokalaemia can occur because of either an absolute reduction of potassium in the body or a relative reduction of potassium in the blood due to the redistribution of potassium. An absolute loss of potassium can arise from decreased intake, frequently related to starvation. It can also result from vomiting, diarrhoea, or alkalosis.
Some insulin-dependent diabetic patients experience a relative reduction of potassium in the blood from the redistribution of potassium. When insulin is administered and glucose is taken up by cells, potassium passes through the cell membrane along with glucose, decreasing the amount of potassium in the blood and IF, which can cause hyperpolarisation of the cell membranes of neurons, reducing their responses to stimuli.
Hyperkalaemia, an elevated potassium blood level, also can impair the function of skeletal muscles, the nervous system, and the heart. Hyperkalaemia can result from increased dietary intake of potassium. In such a situation, potassium from the blood ends up in the ECF in abnormally high concentrations. This can result in a partial depolarisation (excitation) of the plasma membrane of skeletal muscle fibres, neurons, and cardiac cells of the heart, and can also lead to an inability of cells to repolarise. For the heart, this means that it won’t relax after a contraction, and will effectively “seize” and stop pumping blood, which is fatal within minutes. Because of such effects on the nervous system, a person with Hyperkalaemia may also exhibit mental confusion, numbness, and weakened respiratory muscles.
Chloride (Cl–)
Chloride is the predominant extracellular anion. Chloride is a major contributor to the osmotic pressure gradient between the ICF and ECF and plays an important role in maintaining proper hydration. Chloride functions to balance cations in the ECF, maintaining the electrical neutrality of this fluid. The paths of secretion and reabsorption of chloride ions in the renal system follow the paths of sodium ions.
Hypochloraemia, or lower-than-normal blood chloride levels, can occur because of defective renal tubular absorption. Vomiting, diarrhoea and metabolic acidosis can also lead to hypochloraemia. Hyperchloraemia, or higher-than-normal blood chloride levels, can occur due to dehydration, excessive intake of dietary salt (NaCl) or swallowing of sea water, aspirin intoxication, congestive heart failure and the hereditary, chronic lung disease, cystic fibrosis. In people who have cystic fibrosis, chloride levels in sweat are two to five times those of normal levels and analysis of sweat is often used in the diagnosis of the disease.
Bicarbonate (HCO3–)
Bicarbonate is the second most abundant anion in the blood. Its principal function is to maintain your body’s acid-base balance by being part of buffer systems. This role will be discussed in a different section.
Bicarbonate ions result from a chemical reaction that starts with carbon dioxide (CO2) and water, two molecules that are produced at the end of aerobic metabolism. Only a small amount of CO2 can be dissolved in body fluids. Thus, over 90 percent of the CO2 is converted into bicarbonate ions, HCO3–, through the following reactions:
CO2 + H2O ↔ H2CO3 ↔ HCO3– + H+
The bidirectional arrows indicate that the reactions can go in either direction, depending on the concentrations of the reactants and products. Carbon dioxide is produced in large amounts in tissues that have a high metabolic rate. Carbon dioxide is converted into bicarbonate in the cytoplasm of red blood cells through the action of an enzyme called carbonic anhydrase. Bicarbonate is transported in the blood. Once in the lungs, the reactions reverse direction and CO2 is regenerated from bicarbonate to be exhaled as metabolic waste.
Calcium (Ca2+)
It has been estimated that in adults, roughly one kilogram of calcium is found as calcium hydroxyapatite (Ca10[PO4]6[OH]2) inside bones and teeth, which provides hardness to the bone and serves as a mineral reserve for calcium and its salts for the rest of the tissues. Teeth also have a high concentration of calcium within them. A little more than one-half of blood calcium is bound to proteins, leaving the rest in its ionised form. Calcium ions, Ca2+, are necessary for muscle contraction, enzyme activity and blood coagulation. In addition, calcium helps to stabilise cell membranes and is essential for the release of neurotransmitters from neurons and of hormones from endocrine glands.
Calcium is absorbed through the intestines under the influence of activated vitamin D. A deficiency of vitamin D leads to a decrease in absorbed calcium and, eventually, a depletion of calcium stores from the skeletal system, potentially leading to rickets in children and osteomalacia in adults, contributing to osteoporosis.
Hypocalcaemia, or abnormally low calcium blood levels, is seen in hypoparathyroidism, which may follow the removal of the thyroid gland, because the four nodules of the parathyroid gland are embedded in it. Hypercalcemia, or abnormally high calcium blood levels, is seen in primary hyperparathyroidism. Some malignancies may also result in hypercalcemia.
Phosphate ([H2PO4]−, [HPO4]2- and [PO4]3−)
Phosphate is present in the body in three ionic forms: [H2PO4]−, [HPO4]2- and [PO4]3−. The most common form is [HPO4]2−. Bone and teeth bind up 85 percent of the body’s phosphate as part of calcium-phosphate salts. Phosphate is found in phospholipids, such as those that make up the cell membrane, and in ATP, nucleotides and buffers.
Hypophosphataemia, or abnormally low phosphate blood levels, occurs with heavy use of antacids, during alcohol withdrawal and during malnourishment. In the face of phosphate depletion, the kidneys usually conserve phosphate, but during starvation, this conservation is impaired greatly. Hyperphosphataemia, or abnormally increased levels of phosphates in the blood, occurs if there is decreased renal function or in cases of acute lymphocytic leukaemia. Additionally, because phosphate is a major constituent of the ICF, any significant destruction of cells can result in dumping of phosphate into the ECF.
Regulation of Sodium and Potassium
Sodium is reabsorbed from the renal filtrate, and potassium is excreted into the filtrate in the renal collecting tubule. The control of this exchange is governed principally by two hormones—aldosterone and angiotensin II.
Aldosterone
Recall that aldosterone increases the excretion of potassium and the reabsorption of sodium in the distal tubule. Aldosterone is released if blood levels of potassium increase, if blood levels of sodium severely decrease, or if blood pressure decreases. Its net effect is to conserve and increase water levels in the plasma by reducing the excretion of sodium and thus water, from the kidneys. In a negative feedback loop, increased osmolality of the ECF (which follows aldosterone-stimulated sodium absorption) inhibits the release of the hormone (Figure 17.13.1).
Figure 17.13.1. The aldosterone feedback loop. Aldosterone, which is released by the adrenal gland, facilitates reabsorption of Na+ and thus the reabsorption of water.
Angiotensin II
Angiotensin II causes vasoconstriction and an increase in systemic blood pressure. This action increases the glomerular filtration rate, resulting in more material filtered out of the glomerular capillaries and into Bowman’s capsule. Angiotensin II also signals an increase in the release of aldosterone from the adrenal cortex.
In the distal convoluted tubules and collecting ducts of the kidneys, aldosterone stimulates the synthesis and activation of the sodium-potassium pump (Figure 17.13.2). Sodium passes from the filtrate, into and through the cells of the tubules and ducts, into the ECF and then into capillaries. Water follows the sodium due to osmosis. Thus, aldosterone causes an increase in blood sodium levels and blood volume. Aldosterone’s effect on potassium is the reverse of that of sodium; under its influence, excess potassium is pumped into the renal filtrate for excretion from the body.
Figure 17.13.2. The renin-angiotensin-aldosterone system (RAAS). Angiotensin II stimulates the release of aldosterone from the adrenal cortex.
Regulation of Calcium and Phosphate
Calcium and phosphate are both regulated through the actions of three hormones: parathyroid hormone (PTH), dihydroxyvitamin D (calcitriol) and calcitonin. All three are released or synthesised in response to the blood levels of calcium.
PTH is released from the parathyroid gland in response to a decrease in the concentration of blood calcium. The hormone activates osteoclasts to break down bone matrix and release inorganic calcium-phosphate salts. PTH also increases the gastrointestinal absorption of dietary calcium by converting vitamin D into dihydroxyvitamin D (calcitriol), an active form of vitamin D that intestinal epithelial cells require to absorb calcium.
PTH raises blood calcium levels by inhibiting the loss of calcium through the kidneys. PTH also increases the loss of phosphate through the kidneys.
Calcitonin is released from the thyroid gland in response to elevated blood levels of calcium. The hormone increases the activity of osteoblasts, which remove calcium from the blood and incorporate calcium into the bony matrix.
Section Review
Electrolytes serve various purposes, such as helping to conduct electrical impulses along cell membranes in neurons and muscles, stabilising enzyme structures, and releasing hormones from endocrine glands. The ions in plasma also contribute to the osmotic balance that controls the movement of water between cells and their environment. Imbalances of these ions can result in various problems in the body and their concentrations are tightly regulated. Aldosterone and angiotensin II control the exchange of sodium and potassium between the renal filtrate and the renal collecting tubule. Calcium and phosphate are regulated by PTH, calcitriol and calcitonin.
Identify the most powerful buffer system in the body
Explain the way in which the respiratory system affects blood pH
Proper physiological functioning depends on a very tight balance between the concentrations of acids and bases in the blood. Acid-balance balance is measured using the pH scale, as shown in Figure 17.14.1. A variety of buffering systems permits blood and other bodily fluids to maintain a narrow pH range, even in the face of perturbations. A buffer is a chemical system that prevents a radical change in fluid pH by dampening the change in hydrogen ion concentrations in the case of excess acid or base. Most commonly, the substance that absorbs the ions is either a weak acid, which takes up hydroxyl ions, or a weak base which takes up hydrogen ions.
Figure 17.14.1. The pH scale. This chart shows where many common substances fall on the pH scale.
Buffer Systems in the Body
The buffer systems in the human body are extremely efficient and different systems work at different rates. It takes only seconds for the chemical buffers in the blood to make adjustments to pH. The respiratory tract can adjust the blood pH upward in minutes by exhaling CO2 from the body. The renal system can also adjust blood pH through the excretion of hydrogen ions (H+) and the conservation of bicarbonate, but this process takes hours to days to have an effect.
The buffer systems functioning in blood plasma include plasma proteins, phosphate and bicarbonate and carbonic acid buffers. The kidneys help control acid-base balance by excreting hydrogen ions and generating bicarbonate that helps maintain blood plasma pH within a normal range. Protein buffer systems work predominantly inside cells.
Protein Buffers in Blood Plasma and Cells
Nearly all proteins can function as buffers. Proteins are made up of amino acids, which contain positively charged amino groups and negatively charged carboxyl groups. The charged regions of these molecules can bind hydrogen and hydroxyl ions, and thus function as buffers. Buffering by proteins accounts for two-thirds of the buffering power of the blood and most of the buffering within cells.
Haemoglobin as a Buffer
Haemoglobin is the principal protein inside of red blood cells and accounts for one-third of the mass of the cell. During the conversion of CO2 into bicarbonate, hydrogen ions liberated in the reaction are buffered by haemoglobin, which is reduced by the dissociation of oxygen. This buffering helps maintain normal pH. The process is reversed in the pulmonary capillaries to re-form CO2, which then can diffuse into the air sacs to be exhaled into the atmosphere. This process is discussed in detail in the chapter on the respiratory system.
Phosphate Buffer
Phosphates are found in the blood in two forms: sodium dihydrogen phosphate (Na2H2PO4−), which is a weak acid, and sodium monohydrogen phosphate (Na2HPO42-), which is a weak base. When Na2HPO42- comes into contact with a strong acid, such as HCl, the base picks up a second hydrogen ion to form the weak acid Na2H2PO4− and sodium chloride, NaCl. When Na2HPO42− (the weak acid) comes into contact with a strong base, such as sodium hydroxide (NaOH), the weak acid reverts back to the weak base and produces water. Acids and bases are still present, but they hold onto the ions.
The bicarbonate-carbonic acid buffer works in a fashion similar to phosphate buffers. The bicarbonate is regulated in the blood by sodium, as are the phosphate ions. When sodium bicarbonate (NaHCO3), comes into contact with a strong acid, such as HCl, carbonic acid (H2CO3), which is a weak acid, and NaCl are formed. When carbonic acid comes into contact with a strong base, such as NaOH, bicarbonate and water are formed.
As with the phosphate buffer, a weak acid or weak base captures the free ions, and a significant change in pH is prevented. Bicarbonate ions and carbonic acid are present in the blood in a 20:1 ratio if the blood pH is within the normal range. With 20 times more bicarbonate than carbonic acid, this capture system is most efficient at buffering changes that would make the blood more acidic. This is useful because most of the body’s metabolic wastes, such as lactic acid and ketones, are acids. Carbonic acid levels in the blood are controlled by the expiration of CO2 through the lungs. In red blood cells, carbonic anhydrase forces the dissociation of the acid, rendering the blood less acidic. Because of this acid dissociation, CO2 is exhaled (see equations above). The level of bicarbonate in the blood is controlled through the renal system, where bicarbonate ions in the renal filtrate are conserved and passed back into the blood. However, the bicarbonate buffer is the primary buffering system of the IF surrounding the cells in tissues throughout the body.
Respiratory Regulation of Acid-Base Balance
The respiratory system contributes to the balance of acids and bases in the body by regulating the blood levels of carbonic acid (Figure 17.14.2). CO2 in the blood readily reacts with water to form carbonic acid, and the levels of CO2 and carbonic acid in the blood are in equilibrium. When the CO2 level in the blood rises (as it does when you hold your breath), the excess CO2 reacts with water to form additional carbonic acid, lowering blood pH. Increasing the rate and/or depth of respiration (which you might feel the “urge” to do after holding your breath) allows you to exhale more CO2. The loss of CO2 from the body reduces blood levels of carbonic acid and thereby adjusts the pH upward, toward normal levels. As you might have surmised, this process also works in the opposite direction. Excessive deep and rapid breathing (as in hyperventilation) rids the blood of CO2 and reduces the level of carbonic acid, making the blood too alkaline. This brief alkalosis can be remedied by rebreathing air that has been exhaled into a paper bag. Rebreathing exhaled air will rapidly bring blood pH down toward normal.
Figure 17.14.2. Respiratory regulation of blood pH. The respiratory system can reduce blood pH by removing CO2 from the blood.
The chemical reactions that regulate the levels of CO2 and carbonic acid occur in the lungs when blood travels through the lung’s pulmonary capillaries. Minor adjustments in breathing are usually sufficient to adjust the pH of the blood by changing how much CO2 is exhaled. In fact, doubling the respiratory rate for less than 1 minute, removing “extra” CO2, would increase the blood pH by 0.2. This situation is common if you are exercising strenuously over a period of time. To keep up the necessary energy production, you would produce excess CO2 (and lactic acid if exercising beyond your aerobic threshold). In order to balance the increased acid production, the respiration rate goes up to remove the CO2. This helps to keep you from developing acidosis.
The body regulates the respiratory rate by the use of chemoreceptors, which primarily use CO2 as a signal. Peripheral blood sensors are found in the walls of the aorta and carotid arteries. These sensors signal the brain to provide immediate adjustments to the respiratory rate if CO2 levels rise or fall. Yet other sensors are found in the brain itself. Changes in the pH of CSF affect the respiratory centre in the medulla oblongata, which can directly modulate breathing rate to bring the pH back into the normal range.
Hypercapnia, or abnormally elevated blood levels of CO2, occurs in any situation that impairs respiratory functions, including pneumonia and congestive heart failure. Reduced breathing (hypoventilation) due to drugs such as morphine, barbiturates or ethanol (or even just holding one’s breath) can also result in hypercapnia. Hypocapnia, or abnormally low blood levels of CO2, occurs with any cause of hyperventilation that drives off the CO2, such as salicylate toxicity, elevated room temperatures, fever or hysteria.
Renal Regulation of Acid-Base Balance
The renal regulation of the body’s acid-base balance addresses the metabolic component of the buffering system. Whereas the respiratory system (together with breathing centres in the brain) controls the blood levels of carbonic acid by controlling the exhalation of CO2, the renal system controls the blood levels of bicarbonate. A decrease of blood bicarbonate can result from the inhibition of carbonic anhydrase by certain diuretics or from excessive bicarbonate loss due to diarrhoea. Blood bicarbonate levels are also typically lower in people who have Addison’s disease (chronic adrenal insufficiency), in which aldosterone levels are reduced, and in people who have renal damage, such as chronic nephritis. Finally, low bicarbonate blood levels can result from elevated levels of ketones (common in unmanaged diabetes mellitus), which bind bicarbonate in the filtrate and prevent its conservation.
Bicarbonate ions, HCO3–, found in the filtrate, are essential to the bicarbonate buffer system, yet the cells of the tubule are not permeable to bicarbonate ions. The steps involved in supplying bicarbonate ions to the system are seen in Figure 17.14.3 and are summarised below:
Step 1: Sodium ions are reabsorbed from the filtrate in exchange for H+ by an antiport mechanism in the apical membranes of cells lining the renal tubule.
Step 2: The cells produce bicarbonate ions that can be shunted to peritubular capillaries.
Step 3: When CO2 is available, the reaction is driven to the formation of carbonic acid, which dissociates to form a bicarbonate ion and a hydrogen ion.
Step 4: The bicarbonate ion passes into the peritubular capillaries and returns to the blood. The hydrogen ion is secreted into the filtrate, where it can become part of new water molecules and be reabsorbed as such or removed in the urine.
Figure 17.14.3. Conservation of bicarbonate in the kidney. Tubular cells are not permeable to bicarbonate; thus, bicarbonate is conserved rather than reabsorbed. Steps 1 and 2 of bicarbonate conservation are indicated.
It is also possible that salts in the filtrate, such as sulfates, phosphates, or ammonia, will capture hydrogen ions. If this occurs, the hydrogen ions will not be available to combine with bicarbonate ions and produce CO2. In such cases, bicarbonate ions are not conserved from the filtrate to the blood, which will also contribute to a pH imbalance and acidosis.
The hydrogen ions also compete with potassium to exchange with sodium in the renal tubules. If more potassium is present than normal, potassium, rather than the hydrogen ions, will be exchanged, and increased potassium enters the filtrate. When this occurs, fewer hydrogen ions in the filtrate participate in the conversion of bicarbonate into CO2 and less bicarbonate is conserved. If there is less potassium, more hydrogen ions enter the filtrate to be exchanged with sodium and more bicarbonate is conserved.
Chloride ions are important in neutralising positive ion charges in the body. If chloride is lost, the body uses bicarbonate ions in place of the lost chloride ions. Thus, lost chloride results in an increased reabsorption of bicarbonate by the renal system.
Disorders of the Acid-Base Balance: Ketoacidosis
Diabetic acidosis, or ketoacidosis, occurs most frequently in people with poorly controlled diabetes mellitus. When certain tissues in the body cannot get adequate amounts of glucose, they depend on the breakdown of fatty acids for energy. When acetyl groups break off the fatty acid chains, the acetyl groups then non-enzymatically combine to form ketone bodies, acetoacetic acid, beta-hydroxybutyric acid and acetone, all of which increase the acidity of the blood. In this condition, the brain isn’t supplied with enough of its fuel—glucose—to produce all of the ATP it requires to function.
Ketoacidosis can be severe and, if not detected and treated properly, can lead to diabetic coma, which can be fatal. A common early symptom of ketoacidosis is deep, rapid breathing as the body attempts to drive off CO2 and compensate for the acidosis. Another common symptom is fruity-smelling breath, due to the exhalation of acetone. Other symptoms include dry skin and mouth, a flushed face, nausea, vomiting and stomach pain. Treatment for diabetic coma is ingestion or injection of sugar; its prevention is the proper daily administration of insulin. A person who is diabetic and uses insulin can initiate ketoacidosis if a dose of insulin is missed. Between 2014 and 2015, there were over 7,000 hospitalisations due to diabetic ketoacidosis in Australia, with approximately 85% of patients suffering from type I diabetes. Approximately half of these patients were children and young adults (under 25 years old).
Section Review
A variety of buffering systems exist in the body that helps maintain the pH of the blood and other fluids within a narrow range—between pH 7.35 and 7.45. A buffer is a substance that prevents a radical change in fluid pH by absorbing excess hydrogen or hydroxyl ions. Most commonly, the substance that absorbs the ion is either a weak acid, which takes up a hydroxyl ion (OH–), or a weak base which takes up a hydrogen ion (H+). Several substances serve as buffers in the body, including cell and plasma proteins, haemoglobin, phosphates, bicarbonate ions and carbonic acid. The bicarbonate buffer is the primary buffering system of the IF surrounding the cells in tissues throughout the body. The respiratory and renal systems also play major roles in acid-base homeostasis by removing CO2 and hydrogen ions, respectively, from the body.
Identify the three blood variables considered when making a diagnosis of acidosis or alkalosis
Identify the source of compensation for blood pH problems of a respiratory origin
Identify the source of compensation for blood pH problems of a metabolic/renal origin
Normal arterial blood pH is restricted to a very narrow range of 7.35 to 7.45. A person who has a blood pH below 7.35 is considered to be in acidosis (actually, “physiological acidosis,” because blood is not truly acidic until its pH drops below 7), and a continuous blood pH below 7.0 can be fatal. Acidosis has several symptoms, including headache and confusion, and the individual can become lethargic and easily fatigued (Figure 17.15.1). A person who has a blood pH above 7.45 is considered to be in alkalosis, and a pH above 7.8 is fatal. Some symptoms of alkalosis include cognitive impairment (which can progress to unconsciousness), tingling or numbness in the extremities, muscle twitching and spasm, and nausea and vomiting. Both acidosis and alkalosis can be caused by either metabolic or respiratory disorders.
As discussed earlier in this chapter, the concentration of carbonic acid in the blood is dependent on the level of CO2 in the body and the amount of CO2 gas exhaled through the lungs. Thus, the respiratory contribution to acid-base balance is usually discussed in terms of CO2 (rather than of carbonic acid). Remember that a molecule of carbonic acid is lost for every molecule of CO2 exhaled, and a molecule of carbonic acid is formed for every molecule of CO2 retained.
Figure 17.15.1. Symptoms of acidosis and alkalosis. Symptoms of acidosis affect several organ systems. Both acidosis and alkalosis can be diagnosed using a blood test.
Metabolic acidosis occurs when the blood is too acidic (pH below 7.35) due to too little bicarbonate, a condition called primary bicarbonate deficiency. At the normal pH of 7.40, the ratio of bicarbonate to carbonic acid buffer is 20:1. If a person’s blood pH drops below 7.35, then he or she is in metabolic acidosis. The most common cause of metabolic acidosis is the presence of organic acids or excessive ketones in the blood. Table 17.15.1 lists some other causes of metabolic acidosis.
Table 17.15.1. Common causes of metabolic acidosis and blood metabolites
Cause
Metabolite
Diarrhoea
Bicarbonate
Uraemia
Phosphoric, sulfuric, and lactic acids
Diabetic ketoacidosis
Increased ketones
Strenuous exercise
Lactic acid
Methanol
Formic acid*
Paraldehyde
β-Hydroxybutyric acid*
Isopropanol
Propionic acid*
Ethylene glycol
Glycolic acid, and some oxalic acids and formic acids*
Salicylate/aspirin
Sulfosalicylic acid (SSA)*
* Acid metabolites from ingested chemical
The first three of the eight causes of metabolic acidosis listed are medical (or unusual physiological) conditions. Strenuous exercise can cause temporary metabolic acidosis due to the production of lactic acid. The last five causes result from the ingestion of specific substances. The active form of aspirin is its metabolite, sulfasalicylic acid. An overdose of aspirin causes acidosis due to the acidity of this metabolite. Metabolic acidosis can also result from uraemia, which is the retention of urea and uric acid. Metabolic acidosis can also arise from diabetic ketoacidosis, wherein an excess of ketones is present in the blood. Other causes of metabolic acidosis are a decrease in the excretion of hydrogen ions, which inhibits the conservation of bicarbonate ions, and excessive loss of bicarbonate ions through the gastrointestinal tract due to diarrhoea.
Metabolic Alkalosis: Primary Bicarbonate Excess
Metabolic alkalosis is the opposite of metabolic acidosis. It occurs when the blood is too alkaline (pH above 7.45) due to too much bicarbonate (called primary bicarbonate excess).
A transient excess of bicarbonate in the blood can follow ingestion of excessive amounts of bicarbonate, citrate, or antacids for conditions such as stomach acid reflux—known as heartburn. Cushing’s disease, which is the chronic hypersecretion of adrenocorticotrophic hormone (ACTH) by the anterior pituitary gland, can cause chronic metabolic alkalosis. The oversecretion of ACTH results in elevated aldosterone levels and an increased loss of potassium by urinary excretion. Other causes of metabolic alkalosis include the loss of hydrochloric acid from the stomach through vomiting, potassium depletion due to the use of diuretics for hypertension, and the excessive use of laxatives.
Respiratory acidosis occurs when the blood is overly acidic due to an excess of carbonic acid, resulting from too much CO2 in the blood. Respiratory acidosis can result from anything that interferes with respiration, such as pneumonia, emphysema, or congestive heart failure.
Respiratory alkalosis occurs when the blood is overly alkaline due to a deficiency in carbonic acid and CO2 levels in the blood. This condition usually occurs when too much CO2 is exhaled from the lungs, as occurs in hyperventilation, which is breathing that is deeper or more frequent than normal. An elevated respiratory rate leading to hyperventilation can be due to extreme emotional upset or fear, fever, infections, hypoxia, or abnormally high levels of catecholamines, such as adrenaline and noradrenaline. Surprisingly, aspirin overdose—salicylate toxicity—can result in respiratory alkalosis as the body tries to compensate for initial acidosis.
Compensation Mechanisms
Various compensatory mechanisms exist to maintain blood pH within a narrow range, including buffers, respiration and renal mechanisms. Although compensatory mechanisms usually work very well, when one of these mechanisms is not working properly (like kidney failure or respiratory disease), they have their limits. If the pH and bicarbonate to carbonic acid ratio are changed too drastically, the body may not be able to compensate. Moreover, extreme changes in pH can denature proteins. Extensive damage to proteins in this way can result in disruption of normal metabolic processes, serious tissue damage and ultimately, death.
Respiratory Compensation
Respiratory compensation for metabolic acidosis increases the respiratory rate to drive off CO2 and readjust the bicarbonate to carbonic acid ratio to the 20:1 level. This adjustment can occur within minutes. Respiratory compensation for metabolic alkalosis is not as adept as its compensation for acidosis. The normal response of the respiratory system to elevated pH is to increase the amount of CO2 in the blood by decreasing the respiratory rate to conserve CO2. There is a limit to the decrease in respiration, however, that the body can tolerate. Hence, the respiratory route is less efficient at compensating for metabolic alkalosis than for acidosis.
Metabolic Compensation
Metabolic and renal compensation for respiratory diseases that can create acidosis revolves around the conservation of bicarbonate ions. In cases of respiratory acidosis, the kidney increases the conservation of bicarbonate and secretion of H+ through the exchange mechanism discussed earlier. These processes increase the concentration of bicarbonate in the blood, re-establishing the proper relative concentrations of bicarbonate and carbonic acid. In cases of respiratory alkalosis, the kidneys decrease the production of bicarbonate and reabsorb H+ from the tubular fluid. These processes can be limited by the exchange of potassium by the renal cells, which use a K+-H+ exchange mechanism (antiporter).
Diagnosing Acidosis and Alkalosis
Lab tests for pH, CO2 partial pressure (pCO2) and HCO3– can identify acidosis and alkalosis, indicating whether the imbalance is respiratory or metabolic, and the extent to which compensatory mechanisms are working. The blood pH value, as shown in Table 17.15.2, indicates whether the blood is in acidosis, the normal range, or alkalosis. The pCO2 and total HCO3– values aid in determining whether the condition is metabolic or respiratory, and whether the patient has been able to compensate for the problem. Table 17.15.2 lists the conditions and laboratory results that can be used to classify these conditions. Metabolic acid-base imbalances typically result from kidney disease and the respiratory system usually responds to compensate.
Table 17.15.2. Types of acidosis and alkalosis
pH
pCO2
Total HCO3–
Metabolic acidosis
Decreases
Normal, then decreases
Decreases
Respiratory acidosis
Decreases
Increases
Normal, then increases
Metabolic alkalosis
Increases
Normal, then increases
Increases
Respiratory alkalosis
Increases
Decreases
Normal, then decreases
Reference values (arterial): pH: 7.35-7.45; pCO2: male: 35-48 mmHg, female: 32-45 mmHg; total venous bicarbonate: 22-29 mM.
Metabolic acidosis is problematic, as lower-than-normal amounts of bicarbonate are present in the blood. The pCO2 would be normal at first, but if compensation has occurred, it would decrease as the body re-establishes the proper ratio of bicarbonate and carbonic acid/CO2.
Respiratory acidosis is problematic, as excess CO2 is present in the blood. Bicarbonate levels would be normal at first, but if compensation has occurred, they would increase in an attempt to re-establish the proper ratio of bicarbonate and carbonic acid/CO2.
Alkalosis is characterised by a higher-than-normal pH. Metabolic alkalosis is problematic, as elevated pH and excess bicarbonate are present. The pCO2 would again be normal at first, but if compensation has occurred, it would increase as the body attempts to re-establish the proper ratios of bicarbonate and carbonic acid/CO2.
Respiratory alkalosis is problematic, as CO2 deficiency is present in the bloodstream. The bicarbonate concentration would be normal at first. When renal compensation occurs, however, the bicarbonate concentration in blood decreases as the kidneys attempt to re-establish the proper ratios of bicarbonate and carbonic acid/CO2 by eliminating more bicarbonate to bring the pH into the physiological range.
Section Review
Acidosis and alkalosis describe conditions in which a person’s blood is, respectively, too acidic (pH below 7.35) and too alkaline (pH above 7.45). Each of these conditions can be caused either by metabolic problems related to bicarbonate levels or by respiratory problems related to carbonic acid and CO2 levels. Several compensatory mechanisms allow the body to maintain a normal pH.
abdominal aorta – portion of the aorta inferior to the aortic hiatus and superior to the common iliac arteries.
abdominopelvic cavity – division of the anterior (ventral) cavity that houses the abdominal and pelvic viscera.
abducens nerve – sixth cranial nerve; responsible for contraction of one of the extraocular muscles.
abduction – movement in the coronal plane that moves a limb laterally away from the body; spreading of the fingers.
ABO blood group – blood-type classification based on the presence or absence of A and B carbohydrates on the erythrocyte membrane surface.
absolute refractory period – time during an action period when another action potential cannot be generated because the voltage-gated Na+ channel is inactivated.
absorption – passage of digested products from the intestinal lumen through mucosal cells and into the bloodstream or lacteals.
accessory digestive organ – includes teeth, tongue, salivary glands, gallbladder, liver, and pancreas.
accessory duct – (also, duct of Santorini) duct that runs from the pancreas into the duodenum.
acclimatisation – process of adjustment that the respiratory system makes due to chronic exposure to high altitudes.
acetabular labrum – lip of fibrocartilage that surrounds outer margin of the acetabulum on the hip bone.
acetabulum – large, cup-shaped cavity located on the lateral side of the hip bone; formed by the junction of the ilium, pubis, and ischium portions of the hip bone.
acetylcholine (ACh) – neurotransmitter that binds at a motor end-plate to trigger depolarisation.
acinus – cluster of glandular epithelial cells in the pancreas that secretes pancreatic secretion in the pancreas.
acne – skin condition due to infected sebaceous glands.
acromegaly – disorder in adults caused when abnormally high levels of GH trigger growth of bones in the face, hands, and feet.
acromial end of the clavicle – lateral end of the clavicle that articulates with the acromion of the scapula.
acromial process – acromion of the scapula.
acromioclavicular joint – articulation between the acromion of the scapula and the acromial end of the clavicle.
acromion – flattened bony process that extends laterally from the scapular spine to form the bony tip of the shoulder.
acrosome – cap-like vesicle located at the anterior-most region of a sperm that is rich with lysosomal enzymes capable of digesting the protective layers surrounding the oocyte.
acrosomal reaction – release of digestive enzymes by sperm that enables them to burrow through the corona radiata and penetrate the zona pellucida of an oocyte prior to fertilisation.
actin – protein that makes up most of the thin myofilaments in a sarcomere muscle fibre.
action potential – change in voltage of a cell membrane in response to a stimulus that results in transmission of an electrical signal; unique to neurons and muscle fibres.
activation gate – part of the voltage-gated Na+ channel that opens when the membrane voltage reaches threshold.
active immunity – immunity developed from an individual’s own immune system.
active transport – form of transport across the cell membrane that requires input of cellular energy.
acute inflammation – inflammation occurring for a limited time period; rapidly developing.
acute mountain sickness (AMS) – condition that occurs a result of acute exposure to high altitude due to a low partial pressure of oxygen.
adaptive immune response – relatively slow but very specific and effective immune response controlled by lymphocytes.
adductor tubercle – small, bony bump located on the superior aspect of the medial epicondyle of the femur.
adenylyl cyclase – membrane-bound enzyme that converts ATP to cyclic AMP, creating cAMP, as a result of G-protein activation.
α-dextrin – breakdown product of starch.
α-dextrinase – brush border enzyme that acts on α-dextrins.
adipocytes – lipid storage cells.
adipose tissue – specialised areolar tissue rich in stored fat.
adrenal artery – branch of the abdominal aorta; supplies blood to the adrenal (suprarenal) glands.
adrenal cortex – outer region of the adrenal glands consisting of multiple layers of epithelial cells and capillary networks that produces mineralocorticoids and glucocorticoids.
adrenal glands – endocrine glands located at the top of each kidney that are important for the regulation of the stress response, blood pressure and blood volume, water homeostasis, and electrolyte levels.
adrenal medulla – inner layer of the adrenal glands that plays an important role in the stress response by producing adrenaline and noradrenaline.
adrenal vein – drains the adrenal or suprarenal glands that are immediately superior to the kidneys; the right adrenal vein enters the inferior vena cava directly and the left adrenal vein enters the left renal vein.
adrenaline – primary and most potent catecholamine hormone secreted by the adrenal medulla in response to short-term stress.
adrenocorticotropic hormone (ACTH) – anterior pituitary hormone that stimulates the adrenal cortex to secrete corticosteroid hormones (also called corticotropin).
aerobic respiration – production of ATP in the presence of oxygen.
afferent lymphatic vessels – lead into a lymph node.
afterload – force the ventricles must develop to effectively pump blood against the resistance in the vessels.
agglutination – clustering of cells into masses linked by antibodies.
agranular leukocytes – leukocytes with few visible granules in their cytoplasm; specifically, monocytes, lymphocytes, and NK cells.
ala – (plural = alae) small, flaring structure of a nostril that forms the lateral side of the nares.
alar cartilage – cartilage that supports the apex of the nose and helps shape the nares; it is connected to the septal cartilage and connective tissue of the alae.
alarm reaction – the short-term stress, or the fight-or-flight response, of stage one of the general adaptation syndrome mediated by the hormones adrenaline and noradrenaline.
alar plate – developmental region of the spinal cord that gives rise to the posterior horn of the grey matter.
albinism – genetic disorder that affects the skin, in which there is no melanin production.
albumin – most abundant plasma protein, accounting for most of the osmotic pressure of plasma.
aldosterone – hormone produced and secreted by the adrenal cortex that stimulates sodium and fluid retention and increases blood volume and blood pressure.
alkaloid – substance, usually from a plant source, that is chemically basic with respect to pH and will stimulate bitter receptors.
allantois – finger-like outpocketing of yolk sac forms the primitive excretory duct of the embryo; precursor to the urinary bladder.
alpha cell – pancreatic islet cell type that produces the hormone glucagon.
alpha-helix structure (α-helix) – type of secondary protein structure formed by folding the polypeptide into a helix shape with hydrogen bonds stabilising the structure.
alveolar dead space – air space within alveoli that are unable to participate in gas exchange.
alveolar duct – small tube that leads from the terminal bronchiole to the respiratory bronchiole and is the point of attachment for alveoli.
alveolar macrophage – immune system cell of the alveolus that removes debris and pathogens.
alveolar pore – opening that allows airflow between neighbouring alveoli.
alveolar process of the mandible – upper border of mandibular body that contains the lower teeth.
alveolar process of the maxilla – curved, inferior margin of the maxilla that supports and anchors the upper teeth.
alveolar sac – cluster of alveoli.
alveoli – (of the breast) milk-secreting cells in the mammary gland.
alveolus – small, grape-like sac that performs gas exchange in the lungs.
amacrine cell – type of cell in the retina that connects to the bipolar cells near the outer synaptic layer and provides the basis for early image processing within the retina.
amino acid – a protein’s monomer; has a central carbon or alpha carbon to which an amino group, a carboxyl group, a hydrogen, and an R group or side chain is attached; the R group is different for all 20 common amino acids.
amnion – transparent membranous sac that encloses the developing foetus and fills with amniotic fluid.
amniotic cavity – cavity that opens between the inner cell mass and the trophoblast; develops into amnion.
aminopeptidase – brush border enzyme that acts on proteins.
amphiarthrosis – slightly mobile joint.
amphipathic – descr.ibes a molecule that exhibits a difference in polarity between its two ends, resulting in a difference in water solubility.
ampulla – in the ear, the structure at the base of a semicircular canal that contains the hair cells and cupula for transduction of rotational movement of the head.
ampulla – (of the uterine tube) middle portion of the uterine tube in which fertilisation often occurs
amygdala – nucleus deep in the temporal lobe of the cerebrum that is related to memory and emotional behaviour.
anaemia – deficiency of red blood cells or haemoglobin.
anagen – active phase of the hair growth cycle.
anal canal – final segment of the large intestine.
anal column – long fold of mucosa in the anal canal.
anal sinus – recess between anal columns.
anaphase – third stage of mitosis (and meiosis), during which sister chromatids separate into two new nuclear regions of a dividing cell.
anaphylactic shock – type of shock that follows a severe allergic reaction and results from massive vasodilation.
anastomosis – (plural = anastomoses) area where vessels unite to allow blood to circulate even if there may be partial blockage in another branch.
anatomical dead space – air space present in the airway that never reaches the alveoli and therefore never participates in gas exchange.
anatomical neck – line on the humerus located around the outside margin of the humeral head.
anatomical position – standard reference position used for describing locations and directions on the human body.
anatomical sphincter – smooth or skeletal muscle surrounding the lumen of a vessel or hollow organ that can restrict flow when contracted
anatomy – science that studies the form and composition of the body’s structures.
anchoring junction – mechanically attaches adjacent cells to each other or to the basement membrane.
angioblasts – stem cells that give rise to blood vessels.
angiogenesis – development of new blood vessels from existing vessels.
angiotensin-converting enzyme – the enzyme that c.
angle of the mandible – rounded corner located at outside margin of the body and ramus junction.
angle of the rib – portion of rib with greatest curvature; together, the rib angles form the most posterior extent of the thoracic cage.
angiotensin-converting enzyme (ACE) – enzyme produced by the lungs that catalyses the reaction of inactive angiotensin I into active angiotensin II.
angiotensin I – protein produced by the enzymatic action of renin on angiotensinogen; inactive precursor of angiotensin II.
angiotensin II – protein produced by the enzymatic action of ACE on inactive angiotensin I; actively causes vasoconstriction and stimulates aldosterone release by the adrenal cortex.
angiotensinogen – inactive protein in the circulation produced by the liver; precursor of angiotensin I; must be modified by the enzymes renin and ACE to be activated.
ankle joint – joint that separates the leg and foot portions of the lower limb; formed by the articulations between the talus bone of the foot inferiorly, and the distal end of the tibia, medial malleolus of the tibia, and lateral malleolus of the fibula superiorly.
annular ligament – intrinsic ligament of the elbow articular capsule that surrounds and supports the head of the radius at the proximal radioulnar joint.
anosmia – loss of the sense of smell; usually the result of physical disruption of the first cranial nerve.
aqueous humour – watery fluid that fills the anterior chamber containing the cornea, iris, ciliary body, and lens of the eye.
anterior – describes the front or direction toward the front of the body; also referred to as ventral.
anterior arch – anterior portion of the ring-like C1 (atlas) vertebra.
anterior border of the tibia – narrow, anterior margin of the tibia that extends inferiorly from the tibial tuberosity.
anterior cardiac veins – vessels that parallel the small cardiac arteries and drain the anterior surface of the right ventricle; bypass the coronary sinus and drain directly into the right atrium.
anterior cavity – larger body cavity located anterior to the posterior (dorsal) body cavity; includes the serous membrane-lined pleural cavities for the lungs, pericardial cavity for the heart, and peritoneal cavity for the abdominal and pelvic organs; also referred to as ventral cavity.
anterior cerebral artery – arises from the internal carotid artery; supplies the frontal lobe of the cerebrum.
anterior column – white matter between the anterior horns of the spinal cord composed of many different groups of axons of both ascending and descending tracts.
anterior communicating artery – anastomosis of the right and left internal carotid arteries; supplies blood to the brain.
anterior corticospinal tract – division of the corticospinal pathway that travels through the ventral (anterior) column of the spinal cord and controls axial musculature through the medial motor neurons in the ventral (anterior) horn.
anterior cranial fossa – shallowest and most anterior cranial fossa of the cranial base that extends from the frontal bone to the lesser wing of the sphenoid bone.
anterior cruciate ligament – intracapsular ligament of the knee; extends from anterior, superior surface of the tibia to the inner aspect of the lateral condyle of the femur; resists hyperextension of knee.
anterior horn – grey matter of the spinal cord containing multipolar motor neurons, sometimes referred to as the ventral horn.
anterior inferior iliac spine – small, bony projection located on the anterior margin of the ilium, below the anterior superior iliac spine.
anterior interventricular artery – (also, left anterior descending artery or LAD) major branch of the left coronary artery that follows the anterior interventricular sulcus.
anterior interventricular sulcus – sulcus located between the left and right ventricles on the anterior surface of the heart.
anterior longitudinal ligament – ligament that runs the length of the vertebral column, uniting the anterior aspects of the vertebral bodies.
anterior median fissure – deep midline feature of the anterior spinal cord, marking the separation between the right and left sides of the cord.
anterior sacroiliac ligament – strong ligament between the sacrum and the ilium portions of the hip bone that supports the anterior side of the sacroiliac joint.
anterior spinal artery – blood vessel from the merged branches of the vertebral arteries that runs along the anterior surface of the spinal cord.
anterior superior iliac spine – rounded, anterior end of the iliac crest.
anterior talofibular ligament – intrinsic ligament located on the lateral side of the ankle joint, between talus bone and lateral malleolus of fibula; supports talus at the talocrural joint and resists excess inversion of the foot.
anterior tibial artery – branches from the popliteal artery; supplies blood to the anterior tibial region; becomes the dorsalis pedis artery.
anterior tibial vein – forms from the dorsal venous arch; drains the area near the tibialis anterior muscle and leads to the popliteal vein.
anterior (ventral) sacral foramen – one of the series of paired openings located on the anterior (ventral) side of the sacrum.
antibodies – (also, immunoglobulins or gamma globulins) antigen-specific proteins produced by specialised B lymphocytes that protect the body by binding to foreign objects such as bacteria and viruses.
anticoagulant – substance such as heparin that opposes coagulation.
anticodon – consecutive sequence of three nucleotides on a tRNA molecule that is complementary to a specific codon on an mRNA molecule.
antidiuretic hormone (ADH) – hypothalamic hormone that is stored by the posterior pituitary and that signals the kidneys to reabsorb water.
antigen – molecule recognised by the receptors of B and T lymphocytes.
antigenic determinant – (also, epitope) one of the chemical groups recognised by a single type of lymphocyte antigen receptor.
antigen presentation – binding of processed antigen to the protein-binding cleft of a major histocompatibility complex molecule.
antigen processing – internalisation and digestion of antigen in an antigen-presenting cell.
antigen receptor – two-chain receptor by which lymphocytes recognise antigen.
angiogenesis – formation of blood capillary networks.
antithrombin – anticoagulant that inactivates factor X and opposes the conversion of prothrombin (factor II) into thrombin in the common pathway.
antrum – fluid-filled chamber that characterises a mature tertiary (antral) follicle.
anulus fibrosus – tough, fibrous outer portion of an intervertebral disc, which is strongly anchored to the bodies of the adjacent vertebrae.
anuria – absence of urine produced; production of 50 mL or less per day.
aorta – largest artery in the body, originating from the left ventricle and descending to the abdominal region where it bifurcates into the common iliac arteries at the level of the fourth lumbar vertebra; arteries originating from the aorta distribute blood to virtually all tissues of the body.
aortic arch – arc that connects the ascending aorta to the descending aorta; ends at the intervertebral disk between the fourth and fifth thoracic vertebrae.
aortic hiatus – opening in the diaphragm that allows passage of the thoracic aorta into the abdominal region where it becomes the abdominal aorta.
aortic sinuses – small pockets in the ascending aorta near the aortic valve that are the locations of the baroreceptors (stretch receptors) and chemoreceptors that trigger a reflex that aids in the regulation of vascular homeostasis.
aortic valve – (also, aortic semilunar valve) valve located at the base of the aorta.
apex – tip of the external nose.
apical – that part of a cell or tissue which, in general, faces an open space.
apical ectodermal ridge – enlarged ridge of ectoderm at the distal end of a limb bud that stimulates growth and elongation of the limb.
apneustic centre – network of neurons within the pons that stimulate the neurons in the dorsal respiratory group; controls the depth of inspiration.
apocrine secretion – release of a substance along with the apical portion of the cell.
aponeurosis – broad, tendon-like sheet of connective tissue that attaches a skeletal muscle to another skeletal muscle or to a bone.
apoptosis – programmed cell death.
apocrine sweat gland – type of sweat gland that is associated with hair follicles in the armpits and genital regions.
appendicular skeleton – all bones of the upper and lower limbs, plus the girdle bones that attach each limb to the axial skeleton.
appendix – (vermiform appendix) coiled tube attached to the cecum.
aquaporin – protein-forming water channels through the lipid bilayer of the cell; allows water to cross; activation in the collecting ducts is under the control of ADH.
arachnoid granulation – out-pocket of the arachnoid membrane into the dural sinuses that allows for reabsorption of CSF into the blood.
arachnoid mater – middle layer of the meninges named for the spider-web–like trabeculae that extend between it and the pia mater.
arachnoid trabeculae – filaments between the arachnoid and pia mater within the subarachnoid space.
arcuate line of the ilium – smooth ridge located at the inferior margin of the iliac fossa; forms the lateral portion of the pelvic brim.
areola – highly pigmented, circular area surrounding the raised nipple and containing areolar glands that secrete fluid important for lubrication during suckling.
areolar tissue – (also, loose connective tissue) a type of connective tissue proper that shows little specialisation with cells dispersed in the matrix.
arm – region of the upper limb located between the shoulder and elbow joints; contains the humerus bone.
arrector pili – smooth muscle that is activated in response to external stimuli that pull on hair follicles and make the hair “stand up”.
arterial circle – (also, circle of Willis) anastomosis located at the base of the brain that ensures continual blood supply; formed from branches of the internal carotid and vertebral arteries; supplies blood to the brain.
arteriole – (also, resistance vessel) very small artery that leads to a capillary.
arteriovenous anastomosis – short vessel connecting an arteriole directly to a venule and bypassing the capillary beds.
artery – blood vessel that conducts blood away from the heart; may be a conducting or distributing vessel.
articular capsule – connective tissue structure that encloses the joint cavity of a synovial joint.
articular cartilage – thin layer of cartilage covering an epiphysis; reduces friction and acts as a shock absorber.
articular disc – meniscus; a fibrocartilage structure found between the bones of some synovial joints; provides padding or smooths movements between the bones; strongly unites the bones together.
articular tubercle – smooth ridge located on the inferior skull, immediately anterior to the mandibular fossa.
articulation – where two bone surfaces meet.
artificial pacemaker – medical device that transmits electrical signals to the heart to ensure that it contracts and pumps blood to the body.
ascending aorta – initial portion of the aorta, rising from the left ventricle for a distance of approximately 5 cm.
ascending tract – central nervous system fibres carrying sensory information from the spinal cord or periphery to the brain.
ascending colon – first region of the colon.
ascending pathway – fibre structure that relays sensory information from the periphery through the spinal cord and brain stem to other structures of the brain.
association area – region of cortex connected to a primary sensory cortical area that further processes the information to generate more complex sensory perceptions.
astrocyte – star-shaped cell in the central nervous system that regulates ions and uptake and/or breakdown of some neurotransmitters and contributes to the formation of the blood-brain barrier.
atmospheric pressure – amount of force that is exerted by gases in the air surrounding any given surface.
atlantoaxial joint – series of three articulations between the atlas (C1) vertebra and the axis (C2) vertebra, consisting of the joints between the inferior articular processes of C1 and the superior articular processes of C2, and the articulation between the dens of C2 and the anterior arch of C1.
atlanto-occipital joint – articulation between the occipital condyles of the skull and the superior articular processes of the atlas (C1 vertebra).
atlas – first cervical (C1) vertebra.
ATPase – enzyme that hydrolyses ATP to ADP.
atrial natriuretic peptide (ANP) – peptide hormone produced by the walls of the atria in response to high blood pressure, blood volume, or blood sodium that reduces the reabsorption of sodium and water in the kidneys and promotes vasodilation.
atrial reflex – (also, called Bainbridge reflex) autonomic reflex that responds to stretch receptors in the atria that send impulses to the cardioaccelerator area to increase HR when venous flow into the atria increases.
atrioventricular bundle – (also, bundle of His) group of specialised myocardial conductile cells that transmit the impulse from the AV node through the interventricular septum; form the left and right atrioventricular bundle branches.
atrioventricular bundle branches – (also, left or right bundle branches) specialised myocardial conductile cells that arise from the bifurcation of the atrioventricular bundle and pass through the interventricular septum; lead to the Purkinje fibres and also to the right papillary muscle via the moderator band.
atrioventricular (AV) node – clump of myocardial cells located in the inferior portion of the right atrium within the atrioventricular septum; receives the impulse from the SA node, pauses, and then transmits it into specialised conducting cells within the interventricular septum.
atrioventricular septum – cardiac septum located between the atria and ventricles; atrioventricular valves are located here.
atrioventricular valves – one-way valves located between the atria and ventricles; the valve on the right is called the tricuspid valve, and the one on the left is the mitral or bicuspid valve.
atrium – (plural = atria) upper or receiving chamber of the heart that pumps blood into the lower chambers just prior to their contraction; the right atrium receives blood from the systemic circuit that flows into the right ventricle; the left atrium receives blood from the pulmonary circuit that flows into the left ventricle.
atrophy – loss of mass and function.
audition – sense of hearing.
auricle – extension of an atrium visible on the superior surface of the heart.
auricle – fleshy external structure of the ear.
auricular surface of the ilium – roughened area located on the posterior, medial side of the ilium of the hip bone; articulates with the auricular surface of the sacrum to form the sacroiliac joint.
autocrine – chemical signal that elicits a response in the same cell that secreted it.
autolysis – breakdown of cells by their own enzymatic action.
autonomic nervous system (ANS) – functional division of the nervous system that is responsible for homeostatic reflexes that coordinate control of cardiac and smooth muscle, as well as glandular tissue.
autonomic tone – contractile state during resting cardiac activity produced by mild sympathetic and parasympathetic stimulation.
autophagy – lysosomal breakdown of a cell’s own components.
autorhythmicity – ability of cardiac muscle to initiate its own electrical impulse that triggers the mechanical contraction that pumps blood at a fixed pace without nervous or endocrine control.
axial skeleton – central, vertical axis of the body, including the skull, vertebral column, and thoracic cage.
axillary artery – continuation of the subclavian artery as it penetrates the body wall and enters the axillary region; supplies blood to the region near the head of the humerus (humeral circumflex arteries); the majority of the vessel continues into the brachium and becomes the brachial artery.
axillary nerve – systemic nerve of the arm that arises from the brachial plexus.
axillary vein – major vein in the axillary region; drains the upper limb and becomes the subclavian vein.
axis – second cervical (C2) vertebra.
axon – single process of the neuron that carries an electrical signal (action potential) away from the cell body toward a target cell.
axon hillock – tapering of the neuron cell body that gives rise to the axon.
axon segment – single stretch of the axon insulated by myelin and bounded by nodes of Ranvier at either end (except for the first, which is after the initial segment, and the last, which is followed by the axon terminal).
axon terminal – end of the axon, where there are usually several branches extending toward the target cell.
axoplasm – cytoplasm of an axon, which is different in composition than the cytoplasm of the neuronal cell body.
azygos vein – originates in the lumbar region and passes through the diaphragm into the thoracic cavity on the right side of the vertebral column; drains blood from the intercostal veins, oesophageal veins, bronchial veins, and other veins draining the mediastinal region; leads to the superior vena cava.
B
Bachmann’s bundle – (also, interatrial band) group of specialised conducting cells that transmit the impulse directly from the SA node in the right atrium to the left atrium.
Bainbridge reflex – (also, called atrial reflex) autonomic reflex that responds to stretch receptors in the atria that send impulses to the cardioaccelerator area to increase HR when venous flow into the atria increases.
ball-and-socket joint – synovial joint formed between the spherical end of one bone (the ball) that fits into the depression of a second bone (the socket); found at the hip and shoulder joints; functionally classified as a multiaxial joint.
baroreceptor reflex – autonomic reflex in which the cardiac centres monitor signals from the baroreceptor stretch receptors and regulate heart function based on blood flow.
barrier defences – antipathogen defences deriving from a barrier that physically prevents pathogens from entering the body to establish an infection.
Bartholin’s glands – (also, greater vestibular glands) glands that produce a thick mucus that maintains moisture in the vulva area; also referred to as the greater vestibular glands.
basal cell – type of stem cell found in the stratum basale and in the hair matrix that continually undergoes cell division, producing the keratinocytes of the epidermis.
basal cell carcinoma – cancer that originates from basal cells in the epidermis of the skin.
basal forebrain – nuclei of the cerebrum related to modulation of sensory stimuli and attention through broad projections to the cerebral cortex, loss of which is related to Alzheimer’s disease.
basal lamina – thin extracellular layer that lies underneath epithelial cells and separates them from other tissues.
basal nuclei – nuclei of the cerebrum (with a few components in the upper brain stem and diencephalon) that are responsible for assessing cortical movement commands and comparing them with the general state of the individual through broad modulatory activity of dopamine neurons; largely related to motor functions, as evidenced through the symptoms of Parkinson’s and Huntington’s diseases.
basal plate – developmental region of the spinal cord that gives rise to the lateral and anterior horns of grey matter.
base of the metatarsal bone – expanded, proximal end of each metatarsal bone.
basement membrane – in epithelial tissue, a thin layer of fibrous material that anchors the epithelial tissue to the underlying connective tissue; made up of the basal lamina and reticular lamina.
basilar artery – formed from the fusion of the two vertebral arteries; sends branches to the cerebellum, brain stem, and the posterior cerebral arteries; the main blood supply to the brain stem.
basilar membrane – in the ear, the floor of the cochlear duct on which the organ of Corti sits.
basilic vein – superficial vein of the arm that arises from the palmar venous arches, intersects with the median cubital vein, parallels the ulnar vein, and continues into the upper arm; along with the brachial vein, it leads to the axillary vein.
basophils – granulocytes that stain with a basic (alkaline) stain and store histamine and heparin.
B cells – lymphocytes that act by differentiating into an antibody-secreting plasma cell.
bedsore – sore on the skin that develops when regions of the body start necrotising due to constant pressure and lack of blood supply; also called decubitis ulcers.
beta cell – pancreatic islet cell type that produces the hormone insulin.
beta-pleated sheet (β-pleated) – secondary structure in proteins in which hydrogen bonding forms “pleats” between atoms on the polypeptide chain’s backbone.
Betz cells – output cells of the primary motor cortex that cause musculature to move through synapses on cranial and spinal motor neurons.
biaxial joint – type of diarthrosis; a joint that allows for movements within two planes (two axes).
bicipital groove – intertubercular groove; narrow groove located between the greater and lesser tubercles of the humerus.
bicuspid valve – (also, mitral valve or left atrioventricular valve) valve located between the left atrium and ventricle; consists of two flaps of tissue.
bile – alkaline solution produced by the liver and important for the emulsification of lipids.
bile canaliculus – small duct between hepatocytes that collects bile.
bilirubin – yellowish bile pigment produced when iron is removed from haem and the remaining molecule is further broken down into waste products.
biliverdin – green bile pigment produced during the conversion of non-iron haem to bilirubin.
binocular depth cues – indications of the distance of visual stimuli on the basis of slight differences in the images projected onto either retina.
biogenic amine – class of neurotransmitters that are enzymatically derived from amino acids but no longer contain a carboxyl group.
biological macromolecule – large molecule necessary for life that is built from smaller organic molecules.
bipolar – shape of a neuron with two processes extending from the neuron cell body—the axon and one dendrite.
bipolar cell – cell type in the retina that connects the photoreceptors to the RGCs.
blastocoel – fluid-filled cavity of the blastocyst.
blastocyst – term for the conceptus at the developmental stage that consists of about 100 cells shaped into an inner cell mass that is fated to become the embryo and an outer trophoblast that is fated to become the associated foetal membranes and placenta.
blastomere – daughter cell of a cleavage.
blood – liquid connective tissue composed of cellular components—erythrocytes, leukocytes, and platelets—and a fluid extracellular matrix called plasma; component of the cardiovascular system.
blood-brain barrier (BBB) – physiological barrier between the circulatory system and the central nervous system that establishes a privileged blood supply, restricting the flow of substances into the CNS.
blood colloidal osmotic pressure (BCOP) – pressure exerted by colloids suspended in blood within a vessel; a primary determinant is the presence of plasma proteins.
blood hydrostatic pressure – force blood exerts against the walls of a blood vessel or heart chamber.
blood flow – movement of blood through a vessel, tissue, or organ that is usually expressed in terms of volume per unit of time.
blood islands – masses of developing blood vessels and formed elements from mesodermal cells scattered throughout the embryonic disc.
blood pressure – force exerted by the blood against the wall of a vessel or heart chamber; can be described with the more generic term hydrostatic pressure.
blood–testis barrier – tight junctions between Sertoli cells that prevent bloodborne pathogens from gaining access to later stages of spermatogenesis and prevent the potential for an autoimmune reaction to haploid sperm.
blood typing or grouping – blood test for identification of blood type using commercially prepared antibodies and small samples of blood.
B lymphocytes – (also, B cells) lymphocytes that defend the body against specific pathogens and thereby provide specific (or adaptive) immunity.
body – mid-portion of the stomach.
body of uterus – middle section of the uterus.
body of the rib – shaft portion of a rib..
Bohr effect – relationship between blood pH and oxygen dissociation from haemoglobin.
bolus – mass of chewed food.
bone – hard, dense connective tissue that forms the structural elements of the skeleton.
bone marrow – tissue found inside bones; the site of all blood cell differentiation and maturation of B lymphocytes.
bone marrow biopsy – diagnostic test of a sample of red bone marrow.
bone marrow transplant – treatment in which a donor’s healthy bone marrow with its stem cells replaces diseased or damaged bone marrow of a patient.
Bowman’s capsule – cup-shaped sack lined by a simple squamous epithelium (parietal surface) and specialised cells called podocytes (visceral surface) that participate in the filtration process; receives the filtrate which then passes on to the PCTs.
Boyle’s law – relationship between volume and pressure as described by the formula: P1V1 = P2V2.
brachial artery – continuation of the axillary artery in the brachium; supplies blood to much of the brachial region; gives off several smaller branches that provide blood to the posterior surface of the arm in the region of the elbow; bifurcates into the radial and ulnar arteries at the coronoid fossa.
brachial plexus – nerve plexus associated with the lower cervical spinal nerves and first thoracic spinal nerve.
brachial vein – deeper vein of the arm that forms from the radial and ulnar veins in the lower arm; leads to the axillary vein.
brachiocephalic artery – single vessel located on the right side of the body; the first vessel branching from the aortic arch; gives rise to the right subclavian artery and the right common carotid artery; supplies blood to the head, neck, upper limb, and wall of the thoracic region.
brachiocephalic vein – one of a pair of veins that form from a fusion of the external and internal jugular veins and the subclavian vein; subclavian, external and internal jugulars, vertebral, and internal thoracic veins lead to it; drains the upper thoracic region and flows into the superior vena cava.
brain – the large organ of the central nervous system composed of white and grey matter, contained within the cranium and continuous with the spinal cord.
brain case – portion of the skull that contains and protects the brain, consisting of the eight bones that form the cranial base and rounded upper skull.
brain stem – region of the adult brain that includes the midbrain, pons, and medulla oblongata and develops from the mesencephalon, metencephalon, and myelencephalon of the embryonic brain.
bridge – portion of the external nose that lies in the area of the nasal bones.
broad ligament – wide ligament that supports the uterus by attaching laterally to both sides of the uterus and pelvic wall.
Broca’s area – region of the frontal lobe associated with the motor commands necessary for speech production and located only in the cerebral hemisphere responsible for language production, which is the left side in approximately 95 percent of the population.
Brodmann’s areas – mapping of regions of the cerebral cortex based on microscopic anatomy that relates specific areas to functional differences, as described by Brodmann in the early 1900s.
bronchial artery – systemic branch from the aorta that provides oxygenated blood to the lungs in addition to the pulmonary circuit.
bronchial bud – structure in the developing embryo that forms when the laryngotracheal bud extends and branches to form two bulbous structures.
bronchial tree – collective name for the multiple branches of the bronchi and bronchioles of the respiratory system.
bronchial vein – drains the systemic circulation from the lungs and leads to the azygos vein.
bronchiole – branch of bronchi that are 1 mm or less in diameter and terminate at alveolar sacs.
bronchoconstriction – decrease in the size of the bronchiole due to contraction of the muscular wall.
bronchodilation – increase in the size of the bronchiole due to contraction of the muscular wall.
bronchus – tube connected to the trachea that branches into many subsidiaries and provides a passageway for air to enter and leave the lungs.
bronchus-associated lymphoid tissue (BALT) – lymphoid nodule associated with the respiratory tract.
brush border – fuzzy appearance of the small intestinal mucosa created by microvilli.
brush border – formed by microvilli on the surface of certain cuboidal cells; in the kidney it is found in the PCT; increases surface area for absorption in the kidney.
buffy coat – thin, pale layer of leukocytes and platelets that separates the erythrocytes from the plasma in a sample of centrifuged blood.
bulbourethral glands – (also, Cowper’s glands) glands that secrete a lubricating mucus that cleans and lubricates the urethra prior to and during ejaculation.
bulbus cordis – portion of the primitive heart tube that will eventually develop into the right ventricle.
bundle of His – (also, atrioventricular bundle) group of specialised myocardial conductile cells that transmit the impulse from the AV node through the interventricular septum; form the left and right atrioventricular bundle branches.
bursa – connective tissue sac containing lubricating fluid that prevents friction between adjacent structures, such as skin and bone, tendons and bone, or between muscles.
C
calcaneofibular ligament – intrinsic ligament located on the lateral side of the ankle joint, between the calcaneus bone and lateral malleolus of the fibula; supports the talus bone at the ankle joint and resists excess inversion of the foot.
calcaneus – heel bone; posterior, inferior tarsal bone that forms the heel of the foot.
calcitonin – peptide hormone produced and secreted by the parafollicular cells (C cells) of the thyroid gland that functions to decrease blood calcium levels.
calyces – cup-like structures receiving urine from the collecting ducts where it passes on to the renal pelvis and ureter.
callus – thickened area of skin that arises due to constant abrasion.
calmodulin – regulatory protein that facilitates contraction in smooth muscles.
calvaria – (also, skullcap) rounded top of the skull.
canaliculi – (singular = canaliculus) channels within the bone matrix that house one of an osteocyte’s many cytoplasmic extensions that it uses to communicate and receive nutrients.
capacitance – ability of a vein to distend and store blood.
capacitance vessels – veins.
capacitation – process that occurs in the female reproductive tract in which sperm are prepared for fertilisation; leads to increased motility and changes in their outer membrane that improve their ability to release enzymes capable of digesting an oocyte’s outer layers.
capillary – smallest of blood vessels where physical exchange occurs between the blood and tissue cells surrounded by interstitial fluid.
capillary bed – network of 10–100 capillaries connecting arterioles to venules.
capillary hydrostatic pressure (CHP) – force blood exerts against a capillary.
capitate – from the lateral side, the third of the four distal carpal bones; articulates with the scaphoid and lunate proximally, the trapezoid laterally, the hamate medially, and primarily with the third metacarpal distally.
capitulum – knob-like bony structure located anteriorly on the lateral, distal end of the humerus.
capsaicin – molecule that activates nociceptors by interacting with a temperature-sensitive ion channel and is the basis for “hot” sensations in spicy food.
carbaminohemoglobin – compound of carbon dioxide and haemoglobin, and one of the ways in which carbon dioxide is carried in the blood.
carbohydrate – biological macromolecule in which the ratio of carbon to hydrogen and to oxygen is 1:2:1; carbohydrates serve as energy sources and structural support in cells and form arthropods’ cellular exoskeleton.
carbonic anhydrase (CA) – enzyme that catalyses the reaction that causes carbon dioxide and water to form carbonic acid.
cardia – (also, cardiac region) part of the stomach surrounding the cardiac orifice (oesophageal hiatus).
cardiac cycle – period of time between the onset of atrial contraction (atrial systole) and ventricular relaxation (ventricular diastole).
cardiac muscle – heart muscle, under involuntary control, composed of striated cells that attach to form fibres, each cell contains a single nucleus, contracts autonomously.
cardiac notch – depression in the medial surface of the inferior lobe of the left lung where the apex of the heart is located.
cardiac output (CO) – volume of blood pumped by each ventricle during one minute; equals HR multiplied by SV.
cardiac plexus – paired complex network of nerve fibres near the base of the heart that receive sympathetic and parasympathetic stimulations to regulate HR.
cardiac reflexes – series of autonomic reflexes that enable the cardiovascular centres to regulate heart function based upon sensory information from a variety of visceral sensors.
cardiac reserve – difference between maximum and resting CO.
cardiac skeleton – (also, skeleton of the heart) reinforced connective tissue located within the atrioventricular septum; includes four rings that surround the openings between the atria and ventricles, and the openings to the pulmonary trunk and aorta; the point of attachment for the heart valves.
cardiogenic area – area near the head of the embryo where the heart begins to develop 18–19 days after fertilisation.
cardiogenic cords – two strands of tissue that form within the cardiogenic area.
cardiogenic shock – type of shock that results from the inability of the heart to maintain cardiac output.
cardiomyocyte – muscle cell of the heart.
carotid canal – zig-zag tunnel providing passage through the base of the skull for the internal carotid artery to the brain; begins anteromedial to the styloid process and terminates in the middle cranial cavity, near the posterior-lateral base of the sella turcica.
carotid sinuses – small pockets near the base of the internal carotid arteries that are the locations of the baroreceptors and chemoreceptors that trigger a reflex that aids in the regulation of vascular homeostasis.
carpal bone – one of the eight small bones that form the wrist and base of the hand; these are grouped as a proximal row consisting of (from lateral to medial) the scaphoid, lunate, triquetrum, and pisiform bones, and a distal row containing (from lateral to medial) the trapezium, trapezoid, capitate, and hamate bones.
carpal tunnel – passageway between the anterior forearm and hand formed by the carpal bones and flexor retinaculum.
carpometacarpal joint – articulation between one of the carpal bones in the distal row and a metacarpal bone of the hand.
cartilage – semi-rigid connective tissue found on the skeleton in areas where flexibility and smooth surfaces support movement.
cartilaginous joint – joint at which the bones are united by hyaline cartilage (synchondrosis) or fibrocartilage (symphysis).
catagen – transitional phase marking the end of the anagen phase of the hair growth cycle.
cauda equina – bundle of spinal nerve roots that descend from the lower spinal cord below the first lumbar vertebra and lie within the vertebral cavity; has the appearance of a horse’s tail.
caudal – describes a position below or lower than another part of the body proper; near or toward the tail (in humans, the coccyx, or lowest part of the spinal column); also referred to as inferior.
caudate – nucleus deep in the cerebrum that is part of the basal nuclei; along with the putamen, it is part of the striatum.
cavernous sinus – enlarged vein that receives blood from most of the other cerebral veins and the eye socket, and leads to the petrosal sinus.
cecum – pouch forming the beginning of the large intestine.
cell – smallest independently functioning unit of all organisms; in animals, a cell contains cytoplasm, composed of fluid and organelles.
cell cycle – life cycle of a single cell, from its birth until its division into two new daughter cells.
cell junction – point of cell-to-cell contact that connects one cell to another in a tissue.
cell membrane – membrane surrounding all animal cells, composed of a lipid bilayer interspersed with various molecules; also known as plasma membrane.
cellular components (formed elements) – cellular components of blood; that is, erythrocytes, leukocytes, and platelets.
cellulose – polysaccharide that comprises the plants’ cell wall; provides structural support to the cell.
cementum – bone-like tissue covering the root of a tooth.
central canal – longitudinal channel in the centre of each osteon; contains blood vessels, nerves, and lymphatic vessels; also known as the Haversian canal.
central chemoreceptor – one of the specialised receptors that are located in the brain that sense changes in hydrogen ion, oxygen, or carbon dioxide concentrations in the brain.
central nervous system (CNS) – anatomical division of the nervous system located within the cranial and vertebral cavities, namely the brain and spinal cord.
central sulcus – surface landmark of the cerebral cortex that marks the boundary between the frontal and parietal lobes.
central tolerance – B cell tolerance induced in immature B cells of the bone marrow.
central vein – vein that receives blood from hepatic sinusoids.
centriole – small, self-replicating organelle that provides the origin for microtubule growth and moves DNA during cell division.
centromere – region of attachment for two sister chromatids.
centrosome – cellular structure that organises microtubules during cell division.
cephalic flexure – curve in midbrain of the embryo that positions the forebrain ventrally.
cephalic phase – (also, reflex phase) initial phase of gastric secretion that occurs before food enters the stomach.
cephalic vein – superficial vessel in the upper arm; leads to the axillary vein.
cerebellum – region of the adult brain connected primarily to the pons that developed from the metencephalon (along with the pons) and is largely responsible for comparing information from the cerebrum with sensory feedback from the periphery through the spinal cord.
cerebral aqueduct – connection of the ventricular system between the third and fourth ventricles located in the midbrain.
cerebral cortex – outermost layer of grey matter in the brain, where conscious perception takes place.
cerebral hemisphere – one half of the bilaterally symmetrical cerebrum.
cerebral peduncles – segments of the descending motor pathway that make up the white matter of the ventral midbrain.
cerebrospinal fluid (CSF) – circulatory medium within the CNS that is produced by ependymal cells in the choroid plexus filtering the blood.
cerebrovascular accident (CVA) – blockage of blood flow to the brain; also called a stroke.
cerebrum – region of the adult brain that develops from the telencephalon and is responsible for higher neurological functions such as memory, emotion, and consciousness.
cervical curve – posteriorly concave curvature of the cervical vertebral column region; a secondary curve of the vertebral column.
cervical enlargement – region of the ventral (anterior) horn of the spinal cord that has a larger population of motor neurons for the greater number of and finer control of muscles of the upper limb.
cervical plexus – nerve plexus associated with the upper cervical spinal nerves.
cervical vertebrae – seven vertebrae numbered as C1–C7 that are located in the neck region of the vertebral column.
cervix – elongate inferior end of the uterus where it connects to the vagina.
channel protein – membrane-spanning protein that has an inner pore which allows the passage of one or more substances.
chaperone – (also, chaperonin) protein that helps nascent protein in the folding process.
checkpoint – progress point in the cell cycle during which certain conditions must be met in order for the cell to proceed to a subsequence phase.
chemical digestion – enzymatic breakdown of food.
chemical synapse – connection between two neurons, or between a neuron and its target, where a neurotransmitter diffuses across a very short distance.
chemokine – soluble, long-range, cell-to-cell communication molecule.
chemoreceptor – sensory receptor cell that is sensitive to chemical stimuli, such as in taste, smell, or pain.
chief cell – gastric gland cell that secretes pepsinogen.
chief sensory nucleus – component of the trigeminal nuclei that is found in the pons.
chitin – type of carbohydrate that forms the outer skeleton of all arthropods that include crustaceans and insects; it also forms fungi cell walls.
chloride shift – facilitated diffusion that exchanges bicarbonate (HCO3–) with chloride (Cl–) ions.
cholinergic system – neurotransmitter system of acetylcholine, which includes its receptors and the enzyme acetylcholinesterase.
chondrocytes – cells of the cartilage.
chromatin – substance consisting of DNA and associated proteins.
chromosome – condensed version of chromatin.
chronic inflammation – inflammation occurring for long periods of time.
chordae tendineae – string-like extensions of tough connective tissue that extend from the flaps of the atrioventricular valves to the papillary muscles.
chorion – membrane that develops from the syncytiotrophoblast, cytotrophoblast, and mesoderm; surrounds the embryo and forms the foetal portion of the placenta through the chorionic villi.
chorionic membrane – precursor to the chorion; forms from extra-embryonic mesoderm cells.
chorionic villi – projections of the chorionic membrane that burrow into the endometrium and develop into the placenta.
choroid – highly vascular tissue in the wall of the eye that supplies the outer retina with blood/
choroid plexus – specialised structure containing ependymal cells that line blood capillaries and filter blood to produce CSF in the four ventricles of the brain.
chromaffin – neuroendocrine cells of the adrenal medulla.
chyle – lipid-rich lymph inside the lymphatic capillaries of the small intestine.
chylomicron – large lipid-transport compound made up of triglycerides, phospholipids, cholesterol, and proteins.
chyme – soupy liquid created when food is mixed with digestive secretions.
cilia – small appendage on certain cells formed by microtubules and modified for movement of materials across the cellular surface.
ciliary body – smooth muscle structure on the interior surface of the iris that controls the shape of the lens through the zonule fibres.
circadian rhythm – internal perception of the daily cycle of light and dark based on retinal activity related to sunlight.
circle of Willis – (also, arterial circle) anastomosis located at the base of the brain that ensures continual blood supply; formed from branches of the internal carotid and vertebral arteries; supplies blood to the brain.
circular folds – (also, plicae circulares) deep fold in the mucosa and submucosa of the small intestine.
circulatory shock – also simply called shock; a life-threatening medical condition in which the circulatory system is unable to supply enough blood flow to provide adequate oxygen and other nutrients to the tissues to maintain cellular metabolism.
circumduction – circular motion of the arm, thigh, hand, thumb, or finger that is produced by the sequential combination of flexion, abduction, extension, and adduction.
circumflex artery – branch of the left coronary artery that follows coronary sulcus.
cisterna chyli – bag-like vessel that forms the beginning of the thoracic duct.
class switching – ability of B cells to change the class of antibody they produce without altering the specificity for antigen.
clavicle – collarbone; elongated bone that articulates with the manubrium of the sternum medially and the acromion of the scapula laterally.
clavicular notch – paired notches located on the superior-lateral sides of the sternal manubrium, for articulation with the clavicle.
cleavage – form of mitotic cell division in which the cell divides but the total volume remains unchanged; this process serves to produce smaller and smaller cells.
cleavage furrow – contractile ring that forms around a cell during cytokinesis that pinches the cell into two halves.
clitoris – (also, glans clitoris) nerve-rich area of the vulva that contributes to sexual sensation during intercourse.
clonal anergy – process whereby B cells that react to soluble antigens in bone marrow are made non-functional.
clonal deletion – removal of self-reactive B cells by inducing apoptosis.
clonal expansion – growth of a clone of selected lymphocytes.
clonal selection – stimulating growth of lymphocytes that have specific receptors.
clone – group of lymphocytes sharing the same antigen receptor.
closed reduction – manual manipulation of a broken bone to set it into its natural position without surgery.
clotting – also called coagulation; complex process by which blood components form a plug to stop bleeding.
clotting factors – group of 12 identified substances active in coagulation.
coagulation – formation of a blood clot; part of the process of haemostasis.
coccyx – small bone located at inferior end of the adult vertebral column that is formed by the fusion of four coccygeal vertebrae; also referred to as the “tailbone”.
cochlea – auditory portion of the inner ear containing structures to transduce sound stimuli.
cochlear duct – space within the auditory portion of the inner ear that contains the organ of Corti and is adjacent to the scala tympani and scala vestibuli on either side.
codon – consecutive sequence of three nucleotides on an mRNA molecule that corresponds to a specific amino acid.
coeliac trunk – (also, coeliac artery) major branch of the abdominal aorta; gives rise to the left gastric artery, the splenic artery, and the common hepatic artery that forms the hepatic artery to the liver, the right gastric artery to the stomach, and the cystic artery to the gall bladder.
collagen fibres – flexible fibrous proteins that give connective tissue tensile strength.
colloid – viscous fluid in the central cavity of thyroid follicles, containing the glycoprotein thyroglobulin.
colon – part of the large intestine between the cecum and the rectum.
colony-stimulating factors (CSFs) – glycoproteins that trigger the proliferation and differentiation of myeloblasts into granular leukocytes (basophils, neutrophils, and eosinophils).
common bile duct – structure formed by the union of the common hepatic duct and the gallbladder’s cystic duct.
common carotid artery – right common carotid artery arises from the brachiocephalic artery, and the left common carotid arises from the aortic arch; gives rise to the external and internal carotid arteries; supplies the respective sides of the head and neck.
common hepatic artery – branch of the celiac trunk that forms the hepatic artery, the right gastric artery, and the cystic artery.
common hepatic duct – duct formed by the merger of the two hepatic ducts.
common iliac artery – branch of the aorta that leads to the internal and external iliac arteries.
common iliac vein – one of a pair of veins that flows into the inferior vena cava at the level of L5; the left common iliac vein drains the sacral region; divides into external and internal iliac veins near the inferior portion of the sacroiliac joint
common pathway – final coagulation pathway activated either by the intrinsic or the extrinsic pathway, and ending in the formation of a blood clot.
compact bone – dense osseous tissue that can withstand compressive forces.
complement – enzymatic cascade of constitutive blood proteins that have antipathogen effects, including the direct killing of bacteria.
compliance – degree to which a blood vessel can stretch as opposed to being rigid.
concentration gradient – difference in the concentration of a substance between two regions.
concentric contraction – muscle contraction that shortens the muscle to move a load.
conceptus – pre-implantation stage of a fertilised egg and its associated membran.
conducting zone – region of the respiratory system that includes the organs and structures that provide passageways for air and are not directly involved in gas exchange.
condylar process of the mandible – thickened upward projection from posterior margin of mandibular ramus.
condyle – oval-shaped process located at the top of the condylar process of the mandible.
condyloid joint – synovial joint in which the shallow depression at the end of one bone receives a rounded end from a second bone or a rounded structure formed by two bones; found at the metacarpophalangeal joints of the fingers or the radiocarpal joint of the wrist; functionally classified as a biaxial joint.
cone photoreceptor – one of the two types of retinal receptor cell that is specialised for colour vision through the use of three photopigments distributed through three separate populations of cells.
connective tissue – type of tissue that serves to hold in place, connect, and integrate the body’s organs and systems.
connective tissue membrane – connective tissue that encapsulates organs and lines movable joints.
connective tissue proper – connective tissue containing a viscous matrix, fibres, and cells.
constant region domain – part of a lymphocyte antigen receptor that does not vary much between different receptor types.
continuous capillary – most common type of capillary, found in virtually all tissues except epithelia and cartilage; contains very small gaps in the endothelial lining that permit exchange.
continuous conduction – slow propagation of an action potential along an unmyelinated axon owing to voltage-gated Na+ channels located along the entire length of the cell membrane.
contractility – ability to shorten (contract) forcibly.
contraction phase – twitch contraction phase when tension increases.
contralateral – word meaning “on the opposite side,” as in axons that cross the midline in a fibre tract.
control centre – compares values to their normal range; deviations cause the activation of an effector.
coracoclavicular ligament – strong band of connective tissue that anchors the coracoid process of the scapula to the lateral clavicle; provides important indirect support for the acromioclavicular joint.
coracohumeral ligament – intrinsic ligament of the shoulder joint; runs from the coracoid process of the scapula to the anterior humerus.
coracoid process – short, hook-like process that projects anteriorly and laterally from the superior margin of the scapula.
corn – type of callus that is named for its shape and the elliptical motion of the abrasive force.
cornea – fibrous covering of the anterior region of the eye that is transparent so that light can pass through it.
corneal reflex – protective response to stimulation of the cornea causing contraction of the orbicularis oculi muscle resulting in blinking of the eye.
corona radiata – in an oocyte, a layer of granulosa cells that surrounds the oocyte and that must be penetrated by sperm before fertilisation can occur.
coronal suture – joint that unites the frontal bone to the right and left parietal bones across the top of the skull.
coronary arteries – branches of the ascending aorta that supply blood to the heart; the left coronary artery feeds the left side of the heart, the left atrium and ventricle, and the interventricular septum; the right coronary artery feeds the right atrium, portions of both ventricles, and the heart conduction system.
coronary sinus – large, thin-walled vein on the posterior surface of the heart that lies within the atrioventricular sulcus and drains the heart myocardium directly into the right atrium.
coronary sulcus – sulcus that marks the boundary between the atria and ventricles.
coronary veins – vessels that drain the heart and generally parallel the large surface arteries.
coronoid fossa – depression on the anterior surface of the humerus above the trochlea; this space receives the coronoid process of the ulna when the elbow is maximally flexed.
coronoid process of the mandible – flattened upward projection from the anterior margin of the mandibular ramus.
coronoid process of the ulna – projecting bony lip located on the anterior, proximal ulna; forms the inferior margin of the trochlear notch.
corpus albicans – non-functional structure remaining in the ovarian stroma following structural and functional regression of the corpus luteum.
corpus callosum – large white matter structure that connects the right and left cerebral hemispheres.
corpus cavernosum – either of two columns of erectile tissue in the penis that fill with blood during an erection.
corpus luteum – transformed follicle after ovulation that secretes progesterone.
corpus spongiosum – (plural = corpora cavernosa) column of erectile tissue in the penis that fills with blood during an erection and surrounds the penile urethra on the ventral portion of the penis.
cortical nephrons – nephrons with loops of Henle that do not extend into the renal medulla.
cortical reaction – following fertilisation, the release of cortical granules from the oocyte’s plasma membrane into the zona pellucida creating a fertilisation membrane that prevents any further attachment or penetration of sperm; part of the slow block to polyspermy.
cortex – in hair, the second or middle layer of keratinocytes originating from the hair matrix, as seen in a cross-section of the hair bulb.
corticobulbar tract – connection between the cortex and the brain stem responsible for generating movement.
corticospinal tract – connection between the cortex and the spinal cord responsible for generating movement.
cortisol – glucocorticoid important in gluconeogenesis, the catabolism of glycogen, and downregulation of the immune system.
costal cartilage – hyaline cartilage structure attached to the anterior end of each rib that provides for either direct or indirect attachment of most ribs to the sternum.
costal facet – site on the lateral sides of a thoracic vertebra for articulation with the head of a rib.
costal groove – shallow groove along the inferior margin of a rib that provides passage for blood vessels and a nerve.
costoclavicular ligament – band of connective tissue that unites the medial clavicle with the first rib.
coxal bone – hip bone.
cranial – describes a position above or higher than another part of the body proper; also referred to as superior.
cranial cavity – division of the posterior (dorsal) cavity that houses the brain.
cranial nerve – one of twelve nerves connected to the brain that are responsible for sensory or motor functions of the head and neck.
cranial nerve ganglion – sensory ganglion of cranial nerves.
cranium – skull.
creatine phosphate – phosphagen used to store energy from ATP and transfer it to muscle.
cribriform plate – small, flattened areas with numerous small openings, located to either side of the midline in the floor of the anterior cranial fossa; formed by the ethmoid bone.
cricoid cartilage – portion of the larynx composed of a ring of cartilage with a wide posterior region and a thinner anterior region; attached to the oesophagus.
crista galli – small upward projection located at the midline in the floor of the anterior cranial fossa; formed by the ethmoid bone.
cross-matching – blood test for determination of compatibility between donor RBC and patient’s plasma.
counter current multiplier system – involves the descending and ascending loops of Henle directing forming urine in opposing directions to create a concentration gradient when combined with variable permeability and sodium pumping.
crown – portion of tooth visible superior to the gum line.
cuboid – tarsal bone that articulates posteriorly with the calcaneus bone, medially with the lateral cuneiform bone, and anteriorly with the fourth and fifth metatarsal bones.
cupula – specialised structure within the base of a semicircular canal that bends the stereocilia of hair cells when the head rotates by way of the relative movement of the enclosed fluid.
cuspid – (also, canine) pointed tooth used for tearing and shredding food.
cutaneous membrane – skin; epithelial tissue made up of a stratified squamous epithelial cells that cover the outside of the body.
cuticle – in hair, the outermost layer of keratinocytes originating from the hair matrix, as seen in a cross-section of the hair bulb.
cyclic adenosine monophosphate (cAMP) – second messenger that, in response to adenylyl cyclase activation, triggers a phosphorylation cascade.
cyclin – one of a group of proteins that function in the progression of the cell cycle.
cyclin-dependent kinase (CDK) – one of a group of enzymes associated with cyclins that help them perform their functions.
cystic artery – branch of the common hepatic artery; supplies blood to the gall bladder.
cystic duct – duct through which bile drains and enters the gallbladder.
cytokine – soluble, short-range, cell-to-cell communication molecule.
cytokines – class of proteins that act as autocrine or paracrine signalling molecules; in the cardiovascular system, they stimulate the proliferation of progenitor cells and help to stimulate both nonspecific and specific resistance to disease.
cytokinesis – final stage in cell division, where the cytoplasm divides to form two separate daughter cells.
cytoplasm – internal material between the cell membrane and nucleus of a cell, mainly consisting of a water-based fluid called cytosol, within which are all the other organelles and cellular solute and suspended materials.
cytoskeleton – “skeleton” of a cell; formed by rod-like proteins that support the cell’s shape and provide, among other functions, locomotive abilities.
cytosol – clear, semi-fluid medium of the cytoplasm, made up mostly of water.
cytotoxic T cells (Tc) – T lymphocytes with the ability to induce apoptosis in target cells.
D
Dalton’s law – statement of the principle that a specific gas type in a mixture exerts its own pressure, as if that specific gas type was not part of a mixture of gases.
deciduous tooth – one of 20 “baby teeth”.
decussate – to cross the midline, as in fibres that project from one side of the body to the other.
deep – describes a position farther from the surface of the body.
deep femoral artery – branch of the femoral artery; gives rise to the lateral circumflex arteries.
deep femoral vein – drains blood from the deeper portions of the thigh and leads to the femoral vein.
defaecation – elimination of undigested substances from the body in the form of faeces.
defensins – antimicrobial proteins released from neutrophils and macrophages that create openings in the plasma membranes to kill cells.
deglutition – three-stage process of swallowing.
dehydration – state of containing insufficient water in blood and other tissues.
dehydration synthesis – (also, condensation) reaction that links monomer molecules, releasing a water molecule for each bond formed..
delayed hypersensitivity – (type IV) T cell-mediated immune response against pathogens infiltrating interstitial tissues, causing cellular infiltrate.
delta cell – minor cell type in the pancreas that secretes the hormone somatostatin.
deltoid ligament – broad intrinsic ligament located on the medial side of the ankle joint; supports the talus at the talocrural joint and resists excess eversion of the foot.
deltoid tuberosity – roughened, V-shaped region located laterally on the mid-shaft of the humerus.
denaturation – loss of shape in a protein as a result of changes in temperature, pH, or chemical exposure.
dendrite – one of many branchlike processes that extends from the neuron cell body and functions as a contact for incoming signals (synapses) from other neurons or sensory cells.
dens – bony projection (odontoid process) that extends upward from the body of the C2 (axis) vertebra.
dens – tooth.
dense body – sarcoplasmic structure that attaches to the sarcolemma and shortens the muscle as thin filaments slide past thick filaments.
dense connective tissue – connective tissue proper that contains many fibres that provide both elasticity and protection.
dentin – bone-like tissue immediately deep to the enamel of the crown or cementum of the root of a tooth.
dentition – set of teeth.
deoxyhemoglobin – molecule of haemoglobin without an oxygen molecule bound to it.
deoxyribonuclease – pancreatic enzyme that digests DNA.
deoxyribonucleic acid (DNA) – double-helical molecule that carries the cell’s hereditary information.
depolarise – to reduce the voltage difference between the inside and outside of a cell’s plasma membrane (the sarcolemma for a muscle fibre), making the inside less negative than at rest.
depolarisation – change in a cell membrane potential from rest toward zero.
depression – downward (inferior) motion of the scapula or mandible.
dermal papilla – (plural = dermal papillae) extension of the papillary layer of the dermis that increases surface contact between the epidermis and dermis.
dermis – layer of skin between the epidermis and hypodermis, composed mainly of connective tissue and containing blood vessels, hair follicles, sweat glands, and other structures.
descending aorta – portion of the aorta that continues downward past the end of the aortic arch; subdivided into the thoracic aorta and the abdominal aorta.
descending colon – part of the colon between the transverse colon and the sigmoid colon.
descending tract – central nervous system fibres carrying motor commands from the brain to the spinal cord or periphery.
desmosome – structure that forms an impermeable junction between cells.
detrusor muscle – smooth muscle in the bladder wall; fibres run in all directions to reduce the size of the organ when emptying it of urine.
diacylglycerol (DAG) – molecule that, like cAMP, activates protein kinases, thereby initiating a phosphorylation cascade.
diapedesis – (also, emigration) process by which leukocytes squeeze through adjacent cells in a blood vessel wall to enter tissues.
diaphysis – tubular shaft that runs between the proximal and distal ends of a long bone.
diarthrosis – freely mobile joint.
diastole – period of time when the heart muscle is relaxed and the chambers fill with blood.
diastolic pressure – lower number recorded when measuring arterial blood pressure; represents the minimal value corresponding to the pressure that remains during ventricular relaxation.
diabetes mellitus – condition caused by destruction or dysfunction of the beta cells of the pancreas or cellular resistance to insulin that results in abnormally high blood glucose levels.
diencephalon – region of the adult brain that retains its name from embryonic development and includes the thalamus and hypothalamus.
diffusion – movement of a substance from an area of higher concentration to one of lower concentration.
digital arteries – formed from the superficial and deep palmar arches; supply blood to the digits.
digital veins – drain the digits and feed into the palmar arches of the hand and dorsal venous arch of the foot.
dihydroxyvitamin D – active form of vitamin D required by the intestinal epithelial cells for the absorption of calcium.
dipeptidase – brush border enzyme that acts on proteins.
diploë – layer of spongy bone, that is sandwiched between two the layers of compact bone found in flat bones.
diploid – condition marked by the presence of a double complement of genetic material (two sets of chromosomes, one set inherited from each of two parents).
direct pathway – connections within the basal nuclei from the striatum to the globus pallidus internal segment and substantia nigra pars reticulata that disinhibit the thalamus to increase cortical control of movement.
disaccharide – two sugar monomers that a glycosidic bond links.
disinhibition – disynaptic connection in which the first synapse inhibits the second cell, which then stops inhibiting the final target.
distal – describes a position farther from the point of attachment or the trunk of the body.
distal convoluted tubules – portions of the nephron distal to the loop of Henle that receive hyposmotic filtrate from the loop of Henle and empty into collecting ducts.
distal radioulnar joint – articulation between the head of the ulna and the ulnar notch of the radius.
distal tibiofibular joint – articulation between the distal fibula and the fibular notch of the tibia.
diuresis – excess production of urine.
diuretic – compound that increases urine output, leading to decreased water conservation.
DNA polymerase – enzyme that functions in adding new nucleotides to a growing strand of DNA during DNA replication.
DNA replication – process of duplicating a molecule of DNA.
dorsal – describes the back or direction toward the back of the body; also referred to as posterior.
dorsal arch – (also, arcuate arch) formed from the anastomosis of the dorsalis pedis artery and medial and plantar arteries; branches supply the distal portions of the foot and digits.
dorsal cavity – posterior body cavity that houses the brain and spinal cord; also referred to the posterior body cavity.
dorsal column system – ascending tract of the spinal cord associated with fine touch and proprioceptive sensations.
dorsal (posterior) nerve root – axons entering the posterior horn of the spinal cord.
dorsal respiratory group (DRG) – region of the medulla oblongata that stimulates the contraction of the diaphragm and intercostal muscles to induce inspiration.
dorsal (posterior) root ganglion – sensory ganglion attached to the posterior nerve root of a spinal nerve.
dorsal stream – connections between cortical areas from the occipital to parietal lobes that are responsible for the perception of visual motion and guiding movement of the body in relation to that motion.
dorsalis pedis artery – forms from the anterior tibial artery; branches repeatedly to supply blood to the tarsal and dorsal regions of the foot.
dorsal venous arch – drains blood from digital veins and vessels on the superior surface of the foot.
dorsiflexion – movement at the ankle that brings the top of the foot toward the anterior leg.
dorsum nasi – intermediate portion of the external nose that connects the bridge to the apex and is supported by the nasal bone.
downregulation – decrease in the number of hormone receptors, typically in response to chronically excessive levels of a hormone.
ductus arteriosus – shunt in the foetal pulmonary trunk that diverts oxygenated blood back to the aorta.
ductus deferens – (also, vas deferens) duct that transports sperm from the epididymis through the spermatic cord and into the ejaculatory duct; also referred as the vas deferens.
ductus venosus – shunt that causes oxygenated blood to bypass the foetal liver on its way to the inferior vena cava.
duodenal gland – (also, Brunner’s gland) mucous-secreting gland in the duodenal submucosa.
duodenum – first part of the small intestine, which starts at the pyloric sphincter and ends at the jejunum.
dura mater – tough, fibrous, outer layer of the meninges that is attached to the inner surface of the cranium and vertebral column and surrounds the entire CNS.
dural sinus – any of the venous structures surrounding the brain, enclosed within the dura mater, which drain blood from the CNS to the common venous return of the jugular veins.
E
early induced immune response – includes antimicrobial proteins stimulated during the first several days of an infection.
ear ossicles – three small bones located in the middle ear cavity that serve to transmit sound vibrations to the inner ear.
eccentric contraction – muscle contraction that lengthens the muscle as the tension is diminished.
eccrine sweat gland – type of sweat gland that is common throughout the skin surface; it produces a hypotonic sweat for thermoregulation.
ectoderm – outermost embryonic germ layer from which the epidermis and the nervous tissue derive.
ectopic pregnancy – implantation of an embryo outside of the uterus.
eczema – skin condition due to an allergic reaction, which resembles a rash.
effector – organ that can cause a change in a value.
efferent lymphatic vessels – lead out of a lymph node.
effector protein – enzyme that catalyses the generation of a new molecule, which acts as the intracellular mediator of the signal that binds to the receptor.
effector T cells – immune cells with a direct, adverse effect on a pathogen.
efferent arteriole – arteriole carrying blood from the glomerulus to the capillary beds around the convoluted tubules and loop of Henle; portion of the portal system.
ejaculatory duct – duct that connects the ampulla of the ductus deferens with the duct of the seminal vesicle at the prostatic urethra.
ejection fraction – portion of the blood that is pumped or ejected from the heart with each contraction; mathematically represented by SV divided by EDV.
elastic artery – (also, conducting artery) artery with abundant elastic fibres located closer to the heart, which maintains the pressure gradient and conducts blood to smaller branches.
elastic cartilage – type of cartilage, with elastin as the major protein, characterised by rigid support as well as elasticity
elastic fibres – fibrous protein within connective tissue that contains a high percentage of the protein elastin that allows the fibres to stretch and return to original size.
elasticity – ability to stretch and rebound.
elbow joint – joint located between the upper arm and forearm regions of the upper limb; formed by the articulations between the trochlea of the humerus and the trochlear notch of the ulna, and the capitulum of the humerus and the head of the radius.
electrical gradient – difference in the electrical charge (potential) between two regions.
electrical synapse – connection between two neurons, or any two electrically active cells, where ions flow directly through channels spanning their adjacent cell membranes.
electrocardiogram (ECG) – surface recording of the electrical activity of the heart that can be used for diagnosis of irregular heart function; also abbreviated as EKG.
electrochemical exclusion – principle of selectively allowing ions through a channel on the basis of their charge.
eleidin – clear protein-bound lipid found in the stratum lucidum that is derived from keratohyalin and helps to prevent water loss.
elevation – upward (superior) motion of the scapula or mandible.
embolus – thrombus that has broken free from the blood vessel wall and entered the circulation.
emigration – (also, diapedesis) process by which leukocytes squeeze through adjacent cells in a blood vessel wall to enter tissues.
embryo – developing human during weeks 3–8.
embryonic folding – process by which an embryo develops from a flat disc of cells to a three-dimensional shape resembling a cylinder.
enamel – covering of the dentin of the crown of a tooth.
encapsulated ending – configuration of a sensory receptor neuron with dendrites surrounded by specialised structures to aid in transduction of a particular type of sensation, such as the lamellated corpuscles in the deep dermis and subcutaneous tissue.
end diastolic volume (EDV) – (also, preload) the amount of blood in the ventricles at the end of atrial systole just prior to ventricular contraction.
endocardial tubes – stage in which lumens form within the expanding cardiogenic cords, forming hollow structures.
endocardium – innermost layer of the heart lining the heart chambers and heart valves; composed of endothelium reinforced with a thin layer of connective tissue that binds to the myocardium.
endochondral ossification – process in which bone forms by replacing hyaline cartilage.
endocrine gland – groups of cells that release chemical signals into the intercellular fluid to be picked up and transported to their target organs by blood.
endocrine system – cells, tissues, and organs that secrete hormones as a primary or secondary function and play an integral role in normal bodily processes.
endocytosis – import of material into the cell by formation of a membrane-bound vesicle.
endoderm – innermost embryonic germ layer from which most of the digestive system and lower respiratory system derive.
endometrium – inner lining of the uterus, part of which builds up during the secretory phase of the menstrual cycle and then sheds with menses.
endomysium – loose, and well-hydrated connective tissue covering each muscle fibre in a skeletal muscle.
endoneurium – innermost layer of connective tissue that surrounds individual axons within a nerve.
endoplasmic reticulum (ER) – cellular organelle that consists of interconnected membrane-bound tubules, which may or may not be associated with ribosomes (rough type or smooth type, respectively).
endosteum – delicate membranous lining of a bone’s medullary cavity.
end systolic volume (ESV) – amount of blood remaining in each ventricle following systole.
endothelins – group of vasoconstrictive, 21-amino acid peptides; produced by endothelial cells of the renal blood vessels, mesangial cells, and cells of the DCT.
endothelium – tissue that lines vessels of the lymphatic and cardiovascular system, made up of a simple squamous epithelium.
enteric nervous system (ENS) – neural tissue associated with the digestive system that is responsible for nervous control through autonomic connections.
enteric plexus – neuronal plexus in the wall of the intestines, which is part of the enteric nervous system.
enteroendocrine cell – gastric gland cell that releases hormones.
enterohepatic circulation – recycling mechanism that conserves bile salts.
enteropeptidase – intestinal brush-border enzyme that activates trypsinogen to trypsin.
enzyme – catalyst in a biochemical reaction that is usually a complex or conjugated protein.
eosinophils – granulocytes that stain with eosin; they release antihistamines and are especially active against parasitic worms.
ependymal cell – glial cell type in the CNS responsible for producing cerebrospinal fluid.
epiblast – upper layer of cells of the embryonic disc that forms from the inner cell mass; gives rise to all three germ layers.
epicardial coronary arteries – surface arteries of the heart that generally follow the sulci.
epicardium – innermost layer of the serous pericardium and the outermost layer of the heart wall.
epidermis – outermost tissue layer of the skin.
epididymis – (plural = epididymides) coiled tubular structure in which sperm start to mature and are stored until ejaculation.
epiglottis – leaf-shaped piece of elastic cartilage that is a portion of the larynx that swings to close the trachea during swallowing.
epimysium – outer layer of connective tissue around a skeletal muscle.
epineurium – outermost layer of connective tissue that surrounds an entire nerve.
epiphyseal line – completely ossified remnant of the epiphyseal plate.
epiphyseal plate – (also, growth plate) sheet of hyaline cartilage in the metaphysis of an immature bone; replaced by bone tissue as the organ grows in length.
epiphysis – wide section at each end of a long bone; filled with spongy bone and red marrow.
epiploic appendage – small sac of fat-filled visceral peritoneum attached to teniae coli.
epithalamus – region of the diecephalon containing the pineal gland.
epithelial membrane – epithelium attached to a layer of connective tissue.
epithelial tissue – type of tissue that serves primarily as a covering or lining of body parts, protecting the body; it also functions in absorption, transport, and secretion.
eponychium – nail fold that meets the proximal end of the nail body, also called the cuticle.
epsilon cell – minor cell type in the pancreas that secretes the hormone ghrelin.
equilibrium – sense of balance that includes sensations of position and movement of the head.
erythroblastosis fetalis – disease of Rh factor-positive newborns in Rh-negative mothers with multiple Rh-positive children; resulting from the action of maternal antibodies against foetal blood.
erythrocyte – (also, red blood cell) mature myeloid blood cell that is composed mostly of haemoglobin and functions primarily in the transportation of oxygen and carbon dioxide.
erythropoietin (EPO) – glycoprotein that triggers the bone marrow to produce haemoglobin and RBCs; secreted by the kidney in response to low oxygen levels.
ethmoid air cell – one of several small, air-filled spaces located within the lateral sides of the ethmoid bone, between the orbit and upper nasal cavity.
ethmoid bone – unpaired bone that forms the roof and upper, lateral walls of the nasal cavity, portions of the floor of the anterior cranial fossa and medial wall of orbit, and the upper portion of the nasal septum.
eversion – foot movement involving the intertarsal joints of the foot in which the bottom of the foot is turned laterally, away from the midline.
excitability – ability to undergo neural stimulation.
excitable membrane – cell membrane that regulates the movement of ions so that an electrical signal can be generated.
excitation-contraction coupling – sequence of events from motor neuron signalling to a skeletal muscle fibre to contraction of the fibre’s sarcomeres.
excitatory postsynaptic potential (EPSP) – graded potential in the postsynaptic membrane that is the result of depolarisation and makes an action potential more likely to occur.
executive functions – cognitive processes of the prefrontal cortex that lead to directing goal-directed behaviour, which is a precursor to executing motor commands.
exocrine gland – group of epithelial cells that secrete substances through ducts that open to the skin or to internal body surfaces that lead to the exterior of the body.
exocrine system – cells, tissues, and organs that secrete substances directly to target tissues via glandular ducts.
exocytosis – export of a substance out of a cell by formation of a membrane-bound vesicle.
exon – one of the coding regions of an mRNA molecule that remain after splicing.
expiration – (also, exhalation) process that causes the air to leave the lungs.
expiratory reserve volume (ERV) – amount of air that can be forcefully exhaled after a normal tidal exhalation.
extensibility – ability to lengthen (extend).
extension – movement in the sagittal plane that increases the angle of a joint (straightens the joint); motion involving posterior bending of the vertebral column or returning to the upright position from a flexed position.
external acoustic meatus – ear canal opening located on the lateral side of the skull.
external anal sphincter – voluntary skeletal muscle sphincter in the anal canal.
external callus – collar of hyaline cartilage and bone that forms around the outside of a fracture.
external carotid artery – arises from the common carotid artery; supplies blood to numerous structures within the face, lower jaw, neck, oesophagus, and larynx.
external ear – structures on the lateral surface of the head, including the auricle and the ear canal back to the tympanic membrane.
external elastic membrane – membrane composed of elastic fibres that separates the tunica media from the tunica externa; seen in larger arteries.
external iliac artery – branch of the common iliac artery that leaves the body cavity and becomes a femoral artery; supplies blood to the lower limbs.
external iliac vein – formed when the femoral vein passes into the body cavity; drains the legs and leads to the common iliac vein.
external jugular vein – one of a pair of major veins located in the superficial neck region that drains blood from the more superficial portions of the head, scalp, and cranial regions, and leads to the subclavian vein.
external nose – region of the nose that is easily visible to others.
external occipital protuberance – small bump located at the midline on the posterior skull.
external respiration – gas exchange that occurs in the alveoli.
external root sheath – outer layer of the hair follicle that is an extension of the epidermis, which encloses the hair root.
external urinary sphincter – skeletal muscle; must be relaxed consciously to void urine.
exteroceptor – sensory receptor that is positioned to interpret stimuli from the external environment, such as photoreceptors in the eye or somatosensory receptors in the skin.
extracellular fluid (ECF) – fluid exterior to cells; includes the interstitial fluid, blood plasma, and fluid found in other reservoirs in the body.
extraocular muscles – six skeletal muscles that control eye movement within the orbit.
extrapyramidal system – pathways between the brain and spinal cord that are separate from the corticospinal tract and are responsible for modulating the movements generated through that primary pathway.
extrinsic ligament – ligament located outside of the articular capsule of a synovial joint.
extrinsic pathway – initial coagulation pathway that begins with tissue damage and results in the activation of the common pathway.
F
facet – small, flattened area on a bone for an articulation (joint) with another bone, or for muscle attachment.
facial bones – fourteen bones that support the facial structures and form the upper and lower jaws and the hard palate.
facial nerve – seventh cranial nerve; responsible for contraction of the facial muscles and for part of the sense of taste, as well as causing saliva production.
facilitated diffusion – diffusion of a substance with the aid of a membrane protein.
faeces – semisolid waste product of digestion.
false ribs – vertebrochondral ribs 8–12 whose costal cartilage either attaches indirectly to the sternum via the costal cartilage of the next higher rib or does not attach to the sternum at all.
fascicle – bundle of muscle fibres within a skeletal muscle.
fasciculus cuneatus – lateral division of the dorsal column system composed of fibres from sensory neurons in the upper body.
fasciculus gracilis – medial division of the dorsal column system composed of fibres from sensory neurons in the lower body.
fas ligand – molecule expressed on cytotoxic T cells and NK cells that binds to the fas molecule on a target cell and induces it do undergo apoptosis.
fast glycolytic (FG) – muscle fibre that primarily uses anaerobic glycolysis.
fast oxidative (FO) – intermediate muscle fibre that is between slow oxidative and fast glycolytic fibres.
fauces – portion of the posterior oral cavity that connects the oral cavity to the oropharynx.
Fc region – in an antibody molecule, the site where the two termini of the heavy chains come together; many cells have receptors for this portion of the antibody, adding functionality to these molecules.
femoral artery – continuation of the external iliac artery after it passes through the body cavity; divides into several smaller branches, the lateral deep femoral artery, and the genicular artery; becomes the popliteal artery as it passes posterior to the knee.
femoral circumflex vein – forms a loop around the femur just inferior to the trochanters; drains blood from the areas around the head and neck of the femur; leads to the femoral vein.
femoral nerve – systemic nerve of the anterior leg that arises from the lumbar plexus.
femoral vein – drains the upper leg; receives blood from the great saphenous vein, the deep femoral vein, and the femoral circumflex vein; becomes the external iliac vein when it crosses the body wall.
femoropatellar joint – portion of the knee joint consisting of the articulation between the distal femur and the patella.
femur – thigh bone; the single bone of the thigh.
fenestrated capillary – type of capillary with pores or fenestrations in the endothelium that allow for rapid passage of certain small materials.
fenestrations – small windows through a cell, allowing rapid filtration based on size; formed in such a way as to allow substances to cross through a cell without mixing with cell contents.
ferritin – protein-containing storage form of iron found in the bone marrow, liver, and spleen.
fertilisation – unification of genetic material from male and female haploid gametes.
fertilisation membrane – impenetrable barrier that coats a nascent zygote; part of the slow block to polyspermy.
fibrin – insoluble, filamentous protein that forms the structure of a blood clot.
fibrinogen – plasma protein produced in the liver and involved in blood clotting.
fibrinolysis – gradual degradation of a blood clot.
fibrocyte – less active form of fibroblast.
fibroelastic membrane – specialised membrane that connects the ends of the C-shape cartilage in the trachea; contains smooth muscle fibres.
fibrosis – replacement of muscle fibres by scar tissue.
fibrous joint – joint where the articulating areas of the adjacent bones are connected by fibrous connective tissue.
fibrous tunic – outer layer of the eye primarily composed of connective tissue known as the sclera and cornea.
fibula – thin, non-weight-bearing bone found on the lateral side of the leg.
fibular collateral ligament – extrinsic ligament of the knee joint that spans from the lateral epicondyle of the femur to the head of the fibula; resists hyperextension and rotation of the extended knee.
fibular nerve – systemic nerve of the posterior leg that begins as part of the sciatic nerve.
fibular notch – wide groove on the lateral side of the distal tibia for articulation with the fibula at the distal tibiofibular joint.
fibular vein – drains the muscles and integument near the fibula and leads to the popliteal vein.
filling time – duration of ventricular diastole during which filling occurs.
filtration – in the cardiovascular system, the movement of material from a capillary into the interstitial fluid, moving from an area of higher pressure to lower pressure.
fimbriae – finger-like projections on the distal uterine tubes.
first messenger – hormone that binds to a cell membrane hormone receptor and triggers activation of a second messenger system.
filtration slits – formed by pedicels of podocytes; substances filter between the pedicels based on size.
flagellum – appendage on certain cells formed by microtubules and modified for movement.
flat bone – thin and curved bone; serves as a point of attachment for muscles and protects internal organs.
flatus – gas in the intestine.
flexion – movement in the sagittal plane that decreases the angle of a joint (bends the joint); motion involving anterior bending of the vertebral column.
flexor retinaculum – strong band of connective tissue at the anterior wrist that spans the top of the U-shaped grouping of the carpal bones to form the roof of the carpal tunnel.
floating ribs – vertebral ribs 11–12 that do not attach to the sternum or to the costal cartilage of another rib.
fluid compartment – fluid inside all cells of the body constitutes a compartment system that is largely segregated from other systems.
fluid connective tissue – specialised cells that circulate in a watery fluid containing salts, nutrients, and dissolved proteins.
foetus – developing human during the time from the end of the embryonic period (week 9) to birth.
follicle – ovarian structure of one oocyte and surrounding granulosa (and later theca) cells.
follicle-stimulating hormone (FSH) – anterior pituitary hormone that stimulates the production and maturation of sex cells.
folliculogenesis – development of ovarian follicles from primordial to tertiary under the stimulation of gonadotropins.
fontanelle – expanded area of fibrous connective tissue that separates the brain-case bones of the skull prior to birth and during the first year after birth.
foot – portion of the lower limb located distal to the ankle joint.
foramen lacerum – irregular opening in the base of the skull, located inferior to the exit of carotid canal.
foramen magnum – large opening in the occipital bone of the skull through which the spinal cord emerges and the vertebral arteries enter the cranium.
foramen ovale – opening in the foetal heart that allows blood to flow directly from the right atrium to the left atrium, bypassing the foetal pulmonary circuit.
foramen ovale of the middle cranial fossa – oval-shaped opening in the floor of the middle cranial fossa.
foramen rotundum – round opening in the floor of the middle cranial fossa, located between the superior orbital fissure and foramen ovale.
foramen spinosum – small opening in the floor of the middle cranial fossa, located lateral to the foramen ovale.
forced breathing – (also, hyperpnoea) mode of breathing that occurs during exercise or by active thought that requires muscle contraction for both inspiration and expiration.
forearm – region of the upper limb located between the elbow and wrist joints; contains the radius and ulna bones.
forebrain – anterior region of the adult brain that develops from the prosencephalon and includes the cerebrum and diencephalon.
foregut – endoderm of the embryo towards the head region.
forming urine – filtrate undergoing modifications through secretion and reabsorption before true urine is produced.
fossa – (plural = fossae) shallow depression on the surface of a bone.
fossa ovalis – oval-shaped depression in the interatrial septum that marks the former location of the foramen ovale.
fourth ventricle – the portion of the ventricular system that is in the region of the brain stem and opens into the subarachnoid space through the median and lateral apertures.
fovea – exact centre of the retina at which visual stimuli are focused for maximal acuity, where the retina is thinnest, at which there is nothing but photoreceptors.
fovea capitis – minor indentation on the head of the femur that serves as the site of attachment for the ligament to the head of the femur.
fracture – broken bone.
fracture haematoma – blood clot that forms at the site of a broken bone.
Frank-Starling mechanism – relationship between ventricular stretch and contraction in which the force of heart contraction is directly proportional to the initial length of the muscle fibre.
free nerve ending – configuration of a sensory receptor neuron with dendrites in the connective tissue of the organ, such as in the dermis of the skin, that are most often sensitive to chemical, thermal, and mechanical stimuli.
frontal bone – unpaired bone that forms forehead, roof of orbit, and floor of anterior cranial fossa.
frontal eye field – region of the frontal lobe associated with motor commands to orient the eyes toward an object of visual attention.
frontal lobe – region of the cerebral cortex directly beneath the frontal bone of the cranium.
frontal plane – two-dimensional, vertical plane that divides the body or organ into anterior and posterior portions.
frontal sinus – air-filled space within the frontal bone; most anterior of the paranasal sinuses.
full thickness burns – burn that penetrates and destroys the full thickness of the skin (epidermis and dermis).
functional residual capacity (FRC) – sum of ERV and RV, which is the amount of air that remains in the lungs after a tidal expiration.
fundus – dome-shaped region of the stomach above and to the left of the cardia.
fundus – (of the uterus) domed portion of the uterus that is superior to the uterine tubes.
G
G cell – gastrin-secreting enteroendocrine cell.
gallbladder – accessory digestive organ that stores and concentrates bile.
gamete – haploid reproductive cell that contributes genetic material to form an offspring.
gamma cell (PP cell) – minor cell type in the pancreas that secretes the hormone pancreatic polypeptide.
ganglion – localised collection of neuron cell bodies in the peripheral nervous system.
gap junction – allows cytoplasmic communications to occur between cells.
gastric emptying – process by which mixing waves gradually cause the release of chyme into the duodenum.
gastric gland – gland in the stomach mucosal epithelium that produces gastric juice.
gastric phase – phase of gastric secretion that begins when food enters the stomach.
gastric pit – narrow channel formed by the epithelial lining of the stomach mucosa.
gastric plexuses – neuronal networks in the wall of the stomach that are part of the enteric nervous system.
gastrin – peptide hormone that stimulates secretion of hydrochloric acid and gut motility.
gastrocolic reflex – propulsive movement in the colon activated by the presence of food in the stomach.
gastroileal reflex – long reflex that increases the strength of segmentation in the ileum.
gastrointestinal microbiota – collection of microorganisms in the gastrointestinal tract.
gastrointestinal tract (alimentary canal) – continuous muscular digestive tube that extends from the mouth to the anus.
gastrulation – process of cell migration and differentiation into three primary germ layers following cleavage and implantation.
gated – property of a channel that determines how it opens under specific conditions, such as voltage change or physical deformation.
glassy membrane – layer of connective tissue that surrounds the base of the hair follicle, connecting it to the dermis.
gene – functional length of DNA that provides the genetic information necessary to build a protein.
gene expression – active interpretation of the information coded in a gene to produce a functional gene product.
general adaptation syndrome (GAS) – the human body’s three-stage response pattern to short- and long-term stress.
general sense – any sensory system that is distributed throughout the body and incorporated into organs of multiple other systems, such as the walls of the digestive organs or the skin.
generator potential – graded potential from dendrites of a unipolar cell which generates the action potential in the initial segment of that cell’s axon.
genicular artery – branch of the femoral artery; supplies blood to the region of the knee.
genome – entire complement of an organism’s DNA; found within virtually every cell.
germinal centres – clusters of rapidly proliferating B cells found in secondary lymphoid tissues.
gestation – in human development, the period required for embryonic and foetal development in utero; pregnancy.
gigantism – disorder in children caused when abnormally high levels of GH prompt excessive growth.
gingiva – gum.
glabella – slight depression of frontal bone, located at the midline between the eyebrows.
glans penis – bulbous end of the penis that contains many nerve endings.
glenohumeral joint – shoulder joint; formed by the articulation between the glenoid cavity of the scapula and the head of the humerus.
glenohumeral ligament – one of the three intrinsic ligaments of the shoulder joint that strengthen the anterior articular capsule.
glenoid cavity – (also, glenoid fossa) shallow depression located on the lateral scapula, between the superior and lateral borders.
glenoid labrum – lip of fibrocartilage located around the outside margin of the glenoid cavity of the scapula.
glial cell – one of the various types of neural tissue cells responsible for maintenance of the tissue, and for supporting neurons.
globin – haem-containing globular protein that is a constituent of haemoglobin.
globulins – heterogeneous group of plasma proteins that includes transport proteins, clotting factors, immune proteins, and others.
globus pallidus – nuclei deep in the cerebrum that are part of the basal nuclei and can be divided into the internal and external segments.
glomerular filtration rate (GFR) – rate of renal filtration.
glomerulus – tuft of capillaries surrounded by Bowman’s capsule; filters the blood based on size.
glossopharyngeal nerve – ninth cranial nerve; responsible for contraction of muscles in the tongue and throat and for part of the sense of taste, as well as causing saliva production.
glottis – opening between the vocal folds through which air passes when producing speech.
glucagon – pancreatic hormone that stimulates the catabolism of glycogen to glucose, thereby increasing blood glucose levels.
glucocorticoids – hormones produced by the zona fasciculata of the adrenal cortex that influence glucose metabolism.
gluteal tuberosity – roughened area on the posterior side of the proximal femur, extending inferiorly from the base of the greater trochanter.
glycocalyx – coating of sugar molecules that surrounds the cell membrane.
glycogen – storage carbohydrate in animals.
glycolysis – anaerobic breakdown of glucose to ATP.
glycosidic bond – bond formed by a dehydration reaction between two monosaccharides with eliminating a water molecule.
glycoprotein – protein that has one or more carbohydrates attached.
glycosuria – presence of glucose in the urine; caused by high blood glucose levels that exceed the ability of the kidneys to reabsorb the glucose; usually the result of untreated or poorly controlled diabetes mellitus.
goblet cell – unicellular gland found in columnar epithelium that secretes mucous.
goitre – enlargement of the thyroid gland either as a result of iodine deficiency or hyperthyroidism.
Golgi apparatus – cellular organelle formed by a series of flattened, membrane-bound sacs that functions in protein modification, tagging, packaging, and transport.
gomphosis – type of fibrous joint in which the root of a tooth is anchored into its bony jaw socket by strong periodontal ligaments.
gonadal artery – branch of the abdominal aorta; supplies blood to the gonads or reproductive organs; also described as ovarian arteries or testicular arteries, depending upon the sex of the individual.
gonadal vein – generic term for a vein draining a reproductive organ; may be either an ovarian vein or a testicular vein, depending on the sex of the individual.
gonadotropins – hormones that regulate the function of the gonads.
gonadotropin-releasing hormone (GnRH) – hormone released by the hypothalamus that regulates the production of follicle-stimulating hormone and luteinizing hormone from the pituitary gland.
gonads – reproductive organs (testes in men and ovaries in women) that produce gametes and reproductive hormones.
G protein – guanosine triphosphate (GTP) hydrolase that physically moves from the receptor protein to the effector protein to activate the latter.
graded muscle response – modification of contraction strength.
graded potential – change in the membrane potential that varies in size, depending on the size of the stimulus that elicits it.
graft-versus-host disease – in bone marrow transplants; occurs when the transplanted cells mount an immune response against the recipient.
granular leukocytes – leukocytes with abundant granules in their cytoplasm; specifically, neutrophils, eosinophils, and basophils.
granulosa cells – supportive cells in the ovarian follicle that produce oestrogen.
granzyme – apoptosis-inducing substance contained in granules of NK cells and cytotoxic T cells.
great cardiac vein – vessel that follows the interventricular sulcus on the anterior surface of the heart and flows along the coronary sulcus into the coronary sinus on the posterior surface; parallels the anterior interventricular artery and drains the areas supplied by this vessel.
great cerebral vein – receives most of the smaller vessels from the inferior cerebral veins and leads to the straight sinus.
greater pelvis – (also, greater pelvic cavity or false pelvis) broad space above the pelvic brim defined laterally by the fan-like portion of the upper ilium.
great saphenous vein – prominent surface vessel located on the medial surface of the leg and thigh; drains the superficial portions of these areas and leads to the femoral vein.
greater sciatic foramen – pelvic opening formed by the greater sciatic notch of the hip bone, the sacrum, and the sacrospinous ligament.
greater sciatic notch – large, U-shaped indentation located on the posterior margin of the ilium, superior to the ischial spine.
greater trochanter – large, bony expansion of the femur that projects superiorly from the base of the femoral neck.
greater tubercle – enlarged prominence located on the lateral side of the proximal humerus.
greater wings of sphenoid bone – lateral projections of the sphenoid bone that form the anterior wall of the middle cranial fossa and an area of the lateral skull.
grey matter – regions of the nervous system containing cell bodies of neurons with few or no myelinated axons; typically may be more pink or tan in colour, but called grey in contrast to white matter.
gross anatomy – study of the larger structures of the body, typically with the unaided eye; also referred to macroscopic anatomy.
ground substance – fluid or semi-fluid portion of the matrix.
growth hormone (GH) – anterior pituitary hormone that promotes tissue building and influences nutrient metabolism (also called somatotropin).
gustation – sense of taste.
gustatory receptor cells – sensory cells in the taste bud that transduce the chemical stimuli of gustation.
gyrus – ridge formed by convolutions on the surface of the cerebrum or cerebellum.
G0 phase – phase of the cell cycle, usually entered from the G1 phase; characterised by long or permanent periods where the cell does not move forward into the DNA synthesis phase.
G1 phase – first phase of the cell cycle, after a new cell is born.
G2 phase – third phase of the cell cycle, after the DNA synthesis phase.
H
haem – red, iron-containing pigment to which oxygen binds in haemoglobin.
haemangioblasts – embryonic stem cells that appear in the mesoderm and give rise to both angioblasts and pluripotent stem cells.
haematocrit – (also, packed cell volume) volume percentage of erythrocytes in a sample of centrifuged blood.
haematopoiesis – production of blood cells, which occurs in the red marrow of the bones.
haemocytoblast – hemopoietic stem cell that gives rise to the formed elements of blood.
haemoglobin – oxygen-carrying globular protein in erythrocytes.
haemolysis – destruction (lysis) of erythrocytes and the release of their haemoglobin into circulation.
haemolytic disease of the foetus and newborn (HDFN) – (also, erythroblastosis fetalis) disorder causing agglutination and haemolysis in an Rh+ foetus or newborn of an Rh− mother.
haemopoiesis – production of the formed elements of blood.
haemopoietic growth factors – chemical signals including erythropoietin, thrombopoietin, colony-stimulating factors, and interleukins that regulate the differentiation and proliferation of particular blood progenitor cells.
haemopoietic stem cell – type of pluripotent stem cell that gives rise to the formed elements of blood (haemocytoblast).
haemosiderin – protein-containing storage form of iron found in the bone marrow, liver, and spleen.
haemophilia – genetic disorder characterised by inadequate synthesis of clotting factors.
haemorrhage – excessive bleeding.
haemostasis – physiological process by which bleeding ceases.
hair – keratinous filament growing out of the epidermis.
hair bulb – structure at the base of the hair root that surrounds the dermal papilla.
hair cells – mechanoreceptor cells found in the inner ear that transduce stimuli for the senses of hearing and balance.
hair follicle – cavity or sac from which hair originates.
hair matrix – layer of basal cells from which a strand of hair grows.
hair papilla – mass of connective tissue, blood capillaries, and nerve endings at the base of the hair follicle.
hair root – part of hair that is below the epidermis anchored to the follicle.
hair shaft – part of hair that is above the epidermis but is not anchored to the follicle.
Haldane effect – relationship between the partial pressure of oxygen and the affinity of haemoglobin for carbon dioxide.
hallux – big toe; digit 1 of the foot.
hamate – from the lateral side, the fourth of the four distal carpal bones; articulates with the lunate and triquetrum proximally, the fourth and fifth metacarpals distally, and the capitate laterally.
hand – region of the upper limb distal to the wrist joint.
hard palate – bony structure that forms the roof of the mouth and floor of the nasal cavity, formed by the palatine process of the maxillary bones and the horizontal plate of the palatine bones.
haustrum – small pouch in the colon created by tonic contractions of teniae coli.
haustral contraction – slow segmentation in the large intestine.
head of the femur – rounded, proximal end of the femur that articulates with the acetabulum of the hip bone to form the hip joint.
head of the fibula – small, knob-like, proximal end of the fibula; articulates with the inferior aspect of the lateral condyle of the tibia.
head of the humerus – smooth, rounded region on the medial side of the proximal humerus; articulates with the glenoid fossa of the scapula to form the glenohumeral (shoulder) joint.
head of the metatarsal bone – expanded, distal end of each metatarsal bone.
head of the radius – disc-shaped structure that forms the proximal end of the radius; articulates with the capitulum of the humerus as part of the elbow joint, and with the radial notch of the ulna as part of the proximal radioulnar joint.
head of the rib – posterior end of a rib that articulates with the bodies of thoracic vertebrae.
head of the ulna – small, rounded distal end of the ulna; articulates with the ulnar notch of the distal radius, forming the distal radioulnar joint.
heart block – interruption in the normal conduction pathway.
heart bulge – prominent feature on the anterior surface of the heart, reflecting early cardiac development.
heart rate (HR) – number of times the heart contracts (beats) per minute.
heart sounds – sounds heard via auscultation with a stethoscope of the closing of the atrioventricular valves (“lub”) and semilunar valves (“dub”).
heavy chain – larger protein chain of an antibody.
helicase – enzyme that functions to separate the two DNA strands of a double helix during DNA replication.
helper T cells (Th) – T cells that secrete cytokines to enhance other immune responses, involved in activation of both B and T cell lymphocytes.
hemiazygos vein – smaller vein complementary to the azygos vein; drains the oesophageal veins from the oesophagus and the left intercostal veins, and leads to the brachiocephalic vein via the superior intercostal vein.
Henry’s law – statement of the principle that the concentration of gas in a liquid is directly proportional to the solubility and partial pressure of that gas.
heparin – short-acting anticoagulant stored in mast cells and released when tissues are injured, opposes prothrombin.
hepatic artery – artery that supplies oxygenated blood to the liver.
hepatic artery proper – branch of the common hepatic artery; supplies systemic blood to the liver.
hepatic lobule – hexagonal-shaped structure composed of hepatocytes that radiate outward from a central vein.
hepatic portal system – specialised circulatory pathway that carries blood from digestive organs to the liver for processing before being sent to the systemic circulation.
hepatic portal vein – vein that supplies deoxygenated nutrient-rich blood to the liver.
hepatic sinusoid – blood capillaries between rows of hepatocytes that receive blood from the hepatic portal vein and the branches of the hepatic artery.
hepatic vein – drains systemic blood from the liver and flows into the inferior vena cava.
hepatocytes – major functional cells of the liver.
hepatopancreatic ampulla – (also, ampulla of Vater) bulb-like point in the wall of the duodenum where the bile duct and main pancreatic duct unite.
hepatopancreatic sphincter – (also, sphincter of Oddi) sphincter regulating the flow of bile and pancreatic secretion into the duodenum.
high endothelial venules – vessels containing unique endothelial cells specialised to allow migration of lymphocytes from the blood to the lymph node.
hilum – concave structure on the mediastinal surface of the lungs where blood vessels, lymphatic vessels, nerves, and a bronchus enter the lung.
hindbrain – posterior region of the adult brain that develops from the rhombencephalon and includes the pons, medulla oblongata, and cerebellum.
hinge joint – synovial joint at which the convex surface of one bone articulates with the concave surface of a second bone; includes the elbow, knee, ankle, and interphalangeal joints; functionally classified as a uniaxial joint.
hip bone – coxal bone; single bone that forms the pelvic girdle; consists of three areas, the ilium, ischium, and pubis.
hip joint – joint located at the proximal end of the lower limb; formed by the articulation between the acetabulum of the hip bone and the head of the femur.
hippocampus – grey matter deep in the temporal lobe that is very important for long-term memory formation.
histamine – chemical compound released by mast cells in response to injury that causes vasodilation and endothelium permeability.
histology – microscopic study of tissue architecture, organisation, and function.
histone – family of proteins that associate with DNA in the nucleus to form chromatin.
hole – opening or depression in a bone.
holocrine secretion – release of a substance caused by the rupture of a gland cell, which becomes part of the secretion.
homeostasis – steady state of body systems that living organisms maintain.
homologous – describes two copies of the same chromosome (not identical), one inherited from each parent.
hook of the hamate bone – bony extension located on the anterior side of the hamate carpal bone.
horizontal plate – medial extension from the palatine bone that forms the posterior quarter of the hard palate.
hormone – secretion of an endocrine organ that travels via the bloodstream or lymphatics to induce a response in target cells or tissues in another part of the body.
hormone receptor – protein within a cell or on the cell membrane that binds a hormone, initiating the target cell response.
human chorionic gonadotropin (hCG) – hormone that directs the corpus luteum to survive, enlarge, and continue producing progesterone and oestrogen to suppress menses and secure an environment suitable for the developing embryo.
humerus – single bone of the upper arm.
humeroradial joint – articulation between the capitulum of the humerus and head of the radius.
humeroulnar joint – articulation between the trochlea of humerus and the trochlear notch of the ulna; uniaxial hinge joint that allows for flexion/extension of the forearm.
hyaline cartilage – most common type of cartilage, smooth and made of short collagen fibres embedded in a chondroitin sulphate ground substance.
hydrochloric acid (HCl) – digestive acid secreted by parietal cells in the stomach.
hydrolysis – reaction that causes breakdown of larger molecules into smaller molecules by utilising water.
hydrophilic – describes a substance or structure attracted to water.
hydrophobic – describes a substance or structure repelled by water.
hydrostatic pressure – pressure exerted by a fluid against a wall, caused by its own weight or pumping force.
hymen – membrane that covers part of the opening of the vagina.
hyoid bone – small, U-shaped bone located in upper neck that does not contact any other bone.
hypercalcaemia – condition characterised by abnormally elevated levels of calcium.
hypercapnia – abnormally elevated blood levels of CO2.
hyperplasia – process in which one cell splits to produce new cells.
hyperpnoea – increased rate and depth of ventilation due to an increase in oxygen demand that does not significantly alter blood oxygen or carbon dioxide levels.
hypertension – chronic and persistent blood pressure measurements of 140/90 mm Hg or above.
hyperthyroidism – clinically abnormal, elevated level of thyroid hormone in the blood; characterised by an increased metabolic rate, excess body heat, sweating, diarrhoea, weight loss, and increased heart rate.
hypertonia – abnormally high muscle tone.
hypertonic – describes a solution concentration that is higher than a reference concentration.
hypertrophic cardiomyopathy – pathological enlargement of the heart, generally for no known reason.
hypertrophy – addition of structural proteins to muscle fibres.
hyperventilation – increased ventilation rate that leads to abnormally low blood carbon dioxide levels and high (alkaline) blood pH.
hypervolaemia – abnormally high levels of fluid and blood within the body.
hypoblast – lower layer of cells of the embryonic disc that extend into the blastocoel to form the yolk sac.
hypocalcaemia – condition characterised by abnormally low levels of calcium.
hypophyseal (pituitary) fossa – shallow depression on top of the sella turcica that houses the pituitary (hypophyseal) gland.
hypophyseal portal system – network of blood vessels that enables hypothalamic hormones to travel into the anterior lobe of the pituitary without entering the systemic circulation.
hypothalamus – major region of the diencephalon that is responsible for coordinating autonomic and endocrine control of homeostasis.
hypothyroidism – clinically abnormal, low level of thyroid hormone in the blood; characterised by low metabolic rate, weight gain, cold extremities, constipation, and reduced mental activity.
hypotonia – abnormally low muscle tone caused by the absence of low-level contractions.
hypotonic – describes a solution concentration that is lower than a reference concentration.
hypovolaemia – abnormally low levels of fluid and blood within the body.
hypovolaemic shock – type of circulatory shock caused by excessive loss of blood volume due to haemorrhage or possibly dehydration.
hypoxaemia – below-normal level of oxygen saturation of blood (typically <95 percent).
hypoxia – lack of oxygen supply to the tissues.
I
IgA – antibody whose dimer is secreted by exocrine glands, is especially effective against digestive and respiratory pathogens, and can pass immunity to an infant through breastfeeding.
IgD – class of antibody whose only known function is as a receptor on naive B cells; important in B cell activation.
IgE – antibody that binds to mast cells and causes antigen-specific degranulation during an allergic response.
IgG – main blood antibody of late primary and early secondary responses; passed from mother to unborn child via placenta.
IgM – antibody whose monomer is a surface receptor of naive B cells; the pentamer is the first antibody made blood plasma during primary responses.
iliac crest – curved, superior margin of the ilium.
iliac fossa – shallow depression found on the anterior and medial surfaces of the upper ilium.
ileocaecal sphincter – sphincter located where the small intestine joins with the large intestine.
ileum – end of the small intestine between the jejunum and the large intestine.
iliofemoral ligament – intrinsic ligament spanning from the ilium of the hip bone to the femur, on the superior-anterior aspect of the hip joint.
ilium – superior portion of the hip bone.
immediate hypersensitivity – (type I) IgE-mediated mast cell degranulation caused by crosslinking of surface IgE by antigen.
immune system – series of barriers, cells, and soluble mediators that combine to response to infections of the body with pathogenic organisms.
immunoglobulins – (also, antibodies or gamma globulins) antigen-specific proteins produced by specialised B lymphocytes that protect the body by binding to foreign objects such as bacteria and viruses.
immunological memory – ability of the adaptive immune response to mount a stronger and faster immune response upon re-exposure to a pathogen.
hypoblast – lower layer of cells of the embryonic disc that extend into the blastocoel to form the yolk sac.
inactivation gate – part of a voltage-gated Na+ channel that closes when the membrane potential reaches +30 mV.
incisor – midline, chisel-shaped tooth used for cutting into food.
incontinence – loss of ability to control micturition
incus – (also, anvil) ossicle of the middle ear that connects the malleus to the stapes.
indirect pathway – connections within the basal nuclei from the striatum through the globus pallidus external segment and subthalamic nucleus to the globus pallidus internal segment/substantia nigra pars compacta that result in inhibition of the thalamus to decrease cortical control of movement.
inferior – describes a position below or lower than another part of the body proper; near or toward the tail (in humans, the coccyx, or lowest part of the spinal column); also referred to as caudal.
inferior angle of the scapula – inferior corner of the scapula located where the medial and lateral borders meet.
inferior articular process – bony process that extends downward from the vertebral arch of a vertebra that articulates with the superior articular process of the next lower vertebra.
inferior colliculus – half of the midbrain tectum that is part of the brain stem auditory pathway.
inferior mesenteric artery – branch of the abdominal aorta; supplies blood to the distal segment of the large intestine and rectum.
inferior nasal concha – one of the paired bones that project from the lateral walls of the nasal cavity to form the largest and most inferior of the nasal conchae.
inferior oblique – extraocular muscle responsible for lateral rotation of the eye.
inferior olive – nucleus in the medulla that is involved in processing information related to motor control.
inferior phrenic artery – branch of the abdominal aorta; supplies blood to the inferior surface of the diaphragm.
inferior pubic ramus – narrow segment of bone that passes inferiorly and laterally from the pubic body; joins with the ischial ramus to form the ischiopubic ramus.
inferior rectus – extraocular muscle responsible for looking down.
inferior rotation – movement of the scapula during upper limb adduction in which the glenoid cavity of the scapula moves in a downward direction as the medial end of the scapular spine moves in an upward direction.
inferior vena cava – large systemic vein that returns blood to the heart from the inferior portion of the body.
inflammation – response of tissue to injury.
infraglenoid tubercle – small bump or roughened area located on the lateral border of the scapula, near the inferior margin of the glenoid cavity.
infraorbital foramen – opening located on anterior skull, below the orbit.
infraspinous fossa – broad depression located on the posterior scapula, inferior to the spine.
infratemporal fossa – space on lateral side of skull, below the level of the zygomatic arch and deep (medial) to the ramus of the mandible.
infundibulum – stalk containing vasculature and neural tissue that connects the pituitary gland to the hypothalamus (also called the pituitary stalk).
infundibulum – (of the uterine tube) wide, distal portion of the uterine tube terminating in fimbriae.
ingestion – taking food into the GI tract through the mouth.
inguinal canal – opening in abdominal wall that connects the testes to the abdominal cavity.
inhibin – hormone secreted by the male and female gonads that inhibits FSH production by the anterior pituitary.
inhibitory postsynaptic potential (IPSP) – graded potential in the postsynaptic membrane that is the result of hyperpolarisation and makes an action potential less likely to occur.
initial segment – first part of the axon as it emerges from the axon hillock, where the electrical signals known as action potentials are generated.
innate immune response – rapid but relatively nonspecific immune response.
inner cell mass – cluster of cells within the blastocyst that is fated to become the embryo.
inner ear – structure within the temporal bone that contains the sensory apparati of hearing and balance.
inner segment – in the eye, the section of a photoreceptor that contains the nucleus and other major organelles for normal cellular functions.
inner synaptic layer – layer in the retina where bipolar cells connect to RGCs.
inositol triphosphate (IP3) – molecule that initiates the release of calcium ions from intracellular stores.
inspiration – (also, inhalation) process that causes air to enter the lungs.
inspiratory capacity (IC) – sum of the TV and IRV, which is the amount of air that can maximally be inhaled past a tidal expiration.
inspiratory reserve volume (IRV) – amount of air that enters the lungs due to deep inhalation past the tidal volume.
insulin – pancreatic hormone that enhances the cellular uptake and utilisation of glucose, thereby decreasing blood glucose levels.
insulin-like growth factors (IGF) – protein that enhances cellular proliferation, inhibits apoptosis, and stimulates the cellular uptake of amino acids for protein synthesis.
integral protein – membrane-associated protein that spans the entire width of the lipid bilayer.
integration – nervous system function that combines sensory perceptions and higher cognitive functions (memories, learning, emotion, etc.) to produce a response.
integumentary system – skin and its accessory structures.
interatrial band – (also, Bachmann’s bundle) group of specialised conducting cells that transmit the impulse directly from the SA node in the right atrium to the left atrium.
interatrial septum – cardiac septum located between the two atria; contains the fossa ovalis after birth.
interaural intensity difference – cue used to aid sound localisation in the horizontal plane that compares the relative loudness of sounds at the two ears, because the ear closer to the sound source will hear a slightly more intense sound.
interaural time difference – cue used to help with sound localisation in the horizontal plane that compares the relative time of arrival of sounds at the two ears, because the ear closer to the sound source will receive the stimulus microseconds before the other ear.
intercalated cell – specialised cell of the collecting ducts that secrete or absorb acid or bicarbonate; important in acid–base balance.
intercalated disc – physical junction between adjacent cardiac muscle cells; consisting of desmosomes, specialised linking proteoglycans, and gap junctions that allow passage of ions between the two cells.
intercondylar eminence – irregular elevation on the superior end of the tibia, between the articulating surfaces of the medial and lateral condyles.
intercondylar fossa – deep depression on the posterior side of the distal femur that separates the medial and lateral condyles.
intercostal artery – branch of the thoracic aorta; supplies blood to the muscles of the thoracic cavity and vertebral column.
intercostal nerve – systemic nerve in the thoracic cavity that is found between two ribs.
intercostal vein – drains the muscles of the thoracic wall and leads to the azygos vein.
interferons – early induced proteins made in virally infected cells that cause nearby cells to make antiviral proteins.
interleukins – signalling molecules that may function in haemopoiesis, inflammation, and specific immune responses.
intermediate cuneiform – middle of the three cuneiform tarsal bones; articulates posteriorly with the navicular bone, medially with the medial cuneiform bone, laterally with the lateral cuneiform bone, and anteriorly with the second metatarsal bone.
intermediate filament – type of cytoskeletal filament made of keratin, characterised by an intermediate thickness, and playing a role in resisting cellular tension.
internal acoustic meatus – opening into petrous ridge, located on the lateral wall of the posterior cranial fossa.
internal anal sphincter – involuntary smooth muscle sphincter in the anal canal.
internal callus – fibrocartilaginous matrix, in the endosteal region, between the two ends of a broken bone.
internal capsule – segment of the descending motor pathway that passes between the caudate nucleus and the putamen.
internal carotid artery – arises from the common carotid artery and begins with the carotid sinus; goes through the carotid canal of the temporal bone to the base of the brain; combines with branches of the vertebral artery forming the arterial circle; supplies blood to the brain.
internal elastic membrane – membrane composed of elastic fibres that separates the tunica intima from the tunica media; seen in larger arteries.
internal iliac artery – branch from the common iliac arteries; supplies blood to the urinary bladder, walls of the pelvis, external genitalia, and the medial portion of the femoral region; in females, also provide blood to the uterus and vagina.
internal iliac vein – drains the pelvic organs and integument; formed from several smaller veins in the region; leads to the common iliac vein.
internal jugular vein – one of a pair of major veins located in the neck region that passes through the jugular foramen and canal, flows parallel to the common carotid artery that is more or less its counterpart; primarily drains blood from the brain, receives the superficial facial vein, and empties into the subclavian vein.
internal respiration – gas exchange that occurs at the level of body tissues.
internal root sheath – innermost layer of keratinocytes in the hair follicle that surround the hair root up to the hair shaft.
internal thoracic artery – (also, mammary artery) arises from the subclavian artery; supplies blood to the thymus, pericardium of the heart, and the anterior chest wall.
internal thoracic vein – (also, internal mammary vein) drains the anterior surface of the chest wall and leads to the brachiocephalic vein.
internal urinary sphincter – smooth muscle at the juncture of the bladder and urethra; relaxes as the bladder fills to allow urine into the urethra
internodal pathways – specialised conductile cells within the atria that transmit the impulse from the SA node throughout the myocardial cells of the atrium and to the AV node.
interoceptor – sensory receptor that is positioned to interpret stimuli from internal organs, such as stretch receptors in the wall of blood vessels.
interosseous border of the fibula – small ridge running down the medial side of the fibular shaft; for attachment of the interosseous membrane between the fibula and tibia.
interosseous border of the radius – narrow ridge located on the medial side of the radial shaft; for attachment of the interosseous membrane between the ulna and radius bones.
interosseous border of the tibia – small ridge running down the lateral side of the tibial shaft; for attachment of the interosseous membrane between the tibia and fibula.
interosseous border of the ulna – narrow ridge located on the lateral side of the ulnar shaft; for attachment of the interosseous membrane between the ulna and radius.
interosseous membrane – wide sheet of fibrous connective tissue that fills the gap between two parallel bones, forming a syndesmosis; found between the radius and ulna of the forearm and between the tibia and fibula of the leg.
interosseous membrane of the forearm – sheet of dense connective tissue that unites the radius and ulna bones.
interosseous membrane of the leg – sheet of dense connective tissue that unites the shafts of the tibia and fibula bones.
interphalangeal joint – articulation between adjacent phalanx bones of the hand or foot digits.
interphase – entire life cycle of a cell, excluding mitosis.
interstitial fluid (IF) – fluid in the small spaces between cells not contained within blood vessels.
interstitial fluid colloidal osmotic pressure (IFCOP) – pressure exerted by the colloids within the interstitial fluid.
interstitial fluid hydrostatic pressure (IFHP) – force exerted by the fluid in the tissue spaces.
intertrochanteric crest – short, prominent ridge running between the greater and lesser trochanters on the posterior side of the proximal femur.
intertrochanteric line – small ridge running between the greater and lesser trochanters on the anterior side of the proximal femur.
intertubercular groove (sulcus) – bicipital groove; narrow groove located between the greater and lesser tubercles of the humerus.
interventricular foramina – openings between the lateral ventricles and third ventricle allowing for the passage of CSF.
interventricular septum – cardiac septum located between the two ventricles.
intervertebral disc – structure located between the bodies of adjacent vertebrae that strongly joins the vertebrae; provides padding, weight bearing ability, and enables vertebral column movements.
intervertebral foramen – opening located between adjacent vertebrae for exit of a spinal nerve.
intestinal gland – (also, crypt of Lieberkühn) gland in the small intestinal mucosa that secretes intestinal secretion.
intestinal phase – phase of gastric secretion that begins when chyme enters the intestine.
intestinal secretion – mixture of water and mucus that helps absorb nutrients from chyme.
intra-alveolar pressure – (intrapulmonary pressure) pressure of the air within the alveoli.
intracapsular ligament – ligament that is located within the articular capsule of a synovial joint.
intracellular fluid (ICF) – fluid in the cytosol of cells.
intramembranous ossification – process by which bone forms directly from mesenchymal tissue.
intrapleural pressure – pressure of the air within the pleural cavity.
intrinsic ligament – ligament that is fused to or incorporated into the wall of the articular capsule of a synovial joint.
intrinsic factor – glycoprotein required for vitamin B12 absorption in the small intestine.
intrinsic pathway – initial coagulation pathway that begins with vascular damage or contact with foreign substances, and results in the activation of the common pathway.
intron – non-coding regions of a pre-mRNA transcript that may be removed during splicing.
inulin – plant polysaccharide injected to determine GFR; is neither secreted nor absorbed by the kidney, so its appearance in the urine is directly proportional to its filtration rate.
inversion – foot movement involving the intertarsal joints of the foot in which the bottom of the foot is turned toward the midline.
ionotropic receptor – neurotransmitter receptor that acts as an ion channel gate, and opens by the binding of the neurotransmitter.
ipsilateral – word meaning on the same side, as in axons that do not cross the midline in a fibre tract.
iris – coloured portion of the anterior eye that surrounds the pupil.
irregular bone – bone of complex shape; protects internal organs from compressive forces.
ischaemia – insufficient blood flow to the tissues.
ischial ramus – bony extension projecting anteriorly and superiorly from the ischial tuberosity; joins with the inferior pubic ramus to form the ischiopubic ramus.
ischial spine – pointed, bony projection from the posterior margin of the ischium that separates the greater sciatic notch and lesser sciatic notch.
ischial tuberosity – large, roughened protuberance that forms the posteroinferior portion of the hip bone; weight-bearing region of the pelvis when sitting.
ischiofemoral ligament – intrinsic ligament spanning from the ischium of the hip bone to the femur, on the posterior aspect of the hip joint.
ischiopubic ramus – narrow extension of bone that connects the ischial tuberosity to the pubic body; formed by the junction of the ischial ramus and inferior pubic ramus.
ischium – posteroinferior portion of the hip bone.
isometric contraction – muscle contraction that occurs with no change in muscle length.
isotonic – describes a solution concentration that is the same as a reference concentration.
isotonic contraction – muscle contraction that involves changes in muscle length.
isovolumic contraction – (also, isovolumetric contraction) initial phase of ventricular contraction in which tension and pressure in the ventricle increase, but no blood is pumped or ejected from the heart.
isovolumic ventricular relaxation phase – initial phase of the ventricular diastole when pressure in the ventricles drops below pressure in the two major arteries, the pulmonary trunk, and the aorta, and blood attempts to flow back into the ventricles, producing the dicrotic notch of the ECG and closing the two semilunar valves.
isthmus – narrow, medial portion of the uterine tube that joins the uterus.
J
jejunum – middle part of the small intestine between the duodenum and the ileum.
joint – site at which two or more bones or bone and cartilage come together (articulate).
joint cavity – space enclosed by the articular capsule of a synovial joint that is filled with synovial fluid and contains the articulating surfaces of the adjacent bones.
joint interzone – site within a growing embryonic limb bud that will become a synovial joint.
jugular foramen – irregularly shaped opening located in the lateral floor of the posterior cranial cavity.
jugular (suprasternal) notch – shallow notch located on superior surface of sternal manubrium.
jugular veins – blood vessels that return “used” blood from the head and neck.
juxtaglomerular apparatus (JGA) – located at the juncture of the DCT and the afferent and efferent arterioles of the glomerulus; plays a role in the regulation of renal blood flow and GFR.
juxtaglomerular cell – modified smooth muscle cells of the afferent arteriole; secretes renin in response to a drop in blood pressure.
juxtamedullary nephrons – nephrons adjacent to the border of the cortex and medulla with loops of Henle that extend into the renal medulla.
K
keloid – type of scar that has layers raised above the skin surface.
keratin – type of structural protein that gives skin, hair, and nails its hard, water-resistant properties.
keratinocyte – cell that produces keratin and is the most predominant type of cell found in the epidermis.
keratohyalin – granulated protein found in the stratum granulosum.
kinaesthesia – general sensory perception of movement of the body.
kinetochore – region of a centromere where microtubules attach to a pair of sister chromatids.
knee joint – joint that separates the thigh and leg portions of the lower limb; formed by the articulations between the medial and lateral condyles of the femur, and the medial and lateral condyles of the tibia.
Korotkoff sounds – noises created by turbulent blood flow through the vessels.
kyphosis – (also, humpback or hunchback) excessive posterior curvature of the thoracic vertebral column region.
L
labia majora – hair-covered folds of skin located behind the mons pubis.
labia minora – thin, pigmented, hairless flaps of skin located medial and deep to the labia majora.
labium – lip.
labial frenulum – midline mucous membrane fold that attaches the inner surface of the lips to the gums.
lacrimal bone – paired bones that contribute to the anterior-medial wall of each orbit.
lacrimal duct – duct in the medial corner of the orbit that drains tears into the nasal cavity.
lacrimal fossa – shallow depression in the anterior-medial wall of the orbit, formed by the lacrimal bone that gives rise to the nasolacrimal canal.
lacrimal gland – gland lateral to the orbit that produces tears to wash across the surface of the eye.
lactase – brush border enzyme that breaks down lactose into glucose and galactose.
lacteal – lymphatic capillary in the villi.
lactic acid – product of anaerobic glycolysis.
lactiferous ducts – ducts that connect the mammary glands to the nipple and allow for the transport of milk.
lactiferous sinus – area of milk collection between alveoli and lactiferous duct.
lacunae – (singular = lacuna) small spaces in bone or cartilage tissue that cells occupy.
lambdoid suture – inverted V-shaped joint that unites the occipital bone to the right and left parietal bones on the posterior skull.
lamina – portion of the vertebral arch on each vertebra that extends between the transverse and spinous process.
lamina propria – areolar connective tissue underlying a mucous membrane.
Langerhans cell – specialised dendritic cell found in the stratum spinosum that functions as a macrophage.
lanugo – silk-like hairs that coat the foetus; shed later in foetal development.
large intestine – terminal portion of the gastrointestinal tract.
laryngeal prominence – region where the two lamina of the thyroid cartilage join, forming a protrusion known as “Adam’s apple”.
laryngopharynx – portion of the pharynx bordered by the oropharynx superiorly and oesophagus and trachea inferiorly; serves as a route for both air and food.
laryngotracheal – bud forms from the lung bud, has a tracheal end and bulbous bronchial buds at the distal end.
larynx – cartilaginous structure that produces the voice, prevents food and beverages from entering the trachea, and regulates the volume of air that enters and leaves the lungs.
latch-bridges – subset of a cross-bridge in which actin and myosin remain locked together.
latent period – the time when a twitch does not produce contraction.
lateral – describes the side or direction toward the side of the body.
lateral apertures – pair of openings from the fourth ventricle to the subarachnoid space on either side and between the medulla and cerebellum.
lateral border of the scapula – diagonally oriented lateral margin of the scapula.
lateral circumflex artery – branch of the deep femoral artery; supplies blood to the deep muscles of the thigh and the ventral and lateral regions of the integument.
lateral column – white matter of the spinal cord between the posterior horn on one side and the axons from the anterior horn on the same side; composed of many different groups of axons, of both ascending and descending tracts, carrying motor commands to and from the brain.
lateral condyle of the femur – smooth, articulating surface that forms the distal and posterior sides of the lateral expansion of the distal femur.
lateral condyle of the tibia – lateral, expanded region of the proximal tibia that includes the smooth surface that articulates with the lateral condyle of the femur as part of the knee joint.
lateral corticospinal tract – division of the corticospinal pathway that travels through the lateral column of the spinal cord and controls appendicular musculature through the lateral motor neurons in the ventral (anterior) horn.
lateral cuneiform – most lateral of the three cuneiform tarsal bones; articulates posteriorly with the navicular bone, medially with the intermediate cuneiform bone, laterally with the cuboid bone, and anteriorly with the third metatarsal bone.
lateral epicondyle of the femur – roughened area of the femur located on the lateral side of the lateral condyle.
lateral epicondyle of the humerus – small projection located on the lateral side of the distal humerus.
lateral excursion – side-to-side movement of the mandible away from the midline, toward either the right or left side.
lateral flexion – bending of the neck or body toward the right or left side.
lateral geniculate nucleus – thalamic target of the RGCs that projects to the visual cortex.
lateral horn – region of the spinal cord grey matter in the thoracic, upper lumbar, and sacral regions that is the central component of the sympathetic division of the autonomic nervous system.
lateral malleolus – expanded distal end of the fibula.
lateral meniscus – C-shaped fibrocartilage articular disc located at the knee, between the lateral condyle of the femur and the lateral condyle of the tibia.
lateral plantar artery – arises from the bifurcation of the posterior tibial arteries; supplies blood to the lateral plantar surfaces of the foot.
lateral pterygoid plate – paired, flattened bony projections of the sphenoid bone located on the inferior skull, lateral to the medial pterygoid plate.
lateral rectus – extraocular muscle responsible for abduction of the eye.
lateral (external) rotation – movement of the arm at the shoulder joint or the thigh at the hip joint that moves the anterior surface of the limb away from the midline of the body.
lateral sacral crest – paired irregular ridges running down the lateral sides of the posterior sacrum that was formed by the fusion of the transverse processes from the five sacral vertebrae.
lateral sulcus – surface landmark of the cerebral cortex that marks the boundary between the temporal lobe and the frontal and parietal lobes.
lateral supracondylar ridge – narrow, bony ridge located along the lateral side of the distal humerus, superior to the lateral epicondyle.
lateral tibiofemoral joint – portion of the knee consisting of the articulation between the lateral condyle of the tibia and the lateral condyle of the femur; allows for flexion/extension at the knee.
lateral ventricles – portions of the ventricular system that are in the region of the cerebrum.
leakage channel – ion channel that opens randomly and is not gated to a specific event, also known as a non-gated channel.
leaky tight junctions – tight junctions in which the sealing strands of proteins between the membranes of adjacent cells are fewer in number and incomplete; allows limited intercellular movement of solvent and solutes.
left atrioventricular valve – (also, mitral valve or bicuspid valve) valve located between the left atrium and ventricle; consists of two flaps of tissue.
left colic flexure – (also, splenic flexure) point where the transverse colon curves below the inferior end of the spleen.
left gastric artery – branch of the celiac trunk; supplies blood to the stomach.
leg – portion of the lower limb located between the knee and ankle joints.
lens – component of the eye that focuses light on the retina.
leptin – protein hormone secreted by adipose tissues in response to food consumption that promotes satiety.
lesser pelvis – (also, lesser pelvic cavity or true pelvis) narrow space located within the pelvis, defined superiorly by the pelvic brim (pelvic inlet) and inferiorly by the pelvic outlet.
lesser sciatic foramen – pelvic opening formed by the lesser sciatic notch of the hip bone, the sacrospinous ligament, and the sacrotuberous ligament.
lesser sciatic notch – shallow indentation along the posterior margin of the ischium, inferior to the ischial spine.
lesser trochanter – small, bony projection on the medial side of the proximal femur, at the base of the femoral neck.
lesser tubercle – small, bony prominence located on anterior side of the proximal humerus.
lesser wings of the sphenoid bone – lateral extensions of the sphenoid bone that form the bony lip separating the anterior and middle cranial fossae.
leukaemia – cancer involving leukocytes.
leukocyte – (also, white blood cell) colourless, nucleated blood cell, the chief function of which is to protect the body from disease.
leukocyte esterase – enzyme produced by leukocytes that can be detected in the urine and that serves as an indirect indicator of urinary tract infection.
leukocytosis – excessive leukocyte proliferation.
leukopenia – below-normal production of leukocytes.
levator palpebrae superioris – muscle that causes elevation of the upper eyelid, controlled by fibres in the oculomotor nerve.
Leydig cells – cells between the seminiferous tubules of the testes that produce testosterone, a type of interstitial cell.
ligament – strong band of dense connective tissue spanning between bones.
ligament of the head of the femur – ligament that spans the acetabulum of the hip bone and the fovea capitis of the femoral head.
ligamentum flavum – series of short ligaments that unite the lamina of adjacent vertebrae.
ligand – molecule that binds with specificity to a specific receptor molecule.
ligand-gated channels – another name for an ionotropic receptor for which a neurotransmitter is the ligand.
light chain – small protein chain of an antibody.
limb bud – small elevation that appears on the lateral side of the embryo during the fourth or fifth week of development, which gives rise to an upper or lower limb.
limbic cortex – collection of structures of the cerebral cortex that are involved in emotion, memory, and behaviour and are part of the larger limbic system.
limbic system – structures at the edge (limit) of the boundary between the forebrain and hindbrain that are most associated with emotional behaviour and memory formation.
linea aspera – longitudinally running bony ridge located in the middle third of the posterior femur.
lingual frenulum – mucous membrane fold that attaches the bottom of the tongue to the floor of the mouth.
lingual lipase – digestive enzyme from glands in the tongue that acts on triglycerides.
lingual tonsil – lymphoid tissue located at the base of the tongue.
lingula – small flap of bone located on the inner (medial) surface of mandibular ramus, next to the mandibular foramen.
lipid – macromolecule that is nonpolar and insoluble in water.
lipoprotein lipase – enzyme that breaks down triglycerides in chylomicrons into fatty acids and monoglycerides.
liver – largest gland in the body whose main digestive function is the production of bile.
long bone – cylinder-shaped bone that is longer than it is wide; functions as a lever.
longitudinal fissure – large separation along the midline between the two cerebral hemispheres.
loop of Henle – descending and ascending portions between the proximal and distal convoluted tubules; those of cortical nephrons do not extend into the medulla, whereas those of juxtamedullary nephrons do extend into the medulla.
loose connective tissue – (also, areolar tissue) type of connective tissue proper that shows little specialisation with cells dispersed in the matrix.
lordosis – (also, swayback) excessive anterior curvature of the lumbar vertebral column region.
lower motor neuron – second neuron in the motor command pathway that is directly connected to the skeletal muscle.
lower oesophageal sphincter – smooth muscle sphincter that regulates food movement from the oesophagus to the stomach.
lumbar arteries – branches of the abdominal aorta; supply blood to the lumbar region, the abdominal wall, and spinal cord.
lumbar curve – posteriorly concave curvature of the lumbar vertebral column region; a secondary curve of the vertebral column.
lumbar enlargement – region of the ventral (anterior) horn of the spinal cord that has a larger population of motor neurons for the greater number of muscles of the lower limb.
lumbar plexus – nerve plexus associated with the lumbar spinal nerves.
lumbar puncture – procedure used to withdraw CSF from the lower lumbar region of the vertebral column that avoids the risk of damaging CNS tissue because the spinal cord ends at the upper lumbar vertebrae.
lumbar veins – drain the lumbar portion of the abdominal wall and spinal cord; the superior lumbar veins drain into the azygos vein on the right or the hemiazygos vein on the left; blood from these vessels is returned to the superior vena cava rather than the inferior vena cava.
lumbar vertebrae – five vertebrae numbered as L1–L5 that are located in lumbar region (lower back) of the vertebral column.
lumen – interior of a tubular structure such as a blood vessel or a portion of the alimentary canal through which blood, chyme, or other substances travel.
lunate – from the lateral side, the second of the four proximal carpal bones; articulates with the radius proximally, the capitate and hamate distally, the scaphoid laterally, and the triquetrum medially.
lung – organ of the respiratory system that performs gas exchange.
lung bud – median dome that forms from the endoderm of the foregut.
lunula – basal part of the nail body that consists of a crescent-shaped layer of thick epithelium.
luteinising hormone (LH) – anterior pituitary hormone that triggers ovulation and the production of ovarian hormones in females, and the production of testosterone in males.
lymph – fluid contained within the lymphatic system.
lymphocytes – agranular leukocytes of the lymphoid stem cell line, many of which function in specific immunity.
lymphoid stem cells – type of haemopoietic stem cells that gives rise to lymphocytes, including various T cells, B cells, and NK cells, all of which function in immunity.
lymphoma – form of cancer in which masses of malignant T and/or B lymphocytes collect in lymph nodes, the spleen, the liver, and other tissues.
lymph node – one of the bean-shaped organs found associated with the lymphatic vessels.
lymphatic capillaries – smallest of the lymphatic vessels and the origin of lymph flow.
lymphatic system – network of lymphatic vessels, lymph nodes, and ducts that carries lymph from the tissues and back to the bloodstream.
lymphatic trunks – large lymphatics that collect lymph from smaller lymphatic vessels and empties into the blood via lymphatic ducts.
lymphocytes – white blood cells characterised by a large nucleus and small rim of cytoplasm.
lymphoid nodules – unencapsulated patches of lymphoid tissue found throughout the body.
lysozyme – digestive enzyme with bactericidal properties.
lysosome – membrane-bound cellular organelle originating from the Golgi apparatus and containing digestive enzymes.
M
macrophage – phagocytic cell of the myeloid lineage; a matured monocyte.
macrophage oxidative metabolism – metabolism turned on in macrophages by T cell signals that help destroy intracellular bacteria.
macula – enlargement at the base of a semicircular canal at which transduction of equilibrium stimuli takes place within the ampulla.
macula densa – cells found in the part of the DCT forming the JGA; sense Na+ concentration in the forming urine.
main pancreatic duct – (also, duct of Wirsung) duct through which pancreatic secretion drains from the pancreas.
major duodenal papilla – point at which the hepatopancreatic ampulla opens into the duodenum.
major histocompatibility complex (MHC) – gene cluster whose proteins present antigens to T cells.
malleus – (also, hammer) ossicle that is directly attached to the tympanic membrane.
maltase – brush border enzyme that breaks down maltose and maltotriose into two and three molecules of glucose, respectively.
mammary glands – glands inside the breast that secrete milk.
mandible – unpaired bone that forms the lower jaw bone; the only moveable bone of the skull.
mandibular foramen – opening located on the inner (medial) surface of the mandibular ramus.
mandibular fossa – oval depression located on the inferior surface of the skull.
mandibular notch – large U-shaped notch located between the condylar process and coronoid process of the mandible.
manubrium – expanded, superior portion of the sternum.
marginal arteries – branches of the right coronary artery that supply blood to the superficial portions of the right ventricle.
mass movement – long, slow, peristaltic wave in the large intestine.
mast cell – cell found in the skin and the lining of body cells that contains cytoplasmic granules with vasoactive mediators such as histamine.
mastication – chewing.
mastoid process – large bony prominence on the inferior, lateral skull, just behind the earlobe.
matrix – extracellular material which is produced by the cells embedded in it, containing ground substance and fibres.
maxillary bone – (also, maxilla) paired bones that form the upper jaw and anterior portion of the hard palate.
maxillary sinus – air-filled space located with each maxillary bone; largest of the paranasal sinuses.
maxillary vein – drains blood from the maxillary region and leads to the external jugular vein.
mean arterial pressure (MAP) – average driving force of blood to the tissues; approximated by taking diastolic pressure and adding 1/3 of pulse pressure.
meatus – one of three recesses (superior, middle, and inferior) in the nasal cavity attached to the conchae that increase the surface area of the nasal cavity.
mechanical digestion – chewing, mixing, and segmentation that prepares food for chemical digestion.
mechanically gated channel – ion channel that opens when a physical event directly affects the structure of the protein.
mechanoreceptor – receptor cell that transduces mechanical stimuli into an electrochemical signal.
meconium – foetal wastes consisting of ingested amniotic fluid, cellular debris, mucus, and bile.
medial – describes the middle or direction toward the middle of the body.
medial border of the scapula – elongated, medial margin of the scapula.
medial condyle of the femur – smooth, articulating surface that forms the distal and posterior sides of the medial expansion of the distal femur.
medial condyle of the tibia – medial, expanded region of the proximal tibia that includes the smooth surface that articulates with the medial condyle of the femur as part of the knee joint.
medial cuneiform – most medial of the three cuneiform tarsal bones; articulates posteriorly with the navicular bone, laterally with the intermediate cuneiform bone, and anteriorly with the first and second metatarsal bones.
medial epicondyle of the femur – roughened area of the distal femur located on the medial side of the medial condyle.
medial epicondyle of the humerus – enlarged projection located on the medial side of the distal humerus.
medial excursion – side-to-side movement that returns the mandible to the midline.
medial geniculate nucleus – thalamic target of the auditory brain stem that projects to the auditory cortex.
medial lemniscus – fibre tract of the dorsal column system that extends from the nuclei gracilis and cuneatus to the thalamus, and decussates.
medial malleolus – bony expansion located on the medial side of the distal tibia.
medial meniscus – C-shaped fibrocartilage articular disc located at the knee, between the medial condyle of the femur and medial condyle of the tibia.
medial plantar artery – arises from the bifurcation of the posterior tibial arteries; supplies blood to the medial plantar surfaces of the foot.
medial pterygoid plate – paired, flattened bony projections of the sphenoid bone located on the inferior skull medial to the lateral pterygoid plate; form the posterior portion of the nasal cavity lateral wall.
medial rectus – extraocular muscle responsible for adduction of the eye.
medial (internal) rotation – movement of the arm at the shoulder joint or the thigh at the hip joint that brings the anterior surface of the limb toward the midline of the body.
medial tibiofemoral joint – portion of the knee consisting of the articulation between the medial condyle of the tibia and the medial condyle of the femur; allows for flexion/extension at the knee.
median antebrachial vein – vein that parallels the ulnar vein but is more medial in location; intertwines with the palmar venous arches.
median aperture – singular opening from the fourth ventricle into the subarachnoid space at the midline between the medulla and cerebellum.
median cubital vein – superficial vessel located in the antecubital region that links the cephalic vein to the basilic vein in the form of a v; a frequent site for a blood draw.
median nerve – systemic nerve of the arm, located between the ulnar and radial nerves.
median sacral artery – continuation of the aorta into the sacrum.
median sacral crest – irregular ridge running down the midline of the posterior sacrum that was formed from the fusion of the spinous processes of the five sacral vertebrae.
mediastinal artery – branch of the thoracic aorta; supplies blood to the mediastinum.
medulla – in hair, the innermost layer of keratinocytes originating from the hair matrix.
medulla – inner region of kidney containing the renal pyramids.
medullary cavity – hollow region of the diaphysis; filled with yellow marrow.
megakaryocyte – bone marrow cell that produces platelets.
Meissner corpuscle – (also, tactile corpuscle) receptor in the skin that responds to light touch.
melanin – pigment that determines the colour of hair and skin.
melanocyte – cell found in the stratum basale of the epidermis that produces the pigment melanin.
melanoma – type of skin cancer that originates from the melanocytes of the skin.
melanosome – intercellular vesicle that transfers melanin from melanocytes into keratinocytes of the epidermis.
melatonin – amino acid–derived hormone that is secreted in response to low light and causes drowsiness.
membrane potential – distribution of charge across the cell membrane, based on the charges of ions.
memory cell – type of B or T lymphocyte that forms after exposure to a pathogen.
memory T cells – long-lived immune cell reserved for future exposure to a pathogen.
menarche – first menstruation in a pubertal female.
meninges – protective outer coverings of the CNS composed of connective tissue.
meniscus – articular disc.
menses – shedding of the inner portion of the endometrium out though the vagina; also referred to as menstruation.
menses phase – phase of the menstrual cycle in which the endometrial lining is shed.
menstrual cycle – approximately 28-day cycle of changes in the uterus consisting of a menses phase, a proliferative phase, and a secretory phase.
mental foramen – opening located on the anterior-lateral side of the mandibular body.
mental protuberance – inferior margin of anterior mandible that forms the chin.
Merkel cell – receptor cell in the stratum basale of the epidermis that responds to the sense of touch.
merocrine secretion – release of a substance from a gland via exocytosis.
mesangial – contractile cells found in the glomerulus; can contract or relax to regulate filtration rate.
mesencephalic nucleus – component of the trigeminal nuclei that is found in the midbrain.
mesencephalon – primary vesicle of the embryonic brain that does not significantly change through the rest of embryonic development and becomes the midbrain.
mesenchymal cell – adult stem cell from which most connective tissue cells are derived.
mesenchyme – embryonic tissue from which connective tissue cells derive.
mesoappendix – mesentery of the appendix.
mesoderm – middle embryonic germ layer from which connective tissue, muscle tissue, and some epithelial tissue derive.
mesothelium – simple squamous epithelial tissue which covers the major body cavities and is the epithelial portion of serous membranes.
messenger RNA (mRNA) – nucleotide molecule that serves as an intermediate in the genetic code between DNA and protein.
metabolic acidosis – condition wherein a deficiency of bicarbonate causes the blood to be overly acidic.
metabolic alkalosis – condition wherein an excess of bicarbonate causes the blood to be overly alkaline.
metabotropic receptor – neurotransmitter receptor that involves a complex of proteins that cause metabolic changes in a cell.
metacarpal bone – one of the five long bones that form the palm of the hand; numbered 1–5, starting on the lateral (thumb) side of the hand.
metacarpophalangeal joint – articulation between the distal end of a metacarpal bone of the hand and a proximal phalanx bone of the thumb or a finger.
metaphase – second stage of mitosis (and meiosis), characterised by the linear alignment of sister chromatids in the centre of the cell.
metaphase plate – linear alignment of sister chromatids in the centre of the cell, which takes place during metaphase.
metarteriole – short vessel arising from a terminal arteriole that branches to supply a capillary bed.
metastasis – spread of cancer cells from a source to other parts of the body.
metatarsal bone – one of the five elongated bones that forms the anterior half of the foot; numbered 1–5, starting on the medial side of the foot.
metatarsophalangeal joint – articulation between a metatarsal bone of the foot and the proximal phalanx bone of a toe.
metencephalon – secondary vesicle of the embryonic brain that develops into the pons and the cerebellum.
MHC class I – found on most cells of the body, it binds to the CD8 molecule on T cells.
MHC class II – found on macrophages, dendritic cells, and B cells, it binds to CD4 molecules on T cells.
MHC polygeny – multiple MHC genes and their proteins found in body cells.
MHC polymorphism – multiple alleles for each individual MHC locus.
micelle – tiny lipid-transport compound composed of bile salts and phospholipids with a fatty acid and monoacylglyceride core.
microcirculation – blood flow through the capillaries.
microfilament – the thinnest of the cytoskeletal filaments; composed of actin subunits that function in muscle contraction and cellular structural support.
microglia – glial cell type in the CNS that serves as the resident component of the immune system.
microscopic anatomy – study of very small structures of the body using magnification.
microtubule – the thickest of the cytoskeletal filaments, composed of tubulin subunits that function in cellular movement and structural support.
microvillus – small projection of the plasma membrane of the absorptive cells of the small intestinal mucosa.
micturition – also called urination or voiding
midbrain – middle region of the adult brain that develops from the mesencephalon.
midcarpal joint – articulation between the proximal and distal rows of the carpal bones; contributes to movements of the hand at the wrist.
middle cardiac vein – vessel that parallels and drains the areas supplied by the posterior interventricular artery; drains into the great cardiac vein.
middle cerebral artery – another branch of the internal carotid artery; supplies blood to the temporal and parietal lobes of the cerebrum.
middle cranial fossa – centrally located cranial fossa that extends from the lesser wings of the sphenoid bone to the petrous ridge.
middle ear – space within the temporal bone between the ear canal and bony labyrinth where the ossicles amplify sound waves from the tympanic membrane to the oval window.
middle nasal concha – nasal concha formed by the ethmoid bone that is located between the superior and inferior conchae.
middle sacral vein – drains the sacral region and leads to the left common iliac vein.
migrating motility complex – form of peristalsis in the small intestine.
mineralocorticoids – hormones produced by the zona glomerulosa cells of the adrenal cortex that influence fluid and electrolyte balance.
mitochondrion – one of the cellular organelles bound by a double lipid bilayer that function primarily in the production of cellular energy (ATP).
mitosis – division of genetic material, during which the cell nucleus breaks down and two new, fully functional, nuclei are formed.
mitotic phase – phase of the cell cycle in which a cell undergoes mitosis.
mitotic spindle – network of microtubules, originating from centrioles, that arranges and pulls apart chromosomes during mitosis.
mitral valve – (also, left atrioventricular valve or bicuspid valve) valve located between the left atrium and ventricle; consists of two flaps of tissue.
mixing wave – unique type of peristalsis that occurs in the stomach.
modelling – process, during bone growth, by which bone is resorbed on one surface of a bone and deposited on another.
moderator band – band of myocardium covered by endocardium that arises from the inferior portion of the interventricular septum in the right ventricle and crosses to the anterior papillary muscle; contains conductile fibres that carry electrical signals followed by contraction of the heart.
molar – tooth used for crushing and grinding food.
monocytes – agranular leukocytes of the myeloid stem cell line that circulate in the bloodstream; tissue monocytes are macrophages.
monomer – smallest unit of larger molecules that are polymers.
monosaccharide – single unit or monomer of carbohydrates.
mons pubis – mound of fatty tissue located at the front of the vulva.
morula – tightly packed sphere of blastomeres that has reached the uterus but has not yet implanted itself.
motilin – hormone that initiates migrating motility complexes.
motility – movement of food through the GI tract.
motor end-plate – sarcolemma of muscle fibre at the neuromuscular junction, with receptors for the neurotransmitter acetylcholine.
motor unit – motor neuron and the group of muscle fibres it innervates.
mucosa – innermost lining of the gastrointestinal tract.
mucosa-associated lymphoid tissue (MALT) – lymphoid nodule associated with the mucosa.
mucosal barrier – protective barrier that prevents gastric juice from destroying the stomach itself.
mucous connective tissue – specialised loose connective tissue present in the umbilical cord.
mucous gland – group of cells that secrete mucous, a thick, slippery substance that keeps tissues moist and acts as a lubricant.
mucous membrane – tissue membrane that is covered by protective mucous and lines tissue exposed to the outside environment.
mucous neck cell – gastric gland cell that secretes a uniquely acidic mucus.
Müllerian duct – duct system present in the embryo that will eventually form the internal female reproductive structures.
multiaxial joint – type of diarthrosis; a joint that allows for movements within three planes (three axes).
multimodal integration area – region of the cerebral cortex in which information from more than one sensory modality is processed to arrive at higher level cortical functions such as memory, learning, or cognition.
multipolar – shape of a neuron that has multiple processes—the axon and two or more dendrites.
multipotent – describes the condition of being able to differentiate into different types of cells within a given cell lineage or small number of lineages, such as a red blood cell or white blood cell.
muscarinic receptor – type of acetylcholine receptor protein that is characterised by also binding to muscarine and is a metabotropic receptor.
muscle tissue – type of tissue that is capable of contracting and generating tension in response to stimulation; produces movement.
muscle tension – force generated by the contraction of the muscle; tension generated during isotonic contractions and isometric contractions.
muscle tone – low levels of muscle contraction that occur when a muscle is not producing movement.
muscular artery – (also, distributing artery) artery with abundant smooth muscle in the tunica media that branches to distribute blood to the arteriole network.
muscularis – muscle (skeletal or smooth) layer of the gastrointestinal tract wall.
murmur – unusual heart sound detected by auscultation; typically related to septal or valve defects.
mutation – change in the nucleotide sequence in a gene within a cell’s DNA.
myelencephalon – secondary vesicle of the embryonic brain that develops into the medulla.
myelin – layer of lipid inside some neuroglial cells that wraps around the axons of some neurons.
myelin sheath – lipid-rich layer of insulation that surrounds an axon, formed by oligodendrocytes in the CNS and Schwann cells in the PNS; facilitates the transmission of electrical signals.
myeloid stem cells – type of haemopoietic stem cell that gives rise to some formed elements, including erythrocytes, megakaryocytes that produce platelets, and a myeloblast lineage that gives rise to monocytes and three forms of granular leukocytes (neutrophils, eosinophils, and basophils).
myenteric plexus – (plexus of Auerbach) major nerve supply to gastrointestinal tract wall; controls motility.
mylohyoid line – bony ridge located along the inner (medial) surface of the mandibular body.
myoblast – muscle-forming stem cell.
myocardial conducting cells – specialised cells that transmit electrical impulses throughout the heart and trigger contraction by the myocardial contractile cells.
myocardial contractile cells – bulk of the cardiac muscle cells in the atria and ventricles that conduct impulses and contract to propel blood.
myocardium – thickest layer of the heart composed of cardiac muscle cells built upon a framework of primarily collagenous fibres and blood vessels that supply it and the nervous fibres that help to regulate it.
myocyte – muscle cells.
myofibril – long, cylindrical organelle that runs parallel within the muscle fibre and contains the sarcomeres.
myogenic mechanism – mechanism by which smooth muscle responds to stretch by contracting; an increase in blood pressure causes vasoconstriction and a decrease in blood pressure causes vasodilation so that blood flow downstream remains steady.
myogenic response – constriction or dilation in the walls of arterioles in response to pressures related to blood flow; reduces high blood flow or increases low blood flow to help maintain consistent flow to the capillary network.
myogram – instrument used to measure twitch tension.
myometrium – smooth muscle layer of uterus that allows for uterine contractions during labour and expulsion of menstrual blood.
myosin – protein that makes up most of the thick cylindrical myofilament within a sarcomere muscle fibre.
myotube – fusion of many myoblast cells.
N
nail bed – layer of epidermis upon which the nail body forms.
nail body – main keratinous plate that forms the nail.
nail cuticle – fold of epithelium that extends over the nail bed, also called the eponychium.
nail fold – fold of epithelium at that extend over the sides of the nail body, holding it in place.
nail root – part of the nail that is lodged deep in the epidermis from which the nail grows.
naïve lymphocyte – mature B or T cell that has not yet encountered antigen for the first time.
naris – (plural = nares) opening of the nostrils.
nasal bone – bone of the skull that lies under the root and bridge of the nose and is connected to the frontal and maxillary bones.
nasal cavity – opening through skull for passage of air.
nasal conchae – curved bony plates that project from the lateral walls of the nasal cavity; include the superior and middle nasal conchae, which are parts of the ethmoid bone, and the independent inferior nasal conchae bone.
nasal septum – wall composed of bone and cartilage that separates the left and right nasal cavities.
nasolacrimal canal – passage for drainage of tears that extends downward from the medial-anterior orbit to the nasal cavity, terminating behind the inferior nasal conchae.
nasopharynx – portion of the pharynx flanked by the conchae and oropharynx that serves as an airway.
natural killer (NK) cells – cytotoxic lymphocytes capable of recognising cells that do not express “self” proteins on their plasma membrane or that contain foreign or abnormal markers; provide generalised, nonspecific immunity.
navicular – tarsal bone that articulates posteriorly with the talus bone, laterally with the cuboid bone, and anteriorly with the medial, intermediate, and lateral cuneiform bones.
neck of the femur – narrowed region located inferior to the head of the femur.
neck of the radius – narrowed region immediately distal to the head of the radius.
neck of the rib – narrowed region of a rib, next to the rib head.
necrosis – accidental death of cells and tissues.
negative feedback – homeostatic mechanism that tends to stabilise an upset in the body’s physiological condition by preventing an excessive response to a stimulus, typically as the stimulus is removed.
negative inotropic factors – factors that negatively impact or lower heart contractility.
negative selection – selection against thymocytes in the thymus that react with self-antigen.
neonatal (congenital) hypothyroidism – condition characterised by cognitive deficits, short stature, and other signs and symptoms in people born to women who were iodine-deficient during pregnancy.
nephrons – functional units of the kidney that carry out all filtration and modification to produce urine; consist of renal corpuscles, proximal and distal convoluted tubules, and descending and ascending loops of Henle; drain into collecting ducts.
nerve – cord-like bundle of axons located in the peripheral nervous system that transmits sensory input and response output to and from the central nervous system.
nerve plexus – network of nerves without neuronal cell bodies included.
nervi vasorum – small nerve fibres found in arteries and veins that trigger contraction of the smooth muscle in their walls.
nervous tissue – type of tissue that is capable of sending and receiving impulses through electrochemical signals.
net filtration pressure (NFP) – force driving fluid out of the capillary and into the tissue spaces; equal to the difference of the capillary hydrostatic pressure and the blood colloidal osmotic pressure.
neural crest – tissue that detaches from the edges of the neural groove and migrates through the embryo to develop into peripheral structures of both nervous and non-nervous tissues.
neural fold – elevated edge of the neural groove.
neural groove – region of the neural plate that folds into the dorsal surface of the embryo and closes off to become the neural tube.
neural plate – thickened layer of neuroepithelium that runs longitudinally along the dorsal surface of an embryo and gives rise to nervous system tissue.
neural tube – precursor to structures of the central nervous system, formed by the invagination and separation of neuroepithelium.
neural tunic – layer of the eye that contains nervous tissue, namely the retina.
neurulation – embryonic process that establishes the central nervous system.
neuraxis – central axis to the nervous system, from the posterior to anterior ends of the neural tube; the inferior tip of the spinal cord to the anterior surface of the cerebrum.
neuroglia – supportive neural cells.
neurogenic shock – type of shock that occurs with cranial or high spinal injuries that damage the cardiovascular centres in the medulla oblongata or the nervous fibres originating from this region.
neuron – neural tissue cell that is primarily responsible for generating and propagating electrical signals into, within, and out of the nervous system.
neuropeptide – neurotransmitter type that includes protein molecules and shorter chains of amino acids.
neutralisation – inactivation of a virus by the binding of specific antibody.
neurotransmitter – signalling chemical released by nerve terminals that bind to and activate receptors on target cells.
neutrophils – granulocytes that stain with a neutral dye and are the most numerous of the leukocytes; especially active against bacteria.
nicotinic receptor – type of acetylcholine receptor protein that is characterised by also binding to nicotine and is an ionotropic receptor.
nociceptor – receptor cell that senses pain stimuli.
node of Ranvier – gap between two myelinated regions of an axon, allowing for strengthening of the electrical signal as it propagates down the axon.
nonspecific channel – channel that is not specific to one ion over another, such as a nonspecific cation channel that allows any positively charged ion across the membrane.
noradrenaline – secondary catecholamine hormone secreted by the adrenal medulla in response to short-term stress.
normal range – range of values around the set point that do not cause a reaction by the control centre.
notochord – rod-like structure along dorsal side of the early embryo; largely disappears during later development but does contribute to formation of the intervertebral discs.
nuchal ligament – expanded portion of the supraspinous ligament within the posterior neck; interconnects the spinous processes of the cervical vertebrae and attaches to the base of the skull.
nucleus – cell’s central organelle; contains the cell’s DNA.
nuclear envelope – membrane that surrounds the nucleus; consisting of a double lipid-bilayer.
nuclear pore – one of the small, protein-lined openings found scattered throughout the nuclear envelope.
nucleic acid – biological macromolecule that carries the cell’s genetic blueprint and carries instructions for the cell’s functioning.
nucleotide – monomer of nucleic acids; contains a pentose sugar, one or more phosphate groups, and a nitrogenous base.
nucleolus – small region of the nucleus that functions in ribosome synthesis.
nucleosidase – brush border enzyme that digests nucleotides.
nucleosome – unit of chromatin consisting of a DNA strand wrapped around histone proteins.
nucleus cuneatus – medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the upper body and arms.
nucleus gracilis – medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the lower body and legs.
nucleus pulposus – gel-like central region of an intervertebral disc; provides for padding, weight-bearing, and movement between adjacent vertebrae.
nutrient foramen – small opening in the middle of the external surface of the diaphysis, through which an artery enters the bone to provide nourishment.
O
obstructive shock – type of shock that occurs when a significant portion of the vascular system is blocked.
obturator foramen – large opening located in the anterior hip bone, between the pubis and ischium regions.
occipital bone – unpaired bone that forms the posterior portions of the brain case and base of the skull.
occipital condyle – paired, oval-shaped bony knobs located on the inferior skull, to either side of the foramen magnum.
occipital lobe – region of the cerebral cortex directly beneath the occipital bone of the cranium.
occipital sinus – enlarged vein that drains the occipital region near the falx cerebelli and flows into the left and right transverse sinuses, and also into the vertebral veins.
oculomotor nerve – third cranial nerve; responsible for contraction of four of the extraocular muscles, the muscle in the upper eyelid, and pupillary constriction.
odourant molecules – volatile chemicals that bind to receptor proteins in olfactory neurons to stimulate the sense of smell.
oesophagus – muscular tube that runs from the pharynx to the stomach.
oesophageal artery – branch of the thoracic aorta; supplies blood to the oesophagus.
oesophageal plexus – neuronal plexus in the wall of the oesophagus that is part of the enteric nervous system.
oesophageal vein – drains the inferior portions of the oesophagus and leads to the azygos vein.
oestrogens – class of predominantly female sex hormones important for the development and growth of the female reproductive tract, secondary sex characteristics, the female reproductive cycle, and the maintenance of pregnancy.
olecranon fossa – large depression located on the posterior side of the distal humerus; this space receives the olecranon process of the ulna when the elbow is fully extended.
olecranon process – expanded posterior and superior portions of the proximal ulna; forms the bony tip of the elbow.
olfaction – special sense responsible for smell, which has a unique, direct connection to the cerebrum.
olfactory bulb – central target of the first cranial nerve; located on the ventral surface of the frontal lobe in the cerebrum.
olfactory epithelium – region of the nasal epithelium where olfactory neurons are located.
olfactory nerve – first cranial nerve; responsible for the sense of smell.
olfactory pit – invaginated ectodermal tissue in the anterior portion of the head region of an embryo that will form the nasal cavity.
olfactory sensory neuron – receptor cell of the olfactory system, sensitive to the chemical stimuli of smell, the axons of which compose the first cranial nerve.
oligodendrocyte – neuroglial cell that produces myelin in the brain.
oligopotent – describes the condition of being more specialised than multipotency; the condition of being able to differentiate into one of a few possible cell types.
oliguria – below normal urine production of 400–500 mL/day.
omega fat – type of polyunsaturated fat that the body requires; numbering the carbon omega starts from the methyl end or the end that is farthest from the carboxylic end.
oocyte – cell that results from the division of the oogonium and undergoes meiosis I at the LH surge and meiosis II at fertilisation to become a haploid ovum.
oogenesis – process by which oogonia divide by mitosis to primary oocytes, which undergo meiosis to produce the secondary oocyte and, upon fertilisation, the ovum.
oogonia – ovarian stem cells that undergo mitosis during female foetal development to form primary oocytes.
open reduction – surgical exposure of a bone to reset a fracture.
ophthalmic artery – branch of the internal carotid artery; supplies blood to the eyes.
opposition – thumb movement that brings the tip of the thumb in contact with the tip of a finger.
opsin – protein that contains the photosensitive cofactor retinal for phototransduction.
opsonisation – enhancement of phagocytosis by the binding of antibody or antimicrobial protein.
optic canal – opening spanning between middle cranial fossa and posterior orbit.
optic chiasm – decussation point in the visual system at which medial retina fibres cross to the other side of the brain.
optic disc – spot on the retina at which RGC axons leave the eye and blood vessels of the inner retina pass.
optic nerve – second cranial nerve; responsible for visual sensation.
optic tract – name for the fibre structure containing axons from the retina posterior to the optic chiasm representing their CNS location.
orbit – bony socket that contains the eyeball and associated muscles.
oral cavity – (also, buccal cavity) mouth.
oral vestibule – part of the mouth bounded externally by the cheeks and lips, and internally by the gums and teeth.
organ – functionally distinct structure composed of two or more types of tissues.
organelle – any of several different types of membrane-enclosed specialised structures in the cell that perform specific functions for the cell.
organism – living being that has a cellular structure and that can independently perform all physiologic functions necessary for life.
organ of Corti – structure in the cochlea in which hair cells transduce movements from sound waves into electrochemical signals.
organogenesis – development of the rudimentary structures of all of an embryo’s organs from the germ layers.
organ system – group of organs that work together to carry out a particular function.
oropharynx – portion of the pharynx flanked by the nasopharynx, oral cavity, and laryngopharynx that is a passageway for both air and food.
orthopaedist – doctor who specialises in diagnosing and treating musculoskeletal disorders and injuries.
orthostatic reflex – sympathetic function that maintains blood pressure when standing to offset the increased effect of gravity.
osmoreceptor – receptor cell that senses differences in the concentrations of bodily fluids on the basis of osmotic pressure.
osmosis – diffusion of water molecules down their concentration gradient across a selectively permeable membrane.
osseous tissue – bone tissue; a hard, dense connective tissue that forms the structural elements of the skeleton.
ossicles – three small bones in the middle ear.
ossification – (also, osteogenesis) bone formation.
ossification centre – cluster of osteoblasts found in the early stages of intramembranous ossification.
osteoblast – cell responsible for forming new bone.
osteoclast – cell responsible for resorbing bone.
osteocyte – primary cell in mature bone; responsible for maintaining the matrix.
osteogenic cell – undifferentiated cell with high mitotic activity; the only bone cells that divide; they differentiate and develop into osteoblasts.
osteoid – uncalcified bone matrix secreted by osteoblasts.
osteomalacia – softening of bones due to a lack of mineralisation with calcium and phosphate; most often due to lack of vitamin D; in children, osteomalacia is termed rickets; not to be confused with osteoporosis.
osteon – (also, Haversian system) basic structural unit of compact bone; made of concentric layers of calcified matrix.
osteoporosis – disease characterised by a decrease in bone mass; occurs when the rate of bone resorption exceeds the rate of bone formation, a common occurrence as the body ages.
otolith – layer of calcium carbonate crystals located on top of the otolithic membrane.
otolithic membrane – gelatinous substance in the utricle and saccule of the inner ear that contains calcium carbonate crystals and into which the stereocilia of hair cells are embedded.
outer segment – in the eye, the section of a photoreceptor that contains opsin molecules that transduce light stimuli.
outer synaptic layer – layer in the retina at which photoreceptors connect to bipolar cells
oval window – membrane at the base of the cochlea where the stapes attaches, marking the beginning of the scala vestibuli.
ovarian artery – branch of the abdominal aorta; supplies blood to the ovary, uterine (Fallopian) tube, and uterus.
ovarian cycle – approximately 28-day cycle of changes in the ovary consisting of a follicular phase and a luteal phase.
ovarian vein – drains the ovary; the right ovarian vein leads to the inferior vena cava and the left ovarian vein leads to the left renal vein.
ovaries – female gonads that produce oocytes and sex steroid hormones (notably oestrogen and progesterone).
ovulation – release of a secondary oocyte and associated granulosa cells from an ovary.
ovum – haploid female gamete resulting from completion of meiosis II at fertilisation.
oxyhaemoglobin – molecule of haemoglobin to which oxygen is bound.
oxygen debt – amount of oxygen needed to compensate for ATP produced without oxygen during muscle contraction.
oxygen– haemoglobin dissociation curve – graph that describes the relationship of partial pressure to the binding and disassociation of oxygen to and from haem.
oxytocin – hypothalamic hormone stored in the posterior pituitary gland and important in stimulating uterine contractions in labour, milk ejection during breastfeeding, and feelings of attachment (also produced in males).
P
pacemaker – cluster of specialised myocardial cells known as the SA node that initiates the sinus rhythm.
pacesetter cell – cell that triggers action potentials in smooth muscle.
Pacinian corpuscle – (also, lamellated corpuscle) receptor in the skin that responds to vibration.
packed cell volume (PCV) – (also, haematocrit) volume percentage of erythrocytes present in a sample of centrifuged blood.
palatine bone – paired bones that form the posterior quarter of the hard palate and a small area in floor of the orbit.
palatine process – medial projection from the maxilla bone that forms the anterior three quarters of the hard palate.
palatine tonsil – one of the paired structures composed of lymphoid tissue located anterior to the uvula at the roof of isthmus of the fauces.
palatoglossal arch – muscular fold that extends from the lateral side of the soft palate to the base of the tongue.
palatopharyngeal arch – muscular fold that extends from the lateral side of the soft palate to the side of the pharynx.
palmar arches – superficial and deep arches formed from anastomoses of the radial and ulnar arteries; supply blood to the hand and digital arteries.
palmar venous arches – drain the hand and digits, and feed into the radial and ulnar veins.
palpebral conjunctiva – membrane attached to the inner surface of the eyelids that covers the anterior surface of the cornea.
pancreas – accessory digestive organ that secretes pancreatic secretion.
pancreatic amylase – enzyme secreted by the pancreas that completes the chemical digestion of carbohydrates in the small intestine.
pancreatic islets – specialised clusters of pancreatic cells that have endocrine functions; also called islets of Langerhans.
pancreatic lipase – enzyme secreted by the pancreas that participates in lipid digestion.
pancreatic nuclease – enzyme secreted by the pancreas that participates in nucleic acid digestion.
pancreatic secretion – secretion of the pancreas containing digestive enzymes and bicarbonate.
papilla – for gustation, a bump-like projection on the surface of the tongue that contains taste buds.
papillary layer – superficial layer of the dermis, made of loose, areolar connective tissue.
papillary muscle – extension of the myocardium in the ventricles to which the chordae tendineae attach.
paracrine – chemical signal that elicits a response in neighbouring cells; also called paracrine factor.
paranasal sinus – one of the cavities within the skull that is connected to the conchae that serve to warm and humidify incoming air, produce mucus, and lighten the weight of the skull; consists of frontal, maxillary, sphenoidal, and ethmoidal sinuses.
parathyroid glands – small, round glands embedded in the posterior thyroid gland that produce parathyroid hormone (PTH).
parathyroid hormone (PTH) – peptide hormone produced and secreted by the parathyroid glands in response to low blood calcium levels.
paravertebral ganglia – autonomic ganglia superior to the sympathetic chain ganglia.
parenchyma – functional cells of a gland or organ, in contrast with the supportive or connective tissue of a gland or organ.
parietal bone – paired bones that form the upper, lateral sides of the skull.
parietal branches – (also, somatic branches) group of arterial branches of the thoracic aorta; includes those that supply blood to the thoracic cavity, vertebral column, and the superior surface of the diaphragm.
parietal cell – gastric gland cell that secretes hydrochloric acid and intrinsic factor.
parietal lobe – region of the cerebral cortex directly beneath the parietal bone of the cranium.
parietal pleura – outermost layer of the pleura that connects to the thoracic wall, mediastinum, and diaphragm.
parieto-occipital sulcus – groove in the cerebral cortex representing the border between the parietal and occipital cortices.
parotid gland – one of a pair of major salivary glands located inferior and anterior to the ears.
partial pressure – force exerted by each gas in a mixture of gases.
partial thickness burns – partial-thickness burn that injures the epidermis and a portion of the dermis.
passive immunity – transfer of immunity to a pathogen to an individual that lacks immunity to this pathogen usually by the injection of antibodies.
passive transport – form of transport across the cell membrane that does not require input of cellular energy.
patella – kneecap; the largest sesamoid bone of the body; articulates with the distal femur.
patellar ligament – ligament spanning from the patella to the anterior tibia; serves as the final attachment for the quadriceps femoris muscle.
patellar surface – smooth groove located on the anterior side of the distal femur, between the medial and lateral condyles; site of articulation for the patella.
pattern recognition receptor (PRR) – leukocyte receptor that binds to specific cell wall components of different bacterial species.
pectinate line – horizontal line that runs like a ring, perpendicular to the inferior margins of the anal sinuses.
pectinate muscles – muscular ridges seen on the anterior surface of the right atrium.
pectineal line – narrow ridge located on the superior surface of the superior pubic ramus.
pectoral girdle – shoulder girdle; the set of bones, consisting of the scapula and clavicle, which attaches each upper limb to the axial skeleton.
pedicels – finger-like projections of podocytes surrounding glomerular capillaries; interdigitate to form a filtration membrane.
pedicle – portion of the vertebral arch that extends from the vertebral body to the transverse process.
pelvic brim – pelvic inlet; the dividing line between the greater and lesser pelvic regions; formed by the superior margin of the pubic symphysis, the pectineal lines of each pubis, the arcuate lines of each ilium, and the sacral promontory.
pelvic girdle – hip girdle; consists of a single hip bone, which attaches a lower limb to the sacrum of the axial skeleton.
pelvic inlet – pelvic brim.
pelvic outlet – inferior opening of the lesser pelvis; formed by the inferior margin of the pubic symphysis, right and left ischiopubic rami and sacrotuberous ligaments, and the tip of the coccyx.
pelvis – ring of bone consisting of the right and left hip bones, the sacrum, and the coccyx.
penis – male organ of copulation.
pepsinogen – inactive form of pepsin.
peptide bond – bond formed between two amino acids by a dehydration reaction.
perforating canal – (also, Volkmann’s canal) channel that branches off from the central canal and houses vessels and nerves that extend to the periosteum and endosteum.
perforin – molecule in NK cell and cytotoxic T cell granules that form pores in the membrane of a target cell.
perfusion – distribution of blood into the capillaries so the tissues can be supplied.
pericardial artery – branch of the thoracic aorta; supplies blood to the pericardium.
pericardial cavity – cavity surrounding the heart filled with a lubricating serous fluid that reduces friction as the heart contracts.
pericardial sac – (also, pericardium) membrane that separates the heart from other mediastinal structures; consists of two distinct, fused sublayers: the fibrous pericardium and the parietal pericardium.
pericardium – (also, pericardial sac) membrane that separates the heart from other mediastinal structures; consists of two distinct, fused sublayers: the fibrous pericardium and the parietal pericardium.
perichondrium – membrane that covers cartilage.
pericyte – stem cell that regenerates smooth muscle cells.
perimetrium – outer epithelial layer of uterine wall.
perimysium – connective tissue that bundles skeletal muscle fibres into fascicles within a skeletal muscle.
perineurium – layer of connective tissue surrounding fascicles within a nerve.
periodontal ligament – band of dense connective tissue that anchors the root of a tooth into the bony jaw socket.
peripheral chemoreceptor – one of the specialised receptors located in the aortic arch and carotid arteries that sense changes in pH, carbon dioxide, or oxygen blood levels.
peripheral nervous system (PNS) – anatomical division of the nervous system that is largely outside the cranial and vertebral cavities, namely all parts except the brain and spinal cord.
peripheral protein – membrane-associated protein that does not span the width of the lipid bilayer, but is attached peripherally to integral proteins, membrane lipids, or other components of the membrane.
peripheral tolerance – mature B cell made tolerant by lack of T cell help.
periosteum – fibrous membrane covering the outer surface of bone and continuous with ligaments.
peristalsis – muscular contractions and relaxations that propel food through the GI tract.
peritoneum – serous membrane that lines the abdominopelvic cavity and covers the organs found there.
peritubular capillaries – second capillary bed of the renal portal system; surround the proximal and distal convoluted tubules; associated with the vasa recta
permanent tooth – one of 32 adult teeth.
peroxisome – membrane-bound organelle that contains enzymes primarily responsible for detoxifying harmful substances.
perpendicular plate of the ethmoid bone – downward, midline extension of the ethmoid bone that forms the superior portion of the nasal septum.
petrosal sinus – enlarged vein that receives blood from the cavernous sinus and flows into the internal jugular vein.
petrous ridge – petrous portion of the temporal bone that forms a large, triangular ridge in the floor of the cranial cavity, separating the middle and posterior cranial fossae; houses the middle and inner ear structures.
phagocytosis – endocytosis of large particles.
phalanx bone of the foot – (plural = phalanges) one of the 14 bones that form the toes; these include the proximal and distal phalanges of the big toe, and the proximal, middle, and distal phalanx bones of toes two through five.
phalanx bone of the hand – (plural = phalanges) one of the 14 bones that form the thumb and fingers; these include the proximal and distal phalanges of the thumb, and the proximal, middle, and distal phalanx bones of the fingers two through five.
pharyngeal tonsil – structure composed of lymphoid tissue located in the nasopharynx.
pharynx – region of the conducting zone that forms a tube of skeletal muscle lined with respiratory epithelium; located between the nasal conchae and the oesophagus and trachea.
philtrum – concave surface of the face that connects the apex of the nose to the top lip.
phosphatase – brush border enzyme that digests nucleotides.
phosphodiester – linkage covalent chemical bond that holds together the polynucleotide chains with a phosphate group linking neighbouring nucleotides’ two pentose sugars.
phosphodiesterase (PDE) – cytosolic enzyme that deactivates and degrades cAMP.
photoisomerisation – chemical change in the retinal molecule that alters the bonding so that it switches from the 11-cis-retinal isomer to the all-trans-retinal isomer.
phosphorylation cascade – signalling event in which multiple protein kinases phosphorylate the next protein substrate by transferring a phosphate group from ATP to the protein.
photon – individual “packet” of light.
photoreceptor – receptor cell specialised to respond to light stimuli.
phrenic nerve – systemic nerve from the cervical plexus that enervates the diaphragm.
phrenic vein – drains the diaphragm; the right phrenic vein flows into the inferior vena cava and the left phrenic vein leads to the left renal vein.
physiology – science that studies the chemistry, biochemistry, and physics of the body’s functions.
physiological sphincter – sphincter consisting of circular smooth muscle indistinguishable from adjacent muscle but possessing differential innervations, permitting its function as a sphincter; structurally weak
pia mater – thin, innermost membrane of the meninges that directly covers the surface of the CNS.
pineal gland – endocrine gland that secretes melatonin, which is important in regulating the sleep-wake cycle.
pinealocyte – cell of the pineal gland that produces and secretes the hormone melatonin.
pinocytosis – endocytosis of fluid.
pisiform – from the lateral side, the fourth of the four proximal carpal bones; articulates with the anterior surface of the triquetrum.
pituitary dwarfism – disorder in children caused when abnormally low levels of GH result in growth retardatio.
pituitary gland – bean-sized organ suspended from the hypothalamus that produces, stores, and secretes hormones in response to hypothalamic stimulation (also called hypophysis).
pivot joint – synovial joint at which the rounded portion of a bone rotates within a ring formed by a ligament and an articulating bone; functionally classified as uniaxial joint.
placenta – organ that forms during pregnancy to nourish the developing foetus; also regulates waste and gas exchange between mother and foetus.
placenta previa – low placement of foetus within uterus causes placenta to partially or completely cover the opening of the cervix as it grows.
placentation – formation of the placenta; complete by weeks 14–16 of pregnancy.
plane – imaginary two-dimensional surface that passes through the body.
plane joint – synovial joint formed between the flattened articulating surfaces of adjacent bones; functionally classified as a multiaxial joint.
plantar arch – formed from the anastomosis of the dorsalis pedis artery and medial and plantar arteries; branches supply the distal portions of the foot and digits.
plantar flexion – foot movement at the ankle in which the heel is lifted off of the ground.
plantar veins – drain the foot and lead to the plantar venous arch.
plantar venous arch – formed from the plantar veins; leads to the anterior and posterior tibial veins through anastomoses.
plasma – in blood, the liquid extracellular matrix composed mostly of water that circulates the formed elements and dissolved materials throughout the cardiovascular system.
plasma cell – differentiated B cell that is actively secreting antibody.
plasma osmolality – ratio of solutes to a volume of solvent in the plasma; plasma osmolality reflects a person’s state of hydration.
plasmin – plasma blood protein active in fibrinolysis.
platelets – (also, thrombocytes) one of the formed elements of blood that consists of cell fragments broken off from megakaryocytes.
platelet plug – adhesion and aggregation of platelets at the site of blood vessel injury.
pleura – serous membrane that lines the pleural cavity and covers the lungs.
pleural cavity – space between the visceral and parietal pleurae.
pleural fluid – substance that acts as a lubricant for the visceral and parietal layers of the pleura during the movement of breathing.
plexus – network of nerves or nervous tissue.
pluripotent – describes the condition of being able to differentiate into a large variety of cell types.
pluripotent stem cell – stem cell that derives from totipotent stem cells and is capable of differentiating into many, but not all, cell types.
pneumotaxic centre – network of neurons within the pons that inhibit the activity of the neurons in the dorsal respiratory group; controls rate of breathing.
podocytes – cells forming finger-like processes; form the visceral layer of Bowman’s capsule; pedicels of the podocytes interdigitate to form a filtration membrane.
polar body – smaller cell produced during the process of meiosis in oogenesis.
pollex – (also, thumb) digit 1 of the hand.
polyclonal response – response by multiple clones to a complex antigen with many determinants.
polycythaemia – elevated level of haemoglobin and RBC, whether adaptive or pathological.
polymer – chain of monomer residues that covalent bonds link; polymerisation is the process of polymer formation from monomers by condensation.
polymorphonuclear – having a lobed nucleus, as seen in some leukocytes.
polynucleotide – long chain of nucleotides.
polypeptide – chain of amino acids linked by peptide bonds.
polyribosome – simultaneous translation of a single mRNA transcript by multiple ribosomes.
polysaccharide – long chain of monosaccharides; may be branched or unbranched.
polyspermy – penetration of an oocyte by more than one sperm.
polyuria – urine production in excess of 2.5 L/day; may be caused by diabetes insipidus, diabetes mellitus, or excessive use of diuretics.
popliteal artery – continuation of the femoral artery posterior to the knee; branches into the anterior and posterior tibial arteries.
popliteal vein – continuation of the femoral vein behind the knee; drains the region behind the knee and forms from the fusion of the fibular and anterior and posterior tibial veins.
porta hepatis – “gateway to the liver” where the hepatic artery and hepatic portal vein enter the liver.
portal triad – bile duct, hepatic artery branch, and hepatic portal vein branch.
postcentral gyrus – ridge just posterior to the central sulcus, in the parietal lobe, where somatosensory processing initially takes place in the cerebrum.
posterior – describes the back or direction toward the back of the body; also referred to as dorsal.
posterior arch – posterior portion of the ring-like C1 (atlas) vertebra.
posterior cardiac vein – vessel that parallels and drains the areas supplied by the marginal artery branch of the circumflex artery; drains into the great cardiac vein.
posterior cavity – posterior body cavity that houses the brain and spinal cord; also referred to as dorsal cavity.
posterior cerebral artery – branch of the basilar artery that forms a portion of the posterior segment of the arterial circle; supplies blood to the posterior portion of the cerebrum and brain stem.
posterior columns – white matter of the spinal cord that lies between the posterior horns of the grey matter, sometimes referred to as the dorsal column; composed of axons of ascending tracts that carry sensory information up to the brain.
posterior communicating artery – branch of the posterior cerebral artery that forms part of the posterior portion of the arterial circle; supplies blood to the brain.
posterior cranial fossa – deepest and most posterior cranial fossa; extends from the petrous ridge to the occipital bone.
posterior cruciate ligament – intracapsular ligament of the knee; extends from the posterior, superior surface of the tibia to the inner aspect of the medial condyle of the femur; prevents anterior displacement of the femur when the knee is flexed and weight bearing.
posterior horn – grey matter region of the spinal cord in which sensory input arrives, sometimes referred to as the dorsal horn.
posterior inferior iliac spine – small, bony projection located at the inferior margin of the auricular surface on the posterior ilium.
posterior interventricular artery – (also, posterior descending artery) branch of the right coronary artery that runs along the posterior portion of the interventricular sulcus toward the apex of the heart and gives rise to branches that supply the interventricular septum and portions of both ventricles.
posterior interventricular sulcus – sulcus located between the left and right ventricles on the anterior surface of the heart.
posterior longitudinal ligament – ligament that runs the length of the vertebral column, uniting the posterior sides of the vertebral bodies.
posterior median sulcus – midline feature of the posterior spinal cord, marking the separation between right and left sides of the cord.
posterior (dorsal) sacral foramen – one of the series of paired openings located on the posterior (dorsal) side of the sacrum.
posterior sacroiliac ligament – strong ligament spanning the sacrum and ilium of the hip bone that supports the posterior side of the sacroiliac joint.
posterior superior iliac spine – rounded, posterior end of the iliac crest.
posterior talofibular ligament – intrinsic ligament located on the lateral side of the ankle joint, between the talus bone and lateral malleolus of the fibula; supports the talus at the talocrural joint and resists excess inversion of the foot.
posterior tibial artery – branch from the popliteal artery that gives rise to the fibular or peroneal artery; supplies blood to the posterior tibial region.
posterior tibial vein – forms from the dorsal venous arch; drains the area near the posterior surface of the tibia and leads to the popliteal vein.
posterolateral sulcus – feature of the posterior spinal cord marking the entry of posterior nerve roots and the separation between the posterior and lateral columns of the white matter.
positive chemotaxis – process in which a cell is attracted to move in the direction of chemical stimuli.
positive feedback – mechanism that intensifies a change in the body’s physiological condition in response to a stimulus.
positive inotropic factors – factors that positively impact or increase heart contractility.
positive selection – selection of thymocytes within the thymus that interact with self, but not non-self, MHC molecules.
postsynaptic potential (PSP) – graded potential in the postsynaptic membrane caused by the binding of neurotransmitter to protein receptors.
power stroke – action of myosin pulling actin inward (toward the M line).
precapillary sphincters – circular rings of smooth muscle that surround the entrance to a capillary and regulate blood flow into that capillary.
precentral gyrus – primary motor cortex located in the frontal lobe of the cerebral cortex.
precentral gyrus of the frontal cortex – region of the cerebral cortex responsible for generating motor commands, where the upper motor neuron cell body is located.
prefrontal lobe – specific region of the frontal lobe anterior to the more specific motor function areas, which can be related to the early planning of movements and intentions to the point of being personality-type functions.
preload – (also, end diastolic volume) amount of blood in the ventricles at the end of atrial systole just prior to ventricular contraction.
premolar – (also, bicuspid) transitional tooth used for mastication, crushing, and grinding food.
premotor area – region of the frontal lobe responsible for planning movements that will be executed through the primary motor cortex.
premotor cortex – cortical area anterior to the primary motor cortex that is responsible for planning movements.
prepotential depolarisation – (also, spontaneous depolarisation) mechanism that accounts for the autorhythmic property of cardiac muscle; the membrane potential increases as sodium ions diffuse through the always-open sodium ion channels and causes the electrical potential to rise.
prepuce – (also, foreskin) flap of skin that forms a collar around, and thus protects and lubricates, the glans penis; also referred as the foreskin.
prevertebral ganglia – autonomic ganglia that are anterior to the vertebral column and functionally related to the sympathetic chain ganglia.
primary adaptive response – immune system’s response to the first exposure to a pathogen.
primary curve – anteriorly concave curvatures of the thoracic and sacrococcygeal regions that are retained from the original foetal curvature of the vertebral column.
primary follicles – ovarian follicles with a primary oocyte and one layer of cuboidal granulosa cells.
primary lymphoid organ – site where lymphocytes mature and proliferate; red bone marrow and thymus gland.
primary ossification centre – region, deep in the periosteal collar, where bone development starts during endochondral ossification.
primary sensory cortex – region of the cerebral cortex that initially receives sensory input from an ascending pathway from the thalamus and begins the processing that will result in conscious perception of that modality.
primary structure – linear sequence of amino acids in a protein.
primary union – condition of a wound where the wound edges are close enough to be brought together and fastened if necessary, allowing quicker and more thorough healing.
primary vesicle – initial enlargements of the anterior neural tube during embryonic development that develop into the forebrain, midbrain, and hindbrain.
primitive atrium – portion of the primitive heart tube that eventually becomes the anterior portions of both the right and left atria, and the two auricles.
primitive heart tube – singular tubular structure that forms from the fusion of the two endocardial tubes.
primitive streak – indentation along the dorsal surface of the epiblast through which cells migrate to form the endoderm and mesoderm during gastrulation.
primitive ventricle – portion of the primitive heart tube that eventually forms the left ventricle.
primordial follicles – least developed ovarian follicles that consist of a single oocyte and a single layer of flat (squamous) granulosa cells.
principal cell – found in collecting ducts and possess channels for the recovery or loss of sodium and potassium; under the control of aldosterone; also have aquaporin channels under ADH control to regulate recovery of water.
process – in cells, an extension of a cell body; in the case of neurons, this includes the axon and dendrites.
progesterone – predominantly female sex hormone important in regulating the female reproductive cycle and the maintenance of pregnancy.
projection – bone markings where part of the surface sticks out above the rest of the surface, where tendons and ligaments attach.
prolactin (PRL) – anterior pituitary hormone that promotes development of the mammary glands and the production of breast milk.
proliferative phase – phase of the menstrual cycle in which the endometrium proliferates.
proliferative zone – region of the epiphyseal plate that makes new chondrocytes to replace those that die at the diaphyseal end of the plate and contributes to longitudinal growth of the epiphyseal plate.
promoter – region of DNA that signals transcription to begin at that site within the gene.
pronated position – forearm position in which the palm faces backward.
pronation – forearm motion that moves the palm of the hand from the palm forward to the palm backward position.
prone – face down.
propagation – movement of an action potential along the length of an axon.
prophase – first stage of mitosis (and meiosis), characterised by breakdown of the nuclear envelope and condensing of the chromatin to form chromosomes.
proprioception – general sensory perceptions providing information about location and movement of body parts; the “sense of the self”.
proprioceptor – receptor cell that senses changes in the position and kinaesthetic aspects of the body.
propulsion – voluntary process of swallowing and the involuntary process of peristalsis that moves food through the digestive tract.
prosencephalon – primary vesicle of the embryonic brain that develops into the forebrain, which includes the cerebrum and diencephalon.
prostate gland – doughnut-shaped gland at the base of the bladder surrounding the urethra and contributing fluid to semen during ejaculation.
protein – biological macromolecule comprised of one or more amino acid chains.
protein kinase – enzyme that initiates a phosphorylation cascade upon activation.
proteome – full complement of proteins produced by a cell (determined by the cell’s specific gene expression).
protraction – anterior motion of the scapula or mandible.
proximal – describes a position nearer to the point of attachment or the trunk of the body.
proximal convoluted tubules (PCTs) – tortuous tubules receiving filtrate from Bowman’s capsule; most active part of the nephron in reabsorption and secretion.
proximal radioulnar joint – articulation formed by the radial notch of the ulna and the head of the radius.
proximal tibiofibular joint – articulation between the head of the fibula and the inferior aspect of the lateral condyle of the tibia.
pseudostratified columnar epithelium – tissue that consists of a single layer of irregularly shaped and sized cells that give the appearance of multiple layers; found in ducts of certain glands and the upper respiratory tract.
psychoneuroimmunology – study of the connections between the immune, nervous, and endocrine systems.
pterion – H-shaped suture junction region that unites the frontal, parietal, temporal, and sphenoid bones on the lateral side of the skull.
puberty – life stage during which a male or female adolescent becomes anatomically and physiologically capable of reproduction.
pubic arch – bony structure formed by the pubic symphysis, and the bodies and inferior pubic rami of the right and left pubic bones.
pubic body – enlarged, medial portion of the pubis region of the hip bone.
pubic symphysis – joint formed by the articulation between the pubic bodies of the right and left hip bones.
pubic tubercle – small bump located on the superior aspect of the pubic body.
pubis – anterior portion of the hip bone.
pubofemoral ligament – intrinsic ligament spanning from the pubis of the hip bone to the femur, on the anterior-inferior aspect of the hip joint.
pulmonary arteries – left and right branches of the pulmonary trunk that carry deoxygenated blood from the heart to each of the lungs.
pulmonary capillaries – capillaries surrounding the alveoli of the lungs where gas exchange occurs: carbon dioxide exits the blood and oxygen enters.
pulmonary circuit – blood flow to and from the lungs.
pulmonary plexus – network of autonomic nervous system fibres found near the hilum of the lung.
pulmonary surfactant – substance composed of phospholipids and proteins that reduces the surface tension of the alveoli; made by type II alveolar cells.
pulmonary trunk – large arterial vessel that carries blood ejected from the right ventricle; divides into the left and right pulmonary arteries.
pulmonary valve – (also, pulmonary semilunar valve, the pulmonic valve, or the right semilunar valve) valve at the base of the pulmonary trunk that prevents backflow of blood into the right ventricle; consists of three flaps.
pulmonary veins – veins that carry highly oxygenated blood into the left atrium, which pumps the blood into the left ventricle, which in turn pumps oxygenated blood into the aorta and to the many branches of the systemic circuit.
pulmonary ventilation – exchange of gases between the lungs and the atmosphere; breathing.
pulp cavity – deepest portion of a tooth, containing nerve endings and blood vessels.
pulse – alternating expansion and recoil of an artery as blood moves through the vessel; an indicator of heart rate.
pulse pressure – difference between the systolic and diastolic pressures.
pupil – open hole at the centre of the iris that light passes through into the eye.
purine – type of nitrogenous base in DNA and RNA; adenine and guanine are purines.
Purkinje fibres – specialised myocardial conduction fibres that arise from the bundle branches and spread the impulse to the myocardial contraction fibres of the ventricles.
putamen – nucleus deep in the cerebrum that is part of the basal nuclei; along with the caudate, it is part of the striatum.
P wave – component of the electrocardiogram that represents the depolarisation of the atria.
pyloric antrum – wider, more superior part of the pylorus.
pyloric canal – narrow, more inferior part of the pylorus.
pyloric sphincter – sphincter that controls stomach emptying.
pylorus – lower, funnel-shaped part of the stomach that is continuous with the duodenum.
pyramidal decussation – location at which corticospinal tract fibres cross the midline and segregate into the anterior and lateral divisions of the pathway.
pyramids – segment of the descending motor pathway that travels in the anterior position of the medulla.
pyruvic acid – product of glycolysis that can be used in aerobic respiration or converted to lactic acid.
Q
quaternary structure – association of discrete polypeptide subunits in a protein.
quickening – foetal movements that are strong enough to be felt by the mother
quiet breathing – (also, eupnoea) mode of breathing that occurs at rest and does not require the cognitive thought of the individual.
QRS complex – component of the electrocardiogram that represents the depolarisation of the ventricles and includes, as a component, the repolarisation of the atria.
R
radial artery – formed at the bifurcation of the brachial artery; parallels the radius; gives off smaller branches until it reaches the carpal region where it fuses with the ulnar artery to form the superficial and deep palmar arches; supplies blood to the lower arm and carpal region.
radial collateral ligament – intrinsic ligament on the lateral side of the elbow joint; runs from the lateral epicondyle of humerus to merge with the annular ligament.
radial fossa (anatomical snuff box)– small depression located on the anterior humerus above the capitulum; this space receives the head of the radius when the elbow is maximally flexed sometimes also known as the anatomical snuff box.
radial nerve – systemic nerve of the arm, the distal component of which is located near the radial bone.
radial notch of the ulna – small, smooth area on the lateral side of the proximal ulna; articulates with the head of the radius as part of the proximal radioulnar joint.
radial tuberosity – oval-shaped, roughened protuberance located on the medial side of the proximal radius.
radial vein – parallels the radius and radial artery; arises from the palmar venous arches and leads to the brachial vein.
radiocarpal joint – wrist joint, located between the forearm and hand regions of the upper limb; articulation formed proximally by the distal end of the radius and the fibrocartilaginous pad that unites the distal radius and ulna bone, and distally by the scaphoid, lunate, and triquetrum carpal bones.
radius – bone located on the lateral side of the forearm.
ramus of the mandible – vertical portion of the mandible.
reabsorption – in the cardiovascular system, the movement of material from the interstitial fluid into the capillaries.
reactive oxygen species (ROS) – a group of extremely reactive peroxides and oxygen-containing radicals that may contribute to cellular damage.
receptor – protein molecule that contains a binding site for another specific molecule (called a ligand).
receptor cell – cell that transduces environmental stimuli into neural signals.
receptor-mediated endocytosis – endocytosis of ligands attached to membrane-bound receptors.
receptor potential – graded potential in a specialised sensory cell that directly causes the release of neurotransmitter without an intervening action potential.
recruitment – increase in the number of motor units involved in contraction.
rectal valve – one of three transverse folds in the rectum where faeces is separated from flatus.
rectum – part of the large intestine between the sigmoid colon and anal canal.
red blood cells (RBCs) – (also, erythrocytes) one of the formed elements of blood that transports oxygen.
red marrow – connective tissue in the interior cavity of a bone where haematopoiesis takes place.
red nucleus – midbrain nucleus that sends corrective commands to the spinal cord along the rubrospinal tract, based on disparity between an original command and the sensory feedback from movement.
refractory period – time after the initiation of an action potential when another action potential cannot be generated.
regional anatomy – study of the structures that contribute to specific body regions.
regulatory T cells (Treg) – (also, suppressor T cells) class of CD4 T cells that regulates other T cell responses.
relative refractory period – time during the refractory period when a new action potential can only be initiated by a stronger stimulus than the current action potential because voltage-gated K+ channels are not closed.
relaxation phase – period after twitch contraction when tension decreases.
remodelling – process by which osteoclasts resorb old or damaged bone at the same time as and on the same surface where osteoblasts form new bone to replace that which is resorbed.
renal artery – branch of the abdominal aorta; supplies each kidney.
renal columns – extensions of the renal cortex into the renal medulla; separates the renal pyramids; contains blood vessels and connective tissues.
renal corpuscle – consists of the glomerulus and Bowman’s capsule.
renal cortex – outer part of kidney containing all the nephrons; some nephrons have loops of Henle extending into the medulla.
renal fat pad – adipose tissue between the renal fascia and the renal capsule that provides protective cushioning to the kidney.
renal hilum – recessed medial area of the kidney through which the renal artery, renal vein, ureters, lymphatics, and nerves pass.
renal papillae – medullary area of the renal pyramids where collecting ducts empty urine into the minor calyces.
renal pyramids – six to eight cone-shaped tissues in the medulla of the kidney containing collecting ducts and the loops of Henle of juxtamedullary nephrons.
renal vein – largest vein entering the inferior vena cava; drains the kidneys and leads to the inferior vena cava.
renin – enzyme produced by juxtaglomerular cells in response to decreased blood pressure or sympathetic nervous activity; catalyses the conversion of angiotensinogen into angiotensin I.
repolarisation – return of the membrane potential to its normally negative voltage at the end of the action potential.
reposition – movement of the thumb from opposition back to the anatomical position (next to index finger).
reserve zone – region of the epiphyseal plate that anchors the plate to the osseous tissue of the epiphysis.
residual volume (RV) – amount of air that remains in the lungs after maximum exhalation.
resistance – any condition or parameter that slows or counteracts the flow of blood.
respiratory acidosis – condition wherein an excess of carbonic acid or CO2 causes the blood to be overly acidic.
respiratory alkalosis – condition wherein a deficiency of carbonic acid/CO2 levels causes the blood to be overly alkaline.
respiratory bronchiole – specific type of bronchiole that leads to alveolar sacs.
respiratory cycle – one sequence of inspiration and expiration.
respiratory epithelium – ciliated lining of much of the conducting zone that is specialised to remove debris and pathogens, and produce mucus.
respiratory membrane – alveolar and capillary wall together, which form an air-blood barrier that facilitates the simple diffusion of gases.
respiratory pump – increase in the volume of the thorax during inhalation that decreases air pressure, enabling venous blood to flow into the thoracic region, then exhalation increases pressure, moving blood into the atria.
respiratory rate – total number of breaths taken each minute.
respiratory volume – varying amounts of air within the lung at a given time.
respiratory zone – includes structures of the respiratory system that are directly involved in gas exchange.
response – nervous system function that causes a target tissue (muscle or gland) to produce an event as a consequence to stimuli.
resting membrane potential – the difference in voltage measured across a cell membrane under steady-state conditions, typically -70 mV.
reticular fibres – fine fibrous protein, made of collagen subunits, which cross-link to form supporting “nets” within connective tissue.
reticular formation – diffuse region of grey matter throughout the brain stem that regulates sleep, wakefulness, and states of consciousness.
reticular lamina – matrix containing collagen and elastin secreted by connective tissue; a component of the basement membrane.
reticular layer – deeper layer of the dermis; it has a reticulated appearance due to the presence of abundant collagen and elastin fibres.
reticular tissue – type of loose connective tissue that provides a supportive framework to soft organs, such as lymphatic tissue, spleen, and the liver.
reticulocyte – immature erythrocyte that may still contain fragments of ribosomal RNA.
reticuloendothelial cell – (also, Kupffer cell) phagocyte in hepatic sinusoids that filters out material from venous blood from the gastrointestinal tract.
reticulospinal tract – extrapyramidal connections between the brain stem and spinal cord that modulate movement, contribute to posture, and regulate muscle tone.
retina – nervous tissue of the eye at which phototransduction takes place.
retinal – cofactor in an opsin molecule that undergoes a biochemical change when struck by a photon (pronounced with a stress on the last syllable).
retinal ganglion cell (RGC) – neuron of the retina that projects along the second cranial nerve.
retraction – posterior motion of the scapula or mandible.
retroperitoneal– outside the peritoneal cavity; in the case of the kidney and ureters, between the parietal peritoneum and the abdominal wall.
Rh blood group – blood-type classification based on the presence or absence of the antigen Rh(D) on the erythrocyte membrane surface.
rhodopsin – photopigment molecule found in the rod photoreceptors.
rhombencephalon – primary vesicle of the embryonic brain that develops into the hindbrain, which includes the pons, cerebellum, and medulla.
ribonuclease – pancreatic enzyme that digests RNA.
ribonucleic acid (RNA) – single-stranded, often internally base paired, molecule that is involved in protein synthesis.
ribosome – cellular organelle that functions in protein synthesis.
ribosomal RNA (rRNA) – RNA that makes up the subunits of a ribosome.
ribs – thin, curved bones of the chest wall.
rickets – disease in children caused by vitamin D deficiency, which leads to the weakening of bones.
right atrioventricular valve – (also, tricuspid valve) valve located between the right atrium and ventricle; consists of three flaps of tissue.
right colic flexure – (also, hepatic flexure) point, at the inferior surface of the liver, where the ascending colon turns abruptly to the left.
right gastric artery – branch of the common hepatic artery; supplies blood to the stomach.
right lymphatic duct – drains lymph fluid from the upper right side of body into the right subclavian vein.
RNA polymerase – enzyme that unwinds DNA and then adds new nucleotides to a growing strand of RNA for the transcription phase of protein synthesis.
rod photoreceptor – one of the two types of retinal receptor cell that is specialised for low-light vision.
root – region of the external nose between the eyebrows.
root – portion of a tooth embedded in the alveolar processes beneath the gum line.
rotation – movement of a bone around a central axis (atlantoaxial joint) or around its long axis (proximal radioulnar joint; shoulder or hip joint); twisting of the vertebral column resulting from the summation of small motions between adjacent vertebrae.
rotator cuff – strong connective tissue structure formed by the fusion of four rotator cuff muscle tendons to the articular capsule of the shoulder joint; surrounds and supports superior, anterior, lateral, and posterior sides of the humeral head.
round window – membrane that marks the end of the scala tympani.
rubrospinal tract – descending motor control pathway, originating in the red nucleus, that mediates control of the limbs based on cerebellar processing.
ruga – fold of gastrointestinal tract mucosa and submucosa in the empty stomach and other organs.
rugae – (of the vagina) folds of skin in the vagina that allow it to stretch during intercourse and childbirth.
S
saccharolytic fermentation – anaerobic decomposition of carbohydrates.
saccule – structure of the inner ear responsible for transducing linear acceleration in the vertical plane.
sacral canal – bony tunnel that runs through the sacrum.
sacral foramina – series of paired openings for nerve exit located on both the anterior (ventral) and posterior (dorsal) aspects of the sacrum.
sacral hiatus – inferior opening and termination of the sacral canal.
sacral micturition centre – group of neurons in the sacral region of the spinal cord that controls urination; acts reflexively unless its action is modified by higher brain centres to allow voluntary urination.
sacral plexus – nerve plexus associated with the lower lumbar and sacral spinal nerves.
sacral promontory – anterior lip of the base (superior end) of the sacrum.
sacrococcygeal curve – anteriorly concave curvature formed by the sacrum and coccyx; a primary curve of the vertebral column.
sacroiliac joint – joint formed by the articulation between the auricular surfaces of the sacrum and ilium.
sacrospinous ligament – ligament that spans the sacrum to the ischial spine of the hip bone.
sacrotuberous ligament – ligament that spans the sacrum to the ischial tuberosity of the hip bone.
sacrum – single bone located near the inferior end of the adult vertebral column that is formed by the fusion of five sacral vertebrae; forms the posterior portion of the pelvis.
saddle joint – synovial joint in which the articulating ends of both bones are convex and concave in shape, such as at the first carpometacarpal joint at the base of the thumb; functionally classified as a biaxial joint.
sagittal plane – two-dimensional, vertical plane that divides the body or organ into right and left sides.
sagittal suture – joint that unites the right and left parietal bones at the midline along the top of the skull.
saliva – aqueous solution of proteins and ions secreted into the mouth by the salivary glands.
salivary amylase – digestive enzyme in saliva that acts on starch.
salivary gland – an exocrine gland that secretes a digestive fluid called saliva.
salivation – secretion of saliva.
saltatory conduction – quick propagation of the action potential along a myelinated axon owing to voltage-gated Na+ channels being present only at the nodes of Ranvier.
saphenous nerve – systemic nerve of the lower anterior leg that is a branch from the femoral nerve.
sarcomere – longitudinally, repeating functional unit of skeletal muscle, with all the contractile and associated proteins involved in contraction.
sarcolemma – plasma membrane of a skeletal muscle fibres.
sarcopenia – age-related muscle atrophy.
sarcoplasm – cytoplasm of a muscle cell.
sarcoplasmic reticulum (SR) – specialised smooth endoplasmic reticulum, which stores, releases, and retrieves Ca2+.
satellite cell – stem cell that helps to repair muscle cells.
saturated fatty acid – long-chain hydrocarbon with single covalent bonds in the carbon chain; the number of hydrogen atoms attached to the carbon skeleton is maximised.
scala tympani – portion of the cochlea that extends from the apex to the round window.
scala vestibuli – portion of the cochlea that extends from the oval window to the apex.
scaphoid – from the lateral side, the first of the four proximal carpal bones; articulates with the radius proximally, the trapezoid, trapezium, and capitate distally, and the lunate medially.
scapula – shoulder blade bone located on the posterior side of the shoulder.
scar – collagen-rich skin formed after the process of wound healing that is different from normal skin.
Schwann cell – neuroglial cell that produces myelin in the peripheral nervous system.
sciatic nerve – systemic nerve from the sacral plexus that is a combination of the tibial and fibular nerves and extends across the hip joint and gluteal region into the upper posterior leg.
sciatica – painful condition resulting from inflammation or compression of the sciatic nerve or any of the spinal nerves that contribute to it.
sclera – white of the eye.
sclerotome – medial portion of a somite consisting of mesenchyme tissue that will give rise to bone, cartilage and fibrous connective tissues.
scoliosis – abnormal lateral curvature of the vertebral column.
scrotum – external pouch of skin and muscle that houses the testes.
sebaceous gland – type of oil gland found in the dermis all over the body and helps to lubricate and waterproof the skin and hair by secreting sebum.
sebum – oily substance that is composed of a mixture of lipids that lubricates the skin and hair.
second messenger – molecule that initiates a signalling cascade in response to hormone binding on a cell membrane receptor and activation of a G protein.
secondary adaptive response – immune response observed upon re-exposure to a pathogen, which is stronger and faster than a primary response.
secondary curve – posteriorly concave curvatures of the cervical and lumbar regions of the vertebral column that develop after the time of birth.
secondary follicles – ovarian follicles with a primary oocyte and multiple layers of granulosa cells.
secondary lymphoid organs – sites where lymphocytes mount adaptive immune responses; examples include lymph nodes and spleen.
secondary ossification centre – region of bone development in the epiphyses.
secondary sex characteristics – physical characteristics that are influenced by sex steroid hormones and have supporting roles in reproductive function.
secondary structure – regular structure that proteins form by intramolecular hydrogen bonding between the oxygen atom of one amino acid residue and the hydrogen attached to the nitrogen atom of another amino acid residue.
secondary union – wound healing facilitated by wound contraction.
secondary vesicle – five vesicles that develop from primary vesicles, continuing the process of differentiation of the embryonic brain.
secretory phase – phase of the menstrual cycle in which the endometrium secretes a nutrient-rich fluid in preparation for implantation of an embryo.
section – in anatomy, a single flat surface of a three-dimensional structure that has been cut through.
segmentation – alternating contractions and relaxations of non-adjacent segments of the intestine that move food forward and backward, breaking it apart and mixing it with digestive secretions.
selective permeability – feature of any barrier that allows certain substances to cross but excludes others.
sella turcica – elevated area of sphenoid bone located at midline of the middle cranial fossa.
semen – ejaculatory fluid composed of sperm and secretions from the seminal vesicles, prostate, and bulbourethral glands.
semicircular canals – structures within the inner ear responsible for transducing rotational movement information.
semilunar valves – valves located at the base of the pulmonary trunk and at the base of the aorta.
seminal vesicle – gland that produces seminal fluid, which contributes to semen.
seminiferous tubules – tube structures within the testes where spermatogenesis occurs.
sensation – nervous system function that receives information from the environment and translates it into the electrical signals of nervous tissue.
sensitisation – first exposure to an antigen.
sensor – (also, receptor) reports a monitored physiological value to the control centre.
sensory homunculus – topographic representation of the body within the somatosensory cortex demonstrating the correspondence between neurons processing stimuli and sensitivity.
sensory modality – a particular system for interpreting and perceiving environmental stimuli by the nervous system.
sepsis – (also, septicaemia) organismal-level inflammatory response to a massive infection.
septal cartilage – flat cartilage structure that forms the anterior portion of the nasal septum.
septic shock – (also, blood poisoning) type of shock that follows a massive infection resulting in organism-wide inflammation.
septum – (plural = septa) walls or partitions that divide the heart into chambers.
septum primum – flap of tissue in the foetus that covers the foramen ovale within a few seconds after birth.
seroconversion – clearance of pathogen in the serum and the simultaneous rise of serum antibody.
serous gland – group of cells within the serous membrane that secrete a lubricating substance onto the surface.
serous membrane – membrane that covers organs and reduces friction; also referred to as serosa.
serosa – membrane that covers organs and reduces friction; also referred to as serous membrane.
Sertoli cells – cells that support germ cells through the process of spermatogenesis; a type of sustentacular cell.
serum – blood plasma that does not contain clotting factors.
sesamoid bone – small, round bone embedded in a tendon; protects the tendon from compressive forces.
set point – ideal value for a physiological parameter; the level or small range within which a physiological parameter such as blood pressure is stable and optimally healthful, that is, within its parameters of homeostasis.
severe combined immunodeficiency disease (SCID) – genetic mutation that affects both T cell and B cell arms of the immune response.
shaft of the femur – cylindrically shaped region that forms the central portion of the femur.
shaft of the fibula – elongated, slender portion located between the expanded ends of the fibula.
shaft of the humerus – narrow, elongated, central region of the humerus.
shaft of the radius – narrow, elongated, central region of the radius.
shaft of the tibia – triangular-shaped, central portion of the tibia.
shaft of the ulna – narrow, elongated, central region of the ulna.
short bone – cube-shaped bone that is approximately equal in length, width, and thickness; provides limited motion.
shunt – circulatory shortcut that diverts the flow of blood from one region to another.
sickle cell disease – (also, sickle cell anaemia) inherited blood disorder in which haemoglobin molecules are malformed, leading to the breakdown of RBCs that take on a characteristic sickle shape.
sigmoid colon – end portion of the colon, which terminates at the rectum.
sigmoid sinuses – enlarged veins that receive blood from the transverse sinuses; flow through the jugular foramen and into the internal jugular vein.
simple columnar epithelium – tissue that consists of a single layer of column-like cells; promotes secretion and absorption in tissues and organs.
simple cuboidal epithelium – tissue that consists of a single layer of cube-shaped cells; promotes secretion and absorption in ducts and tubules.
simple squamous epithelium – tissue that consists of a single layer of flat scale-like cells; promotes diffusion and filtration across surface.
sinoatrial (SA) node – known as the pacemaker, a specialised clump of myocardial conducting cells located in the superior portion of the right atrium that has the highest inherent rate of depolarisation that then spreads throughout the heart.
sinus rhythm – normal contractile pattern of the heart.
sinus venosus – develops into the posterior portion of the right atrium, the SA node, and the coronary sinus.
sinusoid capillary – rarest type of capillary, which has extremely large intercellular gaps in the basement membrane in addition to clefts and fenestrations; found in areas such as the bone marrow and liver where passage of large molecules occurs.
sister chromatid – one of a pair of identical chromosomes, formed during DNA replication.
size exclusion – principle of selectively allowing ions through a channel on the basis of their relative size.
skeleton – bones of the body.
skeletal muscle – usually attached to bone, under voluntary control, each cell is a fibre that is multinucleated and striated.
skeletal muscle pump – effect on increasing blood pressure within veins by compression of the vessel caused by the contraction of nearby skeletal muscle.
skeletal system – organ system composed of bones and cartilage that provides for movement, support, and protection.
skull – bony structure that forms the head, face, and jaws, and protects the brain; consists of 22 bones.
small cardiac vein – parallels the right coronary artery and drains blood from the posterior surfaces of the right atrium and ventricle; drains into the great cardiac vein.
small intestine – section of the gastrointestinal tract where most digestion and absorption occurs.
small saphenous vein – located on the lateral surface of the leg; drains blood from the superficial regions of the lower leg and foot, and leads to the popliteal vein.
smooth muscle – under involuntary control, moves internal organs, cells contain a single nucleus, are spindle-shaped, and do not appear striated; each cell is a fibre.
sodium-potassium pump – (also, Na+/K+ ATP-ase) membrane-embedded protein pump that uses ATP to move Na+ out of a cell and K+ into the cell.
soft palate – posterior region of the bottom portion of the nasal cavity that consists of skeletal muscle.
soleal line – small, diagonally running ridge located on the posterior side of the proximal tibia.
solitary nucleus – medullar nucleus that receives taste information from the facial and glossopharyngeal nerves.
soma – in neurons, that portion of the cell that contains the nucleus; the cell body, as opposed to the cell processes (axons and dendrites).
somatic cell – all cells of the body excluding gamete cells.
somatic nervous system (SNS) – functional division of the nervous system that is concerned with conscious perception, voluntary movement, and skeletal muscle reflexes.
somatosensation – general senses related to the body, usually thought of as the senses of touch, which would include pain, temperature, and proprioception.
somites – blocks of paraxial mesoderm cells.
spatial summation – combination of graded potentials across the neuronal cell membrane caused by signals from separate presynaptic elements that add up to initiate an action potential.
special sense – any sensory system associated with a specific organ structure, namely smell, taste, sight, hearing, and balance.
specific gravity – weight of a liquid compared to pure water, which has a specific gravity of 1.0; any solute added to water will increase its specific gravity.
sperm – (also, spermatozoon) male gamete.
spermatic cord – bundle of nerves and blood vessels that supplies the testes; contains ductus deferens.
spermatid – immature sperm cells produced by meiosis II of secondary spermatocytes.
spermatocyte – cell that results from the division of spermatogonium and undergoes meiosis I and meiosis II to form spermatids.
spermatogenesis – formation of new sperm, occurs in the seminiferous tubules of the testes.
spermatogonia – (singular = spermatogonium) diploid precursor cells that become sperm.
spermiogenesis – transformation of spermatids to spermatozoa during spermatogenesis.
S phase – stage of the cell cycle during which DNA replication occurs.
sphenoid bone – unpaired bone that forms the central base of skull.
sphenoid sinus – air-filled space located within the sphenoid bone; most posterior of the paranasal sinuses.
sphygmomanometer – blood pressure cuff attached to a device that measures blood pressure.
spinal accessory nerve – eleventh cranial nerve; responsible for contraction of neck muscles.
spinal cavity – division of the dorsal cavity that houses the spinal cord; also referred to as vertebral cavity.
spinal cord – organ of the central nervous system found within the vertebral cavity and connected with the periphery through spinal nerves; mediates reflex behaviours.
spinal nerve -one of 31 nerves connected to the spinal cord.
spinal trigeminal nucleus – component of the trigeminal nuclei that is found in the medulla.
spine of the scapula – prominent ridge passing mediolaterally across the upper portion of the posterior scapular surface.
spinothalamic tract – ascending tract of the spinal cord associated with pain and temperature sensations.
spinous process – unpaired bony process that extends posteriorly from the vertebral arch of a vertebra.
spiral ganglion – location of neuronal cell bodies that transmit auditory information along the eighth cranial nerve.
spleen – secondary lymphoid organ that filters pathogens from the blood (white pulp) and removes degenerating or damaged blood cells (red pulp).
splenic artery – branch of the celiac trunk; supplies blood to the spleen.
spliceosome – complex of enzymes that serves to splice out the introns of a pre-mRNA transcript.
splicing – the process of modifying a pre-mRNA transcript by removing certain, typically non-coding, regions.
spongy bone – (also, cancellous bone) trabeculated osseous tissue that supports shifts in weight distribution.
spontaneous depolarisation – (also, prepotential depolarisation) the mechanism that accounts for the autorhythmic property of cardiac muscle; the membrane potential increases as sodium ions diffuse through the always-open sodium ion channels and causes the electrical potential to rise.
squamous cell carcinoma – type of skin cancer that originates from the stratum spinosum of the epidermis.
squamous suture – joint that unites the parietal bone to the squamous portion of the temporal bone on the lateral side of the skull.
stage of exhaustion – stage three of the general adaptation syndrome; the body’s long-term response to stress mediated by the hormones of the adrenal cortex.
stage of resistance – stage two of the general adaptation syndrome; the body’s continued response to stress after stage one diminishes.
stapes – (also, stirrup) ossicle of the middle ear that is attached to the inner ear.
starch – storage carbohydrate in plants.
stem cell – cell that is oligo-, multi-, or pluripotency that has the ability to produce additional stem cells rather than becoming further specialised.
stereocilia – array of apical membrane extensions in a hair cell that transduce movements when they are bent.
sternal angle – junction line between manubrium and body of the sternum and the site for attachment of the second rib to the sternum.
sternal end of the clavicle – medial end of the clavicle that articulates with the manubrium of the sternum.
sternoclavicular joint – articulation between the manubrium of the sternum and the sternal end of the clavicle; forms the only bony attachment between the pectoral girdle of the upper limb and the axial skeleton.
sternum – flattened bone located at the centre of the anterior chest.
steroid – type of lipid comprised of four fused hydrocarbon rings forming a planar structure.
stimulus – an event in the external or internal environment that registers as activity in a sensory neuron.
stomach – gastrointestinal tract organ that contributes to chemical and mechanical digestion of food from the oesophagus before releasing it, as chyme, to the small intestine.
straight sinus – enlarged vein that drains blood from the brain; receives most of the blood from the great cerebral vein and flows into the left or right transverse sinus.
stratified columnar epithelium – tissue that consists of two or more layers of column-like cells, contains glands and is found in some ducts.
stratified cuboidal epithelium – tissue that consists of two or more layers of cube-shaped cells, found in some ducts.
stratified squamous epithelium – tissue that consists of multiple layers of cells with the most apical being flat scale-like cells; protects surfaces from abrasion.
stratum basale – deepest layer of the epidermis, made of epidermal stem cells.
stratum corneum – most superficial layer of the epidermis.
stratum granulosum – layer of the epidermis superficial to the stratum spinosum.
stratum lucidum – layer of the epidermis between the stratum granulosum and stratum corneum, found only in thick skin covering the palms, soles of the feet, and digits.
stratum spinosum – layer of the epidermis superficial to the stratum basale, characterised by the presence of desmosomes.
stress-relaxation response – relaxation of smooth muscle tissue after being stretched.
stretch mark – mark formed on the skin due to a sudden growth spurt and expansion of the dermis beyond its elastic limits.
stretch reflex – response to activation of the muscle spindle stretch receptor that causes contraction of the muscle to maintain a constant length.
striation – alignment of parallel actin and myosin filaments which form a banded pattern.
striatum – the caudate and putamen collectively, as part of the basal nuclei, which receive input from the cerebral cortex.
stroke volume (SV) – amount of blood pumped by each ventricle per contraction; also, the difference between EDV and ESV.
styloid process – downward projecting, elongated bony process located on the inferior aspect of the skull.
styloid process of the radius – pointed projection located on the lateral end of the distal radius.
styloid process of the ulna – short, bony projection located on the medial end of the distal ulna.
stylomastoid foramen – opening located on inferior skull, between the styloid process and mastoid process.
subacromial bursa – bursa that protects the supraspinatus muscle tendon and superior end of the humerus from rubbing against the acromion of the scapula.
subarachnoid space – space between the arachnoid mater and pia mater that contains CSF and the fibrous connections of the arachnoid trabeculae.
subclavian artery – right subclavian arises from the brachiocephalic artery, whereas the left subclavian artery arises from the aortic arch; gives rise to the internal thoracic, vertebral, and thyrocervical arteries; supplies blood to the arms, chest, shoulders, back, and central nervous system.
subclavian vein – located deep in the thoracic cavity; becomes the axillary vein as it enters the axillary region; drains the axillary and smaller local veins near the scapular region; leads to the brachiocephalic vein.
subcortical nucleus – all the nuclei beneath the cerebral cortex, including the basal nuclei and the basal forebrain.
subcutaneous bursa – bursa that prevents friction between skin and an underlying bone.
sublingual gland – one of a pair of major salivary glands located beneath the tongue.
submandibular gland – one of a pair of major salivary glands located in the floor of the mouth.
submodality – specific sense within a broader major sense such as sweet as a part of the sense of taste, or colour as a part of vision.
submucosa – layer of dense connective tissue in the gastrointestinal tract wall that binds the overlying mucosa to the underlying muscularis.
submuscular bursa – bursa that prevents friction between bone and a muscle or between adjacent muscles.
submucosal plexus – (plexus of Meissner) nerve supply that regulates activity of glands and smooth muscle.
subpubic angle – inverted V-shape formed by the convergence of the right and left ischiopubic rami; this angle is greater than 80 degrees in females and less than 70 degrees in males.
subscapular bursa – bursa that prevents rubbing of the subscapularis muscle tendon against the scapula.
subscapular fossa – broad depression located on the anterior (deep) surface of the scapula.
subscapular vein – drains blood from the subscapular region and leads to the axillary vein.
subtalar joint – articulation between the talus and calcaneus bones of the foot; allows motions that contribute to inversion/eversion of the foot.
substantia nigra pars compacta – nuclei within the basal nuclei that release dopamine to modulate the function of the striatum; part of the motor pathway.
substantia nigra pars reticulata – nuclei within the basal nuclei that serve as an output centre of the nuclei; part of the motor pathway.
subtendinous bursa – bursa that prevents friction between bone and a muscle tendon.
subthalamus – nucleus within the basal nuclei that is part of the indirect pathway.
sucrase – brush border enzyme that breaks down sucrose into glucose and fructose.
sudoriferous gland – sweat gland.
sulcus – (plural = sulci) fat-filled groove visible on the surface of the heart; coronary vessels are also located in these areas.
summate – to add together, as in the cumulative change in postsynaptic potentials toward reaching threshold in the membrane, either across a span of the membrane or over a certain amount of time.
superficial – describes a position nearer to the surface of the body.
superficial burns – superficial burn that injures only the epidermis.
superior – describes a position above or higher than another part of the body proper; also referred to as cranial.
superior angle of the scapula – corner of the scapula between the superior and medial borders of the scapula.
superior articular process – bony process that extends upward from the vertebral arch of a vertebra that articulates with the inferior articular process of the next higher vertebra.
superior articular process of the sacrum – paired processes that extend upward from the sacrum to articulate (join) with the inferior articular processes from the L5 vertebra.
superior border of the scapula – superior margin of the scapula.
superior colliculus – half of the midbrain tectum that is responsible for aligning visual, auditory, and somatosensory spatial perceptions.
superior oblique – extraocular muscle responsible for medial rotation of the eye.
superior pubic ramus – narrow segment of bone that passes laterally from the pubic body to join the ilium.
superior rectus – extraocular muscle responsible for looking up.
superior rotation – movement of the scapula during upper limb abduction in which the glenoid cavity of the scapula moves in an upward direction as the medial end of the scapular spine moves in a downward direction.
superior sagittal sinus – dural sinus that runs along the top of the longitudinal fissure and drains blood from the majority of the outer cerebrum.
supplemental motor area – cortical area anterior to the primary motor cortex that is responsible for planning movements.
suprachiasmatic nucleus – hypothalamic target of the retina that helps to establish the circadian rhythm of the body based on the presence or absence of daylight.
supraglenoid tubercle – small bump located at the superior margin of the glenoid cavity.
suprascapular notch – small notch located along the superior border of the scapula, medial to the coracoid process.
supraspinous fossa – narrow depression located on the posterior scapula, superior to the spine.
supraspinous ligament – ligament that interconnects the spinous processes of the thoracic and lumbar vertebrae.
superior mesenteric artery – branch of the abdominal aorta; supplies blood to the small intestine (duodenum, jejunum, and ileum), the pancreas, and a majority of the large intestine.
superior nasal concha – smallest and most superiorly located of the nasal conchae; formed by the ethmoid bone.
superior nuchal line – paired bony lines on the posterior skull that extend laterally from the external occipital protuberance.
superior orbital fissure – irregularly shaped opening between the middle cranial fossa and the posterior orbit.
superior phrenic artery – branch of the thoracic aorta; supplies blood to the superior surface of the diaphragm.
superior sagittal sinus – enlarged vein located midsagittally between the meningeal and periosteal layers of the dura mater within the falx cerebri; receives most of the blood drained from the superior surface of the cerebrum and leads to the inferior jugular vein and the vertebral vein.
superior vena cava – large systemic vein that returns blood to the heart from the superior portion of the body.
supine – face up.
supinated position – forearm position in which the palm faces anteriorly (anatomical position).
supination – forearm motion that moves the palm of the hand from the palm backward to the palm forward position.
supportive connective tissue – type of connective tissue that provides strength to the body and protects soft tissue.
supraorbital foramen – opening located on anterior skull, at the superior margin of the orbit.
supraorbital margin – superior margin of the orbit.
surgical neck – region of the humerus where the expanded, proximal end joins with the narrower shaft.
suspensory ligaments – bands of connective tissue that suspend the breast onto the chest wall by attachment to the overlying dermis.
sustentaculum tali – bony ledge extending from the medial side of the calcaneus bone.
suture – junction line at which adjacent bones of the skull are united by fibrous connective tissue.
sympathetic chain ganglia – autonomic ganglia in a chain along the anterolateral aspect of the vertebral column that are responsible for contributing to homeostatic mechanisms of the autonomic nervous system.
symphysis – type of cartilaginous joint where the bones are joined by fibrocartilage.
synapse – narrow junction across which a chemical signal passes from neuron to the next, initiating a new electrical signal in the target cell.
synaptic cleft – space between a nerve (axon) terminal and a motor end-plate.
synaptic end bulb – swelling at the end of an axon where neurotransmitter molecules are released onto a target cell across a synapse.
synarthrosis – immobile or nearly immobile joint.
synchondrosis – type of cartilaginous joint where the bones are joined by hyaline cartilage.
syncytiotrophoblast – superficial cells of the trophoblast that fuse to form a multinucleated body that digests endometrial cells to firmly secure the blastocyst to the uterine wall.
syndesmosis – type of fibrous joint in which two separated, parallel bones are connected by an interosseous membrane.
synaptic cleft – small gap between cells in a chemical synapse where neurotransmitter diffuses from the presynaptic element to the postsynaptic element.
synostosis – site at which adjacent bones or bony components have fused together.
synovial fluid – thick, lubricating fluid that fills the interior of a synovial joint.
synovial joint – joint at which the articulating surfaces of the bones are located within a joint cavity formed by an articular capsule.
synovial membrane – connective tissue membrane that lines the cavities of freely movable joints, producing synovial fluid for lubrication.
systemic anatomy – study of the structures that contribute to specific body systems.
systemic circuit – blood flow to and from virtually all of the tissues of the body.
systemic nerve – nerve in the periphery distal to a nerve plexus or spinal nerve.
systemic oedema – increased fluid retention in the interstitial spaces and cells of the body; can be seen as swelling over large areas of the body, particularly the lower extremities.
systole – period of time when the heart muscle is contracting.
systolic pressure – larger number recorded when measuring arterial blood pressure; represents the maximum value following ventricular contraction.
T
talocrural joint – ankle joint; articulation between the talus bone of the foot and medial malleolus of the tibia, distal tibia, and lateral malleolus of the fibula; a uniaxial hinge joint that allows only for dorsiflexion and plantar flexion of the foot.
talus – tarsal bone that articulates superiorly with the tibia and fibula at the ankle joint; also articulates inferiorly with the calcaneus bone and anteriorly with the navicular bone.
target heart rate – range in which both the heart and lungs receive the maximum benefit from an aerobic workout.
tarsal bone – one of the seven bones that make up the posterior foot; includes the calcaneus, talus, navicular, cuboid, medial cuneiform, intermediate cuneiform, and lateral cuneiform bones.
taste buds – structures within a papilla on the tongue that contain gustatory receptor cells.
T cell – lymphocyte that acts by secreting molecules that regulate the immune system or by causing the destruction of foreign cells, viruses, and cancer cells.
T cell-dependent antigen – antigen that binds to B cells, which requires signals from T cells to make antibody.
T cell-independent antigen – binds to B cells, which do not require signals from T cells to make antibody.
T cell tolerance – process during T cell differentiation where most T cells that recognise antigens from one’s own body are destroyed.
tectorial membrane – component of the organ of Corti that lays over the hair cells, into which the stereocilia are embedded.
tectospinal tract – extrapyramidal connections between the superior colliculus and spinal cord.
tectum – region of the midbrain, thought of as the roof of the cerebral aqueduct, which is subdivided into the inferior and superior colliculi.
tegmentum – region of the midbrain, thought of as the floor of the cerebral aqueduct, which continues into the pons and medulla as the floor of the fourth ventricle.
telencephalon – secondary vesicle of the embryonic brain that develops into the cerebrum.
telogen – resting phase of the hair growth cycle initiated with catagen and terminated by the beginning of a new anagen phase of hair growth.
telophase – final stage of mitosis (and meiosis), preceding cytokinesis, characterised by the formation of two new daughter nuclei.
temporal bone – paired bones that form the lateral, inferior portions of the skull, with squamous, mastoid, and petrous portions.
temporal fossa – shallow space on the lateral side of the skull, above the level of the zygomatic arch.
temporal lobe – region of the cerebral cortex directly beneath the temporal bone of the cranium.
temporal process of the zygomatic bone – short extension from the zygomatic bone that forms the anterior portion of the zygomatic arch.
temporal summation – combination of graded potentials at the same location on a neuron resulting in a strong signal from one input.
temporal vein – drains blood from the temporal region and leads to the external jugular vein.
temporomandibular joint (TMJ) – articulation between the condyle of the mandible and the mandibular fossa and articular tubercle of the temporal bone of the skull; allows for depression/elevation (opening/closing of mouth), protraction/retraction, and side-to-side motions of the mandible.
tendon – dense connective tissue structure that anchors a muscle to bone.
tendon sheath – connective tissue that surrounds a tendon at places where the tendon crosses a joint; contains a lubricating fluid to prevent friction and allow smooth movements of the tendon.
teniae coli – one of three smooth muscle bands that make up the longitudinal muscle layer of the muscularis in all the large intestine except the terminal end.
terminal ganglion – autonomic ganglia that are near or within the walls of organs that are responsible for contributing to homeostatic mechanisms of the autonomic nervous system.
tertiary follicles – (also, antral follicles) ovarian follicles with a primary or secondary oocyte, multiple layers of granulosa cells, and a fully formed antrum.
tertiary structure – a protein’s three-dimensional conformation, including interactions between secondary structural elements; formed from interactions between amino acid side chains.
testes – (singular = testis) male gonads.
testicular artery – branch of the abdominal aorta; will ultimately travel outside the body cavity to the testes and form one component of the spermatic cord.
testicular vein – drains the testes and forms part of the spermatic cord; the right testicular vein empties directly into the inferior vena cava and the left testicular vein empties into the left renal vein.
testosterone – steroid hormone secreted by the male testes and important in the maturation of sperm cells, growth and development of the male reproductive system, and the development of male secondary sex characteristics.
tetanus – a continuous fused contraction.
thalamus – region of the central nervous system that acts as a relay for sensory pathways.
thalassaemia – inherited blood disorder in which the life-span of RBCs is significantly reduced due to unbalanced production of globin chains.
theca cells – androgen-producing cells in a maturing ovarian follicle.
thermoreceptor – type of sensory receptor capable of transducing temperature stimuli into neural action potentials.
thick filament – the thick myosin strands and their multiple heads projecting from the centre of the sarcomere toward, but not all to way to, the Z-discs.
thigh – portion of the lower limb located between the hip and knee joints.
thin filament – thin strands of actin and its troponin-tropomyosin complex projecting from the Z-discs toward the centre of the sarcomere.
third ventricle – portion of the ventricular system that is in the region of the diencephalon.
thoracic aorta – portion of the descending aorta superior to the aortic hiatus.
thoracic cage – consists of 12 pairs of ribs and sternum.
thoracic cavity – division of the anterior (ventral) cavity that houses the heart, lungs, oesophagus, and trachea.
thoracic curve – anteriorly concave curvature of the thoracic vertebral column region; a primary curve of the vertebral column.
thoracic duct – large duct that drains lymph from the lower limbs, left thorax, left upper limb, and the left side of the head.
thoracic wall compliance – ability of the thoracic wall to stretch while under pressure.
thoracic vertebrae – twelve vertebrae numbered as T1–T12 that are located in the thoracic region (upper back) of the vertebral column.
thoroughfare channel – continuation of the metarteriole that enables blood to bypass a capillary bed and flow directly into a venule, creating a vascular shunt.
threshold – membrane voltage at which an action potential is initiated.
thrombin – enzyme essential for conversion of soluble plasma fibrinogen to insoluble fibrin.
thrombosis – excessive clot formation.
thrombus – aggregation of fibrin, platelets, and erythrocytes in an intact artery or vein.
thrombocytes – platelets, one of the formed elements of blood that consists of cell fragments broken off from megakaryocytes.
thrombocytopenia – condition in which there are too few platelets, resulting in abnormal bleeding (haemophilia).
thrombocytosis – condition in which there are too many platelets, resulting in abnormal clotting (thrombosis).
thrombopoietin – hormone secreted by the liver and kidneys that prompts the development of
thymocyte – immature T cell found in the thymus.
thymosins – hormones produced and secreted by the thymus that play an important role in the development and differentiation of T cells.
thymus – primary lymphoid organ; where T lymphocytes proliferate and mature megakaryocytes into thrombocytes (platelets).
thyrocervical artery – arises from the subclavian artery; supplies blood to the thyroid, the cervical region, the upper back, and shoulder.
thyroid cartilage – largest piece of cartilage that makes up the larynx and consists of two lamina.
thyroid gland – large endocrine gland responsible for the synthesis of thyroid hormones.
thyroid-stimulating hormone (TSH) – anterior pituitary hormone that triggers secretion of thyroid hormones by the thyroid gland (also called thyrotropin).
thyroxine – (also, tetraiodothyronine, T4) amino acid–derived thyroid hormone that is more abundant but less potent than T3 and often converted to T3 by target cells.
Th1 cells – cells that secrete cytokines that enhance the activity of macrophages and other cells.
Th2 cells – cells that secrete cytokines that induce B cells to differentiate into antibody-secreting plasma cells.
tibia – shin bone; the large, weight-bearing bone located on the medial side of the leg.
tibial collateral ligament – extrinsic ligament of knee joint that spans from the medial epicondyle of the femur to the medial tibia; resists hyperextension and rotation of extended knee.
tibial nerve – systemic nerve of the posterior leg that begins as part of the sciatic nerve.
tibial tuberosity – elevated area on the anterior surface of the proximal tibia.
tidal volume (TV) – amount of air that normally enters the lungs during quiet breathing.
tight junction – forms an impermeable barrier between cells.
tissue – group of similar or closely related cells that act together to perform a specific function.
tissue factor – protein thromboplastin, which initiates the extrinsic pathway when exposed to clotting factor VII as a result of tissue damage.
tissue membrane – thin layer or sheet of cells that covers the outside of the body, organs, and internal cavities.
tissue typing – typing of MHC molecules between a recipient and donor for use in a potential transplantation procedure.
T lymphocytes – (also, T cells) lymphocytes that provide cellular-level immunity by physically attacking foreign or diseased cells.
tongue – accessory digestive organ of the mouth, the bulk of which is composed of skeletal muscle.
tonsils – lymphoid nodules associated with the nasopharynx.
topographical – relating to positional information.
total dead space – sum of the anatomical dead space and alveolar dead space.
total lung capacity (TLC) – total amount of air that can be held in the lungs; sum of TV, ERV, IRV, and RV.
total pressure – sum of all the partial pressures of a gaseous mixture.
totipotent – embryonic cells that have the ability to differentiate into any type of cell and organ in the body.
totipotent stem cell – embryonic stem cell that is capable of differentiating into any and all cells of the body, enabling the full development of an organism.
trabeculae – (singular = trabecula) spikes or sections of the lattice-like matrix in spongy bone.
trabeculae carneae – ridges of muscle covered by endocardium located in the ventricles.
trachea – tube composed of cartilaginous rings and supporting tissue that connects the lung bronchi and the larynx; provides a route for air to enter and exit the lung.
trachealis muscle – smooth muscle located in the fibroelastic membrane of the trachea.
tract – bundle of axons in the central nervous system having the same function and point of origin.
transcription – process of producing an mRNA molecule that is complementary to a particular gene of DNA.
transcription factor – one of the proteins that regulate the transcription of genes.
transduction – process of changing an environmental stimulus into the electrochemical signals of the nervous system.
trans fat – fat formed artificially by hydrogenating oils, leading to a different arrangement of double bond(s) than those in naturally occurring lipids.
transferrin – plasma protein that binds reversibly to iron and distributes it throughout the body.
transient ischemic attack (TIA) – temporary loss of neurological function caused by a brief interruption in blood flow; also known as a mini-stroke.
transitional epithelium – form of stratified epithelium found in the urinary tract, characterised by an apical layer of cells that change shape in response to the presence of urine.
transfer RNA (tRNA) – molecules of RNA that serve to bring amino acids to a growing polypeptide strand and properly place them into the sequence.
translation – process of producing a protein from the nucleotide sequence code of an mRNA transcript.
transpulmonary pressure – pressure difference between the intrapleural and intra-alveolar pressures.
transverse colon – part of the colon between the ascending colon and the descending colon.
transverse foramen – opening found only in the transverse processes of cervical vertebrae.
transverse plane – two-dimensional, horizontal plane that divides the body or organ into superior and inferior portions.
transverse process – paired bony processes that extends laterally from the vertebral arch of a vertebra.
transverse sinuses – pair of enlarged veins near the lambdoid suture that drain the occipital, sagittal, and straight sinuses, and leads to the sigmoid sinuses.
trapezium – from the lateral side, the first of the four distal carpal bones; articulates with the scaphoid proximally, the first and second metacarpals distally, and the trapezoid medially.
trapezoid – from the lateral side, the second of the four distal carpal bones; articulates with the scaphoid proximally, the second metacarpal distally, the trapezium laterally, and the capitate medially.
treppe – stepwise increase in contraction tension.
triacylglycerol (also, triglyceride) – fat molecule; consists of three fatty acids linked to a glycerol molecule.
triad – the grouping of one T-tubule and two terminal cisternae.
tricuspid valve – term used most often in clinical settings for the right atrioventricular valve.
trigeminal ganglion – sensory ganglion that contributes sensory fibres to the trigeminal nerve.
trigeminal nerve – fifth cranial nerve; responsible for cutaneous sensation of the face and contraction of the muscles of mastication.
trigone – area at the base of the bladder marked by the two ureters in the posterior–lateral aspect and the urethral orifice in the anterior aspect oriented like points on a triangle.
triiodothyronine – (also, T3) amino acid–derived thyroid hormone that is less abundant but more potent than T4.
triplet – consecutive sequence of three nucleotides on a DNA molecule that, when transcribed into an mRNA codon, corresponds to a particular amino acid.
triquetrum – from the lateral side, the third of the four proximal carpal bones; articulates with the lunate laterally, the hamate distally, and has a facet for the pisiform.
trochlea – pulley-shaped region located medially at the distal end of the humerus; articulates at the elbow with the trochlear notch of the ulna.
trochlear nerve – fourth cranial nerve; responsible for contraction of one of the extraocular muscles.
trochlear notch – large, C-shaped depression located on the anterior side of the proximal ulna; articulates at the elbow with the trochlea of the humerus.
trophoblast – fluid-filled shell of squamous cells destined to become the chorionic villi, placenta, and associated foetal membranes.
troponin – regulatory protein that binds to actin, tropomyosin, and calcium.
tropomyosin – regulatory protein that covers myosin-binding sites to prevent actin from binding to myosin.
true ribs – vertebrosternal ribs 1–7 that attach via their costal cartilage directly to the sternum.
true vocal cord – one of the pair of folded, white membranes that have a free inner edge that oscillates as air passes through to produce sound.
truncus arteriosus – portion of the primitive heart that will eventually divide and give rise to the ascending aorta and pulmonary trunk.
tubercle of the rib – small bump on the posterior side of a rib for articulation with the transverse process of a thoracic vertebra.
T-tubule – projection of the sarcolemma into the interior of the cell.
trunk – large vessel that gives rise to smaller vessels.
tubuloglomerular feedback – feedback mechanism involving the JGA; macula densa cells monitor Na+ concentration in the terminal portion of the ascending loop of Henle and act to cause vasoconstriction or vasodilation of afferent and efferent arterioles to alter GFR.
tunica externa – (also, tunica adventitia) outermost layer or tunic of a vessel (except capillaries).
tunica intima – (also, tunica interna) innermost lining or tunic of a vessel.
tunica media – middle layer or tunic of a vessel (except capillaries).
T wave – component of the electrocardiogram that represents the repolarisation of the ventricles.
twitch – single contraction produced by one action potential.
tympanic membrane – ear drum.
type I alveolar cell – squamous epithelial cells that are the major cell type in the alveolar wall; highly permeable to gases.
type II alveolar cell – cuboidal epithelial cells that are the minor cell type in the alveolar wall; secrete pulmonary surfactant.
type I hypersensitivity – immediate response mediated by mast cell degranulation caused by the crosslinking of the antigen-specific IgE molecules on the mast cell surface.
type II hypersensitivity – cell damage caused by the binding of antibody and the activation of complement, usually against red blood cells.
type III hypersensitivity – damage to tissues caused by the deposition of antibody-antigen (immune) complexes followed by the activation of complement.
U
ulna – bone located on the medial side of the forearm.
ulnar artery – formed at the bifurcation of the brachial artery; parallels the ulna; gives off smaller branches until it reaches the carpal region where it fuses with the radial artery to form the superficial and deep palmar arches; supplies blood to the lower arm and carpal region.
ulnar collateral ligament – intrinsic ligament on the medial side of the elbow joint; spans from the medial epicondyle of the humerus to the medial ulna.
ulnar nerve – systemic nerve of the arm located close to the ulna, a bone of the forearm.
ulnar notch of the radius – shallow, smooth area located on the medial side of the distal radius; articulates with the head of the ulna at the distal radioulnar joint.
ulnar tuberosity – roughened area located on the anterior, proximal ulna inferior to the coronoid process.
ulnar vein – parallels the ulna and ulnar artery; arises from the palmar venous arches and leads to the brachial vein.
umami – taste submodality for sensitivity to the concentration of amino acids; also called the savory sense.
umbilical arteries – pair of vessels that runs within the umbilical cord and carries foetal blood low in oxygen and high in waste to the placenta for exchange with maternal blood.
umbilical cord – connection between the developing conceptus and the placenta; carries deoxygenated blood and wastes from the foetus and returns nutrients and oxygen from the mother.
umbilical vein – single vessel that originates in the placenta and runs within the umbilical cord, carrying oxygen- and nutrient-rich blood to the foetal heart.
uniaxial joint – type of diarthrosis; joint that allows for motion within only one plane (one axis).
unipolar – shape of a neuron which has only one process that includes both the axon and dendrite.
unipotent – describes the condition of being committed to a single specialised cell type.
universal RBC donor – individual with type O− blood.
universal RBC recipient – individual with type AB+ blood.
unsaturated fatty acid – long-chain hydrocarbon that has one or more double bonds in the hydrocarbon chain.
upper motor neuron – first neuron in the motor command pathway with its cell body in the cerebral cortex that synapses on the lower motor neuron in the spinal cord.
upper oesophageal sphincter – skeletal muscle sphincter that regulates food movement from the pharynx to the oesophagus.
upregulation – increase in the number of hormone receptors, typically in response to chronically reduced levels of a hormone.
urethra – transports urine from the bladder to the outside environment.
urinalysis – analysis of urine to diagnose disease.
urochrome – haem-derived pigment that imparts the typical yellow colour of urine.
uterine tubes – (also, fallopian tubes or oviducts) ducts that facilitate transport of an ovulated oocyte to the uterus.
uterus – muscular hollow organ in which a fertilised egg develops into a foetus.
utricle – structure of the inner ear responsible for transducing linear acceleration in the horizontal plane.
V
vagina – tunnel-like organ that provides access to the uterus for the insertion of semen and from the uterus for the birth of a baby.
vagus nerve – tenth cranial nerve; responsible for the autonomic control of organs in the thoracic and upper abdominal cavities.
Valsalva’s manoeuvre – voluntary contraction of the diaphragm and abdominal wall muscles and closing of the glottis, which increases intra-abdominal pressure and facilitates defecation.
valve – in the cardiovascular system, a specialised structure located within the heart or vessels that ensures one-way flow of blood.
variable region domain – part of a lymphocyte antigen receptor that varies considerably between different receptor types.
varicosity – enlargement of neurons that release neurotransmitters into synaptic clefts.
vasa recta – branches of the efferent arterioles that parallel the course of the loops of Henle and are continuous with the peritubular capillaries; with the glomerulus, form a portal system.
vasa vasorum – small blood vessels located within the walls or tunics of larger vessels that supply nourishment to and remove wastes from the cells of the vessels.
vasodilation – widening of blood vessels.
vascular shock – type of shock that occurs when arterioles lose their normal muscular tone and dilate dramatically.
vascular shunt – continuation of the metarteriole and thoroughfare channel that allows blood to bypass the capillary beds to flow directly from the arterial to the venous circulation.
vascular spasm – initial step in haemostasis, in which the smooth muscle in the walls of the ruptured or damaged blood vessel contracts.
vascular tone – contractile state of smooth muscle in a blood vessel.
vascular tubes – rudimentary blood vessels in a developing foetus.
vascular tunic – middle layer of the eye primarily composed of connective tissue with a rich blood supply.
vasoconstriction – constriction of the smooth muscle of a blood vessel, resulting in a decreased vascular diameter.
vasodilation – relaxation of the smooth muscle in the wall of a blood vessel, resulting in an increased vascular diameter.
vasomotion – irregular, pulsating flow of blood through capillaries and related structures.
vein – blood vessel that conducts blood toward the heart.
venous reserve – volume of blood contained within systemic veins in the integument, bone marrow, and liver that can be returned to the heart for circulation, if needed.
ventilation – movement of air into and out of the lungs; consists of inspiration and expiration.
ventral – describes the front or direction toward the front of the body; also referred to as anterior.
ventral cavity – larger body cavity located anterior to the posterior (dorsal) body cavity; includes the serous membrane-lined pleural cavities for the lungs, pericardial cavity for the heart, and peritoneal cavity for the abdominal and pelvic organs; also referred to as anterior body cavity.
ventral (anterior) nerve root – axons emerging from the anterior or lateral horns of the spinal cord.
ventral posterior nucleus – nucleus in the thalamus that is the target of gustatory sensations and projects to the cerebral cortex.
ventral respiratory group (VRG) – region of the medulla oblongata that stimulates the contraction of the accessory muscles involved in respiration to induce forced inspiration and expiration.
ventral stream – connections between cortical areas from the occipital lobe to the temporal lobe that are responsible for identification of visual stimuli.
ventricle – one of the primary pumping chambers of the heart located in the lower portion of the heart; the left ventricle is the major pumping chamber on the lower left side of the heart that ejects.
ventricle – central cavity within the brain where CSF is produced and circulates.
ventricular ejection phase – second phase of ventricular systole during which blood is pumped from the ventricle.
venule – small vessel leading from the capillaries to veins.
vernix caseosa – waxy, cheese-like substance that protects the delicate foetal skin until birth.
vertebra – individual bone in the neck and back regions of the vertebral column.
vertebral arch – bony arch formed by the posterior portion of each vertebra that surrounds and protects the spinal cord.
vertebral artery – arises from the subclavian artery and passes through the vertebral foramen through the foramen magnum to the brain; joins with the internal carotid artery to form the arterial circle; supplies blood to the brain and spinal cord.
vertebral column – entire sequence of bones that extend from the skull to the tailbone.
vertebral (spinal) canal – bony passageway within the vertebral column for the spinal cord that is formed by the series of individual vertebral foramina.
vertebral foramen – opening associated with each vertebra defined by the vertebral arch that provides passage for the spinal cord.
vertebral vein – arises from the base of the brain and the cervical region of the spinal cord; passes through the intervertebral foramina in the cervical vertebrae; drains smaller veins from the cranium, spinal cord, and vertebrae, and leads to the brachiocephalic vein; counterpart of the vertebral artery.
vesicle – membrane-bound structure that contains materials within or outside of the cell.
vestibular ganglion – location of neuronal cell bodies that transmit equilibrium information along the eighth cranial nerve.
vestibular fold – part of the folded region of the glottis composed of mucous membrane; supports the epiglottis during swallowing.
vestibular nuclei – targets of the vestibular component of the eighth cranial nerve.
vestibule – in the ear, the portion of the inner ear responsible for the sense of equilibrium.
vestibulocochlear nerve – eighth cranial nerve; responsible for the sensations of hearing and balance.
vestibulo-ocular reflex (VOR) – reflex based on connections between the vestibular system and the cranial nerves of eye movements that ensures images are stabilised on the retina as the head and body move.
vestibulospinal tract – extrapyramidal connections between the vestibular nuclei in the brain stem and spinal cord that modulate movement and contribute to balance based on the sense of equilibrium.
villus – projection of the mucosa of the small intestine.
visceral branches – branches of the descending aorta that supply blood to the viscera.
visceral muscle – smooth muscle found in the walls of visceral organs.
visceral pleura – innermost layer of the pleura that is superficial to the lungs and extends into the lung fissures.
visceral sense – sense associated with the internal organs.
vision – special sense of sight based on transduction of light stimuli.
visual acuity – property of vision related to the sharpness of focus, which varies in relation to retinal position.
vital capacity (VC) – sum of TV, ERV, and IRV, which is all the volumes that participate in gas exchange.
vitamin D – compound that aids absorption of calcium and phosphates in the intestine to improve bone health.
vitiligo – skin condition in which melanocytes in certain areas lose the ability to produce melanin, possibly due an autoimmune reaction that leads to loss of colour in patches.
vitreous humour – viscous fluid that fills the posterior chamber of the eye.
voltage-gated channel – ion channel that opens because of a change in the charge distributed across the membrane where it is located.
voltage-gated sodium channels – membrane proteins that open sodium channels in response to a sufficient voltage change, and initiate and transmit the action potential as Na+ enters through the channel.
voluntary phase – initial phase of deglutition, in which the bolus moves from the mouth to the oropharynx.
vomer bone – unpaired bone that forms the inferior and posterior portions of the nasal septum.
vulva – external female genitalia.
W
wax – lipid comprised of a long-chain fatty acid that is esterified to a long-chain alcohol; serves as a protective coating on some feathers, aquatic mammal fur, and leaves.
wave summation – addition of successive neural stimuli to produce greater contraction.
white blood cells (WBCs) – (also, leukocytes) one of the formed elements of blood that provides defence against disease agents and foreign materials.
white matter – regions of the nervous system containing mostly myelinated axons, making the tissue appear white because of the high lipid content of myelin.
Wolffian duct – duct system present in the embryo that will eventually form the internal male reproductive structures.
working memory – function of the prefrontal cortex to maintain a representation of information that is not in the immediate environment.
wound contraction – process whereby the borders of a wound are physically drawn together.
X
xiphoid process – small process that forms the inferior tip of the sternum.
Y
yellow marrow – connective tissue in the interior cavity of a bone where fat is stored.
yolk sac – membrane associated with primitive circulation to the developing embryo; source of the first blood cells and germ cells and contributes to the umbilical cord structure.
Z
zona fasciculata – intermediate region of the adrenal cortex that produce hormones called glucocorticoids.
zona glomerulosa – most superficial region of the adrenal cortex, which produces the hormones collectively referred to as mineralocorticoids.
zona pellucida – thick, gel-like glycoprotein membrane that coats the oocyte and must be penetrated by sperm before fertilisation can occur.
zona reticularis – deepest region of the adrenal cortex, which produces the steroid sex hormones called androgens.
zone of calcified matrix – region of the epiphyseal plate closest to the diaphyseal end; functions to connect the epiphyseal plate to the diaphysis.
zone of maturation and hypertrophy – region of the epiphyseal plate where chondrocytes from the proliferative zone grow and mature and contribute to the longitudinal growth of the epiphyseal plate.
zonule fibres – fibrous connections between the ciliary body and the lens.
zygapophysial joints – facet joints; plane joints between the superior and inferior articular processes of adjacent vertebrae that provide for only limited motions between the vertebrae.
zygomatic arch – elongated, free-standing arch on the lateral skull, formed anteriorly by the temporal process of the zygomatic bone and posteriorly by the zygomatic process of the temporal bone.
zygomatic bone – cheekbone; paired bones that contribute to the lateral orbit and anterior zygomatic arch.
zygomatic process of the temporal bone – extension from the temporal bone that forms the posterior portion of the zygomatic arch.
zygote – fertilised egg; a diploid cell resulting from the fertilisation of haploid gametes from the male and female lines.
Bibliography
4
American Academy of Dermatology 2012, Tattoos and body piercings, viewed 7 December 2020 from: http://www.aad.org/media-resources/stats-and-facts/prevention-and-care/tattoos-and-body-piercings/.
American Cancer Society 2013, Cancer facts and figures: colorectal cancer: 2011–2013, viewed 7 December 2020, from http://www.cancer.org/Research/CancerFactsFigures/ColorectalCancerFactsFigures/colorectal-cancer-facts-figures-2011-2013-page.
American Cancer Society 2013, Skin cancer: basal and squamous cell, viewed 7 December 2020 from http://www.cancer.org/acs/groups/cid/documents/webcontent/003139-pdf.pdf.
American Red Cross 2013, Blood types, viewed 7 December 2020 from http://www.redcrossblood.org/learn-about-blood/blood-types.
Asthma Australia 2020, Cigarette smoke, viewed 7 December 2020, from https://asthma.org.au/about-asthma/triggers/cigarette-smoke/.
Australian Bureau of Statistics 2013, Australian Health Survey: Biomedical Results for Chronic Diseases, viewed 7 December 2020, from https://www.abs.gov.au/statistics/health/health-conditions-and-risks/australian-health-survey-biomedical-results-chronic-diseases/latest-release.
Australian Cervical Cancer Foundation n.d, Prevention, viewed 7 December 2020, from https://accf.org.au/cervical-health/prevention/#:~:text=There%20are%20two%20cervical%20cancer,in%20both%20men%20and%20women.
Australian Department of Health 2019, Mumps, viewed 7 December 2020, from https://immunisationhandbook.health.gov.au/vaccine-preventable-diseases/mumps.
Australian Government 2020, About the National Cervical Screening Program, viewed 7 December 2020, fromhttps://www.health.gov.au/initiatives-and-programs/national-cervical-screening-program/about-the-national-cervical-screening-program#:~:text=Cervical%20screening%20has%20changed%20in,Pap%20test%20every%20two%20years.
Australian Government 2010, Australia to 2050: future challenges, viewed 7 December 2020, from https://treasury.gov.au/sites/default/files/2019-03/IGR_2010_Overview.pdf.
Australian Institute of Health and Welfare 2016, Diabetic ketoacidosis (DKA) among children and young people with type 1 diabetes, viewed 7 December 2020, from https://www.aihw.gov.au/getmedia/16775f2c-595a-4c94-af26-a7680f316b94/20443.pdf.aspx?inline=true.
Australian Institute of Health and Welfare 2018, Chronic conditions, viewed 7 December 2020, from https://www.aihw.gov.au/getmedia/6bc8a4f7-c251-4ac4-9c05-140a473efd7b/aihw-aus-221-chapter-3-3.pdf.aspx.
Australian Institute of Health and Welfare 2018, Mumps in Australia, viewed 7 December 2020, from https://www.aihw.gov.au/getmedia/c275433d-4ac8-4968-8207-ee48f07118da/aihw-phe-236_Mumps.pdf.aspx.
Australian Institute of Health and Welfare 2019, High blood pressure, viewed December 2020, https://www.aihw.gov.au/reports/risk-factors/high-blood-pressure/contents/high-blood-pressure.
Australian Institute of Health and Welfare 2020, Asthma, viewed 7 December 2020, from https://www.aihw.gov.au/reports/chronic-respiratory-conditions/asthma/contents/asthma.
Australian Institute of Health and Welfare 2020, Diabetes, viewed 7 December 2020, from https://www.aihw.gov.au/reports/diabetes/diabetes-snapshot/contents/how-many-australians-have-diabetes/type-2-diabetes.
Australian Menopause Society 2020, AMS Guide to Equivalent MHT/HRT Doses Australia only, viewed 7 December 2020, from https://www.menopause.org.au/hp/information-sheets/426-ams-guide-to-equivalent-mht-hrt-doses.
Australian Orthopaedic Association n.d, About AOA, viewed 7 December 2020, from https://www.aoa.org.au/about-aoa/about-aoa.
Australian Psychological Society 2019, Pathways to registration as a psychologist, viewed 7 December 2020, from https://asmile.org.au/wp-content/uploads/2019/10/2018-Pathways-to-Registration.pdf.
Australian Society of Plastic Surgeons 2020, Cleft lip & Palate, viewed 7 December 2020, from https://plasticsurgery.org.au/procedures/surgical-procedures/cleft-lip-palate/.
Bagul A, Frost JH, Drage, M 2013, ‘Stem cells and their role in renal ischaemia reperfusion injury,’ American Journal of Nephrology, vol. 37, no. 1, pp. 16-29, viewed 7 December 2020, from http://www.karger.com/Article/FullText/345731.
Beck, J 2012, FYI: how much can a human body sweat before it runs out? Popular Science viewed 7 December 2020 from http://www.popsci.com/science/article/2011-01/fyi-how-much-can-human-body-sweat-it-runs-out.
Beckenmann, R 2010, Australian report on Bisphenol A, Parliament of Australia, viewed 7 December 2020, from https://www.aph.gov.au/About_Parliament/Parliamentary_Departments/Parliamentary_Library/FlagPost/2010/November/Australian_report_on_Bisphenol_A#:~:text=Some%20materials%20containing%20BPA%20have,plastic%20baby%20bottles%20containing%20BPA.
BetterHealth n.d., Blood groups, viewed 7 December 2020, from https://www.betterhealth.vic.gov.au/health/ConditionsAndTreatments/blood-groups?viewAsPdf=true#:~:text=According%20to%20the%20Australian%20Red,AB%20negative%20%E2%80%93%201%20per%20cent.
Bizzintino J, Lee WM, Laing IA, Vang F, Pappas T, Zhang G, Martin AC, Khoo SK, Cox DW, Geelhoed GC, et al 2010, Association between human rhinovirus C and severity of acute asthma in children. Eur Respir J, vol. 37, no. 5, pp. 1037–1042. from: http://erj.ersjournals.com/gca?submit=Go&gca=erj%3B37%2F5%2F1037&allch=.
Boundless 2020, ‘Gomphoses’ in Anatomy and Physiology, viewed 7 December 2020, from https://med.libretexts.org/Bookshelves/Anatomy_and_Physiology/Book%3A_Anatomy_and_Physiology_(Boundless)/8%3A_Joints/8.2%3A_Fibrous_Joints/8.2D%3A_Gomphoses.
Cambridge IVF n.d., Blastocyst vitrification, viewed 7 December 2020, from https://www.cambridge-ivf.org.uk/patients/what-happens/blastocyst-vitrification.
Cancer Australia 2019, Cancer incidence, viewed 7 December 2020, from https://ncci.canceraustralia.gov.au/diagnosis/cancer-incidence/cancer-incidence.
Cancer Australia 2019, Cancer mortality, viewed 7 December 2020, from https://ncci.canceraustralia.gov.au/outcomes/cancer-mortality/cancer-mortality.
Cancer Australia 2020, 5-year relative survival, viewed 7 December 2020, from https://ncci.canceraustralia.gov.au/outcomes/relative-survival-rate/5-year-relative-survival.
Cancer Council n.d,, Understanding your Pap smear or cervical screening test results, viewed 7 December 2020, from https://www.cancer.org.au/cancer-information/causes-and-prevention/early-detection-and-screening/understanding-your-cervical-screening-test-results.
Centers for Disease Control and Prevention 2013, Getting blood pressure under control: high blood pressure is out of control for too many Americans, viewed 7 December 2020, from: http://www.cdc.gov/features/vitalsigns/hypertension/.
Centres for Disease Control and Prevention 2013, Injury prevention and control: traumatic brain injury viewed 7 December 2020, from http://www.cdc.gov/traumaticbraininjury/statistics.html.
Centres for Disease Control and Prevention 2013, Morbidity and mortality weekly report: notifiable diseases and mortality tables, viewed 7 December 2020, from http://www.cdc.gov/mmwr/preview/mmwrhtml/mm6101md.htm?s_cid=mm6101md_w.
CERVARIX: consumer medicine information, viewed 7 December 2020, from https://au.gsk.com/media/480637/cervarix_cmi_au.pdf.
Children’s Health Queensland Hospital and Health Service n.d., Craniosynostosis Fact Sheet, viewed 7 December 2020, from https://www.childrens.health.qld.gov.au/fact-sheet-craniosynostosis/.
Cystic Fibrosis Australia 2020, Cystic Fibrosis, viewed 7 December 2020 https://www.cysticfibrosis.org.au/.
Cystic Fibrosis Foundation n.d., About Cystic Fibrosis, viewed 7 December 2020, https://www.cff.org/What-is-CF/About-Cystic-Fibrosis/#:~:text=More%20than%2030%2C000%20people%20are,is%20age%2018%20or%20older.
Diabetes Australia 2020, Blood glucose monitoring, viewed 7 December 2020, from https://www.diabetesaustralia.com.au/living-with-diabetes/managing-your-diabetes/blood-glucose-monitoring/#:~:text=Glucose%20level%20targets&text=Normal%20blood%20glucose%20levels%20are%20between%204.0%E2%80%937.8mmol%2FL.
Gleason, C & Juul, S 2018, Avery’s Diseases of the Newborn, 10th ed, Elsevier, from https://www.sciencedirect.com/book/9780323401395/averys-diseases-of-the-newborn.
Gray, K & Gibbons, P 2012, Clubfoot: advances in diagnosis and management, Australian Family Physician, vol. 41, no. 5, pp. 299-301, from https://www.racgp.org.au/afp/2012/may/clubfoot/.
Greenvert, J, Bell, S, Guan, Y, Yu, Y 2011, ‘Folic Acid Supplementation and Pregnancy: More Than Just Neural Tube Defect Prevention,’ Reviews in Obstetrics and Gynecology, vol. 4, no. 2, pp. 52-59, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3218540/.
Health Direct n.d, Bacterial vaginosis, viewed 7 December 2020, from https://www.healthdirect.gov.au/bacterial-vaginosis.
Health Direct n.d., Cleft lip and cleft palate, viewed 7 December 2020, from https://www.healthdirect.gov.au/cleft-lip-and-cleft-palate#:~:text=About%20one%20baby%20in%20every,a%20cleft%20lip%20or%20palate.
HealthLinkBC n.d., In Vitro Fertilization for Infertility, viewed 7 December 2020, from https://www.healthlinkbc.ca/health-topics/hw227379.
Herring, M, Putney, L, Wyatt, G, Finkbeiner, W & Hyde, D 2014, ‘Growth of alveoli during postnatal development in humans based on stereological estimation,’ American Journal of Physiology, Lung Cellular and Molecular Physiology, vol. 307, no. 4, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4137164/.
Hypoxico 2020, Hypoxico Altitude to Oxygen Chart, viewed 7 December 2020, from https://hypoxico.com/altitude-to-oxygen-chart/.
International Diabetes Federation 2020, Diabetes facts and figures, viewed 7 December 2020, from https://www.idf.org/aboutdiabetes/what-is-diabetes/facts-figures.html.
Jelkmann, W 2010, Regulation of erythropoietin production, The Journal of Physiology, vol. 589, pp. 1251-1258, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3082088/#:~:text=Erythropoiesis%20is%20a%20slow%2Dacting,days%20before%20reticulocytosis%20becomes%20apparent.
Kolata, G 2012, Severe diet doesn’t prolong life, at least in monkeys, New York Times, viewed 7 December 2020 http://www.nytimes.com/2012/08/30/science/low-calorie-diet-doesnt-prolong-life-study-of-monkeys-finds.html?_r=2&ref=caloricrestriction&.
Kramer, PD 1993, Listening to Prozac, 1st ed, Penguin Books, New York (NY).
Life Fertility Clinic n.d., IVF step-by-step, viewed 7 December 2020, from https://www.lifefertility.com.au/resources/factsheets/ivf-step-by-step/.
Lucile Packard Children’s Hospital at Stanford 2012, Classification and treatment of burns, viewed 7 December 2020 http://www.lpch.org/diseasehealthinfo/healthlibrary/burns/classify.html.
Magoffin, D 2005, ‘Ovarian theca cell’, Int J Biochem Cell Biol, vol. 37, no. 3, pp. 1344-1349, viewed 7 December 2020, from https://pubmed.ncbi.nlm.nih.gov/15833266/#:~:text=Theca%20cells%20are%20the%20endocrine,proteins%20secreted%20from%20growing%20follicles.
Martin RJ, Kraft M, Chu HW, Berns, EA, Cassell GH 2001, A link between chronic asthma and chronic infection, J Allergy Clin Immunol, vol. 107, no. 4, pp. 595-601, viewed 7 December 2020 from http://erj.ersjournals.com/gca?submit=Go&gca=erj%3B37%2F5%2F1037&allch=.
Mater Education 2020, HLT37215 Certificate III in Pathology Collection, viewed 7 December 2020, from https://www.matereducation.qld.edu.au/Course/Certificate-III-in-Pathology-Collection.
Mayo Clinic 2012, Basal cell carcinoma, viewed 7 December 2020 from http://www.mayoclinic.com/health/basal-cell-carcinoma/ds00925/dsection=treatments-and-drugs.
Ming, GL & Song H 2005, ‘Adult neurogenesis in the mammalian central nervous system.’ Annu. Rev. Neurosci, vol. 28, pp. 223–250.
Monash IVF n.d., Monash IVF History, viewed 7 December 2020, from https://monashivf.com/why-monash-ivf/history/.
Monash IVF, n.d., The IVF Process, viewed 7 December 2020, from https://monashivf.com/services/fertility-treatment-options/the-ivf-process/.
Monash University 2015, Australian Cystic Fibrosis Data Registry Annual Report 2015, Victoria, viewed 7 December 2020 https://www.cysticfibrosis.org.au/getmedia/d0718682-f382-4a99-b6e7-f06d31d8bc36/17P-0960-Cystic-Fibrosis-Annual-Report-FINAL.pdf.aspx#:~:text=CF%20is%20a%20common%20inherited,genetic%20mutation%20responsible%20for%20CF.
Monash University 2017, Australian Cystic Fibrosis Data Registry Annual Report 2017, Victoria, viewed 7 December 2020 https://www.cysticfibrosis.org.au/getmedia/24e94d66-29fa-4e3f-8e65-21ee24ed2e5a/ACFDR-2017-Annual-Report_highres_singles.pdf.aspx.
Muscular Dystrophy Australia n.d., Duchenne Muscular Dystrophy, viewed 7 December 2020, from https://www.mda.org.au/disorders/overview/dmd-bmd/.
My Health Career 2014, How long does it take to become a doctor? Viewed 7 December 2020, from https://www.myhealthcareer.com.au/medicine/how-long-does-it-take-to-become-a-fully-qualified-doctor/.
My Health Career 2020, Becoming a dermatologist, viewed 7 December 2020, from https://www.myhealthcareer.com.au/become-a-dermatologist/.
Naftolin, F & Friedenthal, J 2019, ‘Cardiovascular health and the menopausal woman: the role of estrogen and when to begin and end hormone treatment,’ F1000Res, viewed 7 December 2020, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6733383/.
National Center for Biotechnology Information 2004, ‘Style Points and Conventions’ in The NCBI Style Guide, viewed 7 December 2020, from https://www.ncbi.nlm.nih.gov/books/NBK995/#:~:text=in%20vitro%2C%20in%20vivo,Use%20italics.
Paterson K, Horman D & Brumby-Rendell, O n.d., Clubfeet in the top end, viewed 7 December 2020, from http://www.ruralhealth.org.au/13nrhc/images/C3_Paterson_Horman.pdf.
Payscale 2020, Average Forensic Pathologist Salary in Australia, viewed 7 December 2020, from https://www.payscale.com/research/AU/Job=Forensic_Pathologist/Salary.
PayScale 2020, Average Radiographer Hourly Pay in Australia, viewed 7 December 2020, from https://www.payscale.com/research/AU/Job=Radiographer/Hourly_Rate.
PayScale 2020, What Do Physician / Doctor, Cardiologists Do? Viewed 7 December, from https://www.payscale.com/research/AU/Job=Physician_%2F_Doctor%2C_Cardiologis/Salary.
Queensland Health 2016, Forensic pathologist, viewed 7 December 2020, from https://www.health.qld.gov.au/healthsupport/businesses/forensic-and-scientific-services/careers/forensic-pathologist.
Queensland Health 2018, Why do you need to take folic acid when pregnant? Viewed 7 December 2020, from https://www.health.qld.gov.au/news-events/news/why-take-folic-acid-pregnant-spina-bifida#:~:text=You%20should%20take%20a%20supplement,400%20micrograms%20of%20folic%20acid.
Rinninella E, Raoul P, Cintoni, M, Fraceschu F, Miggiano G, Gasbarrini A & Mele M 2019, ‘What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases.’ Microorganisms, vol. 7, no. 1, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6351938/.
Robinson J, Malik A, Parham P, Bodmer JG, Marsh SG 2013, ‘Tissue antigens,’ IMGT/HLA Database, vol. 55. No. 3, pp. 280-287, viewed 7 December 2020 http://europepmc.org/abstract/MED/10777106/reload=0;jsessionid=otkdw3M0TIVSa2zhvimg.
Ross, C, Taylor, C, Yaktine, A & De Valle, H 2011, ‘Dietary Reference Intakes for Calcium and Vitamin D,’ in Dietary Reference Intakes for Calcium and Vitamin D, National Academic Press, Washington DC, from https://www.ncbi.nlm.nih.gov/books/NBK56060/.
Sharma, S & Hashmi, M 2020, Partial pressure of oxygen, StatPearls, viewed 7 December 2020, from https://www.ncbi.nlm.nih.gov/books/NBK493219/#:~:text=Therefore%2C%20at%20sea%20level%20where,7.6%20mm%20Hg%20for%20argon.
Skin Cancer Foundation 2013, Skin cancer facts, viewed 7 December 2020 from http://www.skincancer.org/skin-cancer-information/skin-cancer-facts#top.
Stern, P 2010, ‘Focus issue: getting excited about glia,’ Science, vol. 3, no. 147, pp 330-773. Viewed 7 December http://stke.sciencemag.org/cgi/content/abstract/sigtrans;3/147/eg11.
The Australian and New Zealand Society for Vascular Surgeons n.d, The Australian and New Zealand Society for Vascular Surgeons homepage, from https://www.anzsvs.org.au/.
The Nutrition Source 2012, Fiber and colon cancer: following the scientific trail, Harvard School of Public Health, viewed 7 December 2020, from http://www.hsph.harvard.edu/nutritionsource/nutrition-news/fiber-and-colon-cancer/index.html.
The Royal Australian and New Zealand College of Obstetricians and Gynaecologists n.d., The Royal Australian and New Zealand College of Obstetricians and Gynaecologists homepage, viewed 7 December 2020, from https://ranzcog.edu.au/.
The Royal Australian and New Zealand College of Obstetricians and Gynaecologists n.d, Menopausal Hormone Therapy Advice, viewed 7 December 2020, from https://ranzcog.edu.au/RANZCOG_SITE/media/RANZCOG-MEDIA/Women%27s%20Health/Statement%20and%20guidelines/Clinical%20-%20Gynaecology/Menopausal-Hormone-Therapy-Advice-(C-Gyn-16)-Re-write-July-2015.pdf?ext=.pdf.
The Royal Australasian College of Physicians n.d, Cardiology, viewed 7 December 2020, from https://www.racp.edu.au/trainees/advanced-training/advanced-training-programs/cardiology.
The University of Western Australia 2020, Cardiologist, viewed 7 December 2020, from https://www.uwa.edu.au/study/careers/cardiologist#:~:text=After%20completing%20a%20Doctor%20of,ultimately%20receive%20Fellowship%20which%20normally.
Stinson, F, Boyce, M, Paune, M, Keelan, J 2019, ‘The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth,’ Frontiers in Microbiology, viewed 7 December 2020, from https://www.frontiersin.org/articles/10.3389/fmicb.2019.01124/full.
To, M & Villatoro, V, ‘Red Blood Cell Maturation’ in A Laboratory Guide to Clinical Haematology, viewed 7 December 2020 https://openeducationalberta.ca/mlsci/chapter/red-blood-cell-maturation/.
van Loon FPL, Holmes SJ, Sirotkin B, Williams W, Cochi S, Hadler S, Lindegren ML 1995, Morbidity and Mortality Weekly Report: Mumps surveillance — United States, 1988–1993, viewed 7 December 2020, from http://www.cdc.gov/mmwr/preview/mmwrhtml/00038546.htm.
World Health Organization 2019, Hypertension, viewed 7 December 2020, https://www.who.int/news-room/fact-sheets/detail/hypertension.
World Health Organization 2020, Salt Reduction, viewed 7 December 2020, from https://www.who.int/news-room/fact-sheets/detail/salt-reduction.
World Health Organization n.d., Potassium intake for adults and children, viewed 7 December 2020, from https://www.who.int/elena/titles/guidance_summaries/potassium_intake/en/.
Zhou, Y, Xue, S & Yang, J 2013. ‘Calciomics: integrative studies of Ca2+-binding proteins and their interactomes in biological systems,’ Metallomics, vol. 5, no. 1, pp/ 29-42. Viewed 7 December 2020, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3614492/.